
Coupled influence of precipitation regimes and seedling emergence time on the reproductive strategy in Chloris virgata
- Published
- Accepted
- Received
- Academic Editor
- Enrique Mateos Naranjo
- Subject Areas
- Ecology, Plant Science, Climate Change Biology
- Keywords
- Reproduction, Seed size, Seed yield, Tiller trait
- Copyright
- © 2020 Wang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Coupled influence of precipitation regimes and seedling emergence time on the reproductive strategy in Chloris virgata. PeerJ 8:e8476 https://doi.org/10.7717/peerj.8476
Abstract
Precipitation regime and seedling emergence time both influence plant growth and reproduction. However, little attention has been given to the effects of these combined factors on the reproductive strategy of Chloris virgata, which is a vital species in Songnen grassland. Here, we simulated the changes in the precipitation regime and seedling emergence time to evaluate tiller traits and seed production. The results showed that tiller number behaved similarly among three precipitation regimes when sowed on 15 May (T1), while it increased significantly with precipitation regimes when sowed on 15 June (T2) and 15 July (T3). Tiller number decreased significantly with the seedling emergence time under the same water supply treatment. The proportional allocation of reproductive tiller number to total tiller number was significantly higher at T3 than at T1 and T2. Seed number remained similar under different precipitation regimes at T2 and T3, whereas it was significantly lower under low precipitation than under other water levels at T1. Seed number reached the maximum values at T2 under the same level of precipitation treatment. Seed size was significantly lower under low precipitation compared to other water supply treatments and the lowest values in seed size, about 0.5 mg, occurred at T2 under all the precipitation regimes. The lowest values in spike number were under low precipitation at all seedling emergence times. Seed yield exhibited similar trends with seed size under different precipitation regimes, while the greatest gains in these values were at T1 under all the precipitation regimes. Our findings showed that simulated precipitation regimes and seedling emergence time affected the reproductive strategy of C. virgata. Typical and high precipitation, as well as early seedling emergence, will improve the seed yield and seed quality in this species.
Introduction
Reproduction is one of the most important stages in a plant’s life history and is the result of natural selection. Plants exhibit different reproductive strategies in response to varied environments (Suonan et al., 2017). Therefore, reproductive strategy plays a major role in plant adaptation by maximizing the overall fitness of plants growing with variable resources (Bazzaz et al., 1987).
Seed yield, one of the vital components in plant restoration, can be determined by inflorescence number, seed number per inflorescence and seed size (Hebblethwaite, Wright & Noble, 1980; Wang et al., 2010). Therefore, environmental conditions experienced by individual plants that affect seed yield will ultimately influence vegetation restoration. Another vital factor for determining seedling establishment and regeneration is seed biology, which is thought to be an important contributor in understanding plant ecological processes within communities (Venable & Brown, 1988). Seed biology is mainly affected by seed genotype and parental environment (He et al., 2014). Maternal nutrition has been demonstrated as a vital factor that impacts the competitive ability of offspring (Galloway, 2001). Selection pressure for increased seed size may also arise because larger seeds are better at surviving hazards of seedling establishment such as nutrient shortages (Metcalfe & Grubb, 1997), water availability (Bidgoly et al., 2018; Müller et al., 2019) and temperature (Dewan et al., 2018). Consequently, studies of the reproductive strategies of plants are useful in understanding the adaptation of plants to their natural environment.
Degradation has been severe in the Songnen grassland of China including bare patches that have appeared in this region due to soil salinity and alkalinity. Chloris virgata Sw. is an annual tuft grass with substantial drought and alkali tolerance. Moreover, it is the first species to form a relatively stable and productive single-species dominant community (Zheng & Li, 1999) and is therefore a vital species to restore degraded semi-arid grassland.
Precipitation is one of the major factors that limits seed productivity in arid and semiarid areas. Inevitably, the precipitation regime may profoundly influence reproductive traits in perennial and annual grasses (Wang et al., 2010; Wang et al., 2018). Moreover, soil water content has been used to successfully predict seedling emergence (Ikeda et al., 2019). Spring precipitation effects on the vegetation spring phenology in arid and semi-arid regions (Shen et al., 2015). Consequently, there can be major impacts on seedling emergence time due to early or late spring precipitation in wet or dry years, with seedling emergence time influencing the plant’s fitness within its community (Du & Huang, 2008).
