
Alveolar soft-part sarcoma (ASPS) resembles a mesenchymal stromal progenitor: evidence from meta-analysis of transcriptomic data
- Published
- Accepted
- Received
- Academic Editor
- Kenta Nakai
- Subject Areas
- Bioinformatics, Cell Biology, Genomics, Oncology
- Keywords
- ASPS, Sarcoma, Mesenchymal, Stromal, Microarray, RNA-seq, Meta-analysis, Surfaceome, Genomics, Transcriptome
- Copyright
- © 2020 Stockwin
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Alveolar soft-part sarcoma (ASPS) resembles a mesenchymal stromal progenitor: evidence from meta-analysis of transcriptomic data. PeerJ 8:e9394 https://doi.org/10.7717/peerj.9394
Abstract
Alveolar soft-part sarcoma (ASPS) is an extremely rare malignancy characterized by the unbalanced translocation der(17)t(X;17)(p11;q25). This translocation generates a fusion protein, ASPL-TFE3, that drives pathogenesis through aberrant transcriptional activity. Although considerable progress has been made in identifying ASPS therapeutic vulnerabilities (e.g., MET inhibitors), basic research efforts are hampered by the lack of appropriate in vitro reagents with which to study the disease. In this report, previously unmined microarray data for the ASPS cell line, ASPS-1, was analyzed relative to the NCI sarcoma cell line panel. These data were combined with meta-analysis of pre-existing ASPS patient microarray and RNA-seq data to derive a platform-independent ASPS transcriptome. Results demonstrated that ASPS-1, in the context of the NCI sarcoma cell panel, had some similarities to normal mesenchymal cells and connective tissue sarcomas. The cell line was characterized by high relative expression of transcripts such as CRYAB, MT1G, GCSAML, and SV2B. Notably, ASPS-1 lacked mRNA expression of myogenesis-related factors MYF5, MYF6, MYOD1, MYOG, PAX3, and PAX7. Furthermore, ASPS-1 had a predicted mRNA surfaceome resembling an undifferentiated mesenchymal stromal cell through expression of GPNMB, CD9 (TSPAN29), CD26 (DPP4), CD49C (ITGA3), CD54 (ICAM1), CD63 (TSPAN30), CD68 (SCARD1), CD130 (IL6ST), CD146 (MCAM), CD147 (BSG), CD151 (SFA-1), CD166 (ALCAM), CD222 (IGF2R), CD230 (PRP), CD236 (GPC), CD243 (ABCB1), and CD325 (CDHN). Subsequent re-analysis of ASPS patient data generated a consensus expression profile with considerable overlap between studies. In common with ASPS-1, elevated expression was noted for CTSK, DPP4, GPNMB, INHBE, LOXL4, PSG9, SLC20A1, STS, SULT1C2, SV2B, and UPP1. Transcripts over-expressed only in ASPS patient samples included ABCB5, CYP17A1, HIF1A, MDK, P4HB, PRL, and PSAP. These observations are consistent with that expected for a mesenchymal progenitor cell with adipogenic, osteogenic, or chondrogenic potential. In summary, the consensus data generated in this study highlight the unique and highly conserved nature of the ASPS transcriptome. Although the ability of the ASPL-TFE3 fusion to perturb mRNA expression must be acknowledged, the prevailing ASPS transcriptome resembles that of a mesenchymal stromal progenitor.
Introduction
Alveolar Soft-Part Sarcoma (ASPS) is an extremely rare soft tissue sarcoma of adolescents and young adults (Christopherson, Foote & Stewart, 1952; Paoluzzi & Maki, 2019). ASPS usually manifests as a soft, painless, slow-growing mass and although disease follows an indolent course, it has the potential to metastasize to several sites (Portera Jr et al., 2001). ASPS is characterized by an unbalanced translocation t(X;17)(p11;q25) that fuses the ASPSCR1 and TFE3 genes, generating a fusion protein that drives pathogenesis (Ladanyi et al., 2001). Evidence suggests that the fusion protein accumulates in the nucleus and directs transcriptional activity (Argani et al., 2003; Betschinger et al., 2013; Hirobe et al., 2013). For example, ASPL-TFE3 binds and activates MET transcription, resulting in an overall enhancement in kinase activity in the presence of hepatocyte growth factor (Tsuda et al., 2007). As a consequence, some clinical benefit is being achieved with kinase inhibitors targeting MET (Paoluzzi & Maki, 2019; Schoffski et al., 2018).
Despite this progress, the origin of disease is still the subject of intense speculation (Folpe & Deyrup, 2006). A longstanding hypothesis posits that ASPS has a myogenous origin (Fisher & Reidbord, 1971; Folpe & Deyrup, 2006; Mukai et al., 1983). However, ASPS tumors do not appear to express markers of muscle cell differentiation such as the myogenic nuclear regulatory proteins MyoD1 and myogenin (Gomez et al., 1999; Hoshino et al., 2009; Tallini et al., 1994; Wang et al., 1996). Several transcriptomic studies have also been published that speculate on the origin of disease (Goodwin et al., 2014; Selvarajah et al., 2014; Stockwin et al., 2009; Tanaka et al., 2017). In 2009 we undertook one of the first microarray studies of ASPS and identified expression of several muscle-restricted transcripts (ITGB1BP3/MIBP, MYF5, MYF6, and TRIM63). However, these data were generated using universal RNA as a reference, which may have biased results towards skeletal muscle expressed transcripts (Stockwin et al., 2009). Selvarajah et al. (2014) showed that the transcription factor PAX6 was upregulated in primary ASPS, suggesting a “tentative neural line of differentiation for ASPS”. Goodwin et al. (2014) generated microarray data from a mouse model of ASPS and also human patient samples. These authors speculated that “some mesenchymal progenitor, possibly pericyte/endothelial in character, provides one potential cell of origin”. Similarly, Tanaka et al. (2017) were able to model ASPS through ectopic expression of ASPL-TFE3 in murine embryonic, but not adult, mesenchymal cells. These observations underscore the current lack of clarity with respect to ASPS ontogeny and lend support to the suggestion that ASPS cells represent a “scrambled” phenotype where the ASPL-TFE3 fusion impairs differentiation (Folpe & Deyrup, 2006; Naka et al., 2013).
