
Morphological and DNA sequence data uncover a new millipede species in the Thyropygus opinatus subgroup and assign T. peninsularis to this subgroup (Diplopoda: Spirostreptida: Harpagophoridae)
- Published
- Accepted
- Received
- Academic Editor
- Viktor Brygadyrenko
- Subject Areas
- Biodiversity, Taxonomy, Zoology
- Keywords
- DNA barcoding, Mitochondrial DNA, Phylogeny, Southeast Asia
- Copyright
- © 2025 Pimvichai et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2025. Morphological and DNA sequence data uncover a new millipede species in the Thyropygus opinatus subgroup and assign T. peninsularis to this subgroup (Diplopoda: Spirostreptida: Harpagophoridae) PeerJ 13:e19277 https://doi.org/10.7717/peerj.19277
Abstract
The millipede genus Thyropygus Pocock, 1894 is one of the most diverse genera within the family Harpagophoridae in Southeast Asia. The Thyropygus opinatus subgroup, belonging to the T. allevatus group, is distinguished by the presence of an additional projection on the anterior coxal fold. Here, we describe a new species of the T. opinatus subgroup, Thyropygus payamense sp. nov., from Payam Island, Ranong Province, Thailand, based on morphological and DNA sequence data. The mean interspecific COI divergence between the new species and other Thyropygus species is 0.13 ± 0.02 (range: 0.07–0.16). The new species is distinguished by (1) a small, slender, pointed spine at base of femoral spine, (2) a short, triangular mesal process of the anterior coxal fold, and (3) a short, slender, slightly mesad-curving tibial spine. Additionally, T. peninsularis Hoffman, 1982 is confirmed as a member of the T. opinatus subgroup, because it shares key gonopodal characters with other species in this subgroup, while COI and 16S rRNA sequence data firmly support this new classification, with a mean interspecific COI sequence divergence of 0.13 ± 0.03 (range: 0.07–0.17) from other species in the T. allevatus group. An identification key for all 29 species in the T. opinatus subgroup is provided. Further research is needed to assess the taxonomic status of, and phylogenetic relationships within, this subgroup, which, except for two species, may tentatively represent an endemic species radiation in the peninsular area of Thailand, Malaysia and Myanmar.
Introduction
The millipede genus Thyropygus Pocock, 1894 is widely distributed across Thailand and Southeast Asia and currently comprises 67 recognized species, 46 of which are exclusively found in Thailand (Pimvichai, Enghoff & Backeljau, 2023). Most Thai species belong to the informal T. allevatus group, which was defined by Hoffman (1975) on the basis of two features of the gonopod telopodite: (1) the presence of tibial and femoral spines, and (2) the tibial spine being very long and recurved proximad towards the femoral spine. The T. allevatus group is widely distributed throughout Thailand, Vietnam, Laos, Cambodia, and Peninsular Malaysia (Enghoff, 2005). By combining morphological and DNA sequence data, the T. allevatus group has been further divided into four informal subgroups: (1) the T. opinatus subgroup, (2) the T. induratus subgroup, (3) the T. cuisinieri subgroup, and (4) the T. allevatus subgroup (Pimvichai, Enghoff & Panha, 2009a; Pimvichai, Enghoff & Panha, 2009b; Pimvichai, Enghoff & Panha, 2011a; Pimvichai, Enghoff & Panha, 2011b; Pimvichai, Enghoff & Panha, 2014; Pimvichai et al., 2016; Pimvichai, Enghoff & Backeljau, 2023). Within this system, the T. opinatus subgroup is characterized by the presence of an additional projection on the anterior coxal fold (Pimvichai et al., 2016). The T. opinatus subgroup is primarily distributed in Thailand, with only two species that also occur outside Thailand: T. implicatus (Demange, 1961) in Peninsular Malaysia and T. opinatus (Karsch, 1881) in southern Myanmar (Pimvichai, Enghoff & Panha, 2009a; Pimvichai, Enghoff & Panha, 2009b; Pimvichai, Enghoff & Panha, 2014).
Hitherto, the informal subgroup division of the T. allevatus group appeared well-supported by the overall congruence between morphological and DNA sequence data. Yet, recently this congruence was challenged with the discovery of two Thyropygus species, T. panhai Pimvichai, Enghoff & Backeljau, 2023 and T. somsaki Pimvichai, Enghoff & Backeljau, 2023, that morphologically clearly belong to the T. induratus subgroup, but whose COI sequences do not support this assignment. In fact, including both species in the COI phylogeny made that the monophyly of the T. induratus subgroup was no longer supported (Pimvichai, Enghoff & Backeljau, 2023). Hence extended taxon sampling is important to further explore the congruence between morphological and DNA sequence data, and eventual taxonomic validity, of the informal subgroups within the T. allevatus group.
Against this background, recently collected millipede specimens from Payam Island in the Andaman Sea appeared morphologically to belong to a new species of the T. opinatus subgroup, thus offering an opportunity to test the consistency of this subgroup. The present contribution aims to do so by formally describing and DNA barcoding this new species as Thyropygus payamense sp. nov. In addition, it provides an updated morphological identification key of all species currently assigned to the T. opinatus subgroup and discusses the taxonomic position of T. peninsularis Hoffman, 1982, a species which until recently was assigned to the T. erythropleurus group (Hoffman, 1982; Pimvichai, Enghoff & Panha, 2009a), but whose transfer to the T. opinatus subgroup in the T. allevatus group (Pimvichai, Enghoff & Backeljau, 2023) is here formally confirmed.
Materials & Methods
Specimen collection
In November 2022 live specimens of the new species were hand-collected at Payam Island, Ranong Province, Thailand and preserved in 70% ethanol (n = 3) or stored in a freezer at −20 °C (n = 10). This material has been deposited in the collections of the Museum of Zoology, Chulalongkorn University, Bangkok, Thailand (CUMZ). Another specimen of T. payamense sp. nov. from Payam Island, collected in April 2013 by J. Urbanski and preserved in 70% ethanol, is kept in the Natural History Museum of Denmark (NHMD).
This research was conducted under the approval of the Animal Care and Use regulations (numbers U1-07304-2560 and IACUC-MSU-037/2019) of the National Research Council of Thailand.
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:68E7FD7F-A8E3-4BE9-9B4B-136CDEEBEA88. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central SCIE and CLOCKSS.
Morphology
Gonopods were photographed with a digital camera and drawings were made using a stereomicroscope and photographs. Gonopod terminology of the T. opinatus subgroup follows Pimvichai, Enghoff & Panha (2009a); Pimvichai, Enghoff & Panha (2009b); Pimvichai et al. (2016). A new term is marked in bold:
ac = anterior coxal fold: the main part of gonopod in anterior view; confusingly called posterior coxal fold by Demange (1961) and Hoffman (1975)
aip = additional spine-like process: between lateral and mesal processes of anterior coxal fold
alp = lateral process of anterior coxal fold: the distolateral part of the anterior coxal fold
amp = mesal process of anterior coxal fold: an additional projection on the anterior coxal fold, protruding from its mesal margin
bp = blepharochaete (pl. -ae): the normal form of apical setae, long, slender, stiffened, and usually pigmented, somewhat reminiscent of the mammalian eyelash (Hoffman, 1975)
cr = longitudinal crest in gutter of palette: a crest which runs along the middle of the gutter near the tip of the palette
fe = femoral spine (also fe 1 and fe 2): a usually long, curved spine on the telopodite, originating slightly distal to the point where the telopodite emerges from the coxa
lc = longitudinal crest: a strong longitudinal crest at the mesal margin of amp in posterior view
ll = lamellar lobe: a small, slightly folded lobe at the basis of the apical part of the telopodite
lo = telopodite lobe: a protruding lobe on the telopodite, distal to fe
pa = palette: the distalmost lobe of the apical part, carrying the row of blepharochaetae
pc = posterior coxal fold: the main part of gonopod in posterior view, usually shorter than ac and forming a shelf for accommodation of telopodite shaft
plp = lateral process of posterior coxal fold: the lateral part of the posterior coxal fold, usually digitiform
pmp = mesal process of posterior coxal fold: the mesal part of the posterior coxal fold, usually forming a shelf for accommodation of telopodite shaft
px = paracoxite: the basal, lateral part of the posterior coxal fold
sfe = small spine at the base of femoral spine: an additional small, slender, sharp spine at the base of femoral spine
sl = spatulate lobe: a distinct distal, separate lobe at the apical part, spatulate, sometimes with a distal spine-like process
sls = slender long spine: an additional slender long spine (much longer than ss) at the base of the apical part of telopodite in posterior view
ss = small spine: an additional small spine at the base of the apical part of telopodite in posterior view
st = sternum: a small, usually triangular sclerite between the basal parts of the anterior coxal folds
ti = tibial spine: a usually long spine on the telopodite, originating distal to the femoral spine, at the basis of the apical part of the telopodite, usually curved in the opposite direction of the femoral spine, the two together forming a circle
Apical part: the part of the telopodite distal to the tibial spine
Shelf: the distal surface of the posterior coxal fold.
DNA extraction, amplification, and sequencing
Total genomic DNA was extracted from legs of three specimens using the NucleoSpin Tissue kit (Macherey-Nagel, Düren, Germany) following the manufacturer’s instructions. PCR amplifications and sequencing of the standard mitochondrial COI DNA barcoding fragment (Hebert et al., 2003) and a mitochondrial 16S rRNA fragment were done as described by Pimvichai et al. (2020). The COI fragment was amplified with the primers LCO-1490 and HCO-2198 (Folmer et al., 1994), and the 16S rRNA fragment was amplified with the primers 16Sar and 16Sbr (Kessing et al., 2004). The new COI and 16S rRNA sequences have been deposited in GenBank under accession numbers PV019345 –PV019347 and PV029246 –PV029247. Sample data and voucher codes are provided in Table S1.
DNA sequence analysis
The COI dataset comprised 61 specimens of 33 nominal Thyropygus species and four outgroup species from the harpagophorid subfamily Rhynchoproctinae viz., Anurostreptus barthelemyae Demange, 1961, A. sculptus Demange, 1961, Armatostreptus armatus (Demange, 1983), and Heptischius lactuca Pimvichai, Enghoff & Panha, 2010 (Table S1). The same specimens were used for the 16S rRNA and combined COI + 16S rRNA datasets, except for T. payamense sp. nov. (KPYR3), T. panhi and T. somsaki, of which no 16S rRNA sequences could be obtained.