Therefore, the objective of this study was to investigate the reproductive traits of this important species in response to combined effects of precipitation regime and seedling emergence time. The findings on the reproductive ecology of this species will inform rational regulatory and management policy for grassland restoration.
Methods and Materials
Experimental design and treatment
The experiment was conducted at Jilin Jianzhu University, Jilin Province, China in 2016. C. virgata seeds were obtained from the Greenhouse of Jilin Jianzhu University, Jilin Province, China (44°33N, 123°82 31E) in 2015. Twenty seven PVC pipes, 25 cm diameter and 50 cm height, were filled with saline-sodic soils at different sowing dates in 2016. The soil was collected from the surface layer (0–20 cm) of a grassland near the Northeast Normal University field experiment station, in Changling, Jilin Province, China (123°44 0E, 44°44 0N, 167 m above sea level). The soil total nitrogen was 6.9%, organic carbon 0.4%, pH 8.6, electrical conductivity 91 µs cm−1 and field capacity of 200 g kg−1. And the maximum/minimum temperatures were 32/16 °C (day/night) during the treatment period from 15 June to 15 September. The soil-filled pipes were all buried in the soil and the top edge of each pipe and the soil surface were kept at the same level. Twenty seeds were sown on 15 May (T1), 15 June (T2) and 15 July (T3), respectively, to ensure three seedling emergence times (SETs). Most seedlings emerged 5 d after sowing and 10 uniform and robust seedlings were selected per pipe 10 d after sowing. Three treatments were used to simulate local precipitation regimes (PRs), corresponding to 120 (L: low, soil moisture was 6.0%), 200 (T: typical, soil moisture was 11.0%), and 280 (H: high, soil moisture was 19.0%) mm precipitation, and these were created by adding water every 5 d to the pipes from 30 d after seedling emergence (see Wang et al., 2018). The typical precipitation regime was based on the average amount and frequency of the local region’s precipitation (June–October) over the past 21 years in the Songnen grassland, where C. virgata is widely distributed. The low and high precipitation regimes were 40% lower and higher than the typical precipitation regime, based on data from the Songnen grassland in wet and dry years, respectively. For example, the plants were watered 12.5 mm 5 d intervals (The number of precipitation was 16 from 15 June to 15 September because most precipitations occurred in this period) under typical precipitation regime. Then the precipitation was calculated according to the precipitation depth and cross section of the pipe. There were 3 replicates for each precipitation level at each seedling emergence time, giving 30 individuals per treatment for determining tiller traits and seed production. There was a total of 27 pipes across all treatments for the three seedling emergence times and the pipes were arranged outdoors in a replicated randomized block design under a rain shelter made from transparent 0.2 mm polyethylene sheeting on 15th May, 2016.
Data collection
The plants from each treatment at each seedling emergence time were harvested in mid-September, when all C. virgata individuals were fully mature. Some pipes were damaged by locusts during the course of the experiment and were excluded from the analyses, giving 22–30 individuals. The numbers of reproductive and vegetative tillers of C. virgata were recorded for each individual in each pipe. One first-order tiller was selected randomly from each individual in each pipe. Then the numbers of mature seeds and unripe seeds were counted to obtain seed number. The number of left spikes from each plant were recorded and the spikes were kept in individual paper bags. Fifty seeds were selected randomly from each plant and dried at 65 °C for 48 h and weighed to determine mean seed size. The remaining seeds from each plant were dried at 65 °C for 48 h and weighed to obtain seed yield.
Data analysis
All data were tested for the normality and homogeneity and subjected to analyze by SPSS statistical software (version 17.0, SPSS Inc., Chicago, IL). One-way ANOVA was used to analyze variables such as tiller number, proportion of reproductive tiller number to total tiller number, seed number, spike number, seed size and seed yield. Two-way ANOVA was also used to analyze all data under different precipitation regimes over seedling emergence times. Differences under different precipitation regimes and seedling emergence times were analyzed using the least significant difference (l.s.d.) test.
