
Trypanosoma cruzi-infected triatomines and rodents co-occur in a coastal island of northern Chile
- Published
- Accepted
- Received
- Academic Editor
- Claudio Lazzari
- Subject Areas
- Ecology, Entomology, Parasitology, Zoology, Infectious Diseases
- Keywords
- T. cruzi reservoir in islands, Island T. cruzi cycle, Island T. cruzi hosts, Mepraia, Insular small mammals, Hemiptera:Reduviidae, Trypanosoma cruzi, T. cruzi life cycle
- Copyright
- © 2020 Campos-Soto et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Trypanosoma cruzi-infected triatomines and rodents co-occur in a coastal island of northern Chile. PeerJ 8:e9967 https://doi.org/10.7717/peerj.9967
Abstract
Trypanosoma cruzi, the cause agent of Chagas disease, is transmitted mainly by blood-feeding insects of the subfamily Triatominae. The T. cruzi life cycle alternates between triatomines and mammalian hosts, excluding birds and reptiles. Triatomines of Mepraia genus are wild vectors of T. cruzi in Chile. Mepraia specimens infected with T. cruzi have been detected in Pan de Azúcar and Santa María islands. The most common vertebrates that inhabit these islands are birds and reptiles, and it is unknown whether small mammals are present. Consequently, it is relevant to know whether there are any T. cruzi-infected small mammals on those islands to elucidate the T. cruzi cycle. To clarify this crossroads, islands of northern Chile were explored to determine if T. cruzi-infected triatomines and rodents co-occur in islands of northern Chile. T. cruzi DNA was detected by conventional and real-time PCR in three islands: on Santa María and Pan de Azúcar islands T. cruzi was detected in Mepraia sp samples, while on Pan de Azúcar (6.1%) and Damas islands (15%) was detected in the rodent Abrothrix olivacea. We show for the first time in Chile the occurrence of insular rodents infected with T. cruzi, and a complete T. cruzi life cycle in a coastal island. Our results provide new insights to understand the T. cruzi infection in the wild cycle.
Introduction
Trypanosoma cruzi is the cause agent of Chagas disease, one of the main zoonotic diseases mediated by vectors in America. This parasite is transmitted principally by blood-feeding bugs of the subfamily Triatominae. The T. cruzi life cycle circulates among triatomines and several mammalian host species while birds and reptiles still are considered refractory to T. cruzi infection (Kierszenbaum, Ivanyi & Budzko, 1976; Urdaneta-Morales & McLure, 1981). Mepraia is a genus (Mazza, Gajardo & Jörg, 1940) of Triatominae endemic to arid and semiarid regions of Chile; it plays an important role in the wild cycle of T. cruzi transmission, and its species are potential vectors for humans (Botto-Mahan et al., 2008; Campos-Soto et al., 2016).
Three species are currently included in the genus Mepraia: M. gajardoi, M. parapatrica and M. spinolai (Frías, Henry & Gonzalez, 1998; Frías-Lasserre, 2010). The first two inhabit coastal areas, while M. spinolai inhabits coastal and interior valleys. M. parapatrica is distributed in the coastal desert in an area intermediate between those of M. spinolai and M. gajardoi (Frías-Lasserre, 2010; Campos et al., 2013). Island populations of M. parapatrica have been reported inhabiting Pan de Azúcar Island in the Atacama Region (Sagua et al., 2000; Frías-Lasserre, 2010). Sagua et al. (2000) suggested that triatomines from Pan de Azúcar Island feed mainly on seabirds (78%), marine mammals (15%) and reptiles (7%). Individuals of Mepraia sp. were also reported in Santa María Island in the Antofagasta Region (Rives-Blanchard et al., 2017).
The presence of triatomines infected with T. cruzi, as well as mixed infections with more than one T. cruzi Discrete Typing Unit (DTU), were reported in both islands (Rives-Blanchard et al., 2017). Mixed infections are more common in ecotopes with high infection rate or high diversity of mammals that harbor different T. cruzi lineages (Campos-Soto et al., 2016). However, the most frequent vertebrates that inhabit both islands are lizards, seabirds and few marine mammals such as seawolf, sea otters and migratory cetaceans (R. Campos-Soto, 2018, field observations of this study). Small mammals inhabiting these islands are unknown, despite their potential major role in the T. cruzi life cycle. Given that there are two islands with infected bugs, are there any T. cruzi-infected small mammals on those islands? Therefore, sampling insular triatomines and small mammals as possible reservoirs of T. cruzi in coastal islands of northern Chile is key to clarify this question and the T. cruzi life cycle. We studied two major islands in the north of Chile (Pan de Azúcar and Santa María) together with three additional islands in which hosts of T. cruzi are unknown without previous infection studies. The evidence provided by this study offers new opportunities to examine the T. cruzi life cycle in coastal islands of northern Chile.
Materials & Methods
Areas of small mammal and triatomine collections
Small mammals and Mepraia individuals were collected during the summer (2017 to 2019) in five coastal islands of northern Chile: Santa María Island (distant site 1.9 km from the continent), in Antofagasta Region; Pan de Azúcar Island (distant site 1.8 km from the continent), in Pan de Azúcar National Park, Atacama Region; Chañaral Island (distant site 8.5 km from the continent), in Atacama Region; Damas Island (in a site at 5.5 km from the continent) and Choros Island (distant site 8.6 km from the continent) in Coquimbo Region. The last three islands are included in the Pingüino de Humboldt National Park. Localities and their geographical locations are shown in Fig. 1 and Table 1. Only on Chañaral Island two field activities were carried out.

Figure 1: Sample localities of small mammal and triatomine collection.