In 2011, a multi-year study culminated in the development of an ASPS cell line designated ASPS-1 (Kenney et al., 2011). This reagent provided the first opportunity to study ASPS gene expression without interference from contaminating cell types. Microarray data was subsequently generated for ASPS-1 as part of the NCI sarcoma cell line panel (Teicher et al., 2015). In the current study, ASPS-1 data was mined relative to the entire NCI sarcoma cell line panel. These efforts were combined with re-analysis of microarray and RNA-seq studies focusing on ASPS patient samples (Goodwin et al., 2014; Kummar et al., 2013; Stockwin et al., 2009). In this regard, we aimed to unify current publicly available transcriptomic data into a consensus profile that can be used as a basis for exploring disease ontogeny and therapeutic vulnerabilities. Results obtained in this study show that, at the mRNA level, ASPS resembles a mesenchymal stromal cell (MSC).1
Materials and Methods
This study utilized six datasets downloaded from the gene expression omnibus (https://www.ncbi.nlm.nih.gov/geo/). GSE68591 comprises exon expression data (Affymetrix Human Exon 1.0 v2 ST platform) for the NCI sarcoma cell line panel (includes data from ASPS-1, 67 sarcoma lines, and five normal tissues) (Teicher et al., 2015). GSE13433 comprises mRNA expression data for seven ASPS patient tumors analyzed using the Affymetrix U133 plus 2.0 platform (Stockwin et al., 2009). For the analyses of data from GSE13433, additional U133 plus 2.0 control arrays were obtained from GSE17070 (normal skeletal muscle) and GSE118370 (normal lung). GSE32569 comprises a set of U133 plus 2.0 microarrays generated from six patients pre- and post- treatment with Cediranib (Kummar et al., 2013). Lastly, GSE54729 comprises RNA-seq data (HiSeq 2000) from five ASPS patients and three skeletal muscle controls. The overall study design is illustrated in Fig. 1. For experiments involving Affymetrix human Exon 1.0 ST and U133 plus 2.0 arrays differentially expressed genes were identified using the Transcriptome Analysis Console (TAC 4.0, ThermoFisher Scientific) using standard algorithm and comparison settings (RMA normalization, P < 0.05, FDR <0.05, fold change +/-2). The TAC was also used to generate hierarchical clusters using the automated workflow. For RNA-seq data, differential expression values relative to skeletal muscle were determined using the GSE54729_10408R.txt spreadsheet that accompanies the submission. In detail, normalized FKPM values were averaged for the 5 human ASPS samples and the 3 normal human skeletal muscles samples; fold changes were then calculated from these values. In terms of utilities; the GTEX portal (https://www.gtexportal.org/) multi gene query option (https://gtexportal.org/home/multiGeneQueryPage) was used to inform tissue of origin from the top 50 differentially expressed ASPS-1 transcripts. Similarly, the GTEX Top 50 expressed genes search function (https://gtexportal.org/home/topExpressedGenePage) was used to identify genes expressed selectively in skeletal muscle. The Protein Atlas (https://www.proteinatlas.org/) was used to investigate both mRNA and protein expression in normal and cancerous samples for specific transcripts. The in silico surfaceome (http://wlab.ethz.ch/surfaceome/) (Bausch-Fluck et al., 2018) was used to predict the hierarchy of cell surface protein expression for ASPS-1. In detail, a file containing the published human surfaceome (table_S3_surfaceome.xlsx) was downloaded from http://wlab.ethz.ch/surfaceome/ and merged, using MS excel, with the list of differentially expressed ASPS-1 transcripts. Transcripts appearing in both datasets were then extracted and sorted according to ASPS-1 expression. The VENN diagram utility InteractiVenn (http://www.interactivenn.net/) (Heberle et al., 2015) was used to determine the extent of overlap between the 4 different experimental approaches.
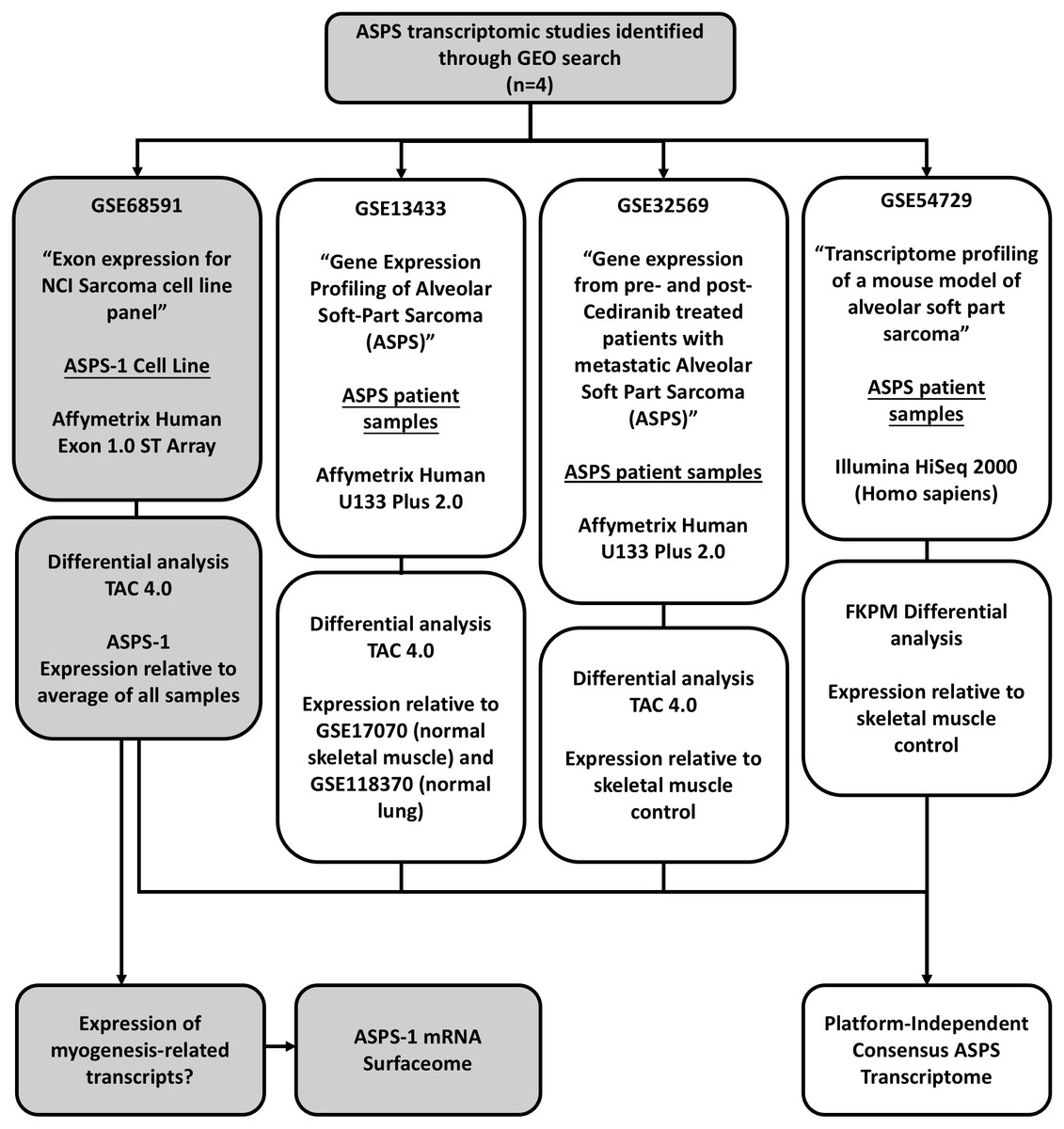
Figure 1: Study design.
This study focuses on the analysis of four publicly available GEO gene-expression datasets. GSE68591 comprises exon level expression data for the NCI sarcoma cell line panel, which includes data for the ASPS cell line ASPS-1. The remaining three studies comprise transcriptomic data for ASPS patient samples. GSE13433 comprises Affymetrix U133 plus 2.0 microarray data from our initial gene expression study of seven ASPS patients. GSE32569 uses the same array platform to study ASPS patient sample responses to Cedirinib. Lastly, GSE54729 comprises Illumina HISeq 2000 RNA-seq data for ASPS patient samples generated as part of an ASPS mouse modeling study. These data were re-analyzed using appropriate controls in order to generate a consensus transcriptome and gain insights into ASPS pathobiology.{kind=link}
Results and Discussion
In the first analysis, ASPS-1 exon-level data from the NCI sarcoma cell line panel was analyzed relative to all samples (cancer and normal). Results shown in Table 1 list the top fifty transcripts over-expressed in ASPS-1 relative to the average (Data S1). Results demonstrated that crystallin alpha beta (CRYAB) mRNA showed the highest expression in ASPS-1 relative to the average (227 fold). Metallothionein 1G (MT1G) was the next most elevated (202 fold). Following this were several lower-abundance transcripts coding for C7orf69, Synaptic Vesicle Glycoprotein 2B (SV2B) and Germinal Center-Associated Signaling and Motility-Like Protein (GCSAML). In common with the initial published report of ASPS-1, results also confirmed expression of GPNMB (Kenney et al., 2011). Likewise, ASPS-1 had some of the highest levels of MET, and VEGFR2 in the panel, both of which are previously noted characteristics of disease (Jun et al., 2010; Stockwin et al., 2009; Tsuda et al., 2007). This analysis served to confirm the ASPS origin of ASPS-1 as outlined in Kenney et al. (2011).