Sequence assembly and editing were performed using CodonCode Aligner (ver. 4.0.4; CodonCode Corporation) to combine forward and reverse reads, identify errors, and resolve ambiguities. All sequences were verified using the Basic Local Alignment Search Tool (BLAST, NCBI) and compared against reference sequences in GenBank. Sequence alignment was conducted using MUSCLE (ver. 3.6; Edgar, 2004; http://www.drive5.com/muscle). The sequences were evaluated for ambiguous nucleotide sites, saturation, and phylogenetic signal using DAMBE (ver. 5.2.65; Xia, 2018; https://dambe.bio.uottawa.ca/DAMBE/dambe.aspx). MEGA11 (ver. 11.0.10; Tamura, Stecher & Kumar, 2021; http://www.megasoftware.net) was used to: (1) screen for stop codons, (2) translate nucleotide sequences into amino acids, and (3) calculate uncorrected pairwise p-distances among sequences.
Phylogenetic analysis
Phylogenetic trees were constructed using maximum likelihood (ML) and Bayesian Inference (BI) approaches.
ML trees were inferred using RAxML (ver. 8.2.12; Stamatakis, 2014; http://www.phylo.org/index.php/tools/raxmlhpc2_tgb.html) via the CIPRES Science Gateway (Miller, Pfeiffer & Schwartz, 2010) and applying the GTR+G substitution model.
BI trees were constructed using MrBayes (ver. 3.2.7a; Huelsenbeck & Ronquist, 2001; http://www.phylo.org/index.php/tools/mrbayes_xsede.html). Substitution models were selected using jModeltest (ver. 2.1.10; Darriba et al., 2012; https://www.github.com/ddarriba/jmodeltest2/releases), with the Akaike Information Criterion (Akaike, 1973) as the selection criterion. The GTR+I+G model was identified as the best fit model for COI (lnL = 11,936.7043, gamma shape = 0.8820), 16S rRNA (–lnL = 8,382.4103, gamma shape = 0.8950), and the combined COI + 16S rRNA dataset (–lnL = 3,392.4942, gamma shape = 0.4530). BI analyses were run for 10 million (combined dataset), 20 million (COI), and 2 million (16S rRNA) generations. The heating parameter was set to 0.01 for all datasets, and trees were sampled every 1,000 generations. Convergence was confirmed by ensuring that the standard deviation of split frequencies was <0.01. The first 1,000 trees were discarded as burn-in, and the final consensus tree was generated from the last 15,002 (combined dataset), 30,002 (COI), and 3,002 (16S rRNA) trees.
Node support was evaluated using posterior probabilities (PP) for BI and bootstrap values (BV) for ML (based on 1,000 replicates). Nodes with BV ≥ 70% or PP ≥ 0.95 were considered well-supported, while BV <70% or PP <0.95 were considered as poorly supported (Hillis & Bull, 1993; San Mauro & Agorreta, 2010).
Results
DNA sequence data and phylogeny
The uncorrected p-distances between the COI sequences (660 bp) of Thyropygus specimens included in this study ranged from 0.00 to 0.18 (Table S2). The mean intraspecific sequence divergence within the T. allevatus group was 0.06 ± 0.03 (range: 0.00–0.12). Mean intraspecific divergence values for individual species of this group were: T. allevatus (two specimens) = 0.00; T. induratus = 0.05 ± 0.02 (range: 0.02–0.07); T. payamense sp. nov. (three specimens) = 0.01 ± 0.02 (range: 0.00–0.01); T. resimus = 0.06 ± 0.04 (range: 0.00–0.10); and T. uncinatus = 0.06 ± 0.03 (range: 0.00–0.12). The mean interspecific sequence divergence within the T. allevatus group (all subgroups included) was 0.14 ± 0.02 (range: 0.02–0.18). The mean interspecific sequence divergence within the T. opinatus subgroup was 0.12 ± 0.03 (range: 0.02–0.17). The mean interspecific sequence divergence in the T. opinatus subgroup without T. payamense sp. nov. = 0.12 ± 0.03 (range: 0.02–0.17). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. opinatus subgroup = 0.11 ± 0.02 (range: 0.07–0.15). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. allevatus group = 0.13 ± 0.02 (range: 0.07–0.16).
The uncorrected p-distances between the 16S rRNA sequences (487 bp) of Thyropygus species ranged from 0.00 to 0.13 (Table S3). The mean intraspecific sequence divergence within the T. allevatus group was 0.02 ± 0.02 (range: 0.00–0.08). Mean intraspecific divergence values for individual species of this group were: T. allevatus (two specimens) = 0.00; T. induratus = 0.03 ± 0.03 (range: 0.01–0.08); T. payamense sp. nov. (two specimens) = 0.00; T. resimus = 0.02 ± 0.01 (range: 0.00–0.03); and T. uncinatus = 0.02 ± 0.01 (range: 0.00–0.04). The mean interspecific sequence divergence within the T. allevatus group (all subgroups included) was 0.08 ± 0.02 (range: 0.00–0.13). The mean interspecific sequence divergence within the T. opinatus subgroup was: 0.05 ± 0.02 (range: 0.00–0.9). The mean interspecific sequence divergence in the T. opinatus subgroup without T. payamense sp. nov. = 0.05 ± 0.02 (range: 0.00–0.09). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. opinatus subgroup = 0.05 ± 0.02 (range: 0.01–0.08). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. allevatus group = 0.08 ± 0.03 (range: 0.01–0.12).
The uncorrected p-distances between the sequences of Thyropygus species in the combined dataset (COI + 16S rRNA, 1,147 bp) ranged from 0.01 to 0.15 (Table S4). The mean intraspecific sequence divergence within the T. allevatus group was 0.04 ± 0.02 (range: 0.00–0.08). Mean intraspecific divergence values for individual species of this group were: T. allevatus (two specimens) = 0.00; T. induratus = 0.04 ± 0.02 (range: 0.02–0.07); T. payamense sp. nov. (two specimens) = 0.00; T. resimus = 0.04 ± 0.03 (range: 0.00–0.07); and T. uncinatus = 0.05 ± 0.02 (range: 0.00–0.08). The mean interspecific sequence divergence within the T. allevatus group (all subgroups included) was 0.11 ± 0.02 (range: 0.01–0.15). The mean interspecific sequence divergence within the T. opinatus subgroup was: 0.09 ± 0.02 (range: 0.01–0.13). The mean interspecific sequence divergence in the T. opinatus subgroup without T. payamense sp. nov. = 0.09 ± 0.03 (range: 0.01–0.13). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. opinatus subgroup = 0.08 ± 0.02 (range: 0.05–0.12). The mean interspecific sequence divergence of T. payamense sp. nov. vs other species in the T. allevatus group = 0.11 ± 0.03 (range: 0.05–0.14).
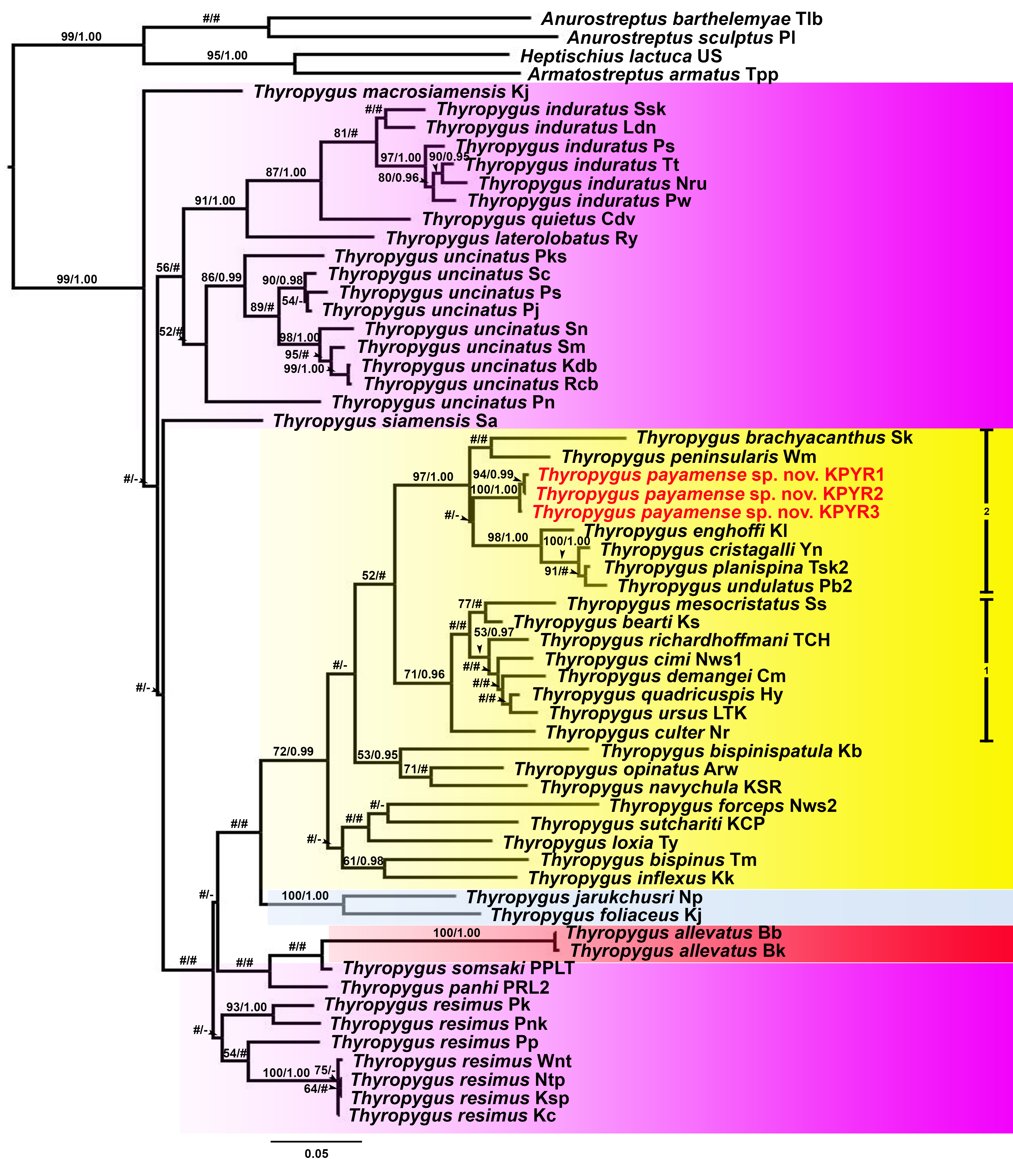
The ML and BI trees (COI and 16S rRNA separately, as well as COI + 16S rRNA combined) were largely congruent with respect to the well-supported nodes (by visual inspection). So, for further discussion, the combined COI + 16S rRNA tree will be used (Fig. 1), while the separate COI and 16S rRNA trees are provided in Figs. S1 and S2.