ISMa: Santa María Island, IPA: Pan de Azúcar Island, ICha: Chañaral Island, ICho: Choros Island, IDa: Damas Island. Attributions: Map data©2020 Google, Image ©2020 TerraMetrics, Image©2020 CNES/Airbus, Data SIO. NOAA. US Navy. NGA. GEBCO. Map was modified by illustrations purposes.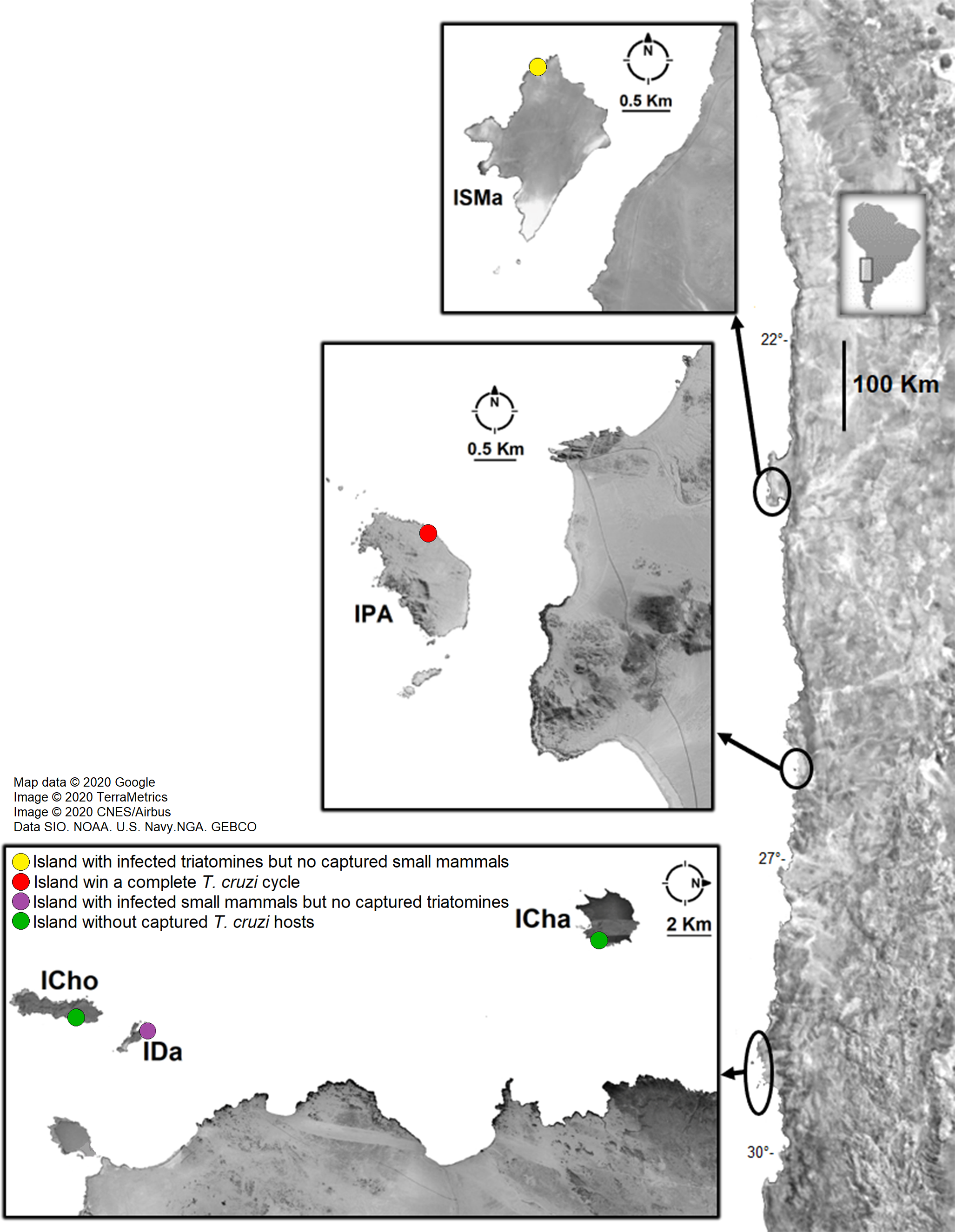{kind=link}
| Island | Latitude/longitude | Host species | ID Host | kDNA PCR | SatDNA qPCR |
|---|---|---|---|---|---|
| Santa María | 23°25′51″S/70°36′31″W | Smam. not found | – | – | – |
| Triat. Mepraia sp | 1isma | + | (-) | ||
| 2isma | + | + | |||
| 3isma | + | + | |||
| 4isma | + | + | |||
| 5isma | (-) | + | |||
| 7isma | (-) | + | |||
| 9isma | + | + | |||
| 10isma | + | (-) | |||
| 12isma | + | + | |||
| 14isma | + | + | |||
| 16isma | + | + | |||
| 18isma | (-) | + | |||
| 22isma | + | + | |||
| 23isma | + | + | |||
| 26isma | (-) | + | |||
| 27isma | (-) | + | |||
| Inf. rate | 11/38 (28.9%) | 14/38 (36.8%) | |||
| Pan de Azúcar | 26°9′6″S/70°41′7″W | Smam. Abrothrix olivaceus | 8Aipa | + | + |
| 25Aipa | + | + | |||
| Inf. rate | 2/33 (6.1%) | 2/33 (6.1%) | |||
| Triat. Mepraia parapatrica | 1ipa | + | + | ||
| 2ipa | + | + | |||
| 5ipa | (-) | + | |||
| 6ipa | + | + | |||
| 7ipa | + | (-) | |||
| 8ipa | (-) | + | |||
| 9ipa | (-) | + | |||
| 10ipa | (-) | + | |||
| 15ipa | (-) | + | |||
| 17ipa | (-) | + | |||
| 19ipa | (-) | + | |||
| 23ipa | (-) | + | |||
| 26ipa | (-) | + | |||
| 29ipa | + | (-) | |||
| 40ipa | + | (-) | |||
| Inf. rate | 6/59 (10.17%) | 12/59 (20%) | |||
| Chañaral | 29°2′17″S/71°34′8″W | Smam. Thylamys elegans | 0/9 | 0/7 | |
| Triat. not found | – | – | – | ||
| Damas | 29°13′49″S/71°31′47″W | Smam. Abrothrix olivaceus | 12ida | + | + |
| 18ida | + | + | |||
| 21ida | + | + | |||
| Inf. rate | 3/20 (15%) | 3/20 (15%) | |||
| Triat. not found | – | – | – | ||
| Choros | 29°2′17″S/71°34′8″W | Smam. not found | – | – | – |
| Triat. not found | – | – | – |
Small mammal and triatomine sampling
Insular rodents were caught with standard Sherman traps (8 × 9 × 23 cm). Trapping effort was 100 traps/night and conducted for two nights at each site. This sampling design was previously shown successful to capture small mammals (Boric-Bargetto et al., 2016). A mixture of oats (900 grs) and vanilla essence (150 ml) was used as bait, which have been used in several studies with efficient results (Torres-Pérez et al., 2004; Boric-Bargetto et al., 2012). Small mammals were anesthetized in the field with isoflurane and 0.2 ml of blood was sampled in a field laboratory. Fresh blood samples were received in a cryotube and conserved in a liquid nitrogen container. Blood samples were taken only from adult rodents; juvenile and pregnant females were released. The captured rodents were marked with a temporal nontoxic-highlighter and released once they were well awake and recovered. Triatomines were collected passively as described in Campos-Soto, Torres-Pérez & Solari (2015) by qualified personnel, when this method was unsuccessful triatomines were collected actively by lifting stones in rock piles and nests. Captured insects were transported to the laboratory and maintained in a climate chamber at 27 ° C with a relative humidity of 50% and a 14:10 h light:dark photoperiod. Then the complete gut of triatomines was dissected and used for DNA extraction.
Ethics statement
