
Two new species of Microlaimidae (Nematoda: Microlaimida) from the Continental Shelf off Northeastern Brazil (Atlantic Ocean) with emended diagnosis and dichotomous key
- Published
- Accepted
- Received
- Academic Editor
- Khor Waiho
- Subject Areas
- Biodiversity, Taxonomy, Zoology
- Keywords
- Spirobolbolaimus, Ixonema, South Atlantic, Taxonomy, Nematode diversity
- Copyright
- © 2024 Manoel et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Two new species of Microlaimidae (Nematoda: Microlaimida) from the Continental Shelf off Northeastern Brazil (Atlantic Ocean) with emended diagnosis and dichotomous key. PeerJ 12:e17976 https://doi.org/10.7717/peerj.17976
Abstract
New species of the genera Spirobolbolaimus and Ixonema (Nematoda: Microlaimidae) have been found in sediment samples collected in the South Atlantic, along the Continental Shelf break off Northeastern Brazil. Different to other Spirobolbolaimus species, S. pernambucanus sp. nov. possesses six outer labial setae and four cephalic setae with approximately the same length. Ixonema gracieleae sp. nov. differs from other species of Ixonema in having somatic setae on peduncles. This is the first time that new species of these taxa have been described for the Brazilian coast. An amendment of the diagnosis and a dichotomous key are proposed for both genera.
Introduction
The family Microlaimidae (Micoletzky, 1922) currently comprises 13 genera and about 150 valid species (Tchesunov, Jeong & Lee, 2021; Nemys, 2024). Records of species belonging to this family are still scarce from the Brazilian coast and are almost all related to Microlaimus De Man, 1880 (Manoel, Neres & Esteves, 2024). The oldest records in which species related to any genus placed in Microlaimidae were originally described for this location date back to the 1950s, when Gerlach described four species of Microlaimus for southeastern coast of Brazil (Gerlach, 1956; Gerlach, 1957a; Gerlach, 1957b). Three decades later, Lima, Neres & Esteves (2022) described three species of Microlaimus from the continental shelf off the Campos Basin, Southeastern Brazil. Recently, Manoel, Neres & Esteves (2024) described three new species of Microlaimus for the Continental Shelf break off Northeastern Brazil.
For other genera of Microlaimidae, such as Spirobolbolaimus Soetaert & Vincx, 1988 and Ixonema Lorenzen, 1971, there are no records of species originally described for the South Atlantic. Spirobolbolaimus was placed in Microlaimidae based on male (two opposite and outstretched testes) and female (two outstretched ovaries) gonads (Soetaert & Vincx, 1988). Spirobolbolaimus is mainly characterized by having outer labial setae longer than the cephalic setae, multispiral amphidial fovea, postamphidial setae in six or eight longitudinal rows, buccal cavity armed with protruding teeth and pharynx with anterior and posterior bulbs (Soetaert & Vincx, 1988; Gourbault & Vincx, 1990; Shi & Xu, 2016). The genus currently has three valid species: S. bathyalis Soetaert & Vincx, 1988 (Mediterranean, Calvi Bay; 280–820 m depth); S. boucherorum Gourbault & Vincx, 1990 (Caribbean Sea, Guadeloupe; 2 m depth) and S. undulatus Shi & Xu, 2016 (China Sea, Nanji Islands; Dasha’ao sand beach).
Ixonema is the only representative of the family Microlaimidae that has three caudal glands with separate outlets, a feature considered rare in free-living marine nematodes (Lorenzen, 1971; Lorenzen, 1994). Often, representatives of this taxon are found with the cuticle covered by particles, such as algae, suggesting that these organisms are capable of carrying their own food (Steyaert et al., 1999). Additionally, this genus is characterized by a narrow and elongated anterior end, amphidial fovea far from the anterior end with a corpus gelatum projecting from the amphidial opening, a small mouth cavity and three minute teeth (Tchesunov, 2014). Only three species have been described for Ixonema: I. sordidum Lorenzen, 1971 (North Sea, German Bight; sublittoral region, depth unspecified); I. powelli Jensen, 1985 (Gulf of Mexico; 72 m depth) and I. deleyi Muthumbi & Vincx, 1999 (Indian Ocean, Kenyan coast; 21–2,007 m depth).
Here we describe two new species, one from Spirobolbolaimus and another from Ixonema, found along the break of the continental shelf off Northeastern Brazil. Amendments to the diagnosis and dichotomous keys are proposed for both genera.