Results
Vegetative and reproductive tiller growth
The ANOVA results showed that seedling emergence time individually had a significant effect on vegetative tiller number, and that precipitation regime and seedling emergence time individually had significant effects on reproductive tiller number. However, their interactions had no effect on vegetative tiller number or reproductive tiller number (Table 1).
| Source | Df | Mean Square | F | Sig. |
|---|---|---|---|---|
| Vegetative tiller number (n plant−1) | ||||
| SET | 2 | 165.964 | 83.377 | .000*** |
| PR | 2 | 0.078 | 0.039 | .962 |
| SET * PR | 4 | 3.402 | 1.709 | .148 |
| Reproductive tiller number (n plant−1) | ||||
| SET | 2 | 16.592 | 11.963 | .000*** |
| PR | 2 | 18.896 | 13.625 | .001*** |
| SET * PR | 4 | 1.741 | 1.255 | .288 |
| Seed number (n spike−1) | ||||
| SET | 2 | 732947.466 | 41.028 | .000*** |
| PR | 2 | 99431.410 | 5.566 | .004** |
| SET * PR | 4 | 22621.282 | 1.266 | .284 |
| Seed size (mg) | ||||
| SET | 2 | 0.313 | 38.160 | .000*** |
| PR | 2 | 0.134 | 16.318 | .000*** |
| SET * PR | 4 | 0.020 | 2.498 | .043* |
| Spike number (n plant−1) | ||||
| SET | 2 | 72.017 | 6.715 | .001*** |
| PR | 2 | 182.704 | 17.036 | .000*** |
| SET * PR | 4 | 13.148 | 1.226 | .300 |
| Seed yield (g m−2) | ||||
| SET | 2 | 61.511 | 36.073 | .000*** |
| PR | 2 | 39.863 | 23.378 | .000*** |
| SET * PR | 4 | 6.003 | 3.52 | .027* |
Growth and reproduction in the C. virgata plants responded differently to different levels of precipitation and seedling emergence times (Fig. 1). The vegetative tiller number was similar under different precipitation regimes at T1 and T2, while significant variations were found among the three precipitation regimes at T3 (Fig. 1A). The vegetative tiller number varied significantly within increasing precipitation and the vegetative tiller number was significantly higher under high precipitation than under low precipitation at T3. There was no significant change in reproductive tiller number with increasing precipitation at T1 (Fig. 1B). At T2 the reproductive tiller number was similar between low and typical precipitation but was significantly higher under high precipitation. At T3 the reproductive tiller number was similar under typical and high precipitation treatments and was significantly higher than under low precipitation. Vegetative tiller number decreased significantly from T1 to T3 under all the precipitation regimes. The highest reproductive tiller numbers occurred at T1 under the low precipitation regime. The reproductive tiller numbers were significantly higher at T1 than at T2 under typical precipitation. Furthermore, changes in seedling emergence time did not affect reproductive tiller number under the high precipitation regime (Fig. 1). The proportion of reproductive tiller number to total tiller number were similar between T1 and T2, while were significantly lower than that at T3 (Fig. 2).
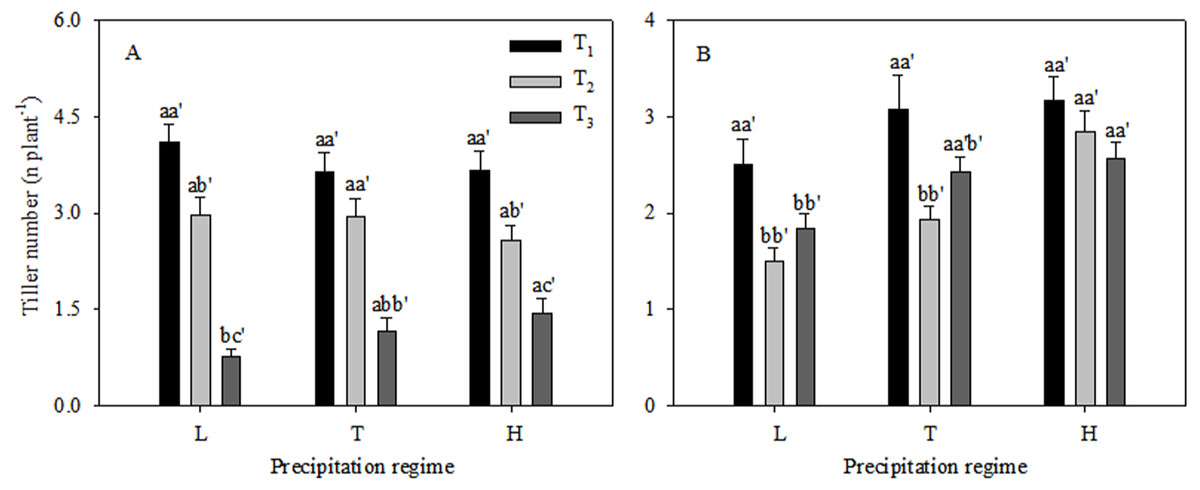
Figure 1: Vegetative tiller number (A) and reproductive tiller number (B) (mean ± 1 s.e.) of C. virgata under different precipitation regimes at different seedling emergence times.