All individuals were manipulated following the standard bioethics and biosafety protocols proposed by the American Society of Mammalogists (Sikes, 2016). Sampling procedures were authorized by the Servicio Agrícola y Ganadero (resolution number: 8353), Corporación Nacional Forestal from Atacama Region (permit number: 049/2017) and Coquimbo Region (permit number: 22/2019). We appreciate the logistical help was provided by Pan de Azúcar National Park and Pingüino de Humboldt National Reserve administrators and their park rangers. The research project that includes this study was approved by the Bioethic and Biosecurity Committee of the Pontificia Universidad Católica de Valparaíso (permit number: BIOEPUCV-A98b-2017).
DNA extraction from triatomines and blood of small mammals
DNA was extracted from blood samples and intestinal contents of triatomines using the DNeasy® Blood & Tissue kit (QIAGEN). The protocol was carried out according to the manufacturer’s instructions; the DNA was eluted twice with 100 µL of elution buffer. All samples were co-extracted with 100 pg of a sequence of Arabidopsis thaliana used as a heterologous internal amplification control (IAC) as previously described in Mc Cabe et al. (2019) to discount loss of DNA or carryover of polymerase chain reaction (PCR) inhibitors.
Kinetoplast DNA conventional PCR assays
Assays were performed for all samples using kinetoplast DNA (kDNA) primers 121 (AAATAATGTACGGGKGAGATGCATGA) and 122 (GGTTCGATTGGGGTTGGTGTAATATA) (Wincker et al., 1994) and the polymerase fast PCR Master Mix SapphireAmp® (Takara). Cycling conditions were 30 s at 94 °C, followed by 40 cycles at 94 °C for 30 s, 55 °C for 30 s and 72 °C for 1 min according to Takara manufacturer’s instructions in a Bioer model TC-96/G/H(b)C LifeEco® thermocycler. Verification of amplification of a variable region of 330 bp of minicircle kDNA was assessed by 2% agarose gel electrophoresis. Each sample was tested twice to confirm the infection with T. cruzi; the sample was considered infected with T. cruzi when at least one of the two amplifications resulted positive.
Satellite DNA real-time PCR assays
Assays were performed using T. cruzi nuclear satellite DNA (satDNA) primers Cruzi 1 (ASTCGGCTGATCGTTTTCGA) and Cruzi 2 (AATTCCTCCAAGCAGCGGATA) (Piron et al., 2007) in a final volume of 20 µL containing 5 µL DNA template as previously described in Muñoz-San Martín et al. (2020). Each sample was tested in duplicate.
Parasite standard calibration curve
T. cruzi DNA standards for absolute quantification were obtained from 105 parasite equivalents/mL (par-eq/mL) of clonal reference strains Dm28c (TcId) and Y (TcII) and 10-fold serial dilutions were performed with nuclease-free water (range between 105 and 101 par-eq/mL) as previously described in Muñoz-San Martín et al. (2020).
Heterologous internal amplification control qPCR Assays
Assays were performed in blood samples using primers IAC Fw (5′ACCGTCATGGAACAG CACGTA 3′) and IAC Rv (5′ CTCCCGCAACAAACCCTATAAAT 3′) Duffy et al., 2013 at a final concentration of 0.2 µM and at a melting temperature of 58 °C as previously described in (Mc Cabe et al., 2019). Quantification of the parasite equivalents from DNA samples was calculated considering the amplification curve of standard T. cruzi DNA and the results were normalized according to the heterologous IAC results.