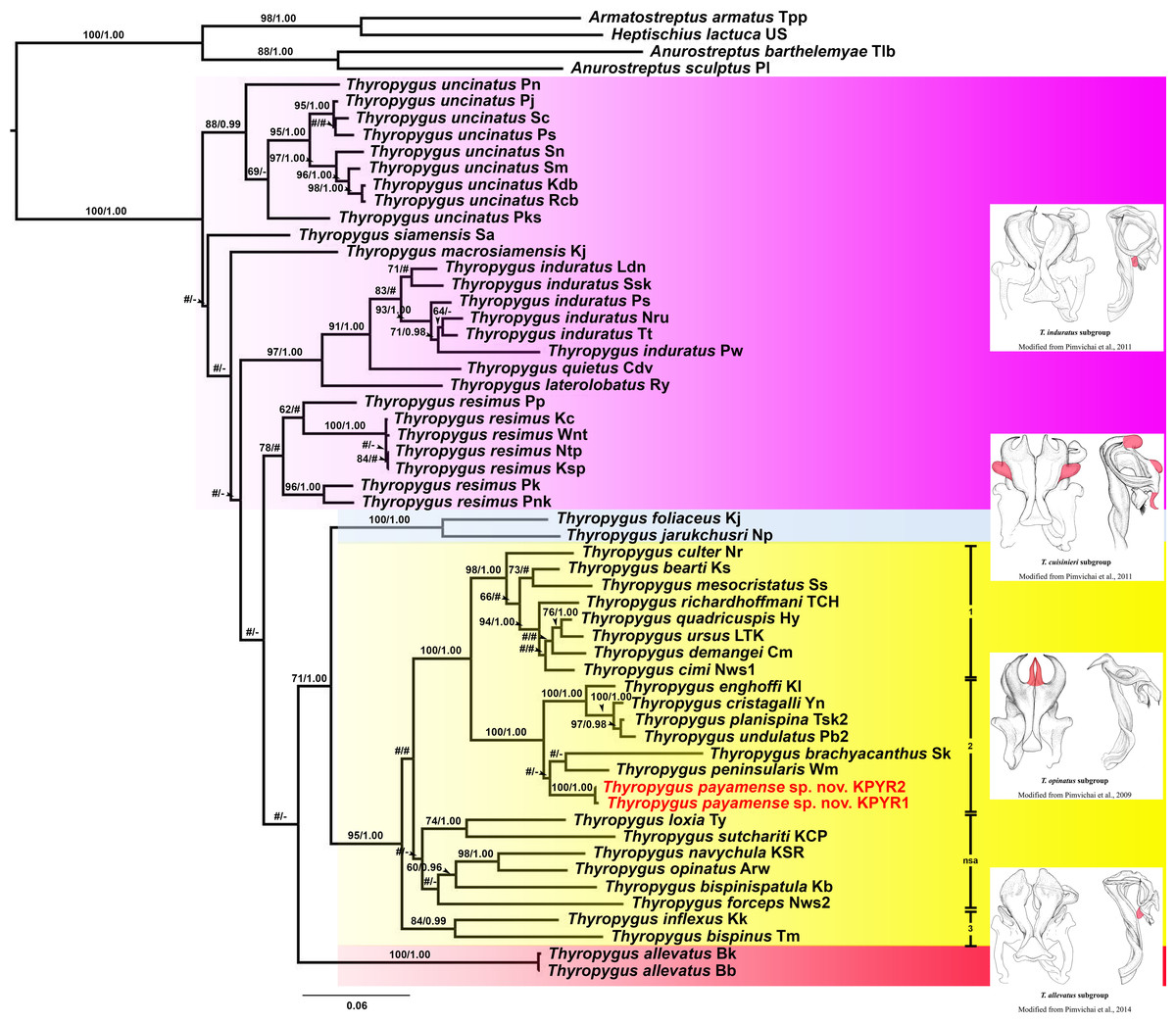
Figure 1: Phylogenetic relationships of Thyropygus species based on maximum likelihood analysis (ML) and Bayesian Inference (BI) of 1,147 bp in the combined COI + 16S rRNA alignment.
Numbers at nodes indicate node support based on bootstrapping (ML)/posterior probabilities (BI). Scale bar = 0.06 substitutions/site. # indicates nodes with < 50% bootstrap support and < 0.95 posterior probability. - indicates non-supported nodes. The colored areas mark the T. induratus subgroup (purple), T. cuisinieri subgroup (blue), T. opinatus subgroup (yellow), and T. allevatus subgroup (red). The vertical lines indicate different clades within the T. opinatus subgroup. Abbreviations after species names refer to the locality names in Table S1. Unique gonopodal characters shared by members of each subgroup are highlighted in red. These characters represent key morphological features that define and differentiate the subgroups.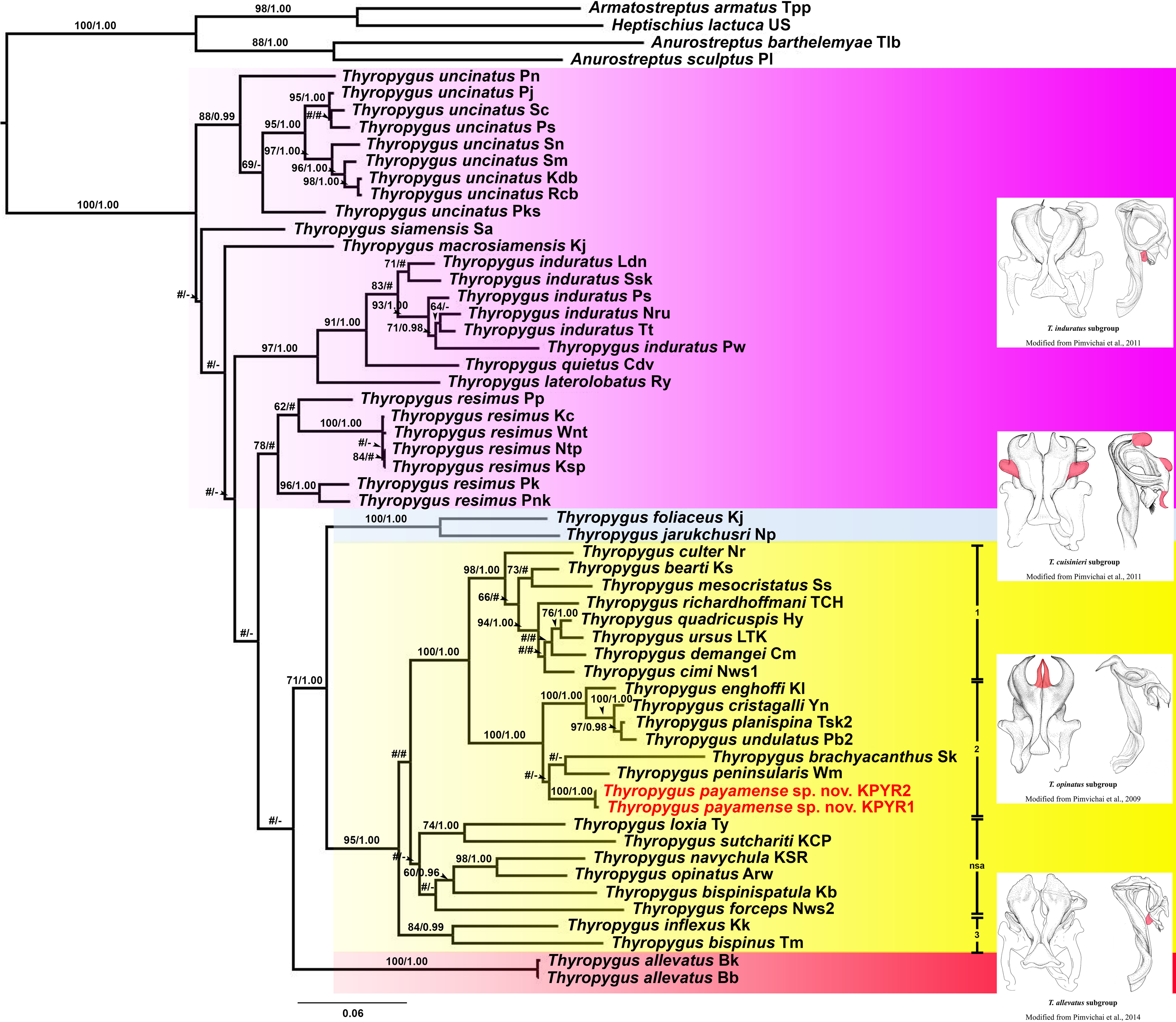{kind=link}
Thyropygus payamense sp. nov. was firmly positioned within the T. opinatus subgroup (Fig. 1), whose monophyly was strongly supported (BV = 95; PP = 1.00). The T. opinatus subgroup was further divided into a non-supported assemblage (nsa) of six species, viz., T. bispinispatula, T. forceps, T. loxia, T. navychula, T. opinatus, and T. sutchariti (Fig. 1: nsa) and three well-supported clades (Fig. 1: 1–3):
Clade 1: was almost maximally supported (BV = 98, PP = 1.00) and comprised eight species from southern Thailand: T. bearti, T. cimi, T. culter, T. demangei, T. mesocristatus, T. quadricuspis, T. richardhoffmani, and T. ursus. This clade was maximally supported as sister group of clade 2.
Clade 2 : was maximally supported (BV = 100, PP = 1.00) and comprised seven species from southern Thailand: T. brachyacanthus, T. cristagalli, T. enghoffi, T. payamense sp. nov., T. peninsularis, T. planispina, and T. undulatus.
Clade 3: was well-supported (BV = 84, PP = 0.99) and comprised two singleton species from northern, central and western Thailand: T. inflexus and T. bispinus. The sister group position of this clade was not well-resolved.