Material and methods
Study area and sampling. This information were previously described in Manoel, Neres & Esteves (2024).
Laboratory processing. In the laboratory, sediment samples were sieved using a 500 µm mesh followed by a 45 µm mesh sieve which was used to retain the meiobenthic organisms. The samples remaining in the 45 µm mesh were extracted with colloidal silica (Somerfield, Warwick & Moens, 2005).
Nematoda were counted (and removed) under a stereomicroscope using a Dolffus plate. All individuals were transferred to a small glass container containing a solution with 99% formaldehyde (4%) + 1% glycerin (Solution 1 –De Grisse, 1969). The methodology of transferring each animal to glycerin was then applied, followed by diaphanization, according to the method described by De Grisse (1969). The individuals were mounted permanently on glass slides, as an adaptation of the method described by Cobb (1920). The genus was identified by using keys provided by Warwick, Platt & Somerfield (1998) and Decraemer & Smol (2006). Species were identified through the comparison of their characteristics with those provided in the original descriptions. Drawings were made with the aid of an Olympus CX 31 optical microscope fitted with a drawing tube. Body measurements were taken using a mechanical map meter. The holotype and one paratype (female) of each species are deposited in the Nematoda Collection of the Museum of Oceanography Prof. Petronio Alves Coelho (MOUFPE), Brazil. Other paratypes are deposited in the Meiofauna Laboratory, Zoology Department, Federal University of Pernambuco (NM LMZOO-UFPE).
The electronic version of this article in Portable Document Format (PDF) will represent a published study according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under the Code from the electronic edition alone. This published research and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org: pub: urn:lsid:zoobank.org:pub:8B5A29A6-0EF8-454C-A62F-8C0B59390B83. The online version of this research is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Results
Systematics
| Class CHROMADOREA Inglis, 1983 |
| Subclass CHROMADORIA Pearse, 1942 |
| Order Microlaimida Leduc, Verdon & Zhao, 2018 |
| Superfamily Microlaimoidea Micoletzky, 1922 |
| Family Microlaimidae Micoletzky, 1922 |
| Genus SpirobolbolaimusSoetaert & Vincx, 1988 |
Diagnosis. (Emended from Shi & Xu, 2016) Microlaimidae. Cuticle annulated. Anterior sensilla in three circles: six inner labial setae papilliform; six stout outer labial setae (sometimes jointed); and four cephalic setae shorter or similar in length to the six outer labial setae. Buccal cavity large, armed with a well-developed dorsal tooth and a pair of ventrosublateral teeth. Additional lateral teeth may be present. Amphidial fovea multispiral and ventrally wound, sclerotized. Postamphidial setae in six or eight longitudinal rows. Pharynx with anterior bulb. Oval posterior bulb present or absent. Females with two outstretched ovaries. Males with two opposed testes. Copulatory apparatus strongly sclerotized. Gubernaculum with or without apophysis. Precloacal supplements (papilliform or small pores) present or absent. Oval, elongated or banana-shaped sperm cells. Tail conical.
Type species: Spirobolbolaimus bathyalis Soetaert & Vincx, 1988.
List of valid species of SpirobolbolaimusSoetaert & Vincx, 1988
| Spirobolbolaimus bathyalisSoetaert & Vincx, 1988 |
| Spirobolbolaimus boucherorumGourbault & Vincx, 1990 |
| Spirobolbolaimus pernambucanussp. nov. |
| Spirobolbolaimus undulatusShi & Xu, 2016 |
Description of new species
Spirobolbolaimus pernambucanus sp. nov.