Following this, ASPS-1 was subjected to hierarchical clustering relative to the entire NCI sarcoma cell line panel (Fig. 2A). Results showed that ASPS-1 was an outlier that did not closely align with any lines/tissues in the panel. Those specimens with the nearest similarity are shown in Fig. 2B; this includes normal cell/tissues such as knee articular chondrocytes, dermal fibroblasts, skeletal muscle, mesenchymal stem cells and osteoblasts. Cancer line relatives included the spindle cell sarcoma Hs 132.T, the fibrosarcomas Hs 93.T and Hs 414.T along with the chondrosarcoma Hs 819. In order to define which changes were guiding this clustering, several breakout analyses were performed. Over-expressed transcripts found in the nearest cluster group and ASPS-1 included GPNMB, CRYAB, FABP3, and CTSK, which are markers of mesenchymal cells (Debnath et al., 2018; Kulterer et al., 2007; Wang Jr et al, 2014; Yu et al., 2016). Transcripts expressed only in ASPS-1 relative to the nearest cluster included SEPT3, C7orf69, MT1G, and ACP5, many of which are implicated in the differentiation of mesenchymal stromal cells (Dohi et al., 2005; Hayman et al., 2000; Moller et al., 2018; Tan et al., 2015). Lastly, the nearest cluster group could be distinguished from ASPS-1 through higher expression of the canonical fibroblast markers GREM1, LOX, THY1, and POSTN (Hortells, Johansen & Yutzey, 2019; Jiang & Rinkevich, 2018; Karagiannis et al., 2012). These data point towards a mesenchymal stromal origin that had not undergone significant fibroblast lineage specialization.
| Gene symbol | Affy ID | Description | FC ASPS-1 vs. Av |
|---|---|---|---|
| CRYAB; FDXACB1 | 3391149 | crystallin alpha B; ferredoxin-fold anticodon binding domain containing 1 | 227.1 |
| MT1G | 3692999 | metallothionein 1G | 202.9 |
| C7orf69 | 3000905 | chromosome 7 open reading frame 69 | 202.0 |
| GCSAML | 2390102 | germinal center-associated, signaling and motility-like | 177.6 |
| SV2B | 3608638 | synaptic vesicle glycoprotein 2B | 132.6 |
| ADGRL4 | 2419432 | adhesion G protein-coupled receptor L4 | 129.9 |
| PLA2G7 | 2955827 | phospholipase A2, group VII | 93.5 |
| SULT1C2 | 2498911 | sulfotransferase family 1C member 2 | 91.3 |
| SLN | 3389954 | sarcolipin | 88.9 |
| CFAP61 | 3878972 | cilia and flagella associated protein 61 | 85.2 |
| PPEF1 | 3970714 | protein phosphatase, EF-hand calcium binding domain 1 | 82.4 |
| ACP5 | 3851072 | acid phosphatase 5, tartrate resistant | 79.8 |
| CD36 | 3010503 | CD36 molecule (thrombospondin receptor) | 79.1 |
| PPARGC1A | 2763550 | peroxisome proliferator-activated receptor gamma, coactivator 1 alpha | 78.9 |
| ASB11 | 4000485 | ankyrin repeat and SOCS box containing 11, E3 ubiquitin protein ligase | 78.1 |
| BMP5 | 2958172 | bone morphogenetic protein 5 | 74.9 |
| PRUNE2 | 3210616 | prune homolog 2 (Drosophila) | 72.3 |
| SUCNR1 | 2648098 | succinate receptor 1 | 66.5 |
| PSG9 | 3864286 | pregnancy specific beta-1-glycoprotein 9 | 66.0 |
| CKMT2 | 2818035 | creatine kinase, mitochondrial 2 (sarcomeric) | 61.7 |
| DPP4 | 2584018 | dipeptidyl-peptidase 4 | 61.2 |
| ABCB1 | 3060182 | ATP binding cassette subfamily B member 1 | 59.7 |
| SCIN | 2990404 | scinderin | 58.7 |
| FABP3 | 2404418 | fatty acid binding protein 3, muscle and heart | 55.8 |
| PRUNE2 | 3210497 | prune homolog 2 (Drosophila) | 55.7 |
| SLC27A2 | 3593575 | solute carrier family 27 (fatty acid transporter), member 2 | 54.7 |
| SLCO4C1 | 2869096 | solute carrier organic anion transporter family, member 4C1 | 51.9 |
| PSG11,5,4,2 | 3863929 | pregnancy specific beta-1-glycoprotein 11,5,4,2 | 50.4 |
| PRLR | 2853102 | prolactin receptor | 47.9 |
| NPY6R | 2830450 | neuropeptide Y receptor Y6 (pseudogene) | 44.6 |
| ANXA3 | 2732844 | annexin A3 | 43.2 |
| TRPC7 | 2876793 | transient receptor potential cation channel, subfamily C, member 7 | 42.9 |
| CD5L | 2439138 | CD5 molecule-like | 41.0 |
| AKR1C2 | 3274758 | aldo-keto reductase family 1, member C2 | 40.9 |
| GPNMB | 2992814 | glycoprotein (transmembrane) nmb | 40.3 |
| IL13RA2 | 4018729 | interleukin 13 receptor, alpha 2 | 39.1 |
| LRRC39 | 2425173 | leucine rich repeat containing 39 | 38.8 |
| CST1 | 3901361 | cystatin SN | 37.4 |
| CDH7 | 3792273 | cadherin 7, type 2 | 36.8 |
| DOK5 | 3889833 | docking protein 5 | 35.1 |
| SEPT3; WBP2NL | 3947227 | septin 3; WBP2 N-terminal like | 34.8 |
| GCNT3 | 3596147 | glucosaminyl (N-acetyl) transferase 3, mucin type | 34.8 |
| ENPP5 | 2955673 | ectonucleotide pyrophosphatase/phosphodiesterase 5 (putative) | 34.7 |
| DOK3 | 2888879 | docking protein 3 | 34.2 |
| LCP1 | 3512874 | lymphocyte cytosolic protein 1 (L-plastin) | 34.0 |
| CDA | 2324084 | cytidine deaminase | 33.4 |
| KLHL4 | 3983324 | kelch-like family member 4 | 33.3 |
| CTSK | 2434609 | cathepsin K | 31.5 |
| LIPC | 3595691 | lipase, hepatic | 31.0 |
| RPSA | 3827218 | ribosomal protein SA | 30.0 |
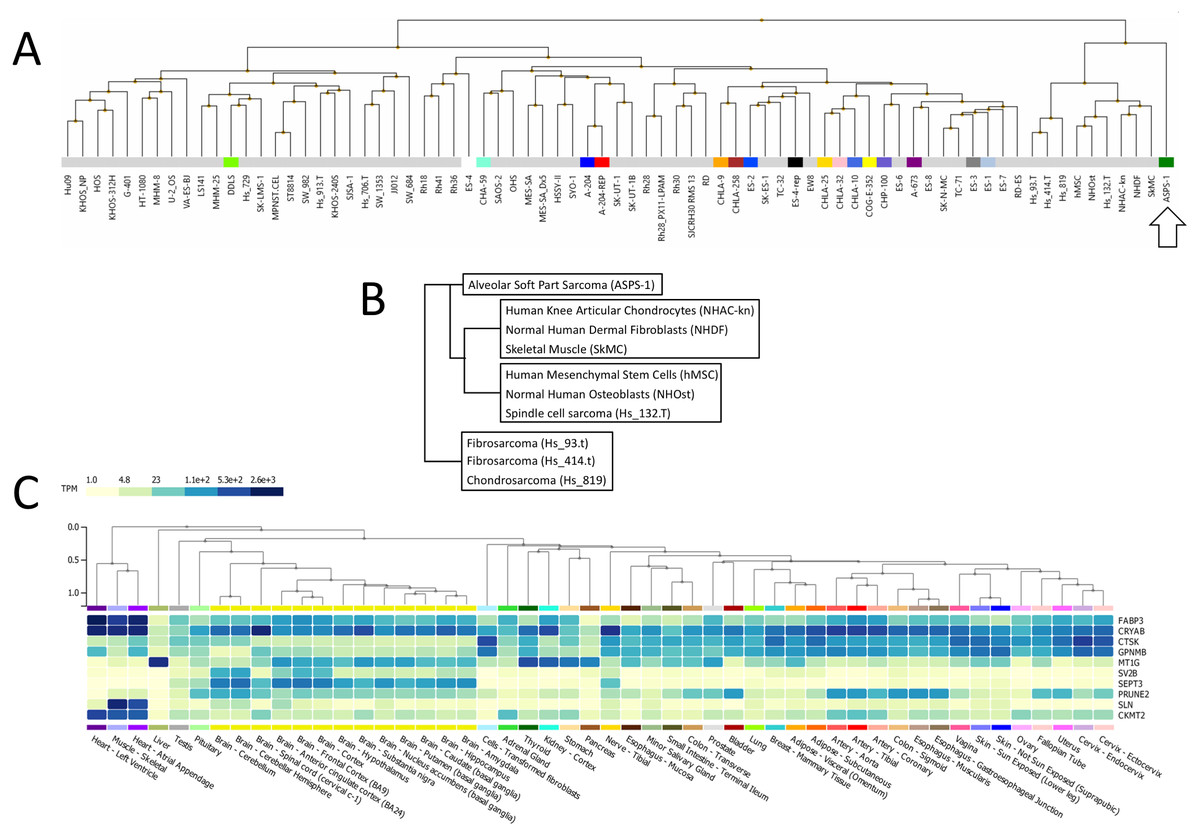
Figure 2: Analysis of ASPS-1 transcriptomic data.