Additionally, the T. cuisinieri subgroup was well-supported (BV = 99, PP = 1.00), consisting of two singleton species: T. foliaceus and T. jarukchusri, that jointly were well-supported as sister taxon of the T. opinatus subgroup. The T. allevatus subgroup was only represented by its nominal species, whose sister group position was not resolved. There was no support for the monophyly of the T. induratus subgroup (Fig. 1: assemblage marked in purple).
In the separate COI tree (Fig. S1), clades 1 and 2 were each well-supported, but their sister group relation was not, while clade 3 was only well-supported in the BI analysis, but its sister group relationship was unresolved. In contrast, clade 1 was not supported in the separate 16S rRNA tree (Fig. S2), while clades 2 and 3 were only well-supported in the ML analysis. Nevertheless, the species of clades 1 and 2 were grouped together in a well-supported overarching clade, while the sister group relationship of clade 3 was unresolved. The six species from the non-supported assemblage in the combined tree, remained as such in either of the separate trees since they appeared scattered throughout the T. opinatus subgroup. The T. cuisinieri subgroup was consistently well-supported by the separate COI and 16S rRNA trees, but its sister group relationships were not. Also the sister group position of T. allevatus remained unresolved, while there was no support for the monophyly of the T. induratus subgroup.
Taxonomy
| Class Diplopoda de Blainville in Gervais, 1844 |
| Order Spirostreptida Brandt, 1833 |
| Suborder Spirostreptidea Brandt, 1833 |
| Family Harpagophoridae Attems, 1909 |
| Genus Thyropygus Pocock, 1894 |
| Informal taxon Thyropygus allevatus group sensu Hoffman (1975) |
| Informal taxon Thyropygus opinatus subgroup sensu Pimvichai et al. (2016) |
Diagnosis. A subgroup of the T. allevatus group. Differing from the T. induratus, T. cuisinieri and T. allevatus subgroups by having an additional projection on the anterior coxal fold (amp).
Included species:
T. bearti Pimvichai, Enghoff & Panha, 2009
T. bifurcus (Demange, 1986)
T. bispinispatula Pimvichai, Enghoff & Panha, 2009
T. bispinus Pimvichai, Enghoff & Panha, 2009
T. brachyacanthus Pimvichai, Enghoff & Panha, 2009
T. casjeekeli Pimvichai, Enghoff & Panha, 2009
T. chelatus Pimvichai, Enghoff & Panha, 2009
T. cimi Pimvichai, Enghoff, Panha & Backeljau, 2016
T. cristagalli Pimvichai, Enghoff & Panha, 2009
T. culter Pimvichai, Enghoff, Panha & Backeljau, 2016
T. demangei Pimvichai, Enghoff & Panha, 2009
T. enghoffi (Demange, 1989)
T. erectus Pimvichai, Enghoff & Panha, 2009
T. floweri Demange, 1961
T. forceps Pimvichai, Enghoff, Panha & Backeljau, 2016
T. implicatus Demange, 1961
T. inflexus (Demange, 1989)
T. loxia Pimvichai, Enghoff & Panha, 2009
T. mesocristatus Pimvichai, Enghoff, Panha & Backeljau, 2016
T. navychula Pimvichai, Enghoff, Panha & Backeljau, 2016
T. opinatus (Karsch, 1881)
T. payamense sp. nov.
T. peninsularis Hoffman, 1982 (see Discussion)
T. planispina Pimvichai, Enghoff, Panha & Backeljau, 2016
T. quadricuspis Pimvichai, Enghoff & Panha, 2009
T. richardhoffmani Pimvichai, Enghoff & Panha, 2009
T. sutchariti Pimvichai, Enghoff, Panha & Backeljau, 2016
T. undulatus Pimvichai, Enghoff, Panha & Backeljau, 2016
T. ursus Pimvichai, Enghoff, Panha & Backeljau, 2016
Species description
Thyropygus payamense sp. nov. (Figs. 2–4)
Material examined. Holotype male (CUMZ-D00155), THAILAND, Ranong Province, Muang Ranong District, Payam Island, Aow Yai, 10 m a.s.l., 9°43′45″N, 98°23′25″E, 13/11/2022, leg. P. Pimvichai, T. Backeljau, B. Segers, K. Breugelmans and S. Saratan. Paratypes five males (CUMZ-D00155-1), eight females (CUMZ-D00155-2), same data as holotype, one male (NHMD 1184744) Thailand, Ranong Province, Muang Ranong District, Payam Island, /04/2013, leg. J. Urbanski.
Etymology. The name refers to Payam Island, the type locality of this species.
Diagnosis. A species of the T. opinatus subgroup in the T. allevatus group. Differs from all other species of the T. opinatus subgroup by having (1) a small, slender, pointed spine (sfe) at base of femoral spine (fe), (2) the mesal process of anterior coxal fold (amp) short, forming a triangular process, and (3) tibial spine (ti) short, slender, slightly curving mesad.
Description. Adult males with 60–61 podous rings, no apodous rings. Length 13–14 cm, width 8.6–9.3 mm. Adult females with 60–62 podous rings, no apodous rings. Length 12–14 cm, width 8.7–9.4 mm.
Colour. Overall colour of living animal (Fig. 2) dark brown. Antennae, legs, epiproct, paraprocts and hypoproct reddish brown.
Gonopods (Figs. 3A–3D). Anterior coxal fold (ac; Fig. 3A): the lateral process (alp) flattened and broad, apically curved caudad and terminating in a short spine, the lateral margin slightly folded; the mesal process (amp) broad at base, apically gradually narrowed, pointed, forming a triangular process, of the height of the lateral process (alp). Posterior coxal fold (pc; Fig. 3B) basally with moderately high paracoxites (px), forming shelf to accommodate telopodite, distally with two processes: mesal process (pmp) very small, directed distolaterad; lateral process (plp) digitiform, directed distad. Telopodite (Figs. 3C–3D) leaving coxite over shelf of posterior coxal fold; the femoral spine (fe) very long, slender, curving backward, with a small, slender, pointed spine (sfe) at its base, in situ resting behind alp; the tibial spine (ti) short, slender, slightly curving mesad; the apical part: spatulate lobe (sl) small, rounded; palette (pa) simple, gutter-like; distally with about 11 brownish blepharochaetae (bp).

Figure 2: Live Thyropygus payamense sp. nov., paratype, male (CUMZ - D00155-1).
{kind=link}
DNA barcodes. The GenBank accession number of the COI barcode of the holotype is PV019345 and that of 16S rRNA is PV029246 (voucher code CUMZ-D00155). The COI barcodes of paratypes are PV019346 –PV019347 (voucher code CUMZ-D00155-1 for a male and voucher codes CUMZ-D00155-2, CUMZ-D00155-2-1 for 2 females). The 16S rRNA barcode of the paratype with voucher code CUMZ-D00155-2 is PV029247.
Distribution. The species is known only from its type locality in Ranong Province, Thailand (Fig. 4). It was collected in Aow Yai, where the specimens were found crawling and hiding underneath leaf litter of coconut trees, jackfruit trees, and other native vegetation.
Key to the 29 currently recognized species of the T. opinatus subgroup; figures underneath a couplet illustrate the relevant gonopodal characteristics referred to in the couplet (updated from Pimvichai et al., 2016) (Supplementary Files)
1. Apical part of telopodite with spatulate lobe (sl)……………………............... …………....2
− Apical part of telopodite with lamellar lobe (ll)……………….………...............…………22
2. Spatulate lobe (sl) distally drawn out into one or two sharp dark brown spine(s)…….…………….………………….…………….…………….…....................3
− Spatulate lobe (sl) distally expanded and/or rounded, spoon-like, without a spine ...…............................……….……………….…………….…………….…………….………9
3. Spatulate lobe (sl) terminating in two sharp brown spines, the outer spine slightly smaller and shorter than the inner one; lateral process of anterior coxal fold (alp) slender, slightly curving mesad; mesal process of anterior coxal fold (amp) almost as long as alp, flattened………….............…………………………………………...T. bispinispatula
− Spatulate lobe (sl) terminating in a single sharp dark brown spine..........………...................4
4. Telopodite without a lobe distal to fe; lateral process of anterior coxal fold (alp) long, slender, regularly curved, tip close to tip of opposite alp, the two together forming a circle; mesal process of anterior coxal fold (amp) straight, shorter than alp; femoral spine (fe) directed distad, pointed……………………………...….................................................. .....T.erectus
− Telopodite distally to fe with a large, round lobe (lo) projecting distolaterally………..........5
5. Lateral process of anterior coxal fold (alp) very slender, regularly curved………………..….6
− Lateral process of anterior coxal fold (alp) different, broader and/or with several apical denticles.....................................................................................................................................................8
6. Mesal margin of lateral process of anterior coxal fold (alp) with fine serrations; mesal process of anterior coxal fold (amp) almost as long as alp, broadly expanded, apically sharp, straight distad, mesal margin forming a strong longitudinal crest (lc) in posterior view……………………..………..…………..…………..……….T. navychula
− Mesal margin of lateral process of anterior coxal fold (alp) without serrations, tip of lateral process close to tip of the opposite side, the two together forming a circle………………………………….............................................................................. .7
7. Mesal process of posterior coxal fold (pmp) strongly developed along anterior-posterior axis.........................................................................................................T. floweri
− Mesal process of posterior coxal fold (pmp): slender, directed distolaterad…....T. forceps
8. Lateral process of anterior coxal fold (alp) broad, apically gradually narrowed; mesal process of anterior coxal fold (amp) almost as long as lateral process (alp), slender, straight, terminally slightly curved, pointed...........................................................................T. opinatus
− Lateral process of anterior coxal fold (alp) apically bent abruptly mesad, tip with serrate margins; mesal process of anterior coxal fold (amp) much shorter than lateral process (alp), directed mesodistad, simple, pointed; mesal process of posterior coxal fold (pmp): strongly developed along anterior-posterior axis………….………...…………...T. implicatus
9. Telopodite with a single femoral spine (fe)…………...………………………....….. …10
− Telopodite with two femoral spines (fe 1 and fe 2)…………………...………..………19
10. Mesal process of anterior coxal fold (amp) short……………………………..………..11
− Mesal process of anterior coxal fold (amp) long, slender ……..……..…..…..…....................13
11. Telopodite with slender tibial spine (ti), not curving mesad; fe curving backward, without small spine; mesal process of anterior coxal fold (amp) very short, pointed......T. peninsularis
− Telopodite with short, slender tibial spine (ti), curving mesad ………................….12
12. Femoral spine (fe) with a small, slender, pointed spine (sfe) at base (Fig. 3C); mesal process of anterior coxal fold (amp) short, forming a triangular process; telopodite distally to fe without a small round lobe (lo)…………………………………………T. payamense sp. nov.