Letters a, b, and c are used for comparisons between different precipitation regimes, while a′, b′ and c′ are used for comparisons between different seedling emergence times. Means with the same letter are not significantly different at P = 0.05.{kind=link}
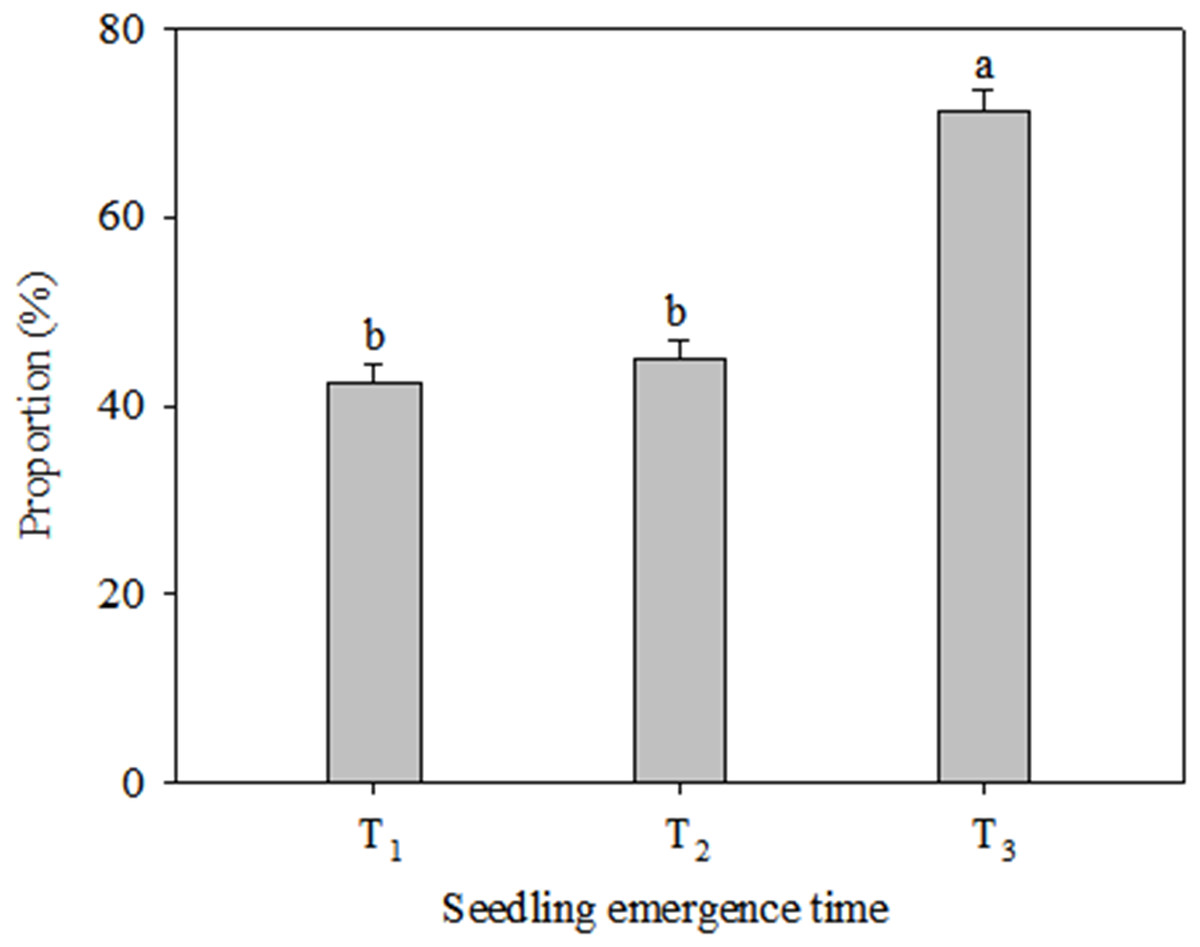
Figure 2: Proportion of reproductive tiller number to total tiller number (mean ± 1 s.e.) of C. virgata at different seedling emergence times.