(A) Heirarchical clustering of the NCI sarcoma cell line panel exon array data (B) tissue or tumor derivation for cluster nearest ASPS-1. (C) GTEX tissue of origin analysis for top fifty ASPS-1 transcripts.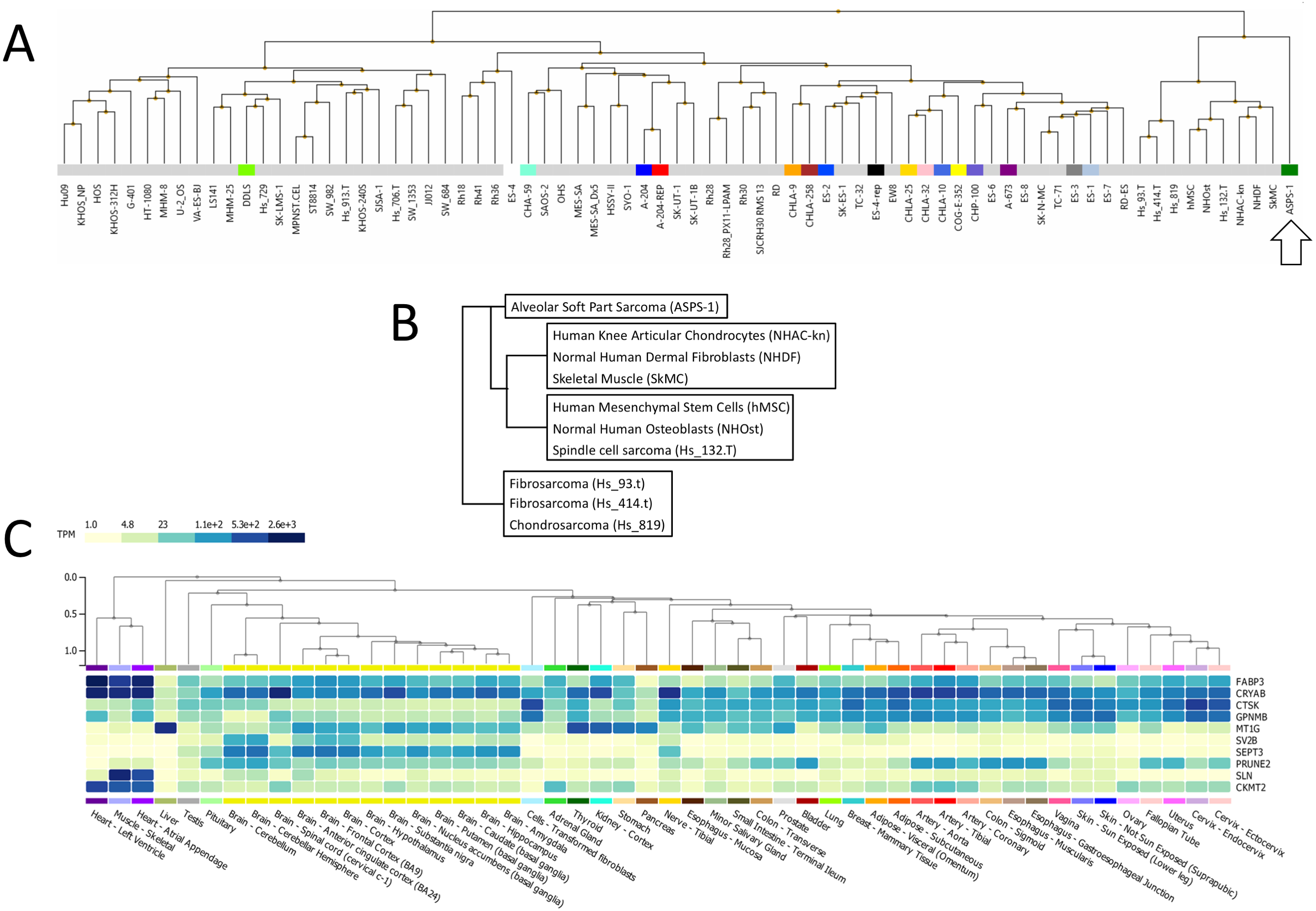{kind=link}
The Genotype-Tissue Expression (GTEx) portal (https://www.gtexportal.org/) was then used in order to determine whether the ASPS-1 expression data suggested a tissue of origin. Results obtained using the top fifty over-expressed transcripts suggested that cardiac/skeletal muscle was a likely origin through expression of transcripts such as CRYAB, FABP3, SLN, and CKMT2 (Fig. 2C). The GTEX database also lists the top 50 transcripts expressed by normal skeletal muscle, where 28 (e.g., creatine kinase M-Type CKM and myoglobin MB) have a considerable degree of muscle-specificity. Results showed that only two transcripts from this set, EEF1A2 and SLN are over-expressed in ASPS-1. As a consequence, we then undertook an expansive study of whether ASPS-1 had any hallmarks of myogenic differentiation. Figure 3 plots raw expression data for all cell lines in the sarcoma panel, where lines are grouped according to histology. Results showed that ASPS-1, unlike some rhabdosarcoma lines, did not express myogenic regulatory factor mRNAs including MYF5, MYF6, MYOD1, MYOG, and similarly did not express PAX3 and PAX7. Taken together these observations suggest that although ASPS-1 has hallmarks of a muscle resident cell, it had not undergone myogenic differentiation.