Genotyping assays
Four DTU real-time PCR genotyping assays were performed for mammal samples positive for T. cruzi (Muñoz Martín, Apt & Zulantay , 2017). Detection of TcI, TcII, TcV, and TcVI was performed using the same primers, concentrations, and controls as previously described in (Muñoz-San Martín et al., 2018). The other assay conditions, including the cycling profile, are described above. Controls were always included in each reaction and each sample was tested in duplicate.
Results
Small mammal and triatomine collection
Small mammals were captured in three islands (Figs. 1 and 2, Table 1). In Pan de Azúcar Island, 49 Abrothrix olivacea (Sigmodontinae) were captured in one sampling night; 33 blood samples were obtained. In Chañaral Island, nine mouse opossums of the species Thylamys elegans (Didelphidae) were captured in two sampling nights in the first field activity (Fig. S1), obtaining nine blood samples. In Damas Island, 48 Abrothrix olivacea were captured in two sampling nights, obtaining 20 blood samples (Table 1, Fig. 2). No small mammals were found in Santa María and Choros Islands. Mepraia triatomines were captured in Santa María Island (N = 38) and Pan de Azúcar Island (N = 59); no triatomines were found in the other three islands. Collected small mammals and triatomines for each island and their geographical coordinates are detailed in Table 1 and Fig. 1.

Figure 2: Sampling sites on islands with detected Trypanosoma cruzi hosts.
(A) Santa María Island. (B) Pan de Azúcar Island. (C) Damas Island. (D) high abundance of Microluphus atacamensis on Santa María Island. (E) Abrothrix olivacea form Pan de Azúcar Island. (F) A. olivacea form Damas Island. Photos A, B, D and E credit: Ricardo Campos-Soto, photos C and F credit: Javier Cruz.{kind=link}
Kinetoplast DNA conventional PCR assays
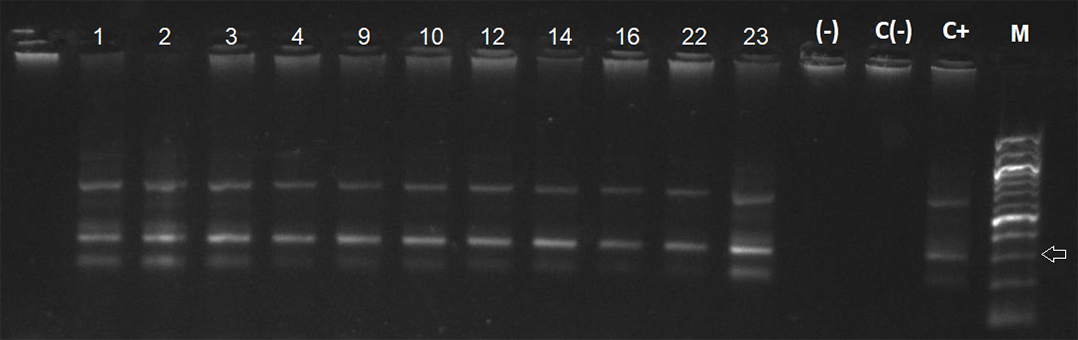
T. cruzi kDNA in small mammals was detected in two Abrothrix olivacea in Pan de Azúcar Island, representing an infection rate of 6.06%. No infected Thylamys elegans were found in Chañaral Island, and three A. olivacea were detected with T. cruzi (infection rate of 15%) in Damas Island (Table 1, Fig. 3). For triatomines, 11 samples of Mepraia were positive for T. cruzi in Santa María Island, representing an infection rate of 28.9% (Fig. S2, Table 1). In Pan de Azúcar Island we detected six samples of Mepraia positive for T. cruzi, representing an infection rate of 10.17% (Fig. S3, Table 1).
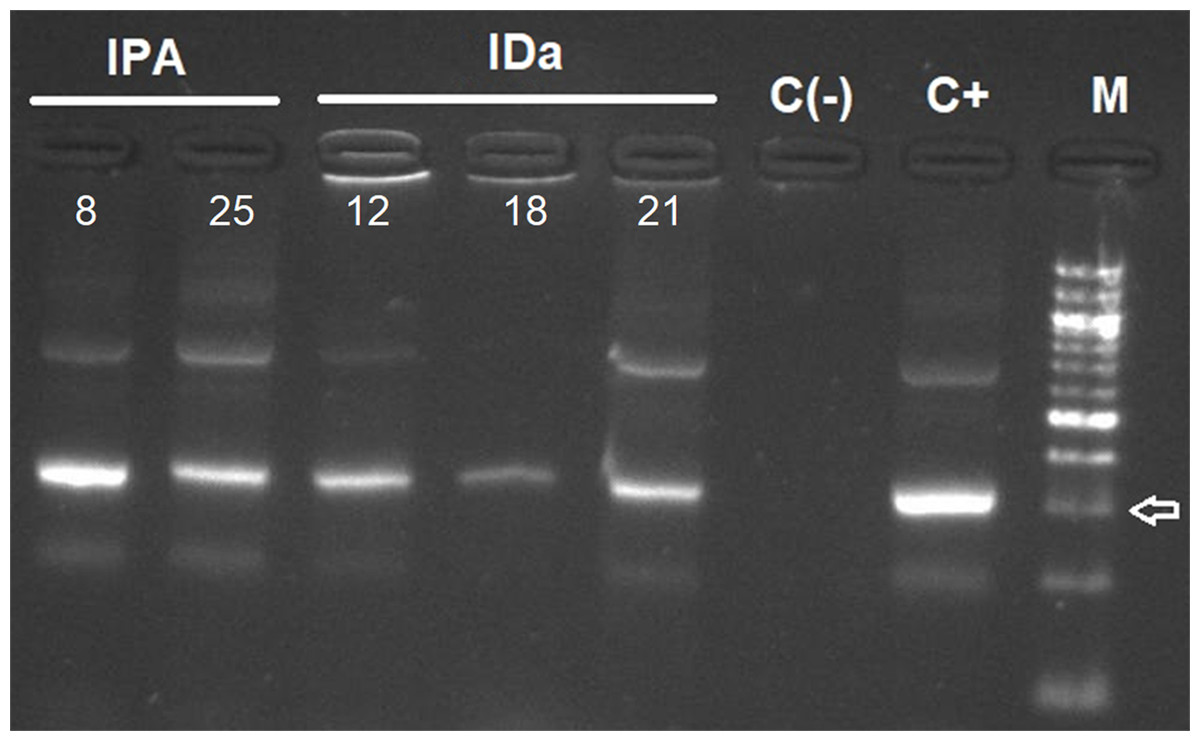
Figure 3: Agarose gel electrophoresis of amplified kDNA by conventional PCR in small mammals (Abrothrix olivacea).