A, b, and c are used for comparisons between different precipitation regimes. Means with the same letter are not significantly different at P = 0.05.{kind=link}
Seed production
The ANOVA results showed that precipitation regime and seedling emergence time individually had significant effects on seed number, seed size, spike number and seed yield (Table 1). Their interactions had an effect on seed size and seed yield. However, their interactions had no effect on seed number and spike number (Table 1).
The seed number and seed size of the C. virgata plants were significantly different under the various precipitation regimes and seedling emergence times (Fig. 3). Seed number remained unchanged among the three precipitation regimes at T2 and T3, whereas seed number was significantly lower under low precipitation than under the typical and high precipitation regimes at T1. Within the same precipitation treatment seed numbers were similar at T1 and T2 and were significantly higher than at T3 (Fig. 3A). Seed size was unchanged from T1 to T3 under typical and high precipitation and was higher than under low precipitation at all sowing times. Within the same precipitation treatment seed size remained unchanged at T1 and T3 and was higher than at T2 (Fig. 3B).

Figure 3: Seed number (A) and seed size (B) (mean ± 1 s.e.) of C. virgata under different precipitation regimes at different sowing times.
A, b, and c are used for comparisons between different precipitation regimes, while a′, b′ and c′ are used for comparisons between different seedling emergence times. Means with the same letter are not significantly different at P = 0.05.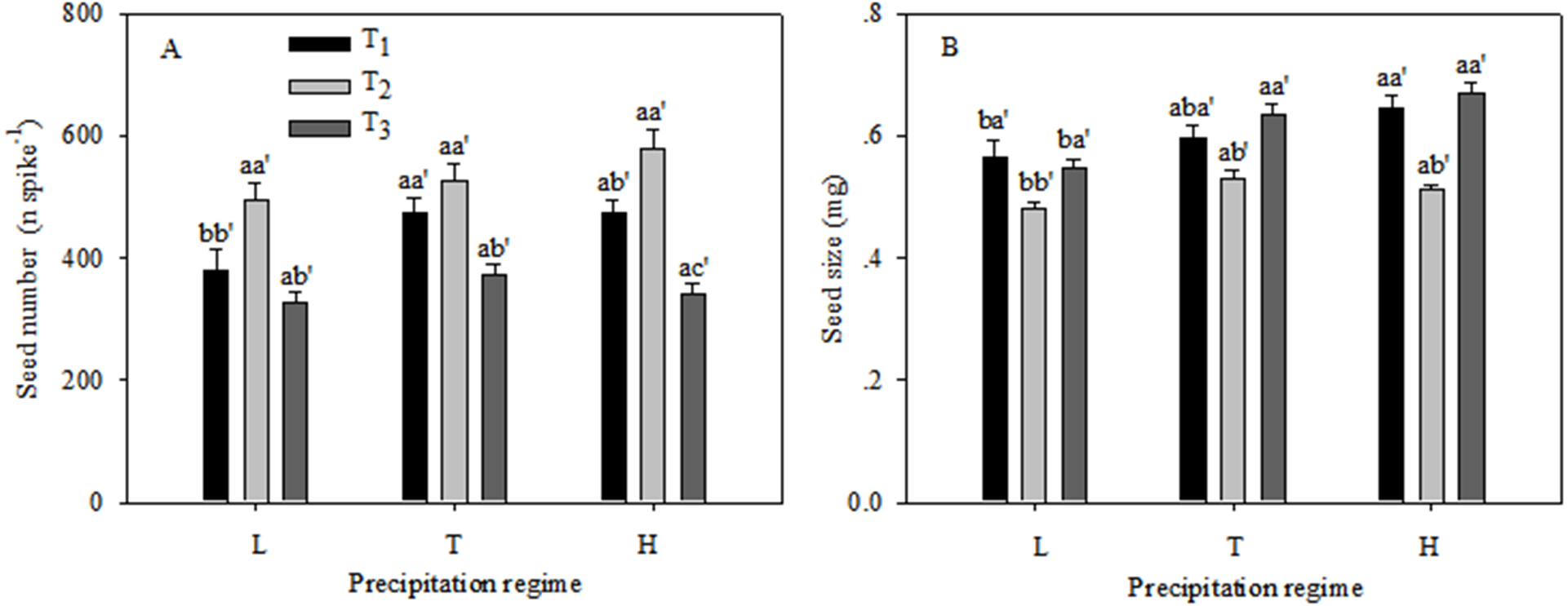{kind=link}
There was a significant difference in spike number among different levels of precipitation at different seedling emergence times (Table 2). Spike number increased from the low to typical precipitation regimes but under high precipitation there was no further significant change at T1. There was a significant difference between low and high precipitation at T2. Spike numbers increased significantly from low to high precipitation at T3. Spike numbers remained unchanged from T1 to T3 under low and high precipitation. While the spike numbers under the typical precipitation regime decreased substantially from T1 to T2. There was no significant difference between T1 and T3, or T2 and T3 under typical precipitation.