− Femoral spine (fe) without a small slender, pointed spine (sfe) at base; telopodite distally to fe with a small round lobe (lo) projecting distolaterally................……………....................T. loxia
13. Lateral process of anterior coxal fold (alp) apically abruptly truncate……….. T. bearti
− Lateral process of anterior coxal fold (alp) apically pointed……………………….……..14
14. Mesal process of anterior coxal fold (amp) shorter than lateral process (alp)...……....…......15
− Mesal process of anterior coxal fold (amp) as long as lateral process (alp)…...….……...16
15. Mesal process of anterior coxal fold (amp) directed obliquely distomesad, slender, straight ............................................................................................................................... T. chelatus
− Mesal process of anterior coxal fold (amp) directed distad, thicker, slightly sigmoid…………………………………………………………….....T. brachyacanthus
16. Mesal process of anterior coxal fold (amp) directed obliquely distomesad, tip overlapping tip of opposite amp; lateral process of posterior coxal fold (plp) a massive, broad lobe, projecting laterad….…………………………………......................……T. sutchariti
− Mesal process of anterior coxal fold (amp) directed distad .... …....…..………………...17
17. Lateral process of anterior coxal fold (alp) apically without a crest; telopodite distally with a rounded lobe (lo); margins of spatulate lobe (sl) terminally meeting in a distinct angle ...................................................................................................................................T. bispinus
− Lateral process of anterior coxal fold (alp) apically with a crest…………….…….…….….18
18. Mesal process of anterior coxal fold (amp) apically irregularly tuberculate; telopodite distally without a rounded lobe (lo).................................................. ..…...................T. inflexus
− Mesal process of anterior coxal fold (amp) slender, straight, its tip pointed, its mesal margin forming a strong longitudinal crest (lc) in posterior view……T. mesocristatus
19. Anterior coxal fold (ac) with an additional spine-like process (aip) between alp and amp; lateral process of anterior coxal fold (alp) broad, mesal margin concave, tip with serrate margins, chicken comb-like; mesal process of anterior coxal fold (amp) much shorter than lateral process (alp), directed mesodistad, simple, pointed; both femoral spines (fe) slender, long.........................................................................................................................T. cristagalli
− Anterior coxal fold (ac) without an additional spine-like process (aip) between alp and amp……………………………………………………...……………….………….......20
20. Lateral process of anterior coxal fold (alp) apically without a crest, flattened, slightly curved, its laterodistal margin coarsely dentate, terminating in a short, sharp, pointed spine; mesal process (amp) much shorter than alp, directed distad, tip curving mesad, pointed; both femoral spines (fe 1, fe 2) long, curving backward; tibial spine (ti) long, not curving in horizontal plane…………………………………………………...……...………………T. culter
− Lateral process of anterior coxal fold (alp) apically with a crest extending caudad…………………………………………………………………………………...21
21. Lateral process (alp) flattened, curving mesad, laterodistal margin coarsely dentate, terminating in a short spine, tip curving against the tip of opposite side; mesal process (amp) much shorter than alp, slender, curving mesad; both femoral spines (fe 1, fe 2) broad, long; tibial spine (ti) long, curving in horizontal plane, not ending in a sharp spine..……….T. undulatus
− Lateral process (alp) regularly curved, terminating in a sharp, slightly upward pointing spine; mesal process (amp) slightly shorter than alp, flattend, straight, directed distad; tibial spine (ti) flattend, short, curving mesad…………………………T. planispina
22. Telopodite with a single femoral spine…………...………………………...…..……………23
− Telopodite with two femoral spines……………………………………….........……………25
23. Lateral process of anterior coxal fold (alp) without an apical crest; mesal process of anterior coxal fold (amp) shorter than and as broad as alp, directed distad; femoral spine (fe) very long and slender..................................................................................T. casjeekeli
− Lateral process of anterior coxal fold (alp), with a sharp crest on the posterior surface near the tip………………………………………………………………..……………...………..24
24. Lateral process of anterior coxal fold (alp) flattened, slightly curved, inflexed; femoral spine (fe) very long, slender, with an additional lamella at base…………. T. quadricuspis
− Lateral process of anterior coxal fold (alp) regularly curved, basally broad, gradually tapering towards end and ending in sharp point; femoral spine (fe) very long, slender, without an additional lamella at base….........………………...……..............................…T. cimi
25. Lateral process of anterior coxal fold (alp) flatten, broad..……………….………………....26
− Lateral process of anterior coxal fold (alp) slender, regularly curved, sickle-shaped ………………...……...……...…………….……………….……………………………....27
26. Lateral process of anterior coxal fold (alp) terminating in a very short external spine and a very long internal one; mesal process of anterior coxal fold (amp) as long as alp; first femoral spine (fe 1) very short, pointed; second femoral spine (fe 2) very long, as long as tibial spine (ti); an additional lamella at both side of base of fe 2.......………………...T. richardhoffmani
− Lateral process (alp) flattened, apically curved laterad as a short spine, lateral margin of alp slightly folded; mesal process (amp) shorter than alp, slender, straight, directed distad, pointed; the first femoral spine (fe 1) very short, directed upward, situated above fe 2, the second fe (fe2) very long, slender, curved downward…………………………………………….T. ursus
27. Mesal margin of lateral process of anterior coxal fold (alp) simple, without a caudad spine or crest; mesal process of anterior coxal fold (amp) much shorter than lateral process (alp), curved, pointed…………………………………………………...……………......T. enghoffi
− Mesal margin of lateral process of anterior coxal fold (alp) with a caudad small spine or crest.................................................................................................................................................28
28. Mesal margin of lateral process of anterior coxal fold (alp) with a small caudad crest; mesal process of anterior coxal fold (amp) slightly shorter than alp, slightly sigmoid, pointed……………………...……………...................…………………..T. bifurcus
−Mesal margin of lateral process of anterior coxal fold (alp) with a short curved caudad spine; mesal process of anterior coxal fold (amp) as long as alp, straight…...................T. demangei
Discussion
Morphologically, Thyropygus payamense sp. nov. undoubtedly belongs to the genus Thyropygus, as it has the diagnostic characteristics of the genus listed by Pimvichai, Enghoff & Panha (2009a). These include: (1) body rings that are not strongly wrinkled dorsally, (2) ozopores begining on body ring 6, (3) very long stigmatic grooves, (4) ventral soft pads on the postfemur and tibia of male walking legs, (5) a triangular gonopod sternum, (6) a gonopod telopodite with a femoral spine and often a tibial spine, (7) a prostatic groove terminating apically on a solenomere or prostatic lobe (apical palette of the telopodite), and (8) a voluminous apical palette that is more or less expanded and forms a gutter-like structure. Within the genus Thyropygus, T. payamense sp. nov. belongs to the T. allevatus group because it has a tibial and a femoral spine on the gonopod telopodite, with the tibial spine being notably long and recurved proximally toward the femoral spine. Finally, it is assigned to the T. opinatus subgroup because it has an additional projection on the anterior coxal fold.
The mean interspecific DNA sequence divergence values of T. payamense sp. nov. relative to other species in the T. allevatus group (mean values: 0.13 for COI and 0.11 for 16S rRNA) or the T. opinatus subgroup (mean values: 0.11 for COI and 0.08 for 16S rRNA) support the species-level distinction of T. payamense sp. nov. since they are of a comparable magnitude as the mean interspecific divergences for other species pairs in this group and subgroup (mean values: 0.12 for COI and 0.09 for 16S rRNA in the T. induratus subgroup; mean values: 0.11 for COI and 0.09 for 16S rRNA in the T. cuisinieri subgroup). The mean interspecific COI divergence values of T. payamense sp. nov. also align well with those observed in some genera of spirobolidan families, such as Pseudospirobolellidae with Coxobolellus Pimvichai, Enghoff, Panha & Backeljau, 2020 (mean 0.11; range: 0.06–0.15) (Pimvichai et al., 2020) and Siliquobolellus Pimvichai, Enghoff, Panha & Backeljau, 2022 (mean: 0.12; range: 0.08–0.15) (Pimvichai et al., 2022) or Pachybolidae with Atopochetus Attems, 1953 (mean: 0.14; range 0.09–0.17) and Litostrophus Chamberlin, 1921 (mean: 0.11; range 0.09–0.11) (Pimvichai et al., 2018).
The combination of its comparative DNA sequence divergence values, its phylogenetic placement as a well-supported clade, and its gonopodal differentiation, provide a solid basis to recognize T. payamense sp. nov. as a well-defined, separate species that complies at least with the morphological, biological, phylogenetic and lineage species concepts.