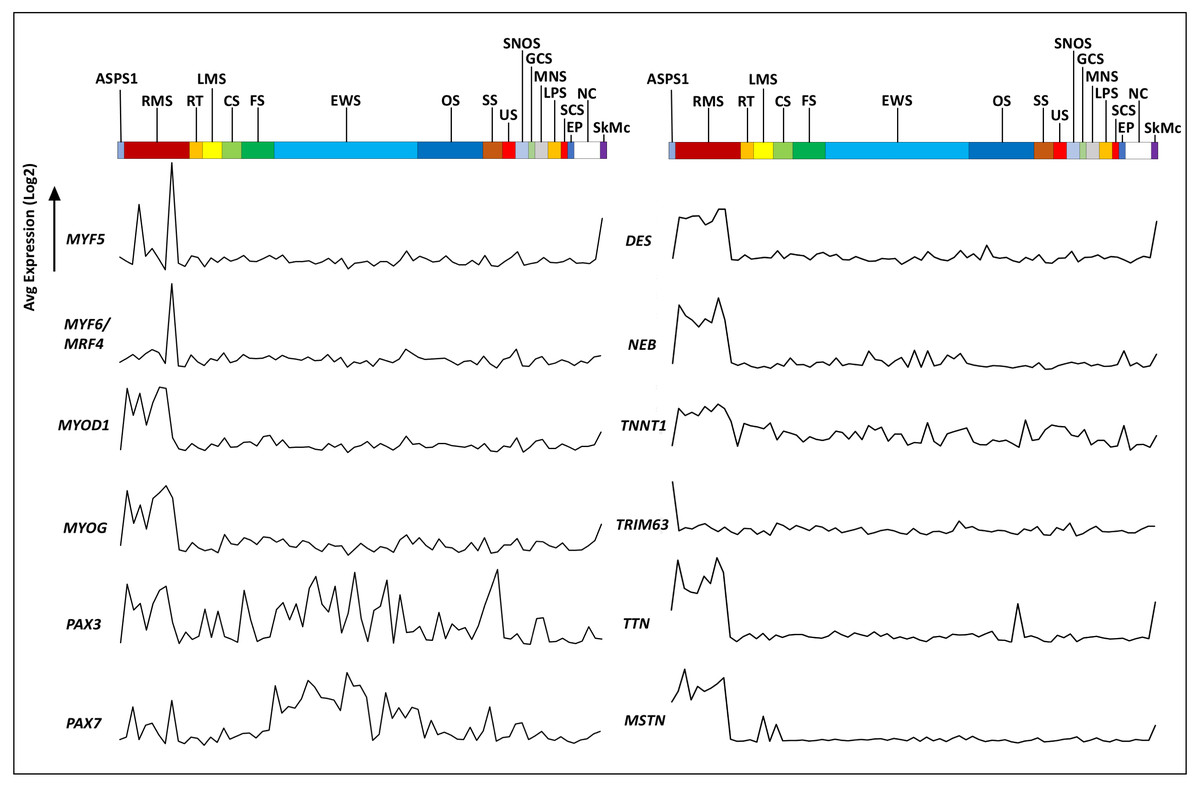
Figure 3: Expression of myogenesis-related transcripts in ASPS-1 relative to the other sarcoma cell lines.
Cell lines in the NCI sarcoma panel were segregated according to disease type and the average of exon expression data plotted for transcripts encoding myogenesis-related transcription factors and muscle structural proteins. ASPS-1 data is shown first; then RMS, Rhabdomyosarcoma; RT, Rhabdoid tumor; LMS, Leiomyosarcoma; CS, Chondrosarcoma; FS, Fibrosarcoma; EWS, Ewings Sarcoma; OS, Osteosarcoma; SS, Synovial Sarcoma; US, Uterine Sarcoma; SNOS, Sarcoma not otherwise specified; GCS, Giant cell sarcoma; MNS, Malignant peripheral nerve sheath; LPS, Liposarcoma; SPS, Spindle cell sarcoma; EP, Epithelioid; NC, Normal cells; SkMc, skeletal muscle cells. For transcripts; MYF5, Myogenic Factor 5, MYF6, Myogenic Factor 6, MYOD1, Myogenic Differentiation 1, MYOG = Myogenin, PAX3, Paired Box 3, PAX7 = Paired Box 7. Structural proteins; DES, desmin, NEB, nebulin, TNNT1, Troponin T1 - Slow Skeletal Type, TRIM63, Tripartite Motif Containing 63, TTN, titin and MSTN, Myostatin. Transcripts evaluated but not shown included; EYA1, LBX1, MEF2B, MEOX2, MITF, MSX1, PITX1, SIM2, SIX1, SIX4, TFE3 and TFEB.{kind=link}
These data also provide a unique opportunity to assess the potential cell surface phenotype of ASPS-1. Bausch-Fluck et al. (2018) identified 2886 proteins that are known, or are predicted by machine learning, to be expressed on the cell surface . Here, ASPS-1 raw microarray data was filtered for these targets and the list sorted in terms of expression. The resultant ASPS-1 ‘surfaceome’ is shown in Data S2. As could be anticipated, GPNMB was the highest expressed mRNA; followed by novel surface makers such as the glutamate transporter SLC38A1 and the amyloid beta (A4) precursor protein APP. In terms of CD antigens, the following mRNAs were highly expressed in ASPS-1; CD9 (TSPAN29), CD26 (DPP4), CD49C (ITGA3), CD54 (ICAM1), CD63 (TSPAN30), CD68 (SCARD1), CD130 (IL6ST), CD146 (MCAM), CD147 (BSG), CD151 (SFA-1), CD166 (ALCAM), CD222 (IGF2R), CD230 (PRP), CD236 (GPC), CD243 (ABCB1), and CD325 (CDHN).
Many of these observations are also compatible with a mesenchymal stromal cell. For example, CD9 (TSPAN29) and CD243 (ABCB1), although widely expressed, are found to varying degrees on MSC (Islam et al., 2005; Kim et al., 2007). CD49C (ITGA3) and CD151 (SFA-1) are both markers of chondrogenic differentiation in MSC (Grogan et al., 2007; Lee et al., 2009a) . Expression of CD54 (ICAM1) can be induced in MSC (Ren et al., 2010), CD63 (TSPAN30) is expressed by bone marrow MSC (McBride et al., 2017), CD68 (SCARD1) expression has been shown on MSC from human umbilical cord (Rocca, Anzalone & Farina, 2009) and CDH2 is a regulator of mesenchymal stem cell fate (Alimperti & Andreadis, 2015). Exosomes expressing Basigin, BSG (CD147), have been shown to promote angiogenesis in MSC (Vrijsen et al., 2016). CD147 is also a major constituent of the pre-crystalline granules present in ASPS (Ladanyi et al., 2002). Expression of Melanoma Cell Adhesion Molecule, MCAM (CD146), mRNA provides strong evidence for an MSC derivation, with several studies demonstrating an important role for this molecule in MSC maintenance and differentiation (Covas et al., 2008; Espagnolle et al., 2014; Jin et al., 2016; Stopp et al., 2013). Likewise, Activated Leukocyte Cell Adhesion Molecule (ALCAM, CD166), is a recognized marker of MSC and implicated in osteogenesis (Bruder et al., 1998; Hu et al., 2016).
In summary, the results presented here demonstrate that the ASPS-1 transcriptome is unique amongst the NCI sarcoma panel, where the closest relatives are normal mesenchymal cells and connective tissue sarcomas. Although ASPS-1 has an expression signature with some similarity to skeletal/cardiac muscle tissue, markers of myogenesis were not detected in this cell line. Furthermore, the ASPS-1 surfaceome does not immediately speak to a tissue derivation but suggests an undifferentiated mesenchymal state.
The next phase of the project involved re-analyzing microarray and RNA-seq data from ASPS tumor resections. GSE13433 comprises microarray data (Affymetrix U133 plus 2.0) for seven patients with primary or metastatic ASPS (Stockwin et al., 2009). In the original study, universal RNA (representing a collection of adult human tissues) was used as a reference. However, patient samples 1,3, 5 and 6 were obtained from skeletal muscle biopsies whereas samples 4 and 7 were isolated from lung. As a consequence, microarray data from normal skeletal muscle and lung represent more appropriate controls. Therefore, skeletal muscle arrays were obtained from GSE17070 and normal lung samples from GSE118370. Patient 2 data, derived from the mandible, was excluded from the analysis for lack of an appropriate control. Two lists of differentially expressed transcripts were then generated for patients 1,3,5,6 vs. skeletal muscle and 4,7 vs. normal lung. Results from these two experiments were largely concordant with the profiles obtained in the original study (Stockwin et al., 2009). However, in Stockwin et al. muscle-differentiation associated transcripts ITGB1BP3/MIBP, MYF5 and MYF6 were identified as overexpressed. In our analysis, only MYF6 was identified, and only in the experiment involving patients 4 and 7 vs. normal lung; supporting our inference that the published study over-emphasized myogenic differentiation in patient ASPS.