| Spirobolbolaimus pernambucanussp. nov. | Holotype male | Male paratype (n = 1) | Females paratypes (n = 3) |
|---|---|---|---|
| Body length | 1,716 | 1,848 | 1,686–1,800 |
| Inner labial papillae length | 2 | – | 2 |
| Outer labial setae length | 7 | 7 | 7 |
| Cephalic setae length | 7 | 7 | 5.5–7 |
| Head diameter at level of the cephalic setae | 34 | 34 | 35–36 |
| Cephalic setae in relation to head diameter (%) | 21% | 21% | 15–19% |
| Distance from anterior end to amphidial fovea | 12 | 13 | 8–10.5 |
| Distance from anterior end to amphidial fovea in relation to head diameter | 0.4 | 0.4 | 0.2–0.3 |
| Amphidial fovea diameter (maximum width) | 12 | 12 | 11–12 |
| Body diameter at level of the amphidial fovea | 34 | 34 | 34.5–37 |
| % of the amphidial fovea diameter in relation to corresponding body diameter | 36% | 36% | 31–35% |
| Pharynx length | 220 | 223 | 226.5–241.5 |
| Position of nerve ring from anterior end | 120 | 133.5 | 125–138 |
| Nerve ring position in relation to pharynx length (%) | 55% | 60% | 55–56% |
| Pharyngeal bulb diameter | 26 | 30 | 28.5–31 |
| Body diameter at level of the pharyngeal bulb | 38 | 40 | 44–46.5 |
| % of basal bulb diameter in relation to corresponding body diameter | 67% | 75% | 64–66% |
| Maximum body diameter | 40 | 44.5 | 46.5–48 |
| Anal or cloacal body diameter | 31 | 36 | 32–33 |
| Tail length | 120 | 114 | 117–131 |
| Length of spicules along arc | 53 | 55 | * |
| Length of spicules along cord | 40 | 42 | * |
| Length of gubernaculum | 19.5 | 19 | * |
| Length of gubernaculum in relation to length of spicules along arc | 37% | 35% | * |
| Length of spicules along arc in relation to cloacal body diameter | 1.7 | 1.5 | * |
| Distance from anterior end to vulva | * | * | 867–984 |
| Position of vulva from anterior end (%) | * | * | 51–55% |
| Body diameter in vulva region | * | * | 46.5–48 |
| Anterior ovary length | * | * | 354–408 |
| Posterior ovary length | * | * | 252–288 |
| Reproductive system length | 927 | 1,104 | 642–666 |
| % of reproductive system in relation to body length | 54% | 60% | 36–39% |
| a | 43 | 42 | 35–39 |
| b | 8 | 8 | 7–7.5 |
| c | 14 | 16 | 13–14 |
| c’ | 4 | 3 | 4 |
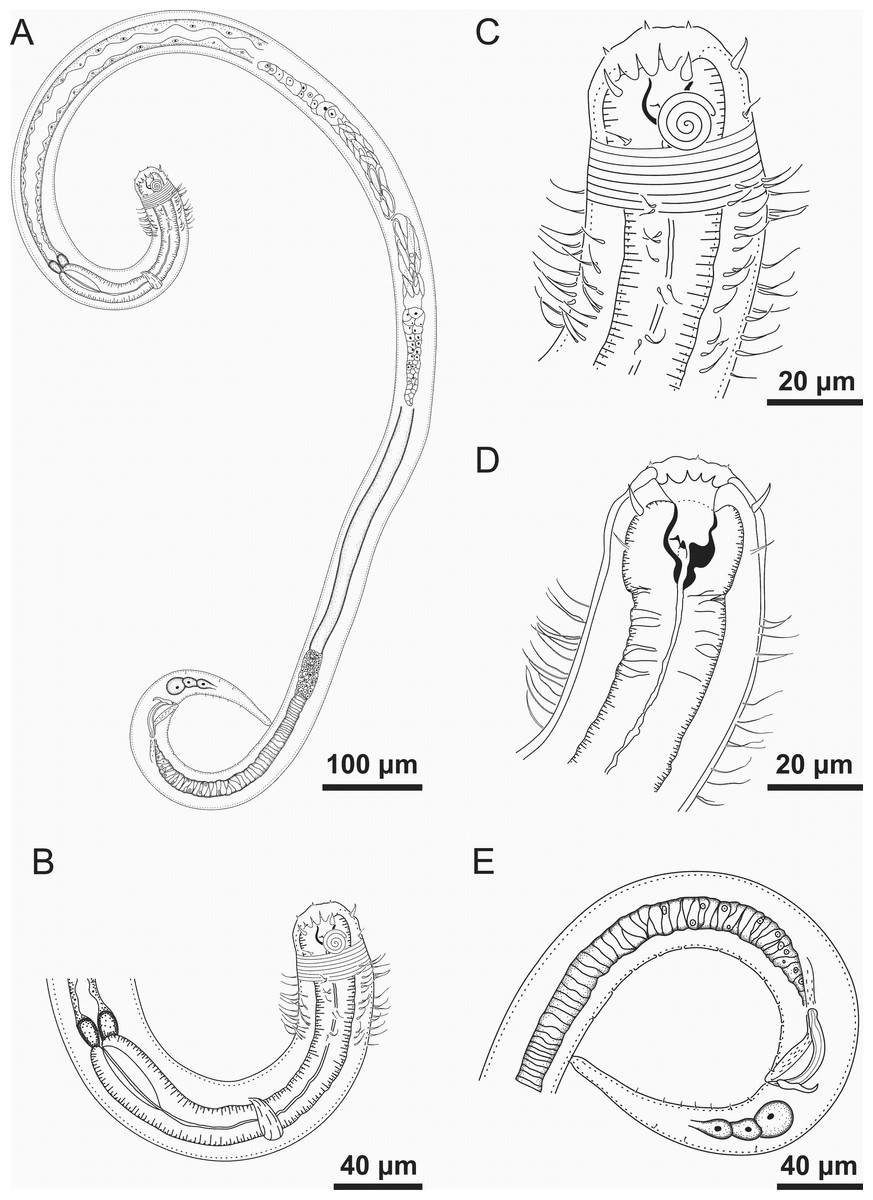
Figure 1: Spirobolbolaimus pernambucanus sp. nov. holotype male.