| T1 | T2 | T3 | |
|---|---|---|---|
| L | 3.5 ± 0.41bA | 2.5 ± 0.33bA | 2.6 ± 0.32cA |
| T | 6.5 ± 1.07aA | 3.5 ± 0.37abB | 4.6 ± 0.61bAB |
| H | 6.1 ± 0.86aA | 4.6 ± 0.40aA | 6.2 ± 0.55aA |
Notes:
Within a column, means followed by the same lower case letter are not significantly different at P = 0.05.
Within a line, means followed by the same capital letter are not significantly different at P = 0.05.
Seed yield was significantly higher under the typical precipitation regime than under low precipitation at T1 and T3, but there was no difference between typical and high precipitation for these two emergence times (Fig. 4). At T2 there was no significant difference in seed yield between low and typical precipitation but there was a marked increase under high precipitation. The highest yield values occurred at T1 under all precipitation regimes.
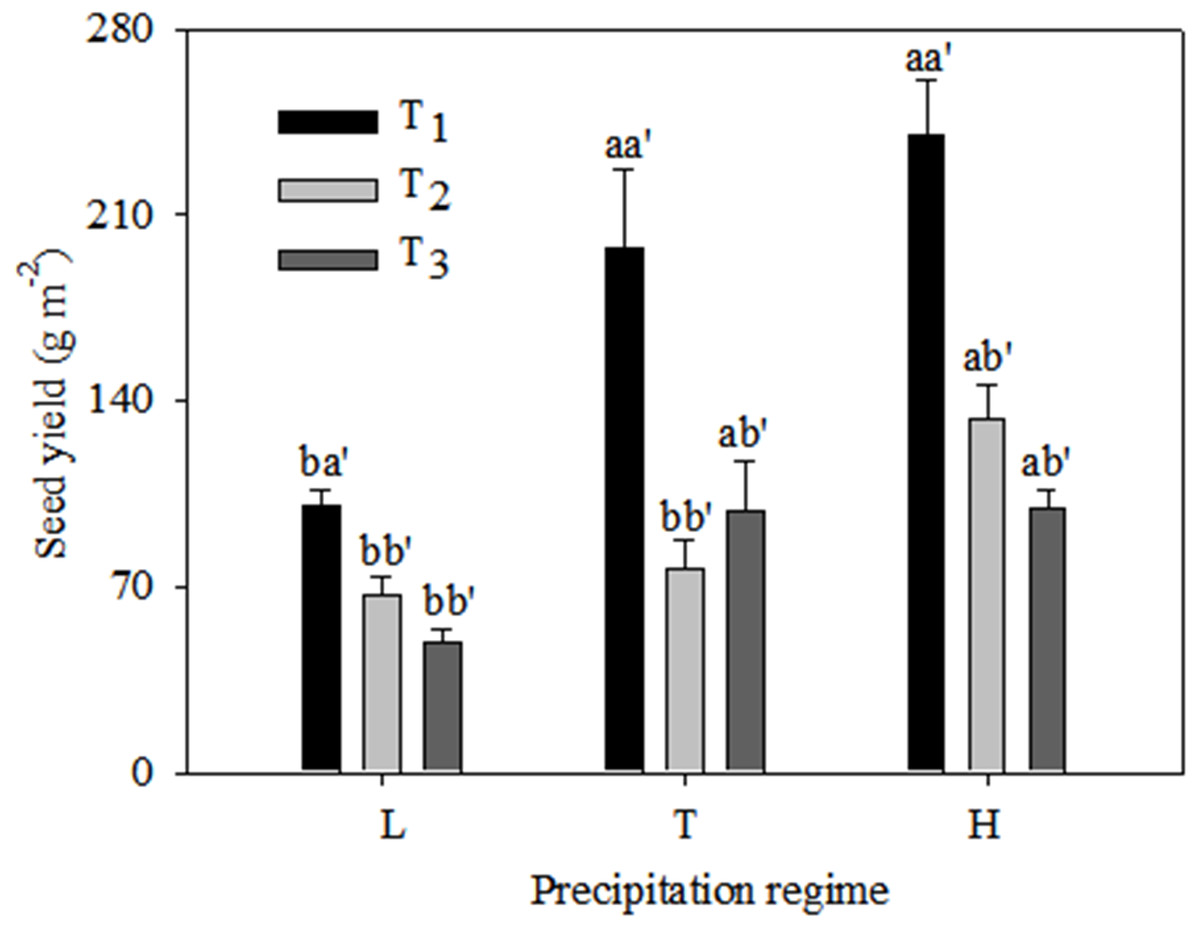
Figure 4: Seed yield (mean ± 1 s.e.) of C. virgata under different precipitation regimes at different seedling emergence times.