GSE32569 is a similar ASPS dataset where U133 plus 2.0 microarrays were generated from patients treated with Cediranib (Kummar et al., 2013). We undertook to use this data to generate a list of differentially expressed transcripts from pre-treatment arrays relative to GSE17070 skeletal muscle samples. Results again showed a similar profile to that obtained from the analysis of GSE13433. The final experiment was performed using RNA-seq data from GSE54729. In this published study, data was generated from five human ASPS tumor samples in order to compare the transcriptome with five mouse tumors generated through ectopic expression of ASPSCR1-TFE3 (Goodwin et al., 2014). Here, FKPM values for the five human tumors and three skeletal muscle controls were used to generate a list of differentially expressed transcripts. The top fifty upregulated transcripts generated from each of the four experiments using GSE13433, GSE32569 and GSE54729 are shown in Table 2. Meta-analysis of data from these four in vivo studies had considerable overlap, emphasizing the consistent upregulation of mRNAs such as GPNMB, ABCB5, PSG9, CYP17A1, PRL, SULT1C2, and SV2B. As with ASPS-1, over-expression of myogenic regulatory factor mRNA was not consistently seen in any of the experiments involving patient samples (results not shown). Lastly, lists of differentially expressed genes from both ASPS-1 and patient sample experiments were combined (at a five-fold cut-off 2 ), and a VENN diagram generated in order to determine the extent of overlap (Fig. 4). Results demonstrated that twenty-five transcripts were elevated in all of the meta-analyses, whereas seventy-three were expressed at the intersection between all in vivo analyses.
| GSE13433 Patients 1,3,5,6 vs. Skeletal muscle | GSE13433 Patient 4,7 vs. Normal lung | GSE32569 Pre-treatment ASPS vs. Skeletal muscle | GSE54729 Patients vs. Skeletal muscle | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AFFY ID | FC | GENE ID | AFFY ID | FC | GENE ID | AFFY ID | FC | GENE ID | Ensembl ID | FC | Gene ID |
| 205502_at | 639.7 | CYP17A1 | 207733_x_at | 608.6 | PSG9 | 211470_s_at | 573.8 | SULT1C2 | ENSG00000159871 | 100.9 | LYPD5 |
| 212992_at | 531.8 | AHNAK2 | 209594_x_at | 601.3 | PSG9 | 205502_at | 420.6 | CYP17A1 | ENSG00000183979 | 423.1 | NPB |
| 205445_at | 434.1 | PRL | 205445_at | 572.3 | PRL | 1554018_at | 375.0 | GPNMB | ENSG00000198203 | 237.6 | SULT1C2 |
| 240717_at | 419.0 | ABCB5 | 205502_at | 537.1 | CYP17A1 | 205342_s_at | 367.4 | SULT1C2 | ENSG00000148795 | 1700.4 | CYP17A1 |
| 205342_s_at | 338.7 | SULT1C2 | 1555786_s_at | 510.2 | LINC00520 | 210809_s_at | 365.1 | POSTN | ENSG00000146678 | 200.6 | IGFBP1 |
| 201850_at | 258.8 | CAPG | 206224_at | 467.6 | CST1 | 238720_at | 351.1 | LOC101927057 | ENSG00000172179 | 357.1 | PRL |
| 211470_s_at | 252.9 | SULT1C2 | 223572_at | 437.6 | HHATL | 210587_at | 341.2 | INHBE | ENSG00000169006 | 142.2 | NTSR2 |
| 238720_at | 248.1 | LOC101927057 | 240717_at | 346.4 | ABCB5 | 206214_at | 262.4 | PLA2G7 | ENSG00000101197 | 113.9 | BIRC7 |
| 1554018_at | 240.8 | GPNMB | 208555_x_at | 322.5 | CST2 | 212992_at | 259.6 | AHNAK2 | ENSG00000170369 | 345.4 | CST2 |
| 205302_at | 207.6 | IGFBP1 | 236972_at | 277.8 | TRIM63 | 1565162_s_a | 241.7 | MGST1 | ENSG00000100167 | 143.8 | SEPT3 |
| 204638_at | 198.2 | ACP5 | 1553663_a_at | 259.2 | NPB | 229831_at | 228.7 | CNTN3 | ENSG00000204632 | 396.4 | HLA-G |
| 206899_at | 170.8 | NTSR2 | 205302_at | 255.8 | IGFBP1 | 200832_s_at | 225.4 | SCD | ENSG00000146070 | 147.4 | PLA2G7 |
| 210587_at | 170.4 | INHBE | 221051_s_at | 200.5 | NMRK2 | 206899_at | 168.5 | NTSR2 | ENSG00000110492 | 313.0 | MDK |
| 212805_at | 157.6 | PRUNE2 | 206239_s_at | 192.8 | SPINK1 | 205302_at | 159.6 | IGFBP1 | ENSG00000227925 | 134.1 | LOC101929771 |
| 209875_s_at | 150.3 | SPP1 | 210587_at | 184.3 | INHBE | 227180_at | 157.3 | ELOVL7 | ENSG00000170373 | 455.2 | CST1 |
| 1557636_a_at | 145.1 | C7orf57 | 206899_at | 171.9 | NTSR2 | 219648_at | 149.3 | MREG | ENSG00000225328 | 118.5 | LINC01594 |
| 210809_s_at | 144.4 | POSTN | 205551_at | 169.9 | SV2B | 210397_at | 148.0 | DEFB1 | ENSG00000118785 | 260.1 | SPP1 |
| 221577_x_at | 134.0 | GDF15 | 219106_s_at | 149.9 | KLHL41 | 209875_s_at | 147.2 | SPP1 | ENSG00000139269 | 137.6 | INHBE |
| 221008_s_at | 133.2 | ETNPPL | 206799_at | 146.3 | SCGB1D2 | 557636_a_a | 145.3 | C7orf57 | ENSG00000102575 | 190.9 | ACP5 |
| 206214_at | 130.1 | PLA2G7 | 229052_at | 143.1 | ANKRD23 | 205825_at | 139.0 | PCSK1 | ENSG00000185518 | 114.7 | SV2B |
| 209035_at | 122.3 | MDK | 221008_s_at | 142.4 | ETNPPL | 219073_s_at | 138.2 | OSBPL10 | ENSG00000205336 | 129.2 | ADGRG1 |
| 202450_s_at | 121.7 | CTSK | 205342_s_at | 125.6 | SULT1C2 | 205343_at | 132.8 | SULT1C2 | ENSG00000185567 | 246.4 | AHNAK2 |
| 200832_s_at | 121.0 | SCD | 212805_at | 124.7 | PRUNE2 | 555778_a_a | 127.5 | POSTN | ENSG00000136235 | 2756.8 | GPNMB |
| 212806_at | 119.7 | PRUNE2 | 1564758_at | 115.7 | LOC643659 | 221008_s_at | 126.1 | ETNPPL | ENSG00000042493 | 172.1 | CAPG |
| 230067_at | 116.8 | FAM124A | 229831_at | 114.5 | CNTN3 | 231736_x_at | 116.1 | MGST1 | ENSG00000143387 | 1718.2 | CTSK |
| 208555_x_at | 116.8 | CST2 | 209738_x_at | 113.4 | PSG6 | 212805_at | 115.1 | PRUNE2 | ENSG00000030582 | 707.5 | GRN |
| 225275_at | 115.8 | EDIL3 | 233389_at | 107.5 | CFAP61 | 558378_a_a | 108.4 | AHNAK2 | ENSG00000107317 | 354.1 | PTGDS |
| 229831_at | 112.1 | CNTN3 | 212992_at | 106.9 | AHNAK2 | 218292_s_at | 98.4 | PRKAG2 | ENSG00000106617 | 161.8 | PRKAG2 |