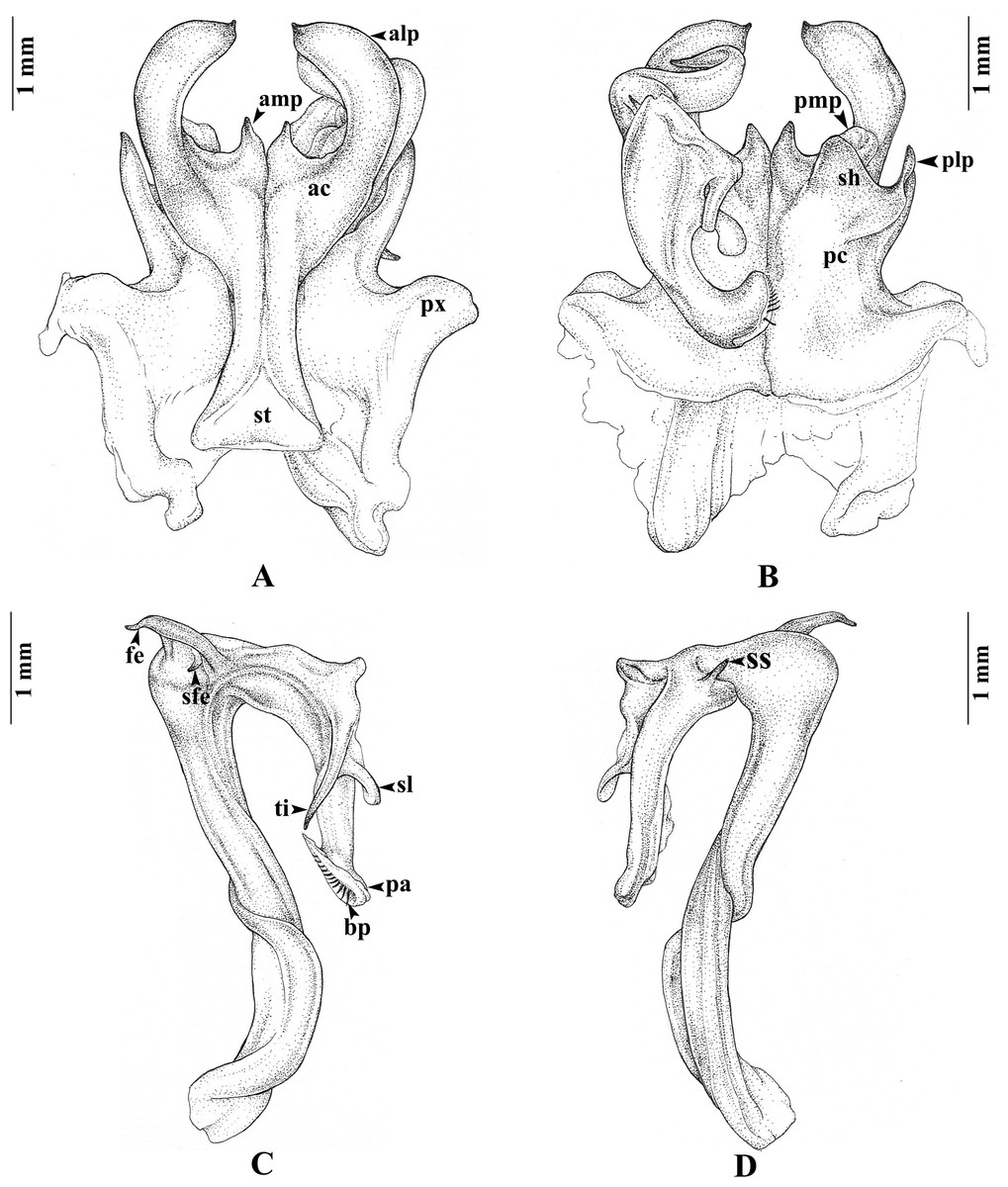
Figure 3: Thyropygus payamense sp. nov., holotype, gonopods (CUMZ - D00155).
(A) Anterior view, left telopodite removed. (B) Posterior view, left telopodite removed. (C) Left telopodite, posterior-mesal view, note the small, slender, pointed spine (sfe). (D) Left telopodite, anterior-lateral view.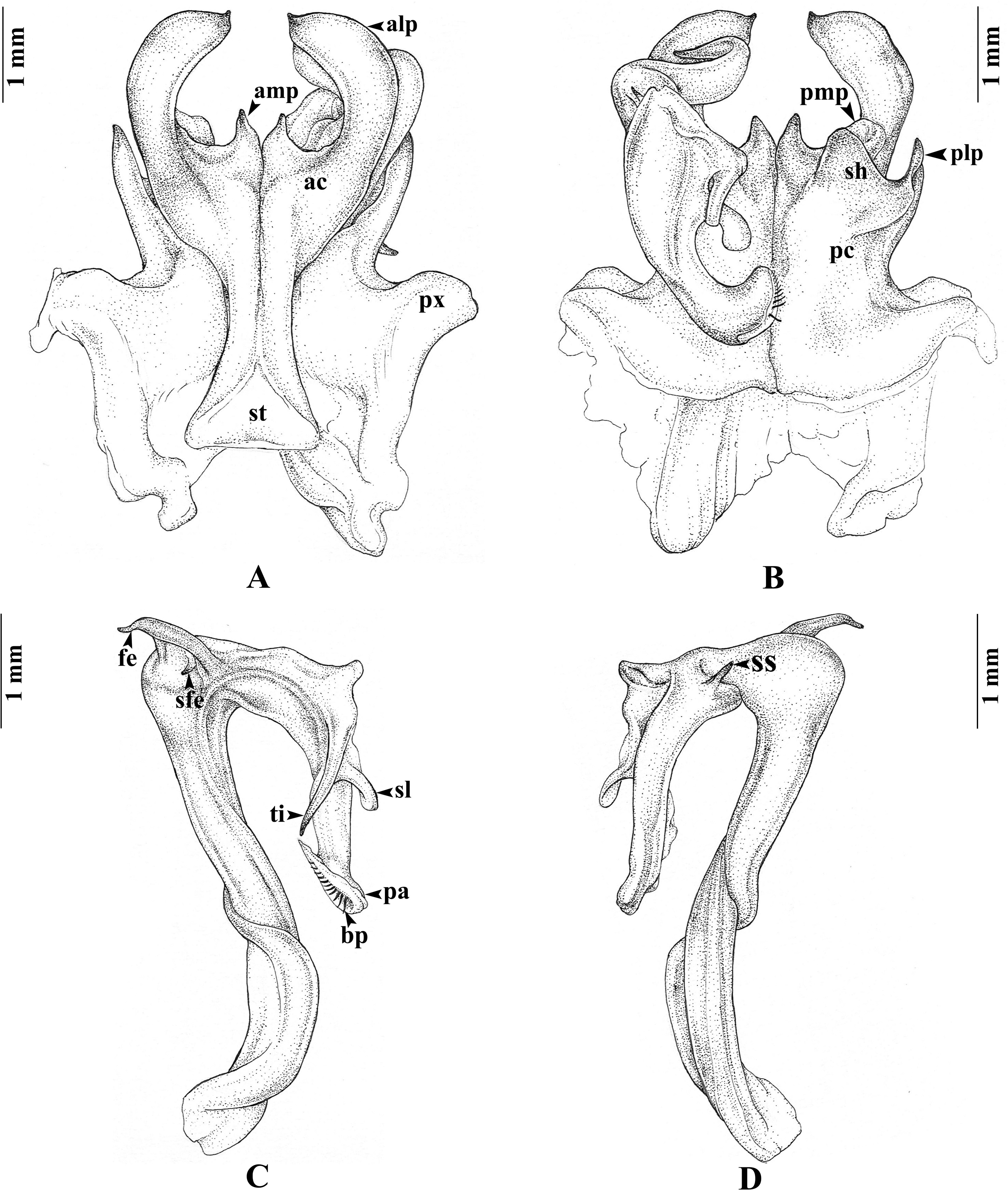{kind=link}
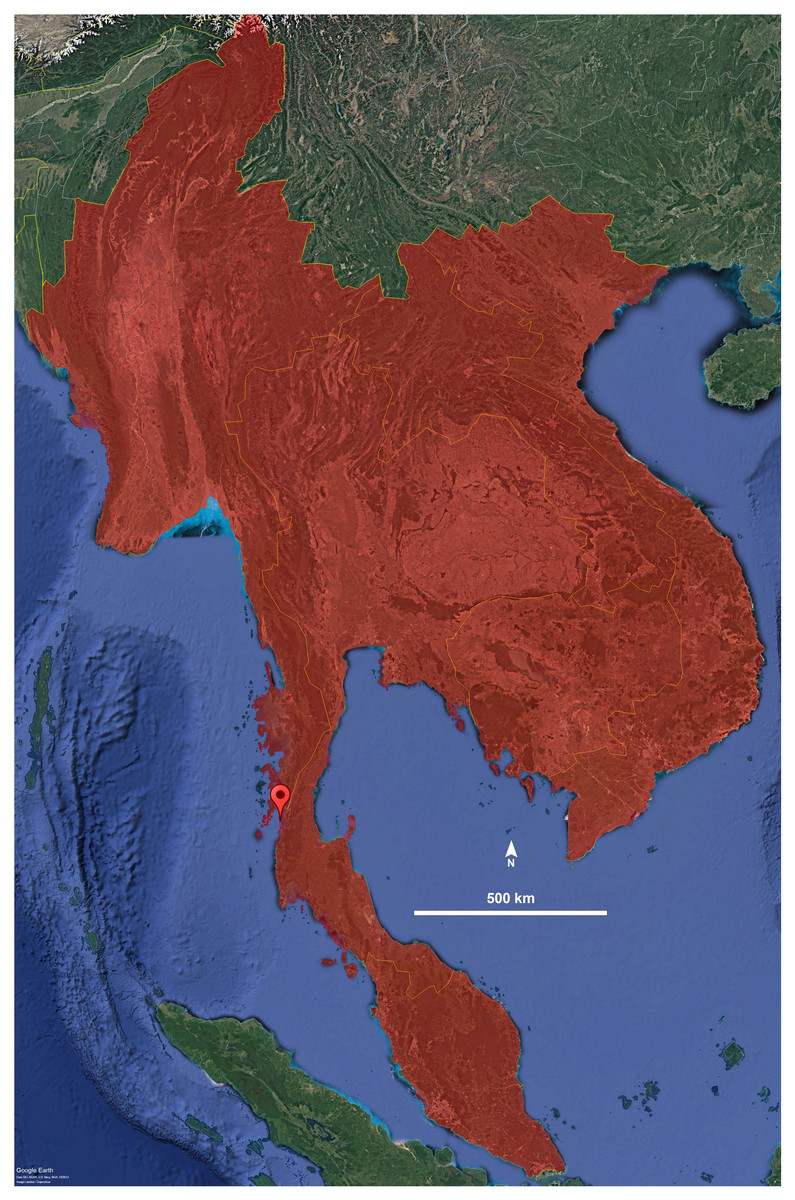
Figure 4: Distribution of the genus Thyropygus.