A, b, and c are used for comparisons between different precipitation regimes, while a′, b′ and c′ are used for comparisons between different seedling emergence times. Means with the same letter are not significantly different at P = 0.05.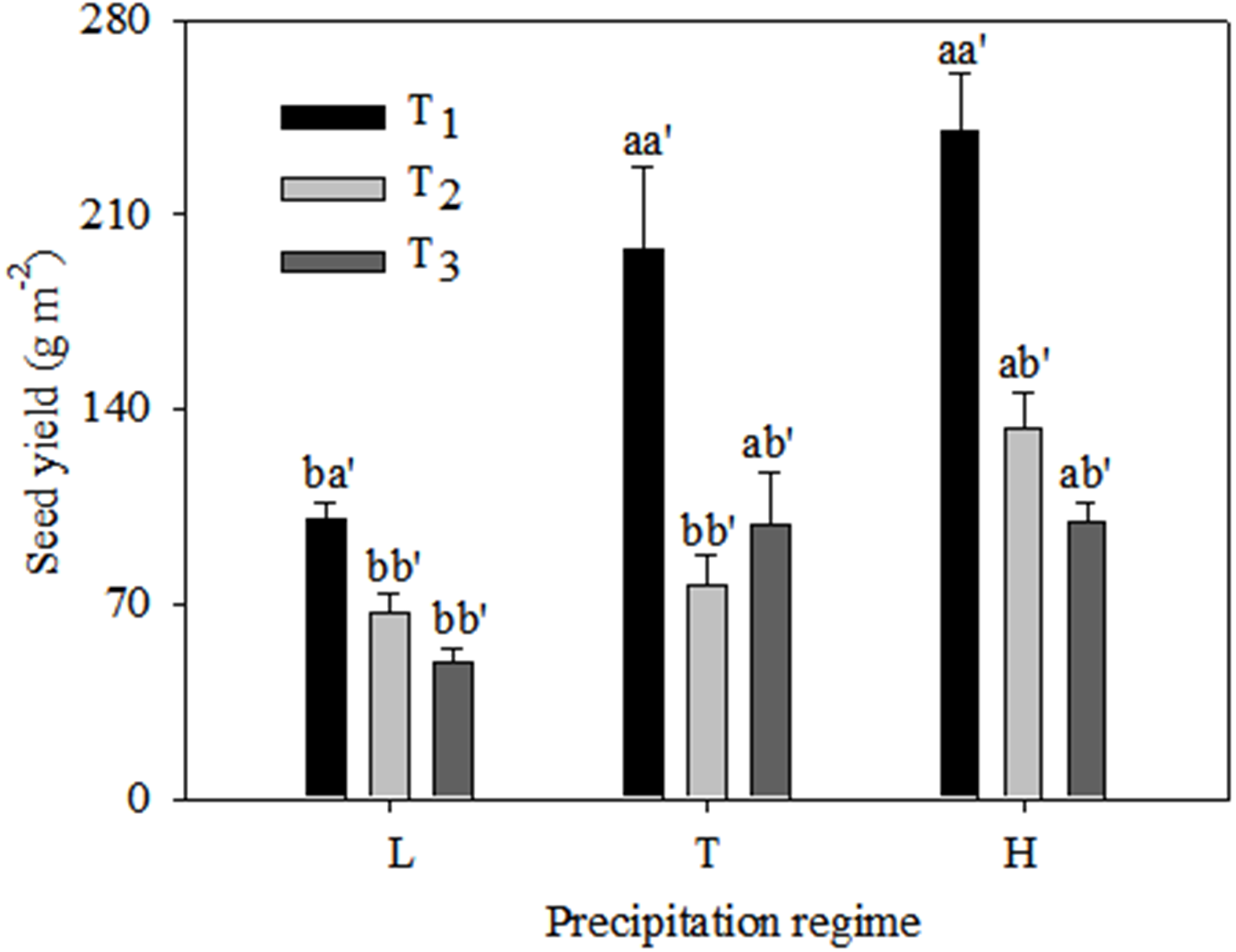{kind=link}
Discussion
Vegetative and reproductive tiller growth
Plants select different reproductive strategies according to their growing environment, and the reproductive strategy selected in a given environment should maximize fitness (Zhang et al., 2018). However, resources are the primary factor determining how plants establish their reproductive machinery (Weiner et al., 2009). Reproductive tiller number is an important characteristic influencing seed yield in many plants (Peng et al., 2015). Whether a bud develops into a reproductive or a vegetative tiller is based on the environment and the evolutionary history of the population (Warringa & Kreuzer, 1996). Our results showed that the trends in tiller number in C. virgata under different precipitation regimes were similar at T1 and T2 but were different at T3. The trends were similar with the results in 2018 when sowed at the early date. It can be concluded that water might be one of the limiting resource for tiller number at T3. Furthermore, the proportional allocation of reproductive tiller number to total tiller number was significantly higher at T3 than at T1 and T2 (Fig. 2). High temperature improved the proportion of reproductive tiller number to total tiller number and the results were consistent with those from previous study (Escalada & Plucknett, 1975). Plant morphology was affected by temperature and daylength, while many plants did not react to critical periods of photoperiod unless their thermal requirements were met. C. virgata produced tiller 10 days later after sowing. The temperature was higher when C. virgata produced tillers at T3 than that at T1 and T2. Delayed sowing and low precipitation reduced C. virgata growth (total tiller number), while maximized it’s reproduction and restoration potential. This strategy has also evolved in monocarpic species and according to optimal allocation theory, such plants should allocate more resources to reproductive parts to improve seed production in unfavorable environments (Ellner, 1987).
Seed production
Seed production are related to plant adaptations to their growing environment. For example, the maternal nutrient environment affects seed size, seed yield and the competitive ability of progeny (Gómez, 2004). In our study, seed size was smaller at T2 compared to the other two emergence times. The larger seed size at T1 and T3 may indicate higher seed quality because large seed size has been associated with higher seed germination ability and seedling survival as reported by other authors (Cowell & Doyle, 1993) and in our earlier work (Wang et al., 2018). This also suggests a management method to produce high quality seeds from C. virgata.