| 227404_s_at | 111.2 | EGR1 | 233238_s_at | 106.0 | CTB-12O2.1 | 221577_x_at | 94.6 | GDF15 | ENSG00000216490 | 275.5 | IFI30 |
| 212841_s_at | 109.4 | PPFIBP2 | 1569072_s_at | 102.9 | ABCB5 | 218404_at | 94.0 | SNX10 | ENSG00000183696 | 183.0 | UPP1 |
| 216834_at | 108.8 | RGS1 | 206994_at | 87.9 | CST4 | 233748_x_at | 90.8 | PRKAG2 | ENSG00000110092 | 192.1 | CCND1 |
| 208792_s_at | 106.1 | CLU | 239205_s_at | 84.7 | CR1; CR1L | 224918_x_at | 89.1 | MGST1 | ENSG00000212443 | 410.2 | SNORA53 |
| 223362_s_at | 105.2 | SEPT3. | 217871_s_at | 84.7 | MIF | 212070_at | 86.0 | ADGRG1 | ENSG00000185585 | 105.1 | OLFML2A |
| 208791_at | 92.7 | CLU | 226086_at | 83.8 | SYT13 | 205551_at | 85.1 | SV2B | ENSG00000130203 | 285.1 | APOE |
| 1558846_at | 92.7 | PNLIPRP3 | 213175_s_at | 82.5 | SNRPB | 244444_at | 84.5 | PKD1L2 | ENSG00000111412 | 119.3 | C12orf49 |
| 230746_s_at | 92.3 | N/A | 221523_s_at | 81.9 | RRAGD | 208965_s_at | 83.6 | IFI16 | ENSG00000206503 | 1571.8 | HLA-A |
| 218292_s_at | 89.1 | PRKAG2 | 243167_at | 77.0 | ABCB5 | 208146_s_at | 82.8 | CPVL | ENSG00000106066 | 239.3 | CPVL |
| 1565162_s_at | 88.4 | MGST1 | 206372_at | 74.4 | MYF6 | 226847_at | 82.8 | FST | ENSG00000138131 | 108.9 | LOXL4 |
| 205825_at | 83.7 | PCSK1 | 209875_s_at | 70.5 | SPP1 | 223484_at | 80.3 | C15orf48 | ENSG00000118508 | 104.3 | RAB32 |
| 226372_at | 82.7 | CHST11 | 244444_at | 67.3 | PKD1L2 | 234983_at | 78.9 | C12orf49 | ENSG00000174080 | 454.1 | CTSF |
| 202503_s_at | 82.6 | KIAA0101 | 205862_at | 65.9 | GREB1 | 240717_at | 78.3 | ABCB5 | ENSG00000169116 | 207.1 | PARM1 |
| 205343_at | 82.0 | SULT1C2 | 222379_at | 65.8 | KCNE4 | 229177_at | 78.1 | C16orf89 | ENSG00000120885 | 187.0 | MIR6843 |
| 205551_at | 81.7 | SV2B | 1554371_at | 60.2 | PKD1L2 | 205844_at | 75.8 | VNN1 | ENSG00000214435 | 114.9 | AS3MT |
| 1569072_s_at | 81.5 | ABCB5 | 205825_at | 58.2 | PCSK1 | 238376_at | 75.5 | LOC100505564 | ENSG00000130208 | 134.3 | APOC1 |
| 227180_at | 79.8 | ELOVL7 | 222714_s_at | 55.6 | LACTB2 | 205445_at | 73.0 | PRL | ENSG00000100644 | 335.4 | HIF1A |
| 231736_x_at | 79.5 | MGST1 | 218619_s_at | 54.6 | SUV39H1 | 242340_at | 71.7 | N/A | ENSG00000135047 | 458.4 | CTSL |
| 202037_s_at | 76.5 | SFRP1 | 236523_at | 54.3 | LOC285556 | 204285_s_at | 71.3 | PMAIP1 | ENSG00000144136 | 134.6 | SLC20A1 |
| 219648_at | 74.5 | MREG | 1557636_a_at | 53.7 | C7orf57 | 204466_s_at | 71.3 | SNCA | ENSG00000101846 | 109.4 | STS |
| 206685_at | 71.8 | HCG4 | 212070_at | 52.9 | ADGRG1 | 203767_s_at | 70.7 | STS | ENSG00000111775 | 242.3 | COX6A1 |
| 210397_at | 71.0 | DEFB1 | 204830_x_at | 52.8 | PSG5 | 222872_x_at | 70.5 | NABP1 | ENSG00000089101 | 164.4 | CFAP61 |
An exploration of the twenty-five conserved transcripts in the context of stem cell biology provides further insights into MSC lineage potential. For example; angiopoietin Like 2 (ANGPTL2) is a regulator of stem cell adipogenesis, chondrogenesis and osteogenesis (Takano et al., 2017; Tanoue et al., 2018). Expression of Cathepsin K (CTSK) is compelling given that in mice CTSK-mGFP cells label the periosteal mesenchyme and have been used to identify periosteal stem cells (Debnath et al., 2018). As previously noted, Dipeptidyl Peptidase 4 (DPP4), also known as CD26, marks mesenchymal preadipocyte progenitors (Merrick et al., 2019). Glycoprotein Nmb (GPNMB) should be recognized as the prototypic cell surface marker for ASPS. As stated, GPNMB is recognized as a marker of mesenchymal cells (Kuci et al., 2019). Interestingly, within protein atlas, the cell line designated ‘ASC diff’, a differentiated adipose-derived mesenchymal stem cell line has the highest expression of GPNMB and also expresses TRIM63, CRYAB, FABP3, and CTSK. These observations would appear to favor the concept that ASPS resembles an MSC capable of adipogenic, chondrogenic or osteogenic differentiation.
Several inferences can also be made for the seventy-three conserved transcripts identified in all ASPS patient experiments. The multi-drug resistance transporter ABCB5, in addition to being expressed by melanoma, also defines a subset of MSC in the cornea and skin (Frank et al., 2003; Ksander et al., 2014; Vander Beken et al., 2019). The hormone prolactin (PRL) has been shown to stimulate proliferation of MSC and also to direct chondrogenic and ostegenic differentiation (Ogueta et al., 2002; Seriwatanachai, Krishnamra & Charoenphandhu, 2012; Surarit, Krishnamra & Seriwatanachai, 2016). Increased expression of the growth factor midkine (MDK) has been noted in previous ASPS gene expression studies and is an MSC survival factor (Stockwin et al., 2009; Zhao et al., 2014). Upregulation of hypoxia-related transcripts such as HIF1A suggests that this pathway is active in ASPS and, although ubiquitous, HIF1A plays an important role in the control of multipotency for MSC (Palomaki et al., 2013). It was similarly interesting that the ASPS patient experiments showed increased expression of THY1 (CD90) relative to control samples. This target is regarded as a classical marker of MSC and has recently been shown to promote osteogenic differentiation over an adipogenic fate (Saalbach & Anderegg, 2019). In summary, re-analysis of microarray and RNA-seq data for ASPS patient samples yielded transcriptomes with considerable overlap between studies irrespective of platform technology; and the final consensus ASPS transcriptome resembles an undifferentiated mesenchymal stromal cell.

Figure 4: VENN diagram showing overlap between analyses.