Droplet indicates the type locality of T. payamense sp. nov. Map generated using Google Earth Pro (Version 7.3.6.9796).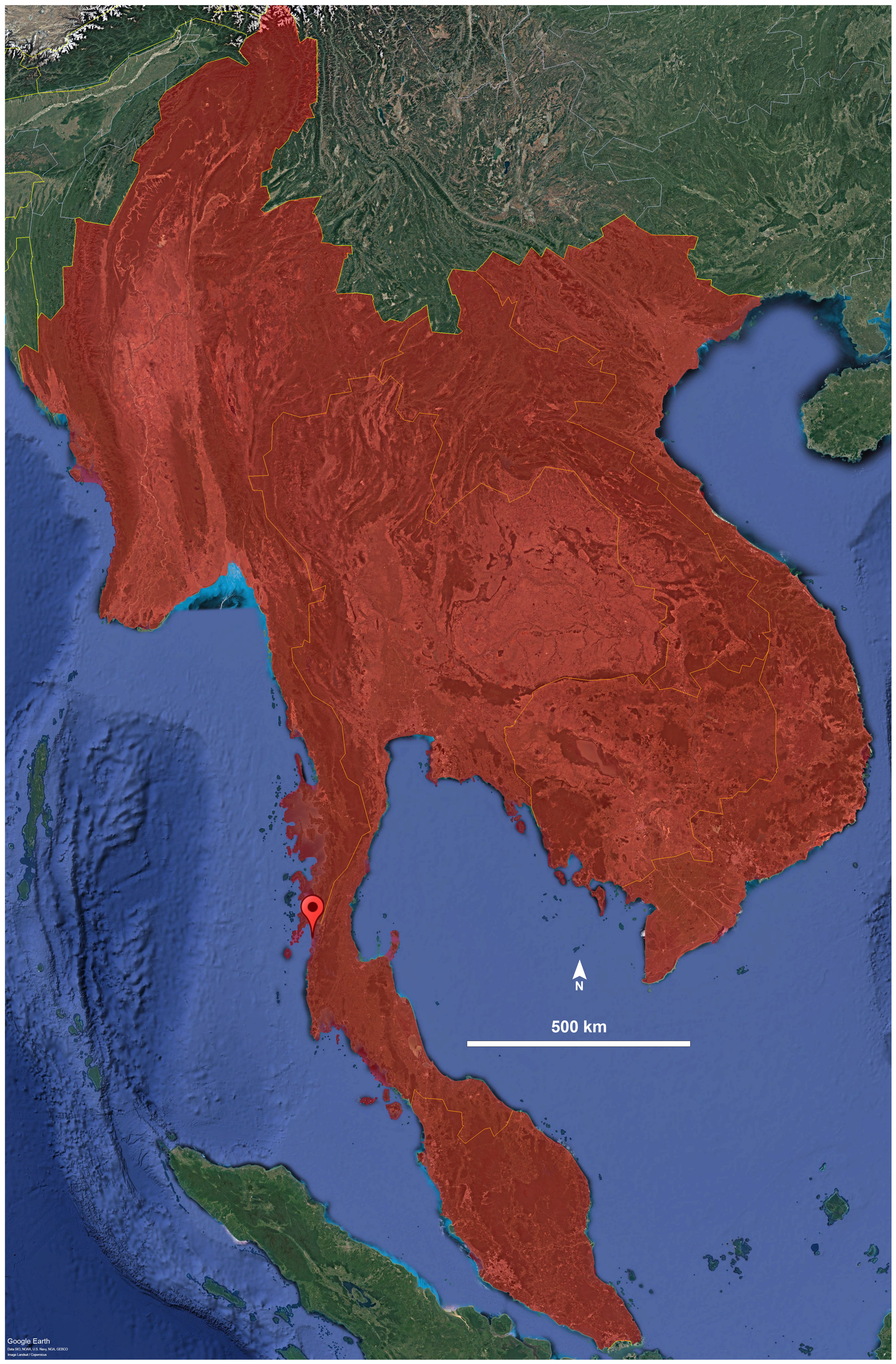{kind=link}
The addition of Thyropygus payamense sp. nov. (and T. peninsularis; see further below) to the T. opinatus subgroup did not affect the strong support for the monophyly of this subgroup, which now comprises 29 species. Hence, the congruence between morphological and DNA sequence data in the T. opinatus subgroup seems to be consistent and robust. It suggests that the defining, shared characters of this multi-species subgroup represent true synapomorphies. This contrasts sharply with the phylogenetic interpretation of the T. induratus subgroup, which was recently questioned because the discovery of two new species that morphologically clearly belong to this subgroup (T. panhai and T. somsaki) obliterated the support of its monophyly as inferred by COI sequence data. Hence the congruence between the morphological and DNA sequence data for the T. induratus subgroup was disrupted (Pimvichai, Enghoff & Backeljau, 2023).
The three clades within the T. opinatus subgroup identified in this study jointly form Clade 1A3 described by Pimvichai et al. (2016), with the inclusion of T. payamense sp. nov. and T. peninsularis. It is striking that the Thai members of the T. opinatus subgroup only occur in southern Thailand (Clades 1, 2, and nsa), except for the two species of clade 3, which are distributed in northern, central and western Thailand. Conversely, no species from the other subgroups of the T. allevatus group were hitherto found in southern Thailand.
Southern Thailand, part of the Sundaland biogeographic region, is characterized by a unique mix of fauna influenced by its peninsular geography, tropical climate, and historical land connections to surrounding regions (Parnell, 2013). As such, the present data tentatively suggest that T. opinatus subgroup clades 1, 2 and the nsa jointly may represent an endemic species radiation in the peninsular area of Thailand, Malaysia and Myanmar. Yet, further phylogeographic analyses incorporating a broader sampling of populations, taxa and DNA markers are needed to infer the precise evolutionary and biogeographical history of these species.
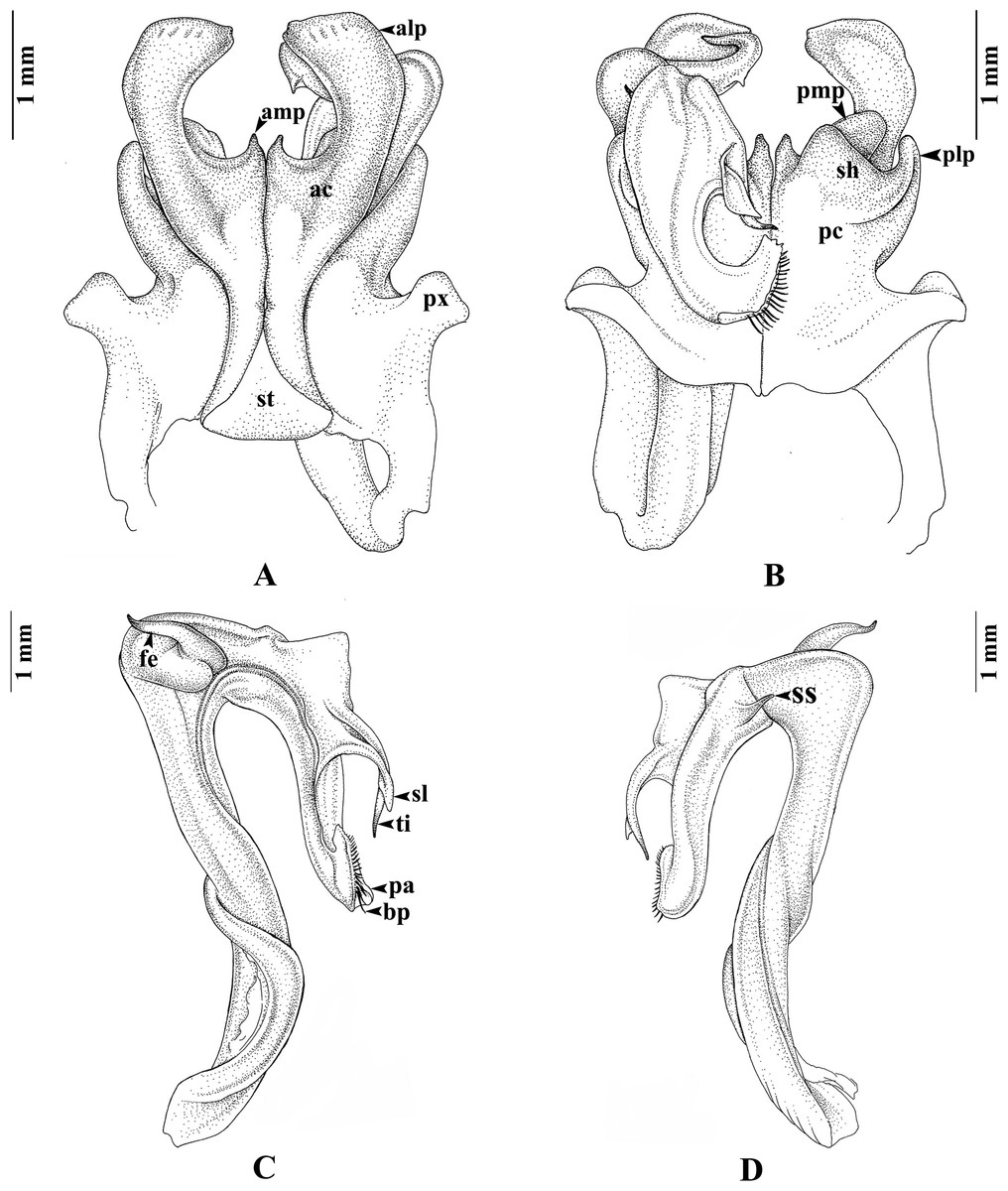
Figure 5: Thyropygus peninsularis, gonopods.