Lanes IPA: positive rodents from Pan de Azúcar Island, IDa: positive rodents from Damas Island. Lanes C(-): negative control, C+: Positive control and M: 100-bp DNA ladder. Arrow indicates 300 bp. Photo credit: Gabriel Díaz-Campusano.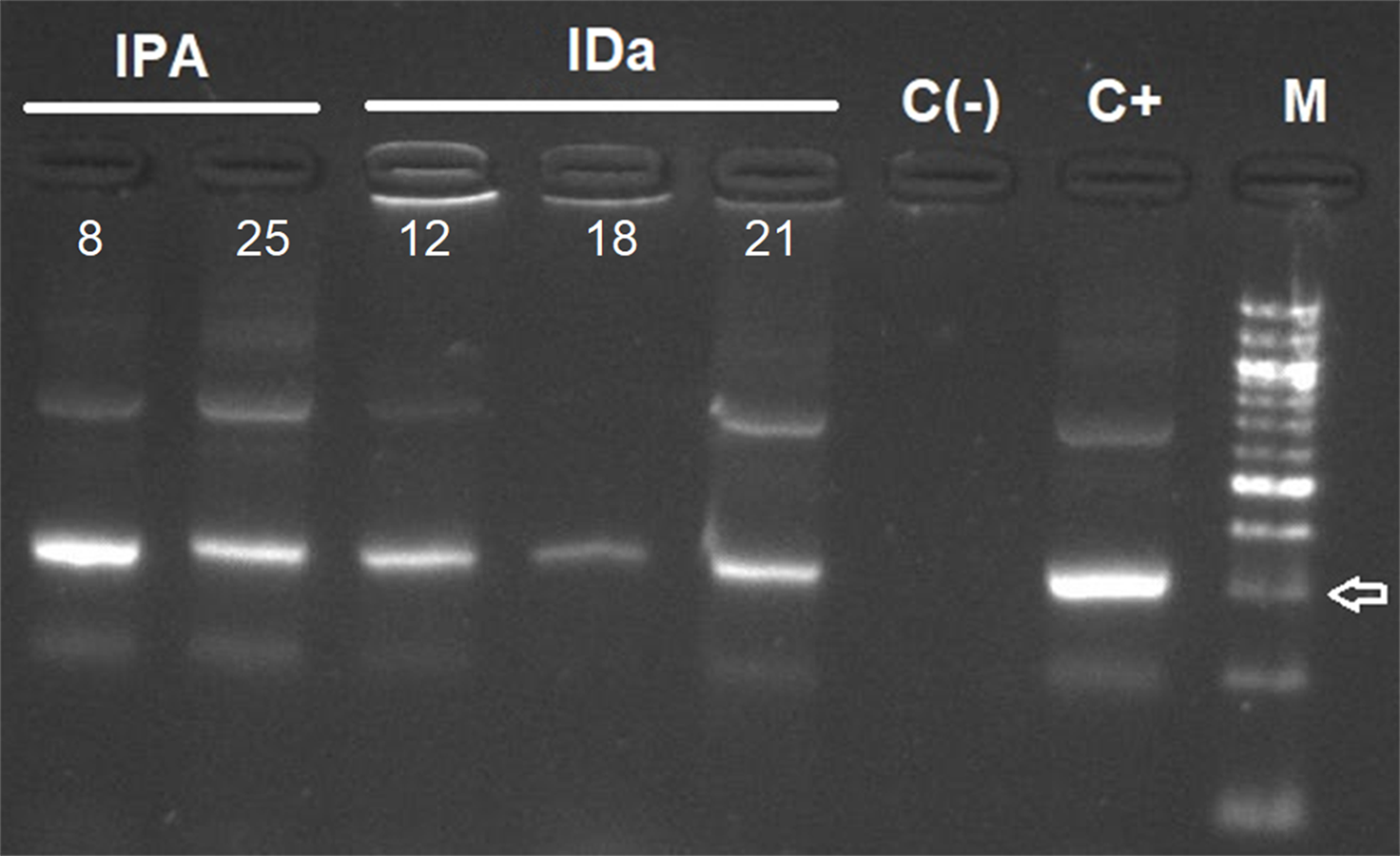{kind=link}
Satellite DNA real-time PCR assays
For T. cruzi satDNA detection in small mammals, all the same A. olivacea samples were positive as in the detection by kDNA (Table 1, Fig. S4). The genomic quantification by real-time PCR only was possible for Abrothrix from Pan de Azúcar Island, with 129 (sample 8Aipa) and 3.6 (sample 25Aipa) par-eq/mL while in Abrothrix from Damas Island low parasitemias were detected (<1 par-eq/mL). For T. cruzi satDNA detection in triatomines, 14 Mepraia samples for Santa María Island were positive, representing an infection rate of 36.8%, while in Pan de Azúcar Island we found 12 positive samples, with an infection rate of 20.33% (Table 1, Fig. S4). Raw data of real-time PCR analyses in small mammals and triatomines are available in Table S1.