Lists of over-expressed transcripts (five-fold cut off) were used to determine extent of overlap between the five datasets. The number of differentially expressed transcripts at five-fold is underlined. Callouts show the 25 transcripts over-expressed in all experiments and the 73 found in all ASPS patient analyses.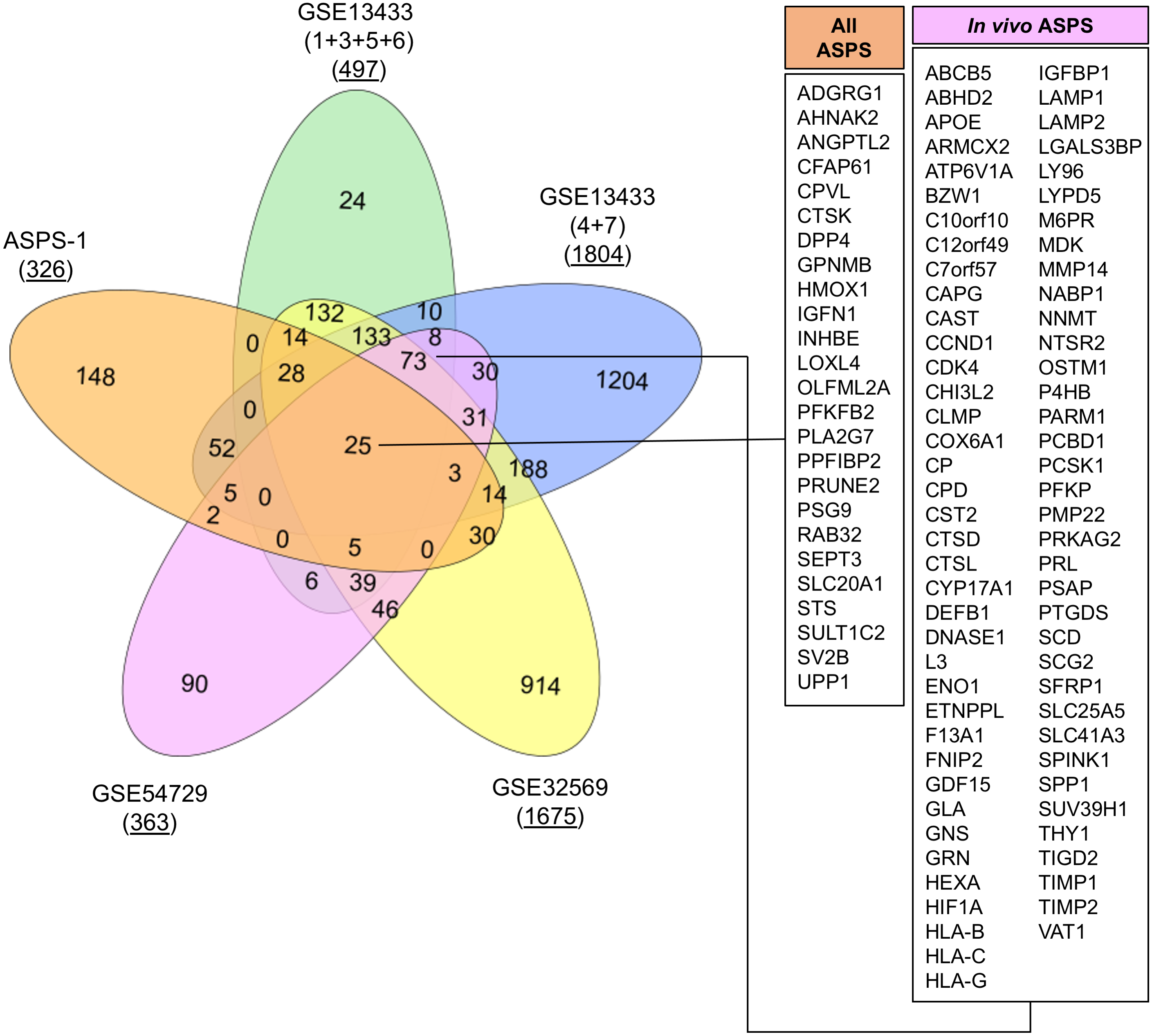{kind=link}
Conclusions
Alveolar-soft part sarcoma is an example of a malignancy that has, despite several immunohistochemical and genomics studies, evaded classification (Fisher & Reidbord, 1971; Folpe & Deyrup, 2006; Gomez et al., 1999; Goodwin et al., 2014; Hoshino et al., 2009; Mukai et al., 1983; Selvarajah et al., 2014; Stockwin et al., 2009; Tallini et al., 1994; Tanaka et al., 2017; Wang et al., 1996). This study was prompted by the public release of exon expression data for the cell line ASPS-1, which offers a unique opportunity to study ASPS in isolation (Kenney et al., 2011; Teicher et al., 2015). We were similarly interested in revisiting the genomic studies of ourselves and others to generate a consensus expression profile independent of platform technology.
The central finding of the current study was that the ASPS transcriptome is indicative of an undifferentiated mesenchymal stromal cell (MSC). Specifically, The ASPS-1 cell line exhibited a mesenchymal expression signature, where expression data clustered with normal and malignant mesenchymal cells within the NCI sarcoma cell line panel. The ASPS-1 surfaceome was similarly suggestive of an undifferentiated mesenchymal cell. Generation of an ASPS consensus transcriptome from previously reported patient studies highlighted the importance of targets such as GPNMB, ABCB5, CSTK, DPP4, BSG, ALCAM, PRL, and CDHN; all of which were consistent with an undifferentiated MSC. Conversely, the ASPS transcriptome lacked expression of myogenesis-related genes and did not feature transcripts indicative of neural, pericyte or endothelial differentiation.
MSC are found in most tissues, these cells are capable of multipotent differentiation into bone, muscle, cartilage, adipocytes, marrow stromal cells, tenocytes, fibroblasts, endothelial and neural cells (Caplan, 2007; Pittenger et al., 2019). Tissues maintain a pool of MSC, with varying degrees of specialization, ready to dynamically replenish differentiated cells in response to signals associated with growth, homeostasis or damage (Rubenstein et al., 2020). Prior to this study, ASPL-TFE3 had already been shown to immortalize embryonic mesenchymal cells (Tanaka et al., 2017). The suggestion that ASPS resembles a mesenchymal stromal progenitor provides a plausible explanation for the failure of previous studies to pinpoint cellular origin, given that the cell retains an undifferentiated state. Evidence from this study favors an MSC capable of adipogenic, osteogenic or chondrogenic differentiation, but not necessarily at the exclusion of other lineages.
If an MSC origin for ASPS could ultimately be confirmed, there would be important consequences for therapeutic development. Foremost is the suggestion that ASPS growth may be inhibited by factors that promote MSC differentiation. For example, several high-throughput studies have identified clinically relevant small molecules capable of promoting or inhibiting differentiation of MSC (Brey et al., 2011; Huang et al., 2008). Re-screening these compounds for their ability to inhibit the growth of ASPS-1 may yield clinically tractable candidates for ASPS treatment. From the authors perspective, the effect of HDAC inhibitors, steroids and retinoids on ASPS-1 growth are of particular interest (Lee et al., 2009b; Salloum, Rubin & Marra, 2013).
The findings of this study have a central caveat; all speculation regarding cellular origin must be moderated until the inference of ASPL-TFE3 is removed. Given the ability of this fusion protein to re-direct transcription, the observed transcriptomes may mask the true cellular origin. For example, GPNMB, CRYAB, CYP17A1, SULT1C2, UPP1 and SV2B have been shown to be up-regulated following expression of ASPL-TFE3 in inducible 293 cells (Kobos et al., 2013). Therefore, the current study only suggests that ASPS resembles an MSC and no firm conclusion can be made regarding origin. A straightforward approach to address this central question involves generating an ASPL-TFE3 knockout in ASPS-1 perhaps with re-introduction of wild-type TFE3 to maintain viability. The resultant line could then be characterized by RNA-seq and FACS phenotyping. These experiments could be accompanied by the addition of defined media to determine whether differentiation can be directed toward specific MSC lineages. In the interim, the data presented here provides a unified picture of ASPS mRNA expression, where considerable similarity with mesenchymal stromal progenitors is evident.