Thyropygus peninsularis, specimen from Wang-Matcha, Kapoe, Ranong, Thailand, gonopods (CUMZ - D00011). (A) Anterior view, left telopodite removed. (B) Posterior view, left telopodite removed. (C) Left telopodite, posterior-mesal view. (D) Left telopodite, anterior-lateral view (modified from Panha, Pimvichai & Enghoff, 2009).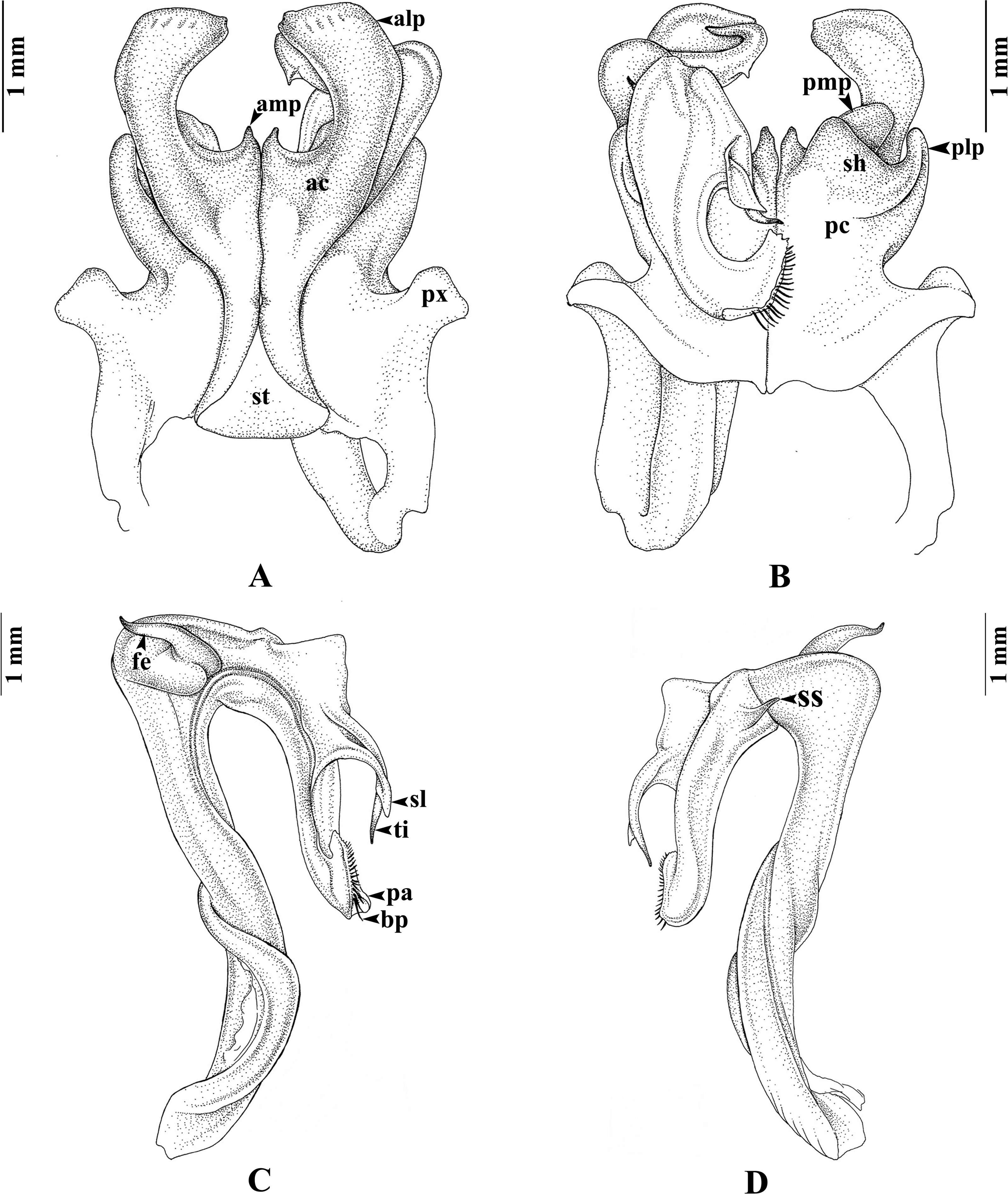{kind=link}
Thyropygus peninsularis was initially suggested to belong to the T. erythropleurus group by Hoffman (1982), because it has no recurved tibial spine proximally directed towards the femoral spine—a defining feature of the T. allevatus group. Therefore, Pimvichai, Enghoff & Panha (2009a) followed Hoffman (1982) and did not include T. peninsularis, in the T. allevatus group. However, T. peninsularis possesses a small spatulate lobe at the apical part of the telopodite, along with a very short additional mesal projection on the gonopod’s anterior coxal lobe (Fig. 5), similar to T. loxia. These features are shared by most species in the T. opinatus subgroup. Furthermore, DNA sequence analysis (COI and 16S rRNA) firmly placed T. peninsularis within the T. opinatus subgroup (Pimvichai, Enghoff & Panha, 2014, present results). Based on these morphological and DNA sequence data, we formally confirm the assignment of T. peninsularis to the T. opinatus subgroup, as was implicitly done by Pimvichai, Enghoff & Backeljau (2023). These findings highlight, once more, the importance of integrating morphological and molecular data for resolving and/or re-interpreting taxonomic ambiguities.
Conclusions
While the support for the monophyly of some species subgroups within the Thyropygus allevatus group disappears by increased species sampling, the high support for the monophyly of the T. opinatus subgroup remains unaffected after increased species sampling by the inclusion of (1) T. payamense sp. nov., described in this study, and (2) T. peninsularis, a species formerly assigned to the T. erythropleurus group, but for which DNA sequence data and a re-interpretation of its gonopod morphology show that it actually belongs to the T. opinatus subgroup. As a consequence, the congruence between the DNA sequence data and the defining synapomorphies in gonopod morphology remains consistent and robust in the T. opinatus subgroup, which now comprises 29 species. While it is too early to draw firm phylogeographic conclusions, these data tentatively suggest that with the exception of T. bispinus and T. inflexus, the T. opinatus subgroup may represent an endemic species radiation in the peninsular area of Thailand, Malaysia and Myanmar. Finally, the results illustrate the importance of combining further species sampling with integrative research to resolve taxonomic ambiguities and explore evolutionary relationships in these millipedes.
Supplemental Information
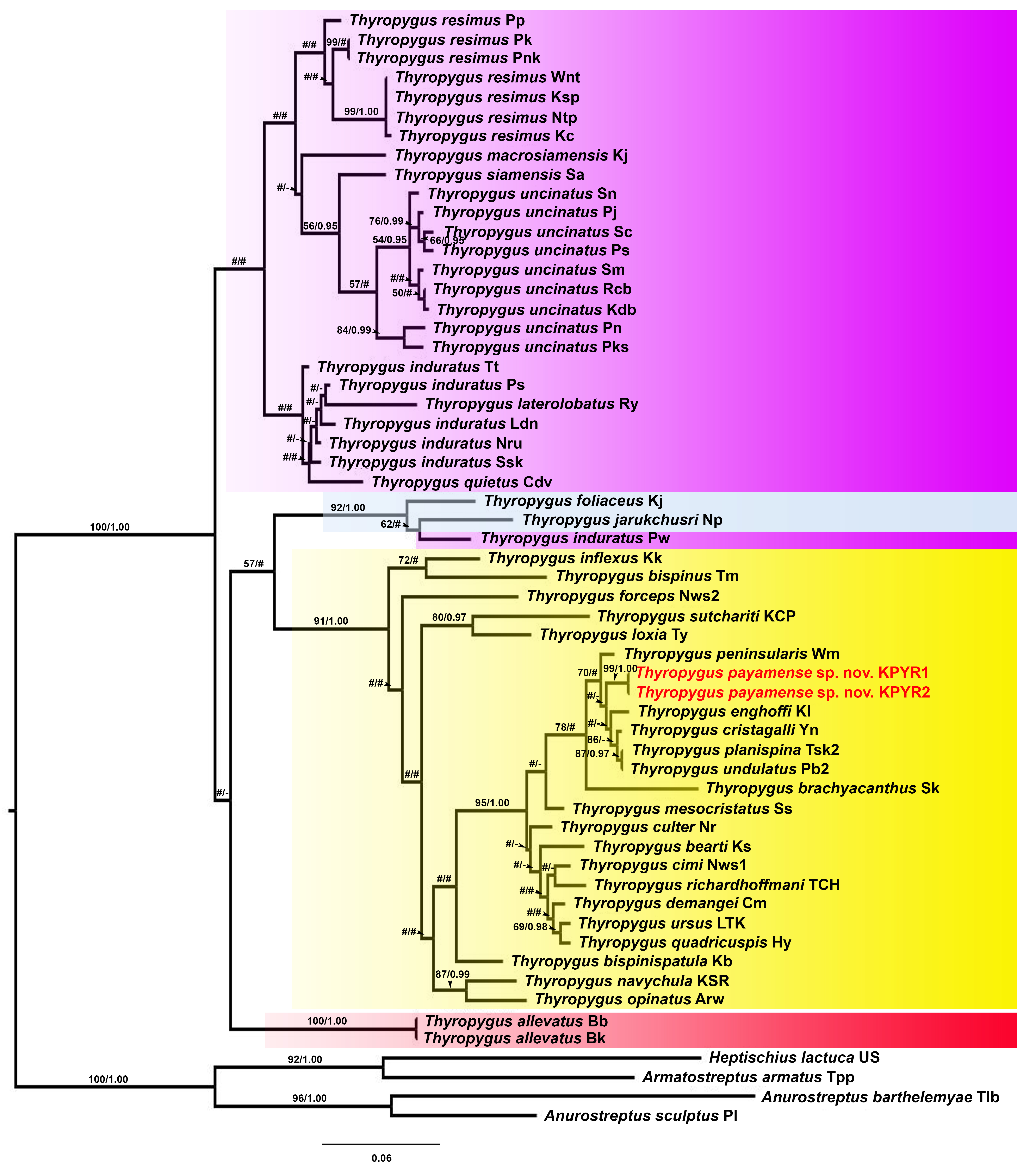
Phylogenetic relationships of Thyropygus species based on maximum likelihood analysis (ML) and Bayesian Inference (BI) of 660 bp of the COI alignment
Numbers at nodes indicate node support based on bootstrapping (ML)/posterior probabilities (BI). Scale bar = 0.05 substitutions/site. # indicates nodes with < 50% bootstrap support and < 0.95 posterior probability. - indicates non - supported nodes. The colored areas mark the T. induratus subgroup (purple), T. cuisinieri subgroup (blue), T. opinatus subgroup (yellow), and T. allevatus subgroup (red). Abbreviations after species names refer to locality names as shown in Table 1.
{kind=link}
Phylogenetic relationships of Thyropygus species based on maximum likelihood analysis (ML) and Bayesian Inference (BI) of 487 bp of the 16S rRNA alignment
Numbers at nodes indicate node support based on bootstrapping (ML)/posterior probabilities (BI). Scale bar = 0.05 substitutions/site. # indicates nodes with <50% bootstrap support and <0.95 posterior probability. - indicates non - supported nodes. The colored areas mark the T. induratus subgroup (purple), T. cuisinieri subgroup (blue), T. opinatus subgroup (yellow), and T. allevatus subgroup (red). Abbreviations after species names refer to locality names as shown in Table 1.
{kind=link}
Specimens from which the COI and 16S rRNA gene fragments were sequenced
CUMZ, Museum of Zoology, Chulalongkorn University, Bangkok, Thailand; NHMW, Naturhistorisches Museum, Vienna, Austria. Names of provinces are capitalized. Abbreviations after species names refer to the isolate of each sequence. GenBank accession numbers are indicated for each species.