Genotyping assays
Genotyping T. cruzi DTU assays were only performed in the Abrothrix from Pan de Azúcar Island because higher parasitemia was detected. We found the TcII DTU in a mixed infection with TcVI in A. olivacea (sample 8Aipa), and a single infection with TcII (sample 25Aipa).
Discussion
The wild cycle of T. cruzi in continental areas has been widely reported, but there is still remains a lack of knowledge in insular areas. A few examples were reported that revealed the enzootic T. cruzi cycle that included mammals and triatomines in islands of Brazil (Grisard et al., 2000; Das Xavier et al., 2014). In Chile, T. cruzi- infected triatomines were reported in Santa María and Pan de Azúcar islands (Rives-Blanchard et al., 2017), raising question related to the mammal hosts involved in this T. cruzi cycle.
On Pan de Azúcar Island, two A. olivacea were positive to T. cruzi by kDNA PCR and satDNA qPCR (infection rate 6.06%, Table 1, Fig. 3). It has been suggested that triatomines on Pan de Azúcar Island feed mainly on sea birds (Sagua et al., 2000), which may explain the low infection rate found in A. olivacea. Interestingly, one of the A. olivacea samples showed a mixed infection with two DTU, TcI and TcII, congruent with the two DTUs previously found in Mepraia on Pan de Azúcar Island (Rives-Blanchard et al., 2017). Therefore, our results confirm that on Pan de Azúcar Island there is a complete T. cruzi life cycle, i.e., a cycle in which T. cruzi circulates through triatomines and mammalian blood.
Three A. olivacea on Damas Island resulted positive for T. cruzi by kDNA PCR and satDNA qPCR (infection rate 15% Table 1, Fig. 3). However, no triatomines were found in our study, a pattern also detected on Chañaral and Choros islands. It has been reported that many triatomines foci can go undetected when vector density is low (Abad-Franch et al., 2014). A likely explanation is that we failed to find triatomines on those islands particularly if they occur at low abundance (sampling bias). However, we sampled intensively using both a passive (Campos-Soto, Torres-Pérez & Solari, 2015) and active method, and the park rangers never reported seeing triatomines (personal communication with park rangers). A study suggests that the current absence of M. spinolai in those islands may be explained by their absence when the islands were formed or that ancient allopatric populations were extinguished (Campos-Soto et al., 2020). If hypothetically triatomines are not present, alternatives to explain the T. cruzi infection in rodents include: (i) Maintenance of the parasite by vertical trans-placental transmission between rodents, which has been reported in humans, bats and mice (Delgado & Santos-Buch, 1978; Añez, Crisante & Soriano, 2009; Ortiz et al., 2012); (ii) Cross-reaction with other trypanosomatids (see below).
On Santa María Island, individuals of Mepraia infected with T. cruzi were previously reported (Rives-Blanchard et al., 2017), but no small mammals were found in our study despite the sampling effort was similar to that performed on Pan de Azúcar and Damas islands. Unlike the other islands, Santa María Island lacks of vegetation (Fig. 2, Fig. S1), and the influence of climatic conditions create an arid and desertic landscape (Jerez, 2000; Cavieres et al., 2002; Clarke, 2006). Also, this island had very few suitable places to set traps (Fig. 2). Under these conditions, detecting small mammals can be difficult, likely impacting our results. Future studies including higher sampling effort both in density and temporal may elucidate this finding. Strikingly, we found triatomines with an infection rate of 28.9% by kDNA PCR and 36.8% by satDNA qPCR (23.6% confirmed by both, Table 1). These values reveal a high infection rate, particularly taking into account that the most abundant vertebrates inhabiting this island are marine birds and reptiles of the genus Microlophus (Fig. 2; R. Campos-Soto, 2018, field observations of this study).
Mixed infection was previously reported in bugs from Santa María Island (Rives-Blanchard et al., 2017). It has been suggested that mixed infections are more frequent in areas with high infection rate and/or there is high diversity of mammals that harbor different T. cruzi lineages (Campos-Soto et al., 2016). This is congruent with our triatomine infection rates but contrasts with our small mammals captures. Despite we cannot confirm the absence of small mammals on Santa María Island, the question of how the T. cruzi cycle is maintained still remains. In the absence of small mammals on Santa María Island, one explanation for our results is the horizontal transmission of T. cruzi among triatomines. For example, coprophagy and cleptohematophagy were reported as (uncommon) mechanisms of transmission among triatomine vectors, mainly by young nymphs (Ryckman, 1951; Schaub, 1988). On the other hand, Rives-Blanchard et al. (2017) showed that there are positive triatomines on Santa María Island, in which T. cruzi DTUs were not identified by hybridization assays. The authors suggested that some TcI or TcII variants did not hybridize with the probes used, or that there are other DTUs not analyzed in their study. Alternatively, there is the possibility that these unidentified DTUs could be another Trypanosoma with cross-reactivity to T. cruzi. In fact, cross-reactivity in parasite detection by PCR and qPCR analyses has been reported between T. cruzi and T. rangeli (Ramírez et al., 2015; Seiringer et al., 2017). The only triatomines that can transmit T. rangeli are Rhodnius and Panstrongylus (Vallejo et al., 2015). These genera do not co-occur with Mepraia, therefore cross-reactivity with T. rangeli is unlikely.