Holotype male: (A) overview; (B) anterior region; (C) anterior end (sensilla disposition, amphidial fovea and cervical setae); (D) anterior end (buccal cavity); (E) posterior region. Image source credit: Alex Manoel.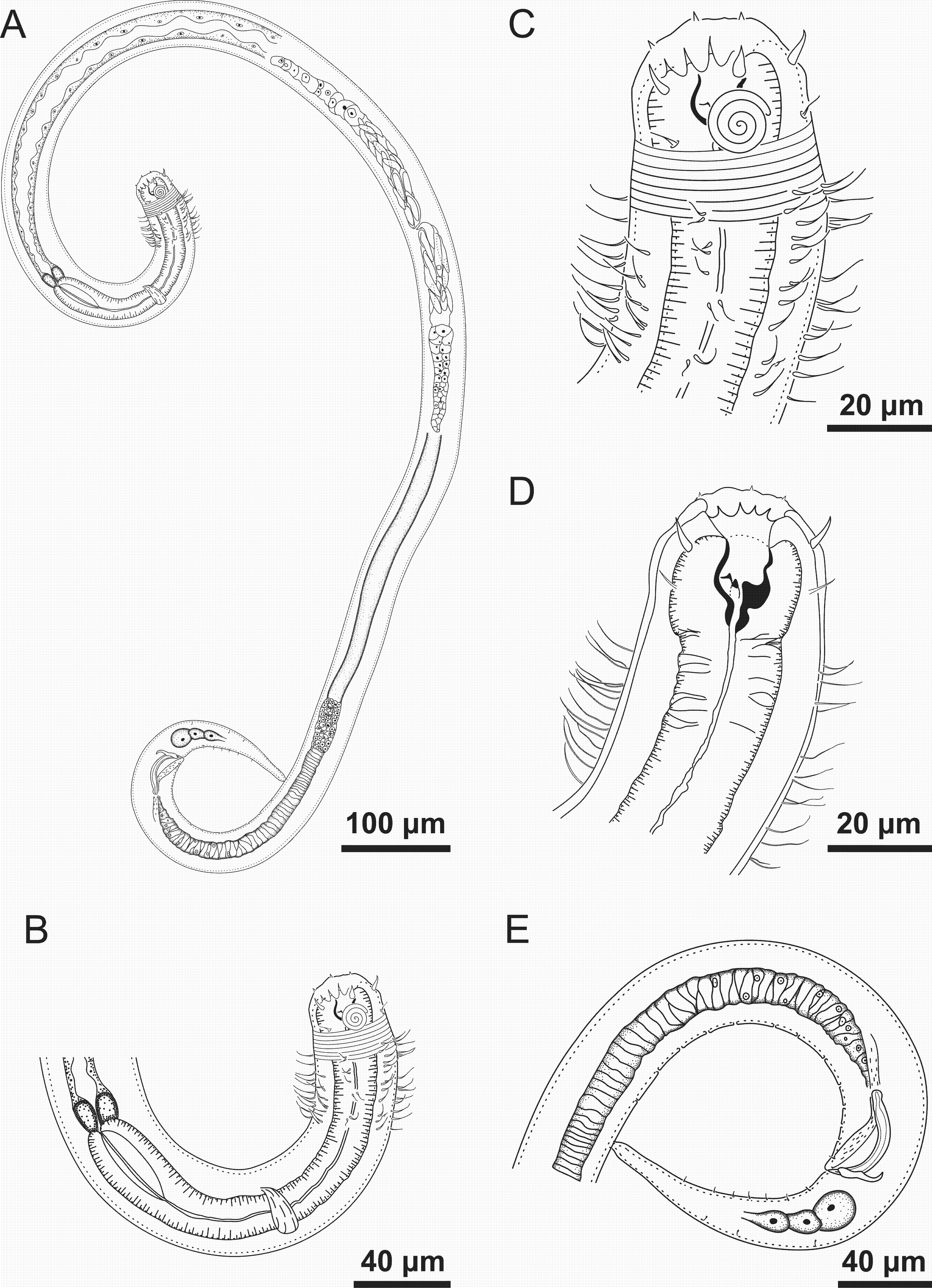{kind=link}
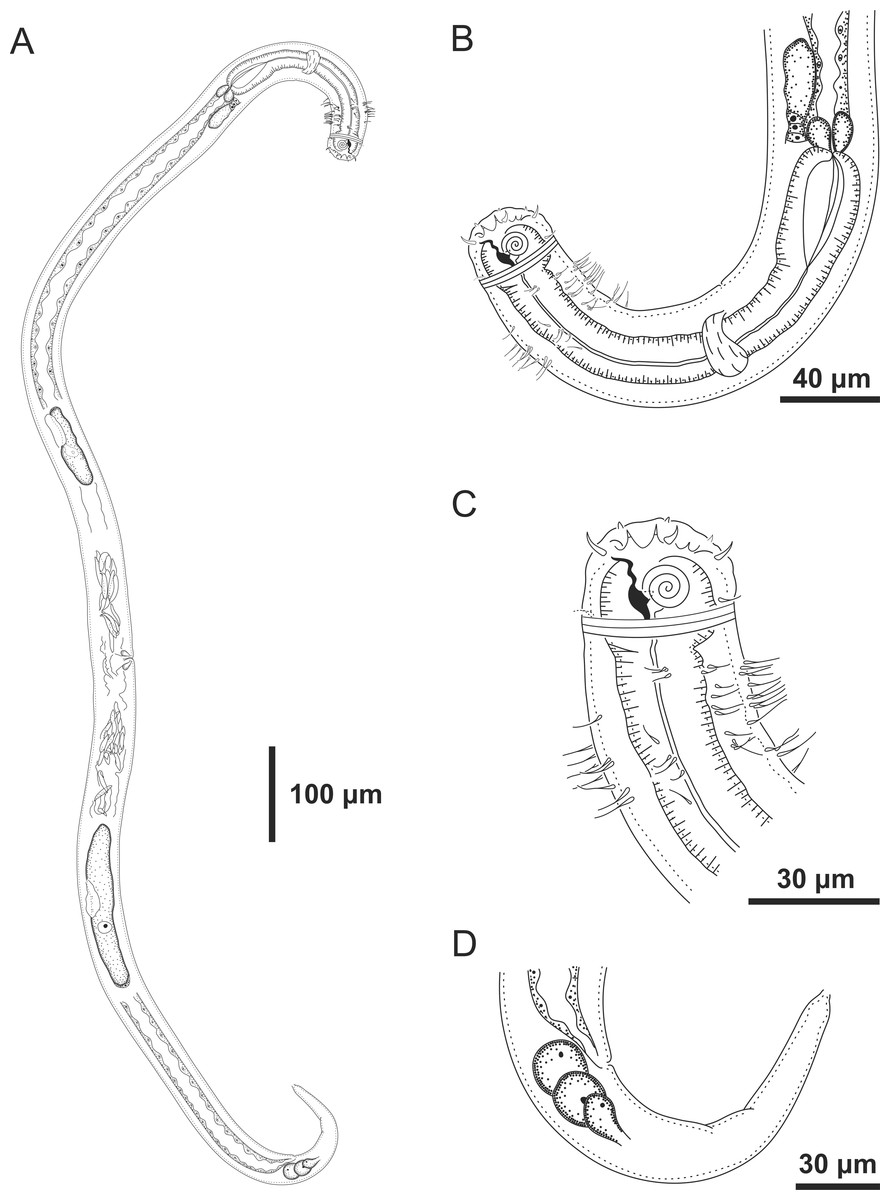
Figure 2: Spirobolbolaimus pernambucanus sp. nov. paratype female.