Moreover, our results showed that when C. virgata was delayed in emergence beyond 30 d or 60 d it produced fewer seeds. It has been demonstrated that sowing date was one of the critical factors for productivity of winter wheat (Triticum aestivum L.) due to different amounts of soil water storage at different sowing times (Ren et al., 2019). Further, Awan & Chauhan (2016) have reported that a delay in emergence would result in fewer seeds returning to the seed bank, which has important implications for vegetation succession and restoration (Shang et al., 2016). However, seed yields were similar among the three water supply treatments and the first-order spike number was one of the most important factors in determining seed yield in our former work (Wang et al., 2018). Seed yields in the current work were higher in typical and high precipitation than under low precipitation. The pattern of seed yield was not consistent with our former work in 2018, and the reason for this might be the different plant densities in the two experiments. However, the first-order spike number (reproductive tiller number) was higher under low precipitation than under typical and high precipitation in these earlier experiments under a density of five plants per pot, whereas the first-order spike number was significantly higher under typical and high precipitation compared to low precipitation with a density of ten plants per pot in the current work (Fig. 1). This indicated the existence of different reproductive strategies under different plant densities. Here, reproductive tiller number was demonstrated to be an important component for C. virgata seed yield, and future work should explore the effect of plant densities on growth and reproductive characteristics.
Conclusion
In conclusion, simulated precipitation regimes and seedling emergence time affect the reproductive strategy of C. virgata. Increased precipitation did increase seed yield in this study, which suggested that reproduction in C. virgata benefits from average and above-average precipitation regimes. In addition, dry years or delayed precipitation during seedling emergence due to climate change might have a negative effect on plant recovery of C. virgata in arid and semiarid areas such as the Songnen grassland. Early sowing in spring might be a strategy to ensure high seed yield and high seed quality by avoiding late-season drought conditions for this species. These results will also provide some important information on management measures for the establishment and recovery of C. virgata in degraded grassland.