Lizard and avian trypanosomes phylogenetically related to T. cruzi has been reported (Hughes & Piontkivska, 2003; Hamilton, Gibson & Stevens, 2007; Viola et al., 2008; Dario et al., 2017). Reptiles of the genus Microlophus and marine birds are found in high abundance on Santa María Island (Fig. 2), thus triatomines may feed mainly of bird and reptile blood. Also, it has been observed that Microlophus actively hunt and feed on these triatomines (R. Campos-Soto, 2018, field observations of this study). Consequently, the triatomines could be hosting a reptilian or avian trypanosome that could have cross-reactivity with T. cruzi. However, reptilian trypanosomes are transmitted by dipterous sandflies and not by triatomines (Adler & Theodor, 1957; Hamilton, Gibson & Stevens, 2007). According to Seiringer et al. (2017), the best T. cruzi diagnosis is a combination of both kinetoplast DNA detection and nuclear satellite DNA by conventional PCR and qPCR assays, respectively. In our study, all small mammals and most of the triatomines were positive for T. cruzi by conventional PCR (targeting kDNA) and qPCR (targeting nuclear satellite DNA), suggesting the absence of cross-reaction with reptilian or avian trypanosomes. Trypanosomes such as T. brucei can exceptionally infect lizards in the wild cycle (Njagu et al., 1999), and chickens experimentally (Minter-Goedbloed, 1981). A study shows that the availability of reptiles is positively related to the T. cruzi infection risk in an endemic area of Chile (San Juan et al., 2020). These antecedents show that the role of endemic reptiles as hosts of T. cruzi remains to be elucidated.
The mechanisms of colonization of Mepraia to the islands of northern Chile are relevant to understand the T. cruzi infection in these areas. The origin of Mepraia populations on Santa María and Pan de Azúcar islands was suggested by mechanisms of vicariance and dispersal, starting about middle-upper Pleistocene. Bidirectional migration rates between these islands and continental populations was inferred (Campos-Soto et al., 2020). Possible means of dispersal include passive transport by marine birds (Schofield et al., 1998; Sagua et al., 2000) and fishermen who sail to the islands carrying infected triatomines and/or eggs in their clothes or backpacks. An additional passive dispersal mechanism might include sea wolves (Otaria flavescens), which could transport nymphs within their pelage (Schofield et al., 1998). The flight of kissing-bugs also may be another dispersal mechanism. However, Mepraia’s nymphs and adult females are wingless while males show wings polymorphism (Schofield et al., 1998; Campos et al., 2011). M. parapatrica and M. gajardoi males are brachypterous (Frías-Lasserre, 2010), with wings shorter or equal than the abdomen length (Campos et al., 2011) and flying capacity not documented, which would allow discard the dispersion by flight of the bugs.
Conclusions
In conclusion, we show for the first time in Chile the occurrence of insular rodents infected with T. cruzi, and a complete T. cruzi life cycle in a coastal island (Pan de Azúcar Island). We also show two different contrasting results: an island (Santa María) with infected triatomines but without captured small mammals, and another island (Damas) with infected rodents but without captured triatomines (Fig. 1, Table 1). Future studies including a greater capture effort targeting the hosts and the vector will help to elucidate the transmission mechanism maintaining the T. cruzi life cycle on those islands. Our study provides new relevant knowledge about the T. cruzi cycle on islands and the role of its hosts and vectors.
Supplemental Information
Island without detected Trypanosoma cruzi hosts
(A) Chañaral Island. (B) Thylamys elegans from Chañaral Island. (C) Choros Island. Photo credits: Ricardo Campos-Soto.
{kind=link}
Agarose gel electrophoresis of amplified kDNA by conventional PCR in triatomines from Santa María Island
Lanes C(-): negative control, C+: Positive control and M: 100-bp DNA ladder. Arrow indicates 300 bp. Photo credit: Gabriel Díaz-Campusano.
{kind=link}
Agarose gel electrophoresis of amplified kDNA by conventional PCR in triatomines from Pan de Azúcar Island
Lanes C(-): negative control, C+: Positive control and *: indicates positive sample. Photo credit: Nicol Quiroga.
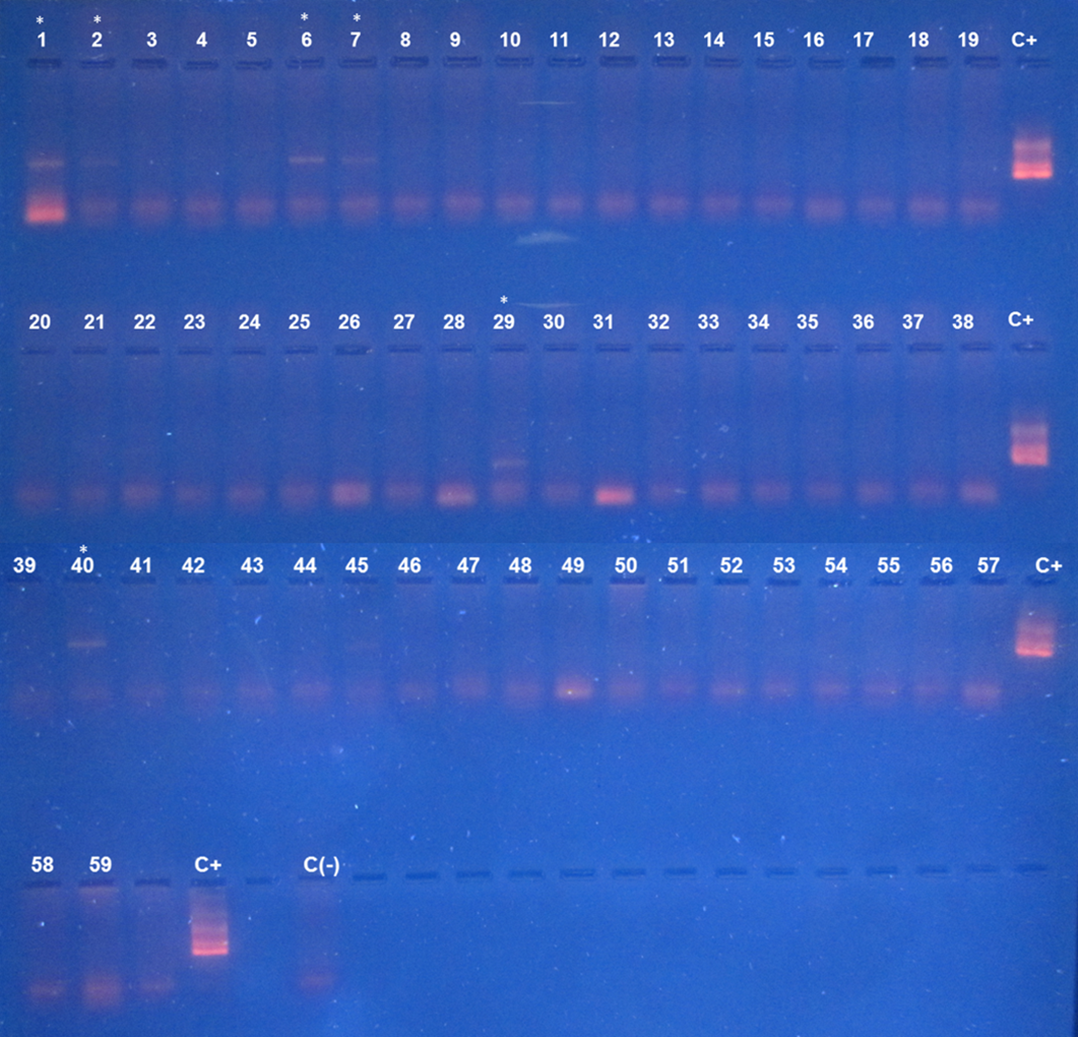{kind=link}
Cycle threshold (Ct) values of positive Satellite DNA real-time PCR assays in small mammals and triatomines from islands
(A) Blood samples of Abrothrix olivacea. (B) Intestinal contents samples of Mepraia sp. The line inside the box represents the median, and the box extends from the lower to the upper quartiles. Whiskers indicate min to max and dots represent the samples. Figure credit: Catalina Muños San-Martín.
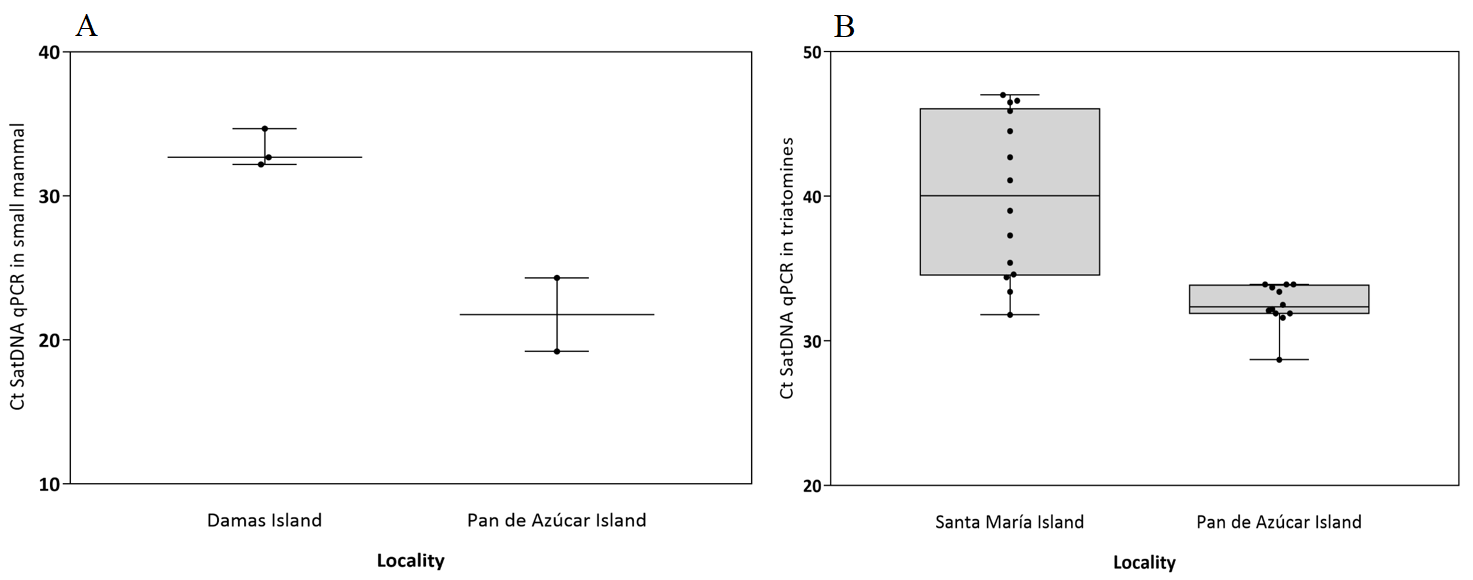{kind=link}
Raw data result of qPCR analyses in small mammals and triatomines
The T. cruzi column refers to the result of satellite DNA amplification; the Ct is the cycle number at which the fluorescence generated within a reaction crosses the threshold, the par-eq/mL is the absolute quantification result. The IAC column shows the result of a sequence of Arabidopsis thaliana DNA amplification. Normalization: IAC results were used for normalization of the quantification result (small mammals only).