Paratype female: (A) overview; (B) anterior region; (C) anterior end (sensilla disposition, amphidial fovea and cervical setae); (D) tail. Image source credit: Alex Manoel.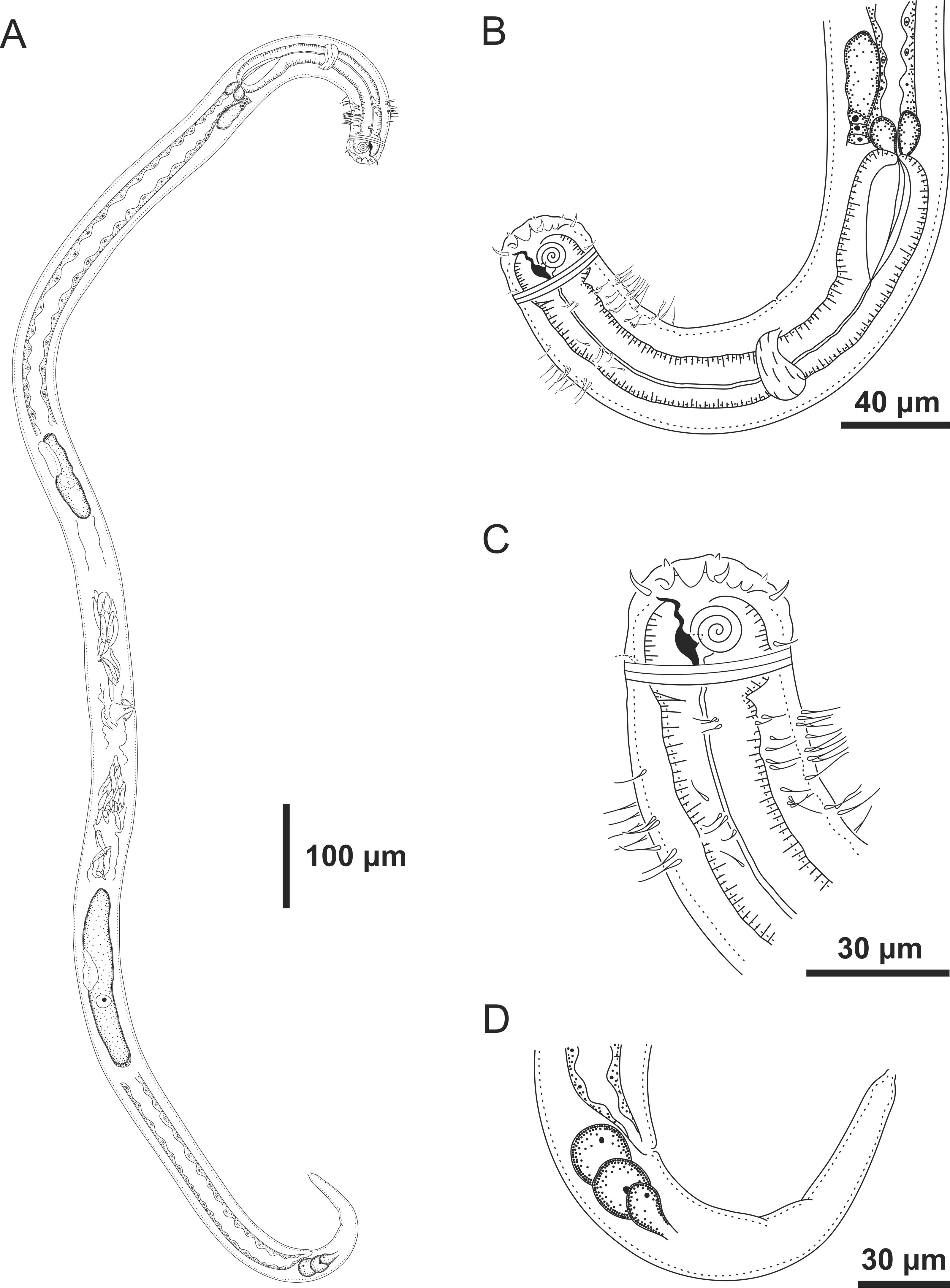{kind=link}
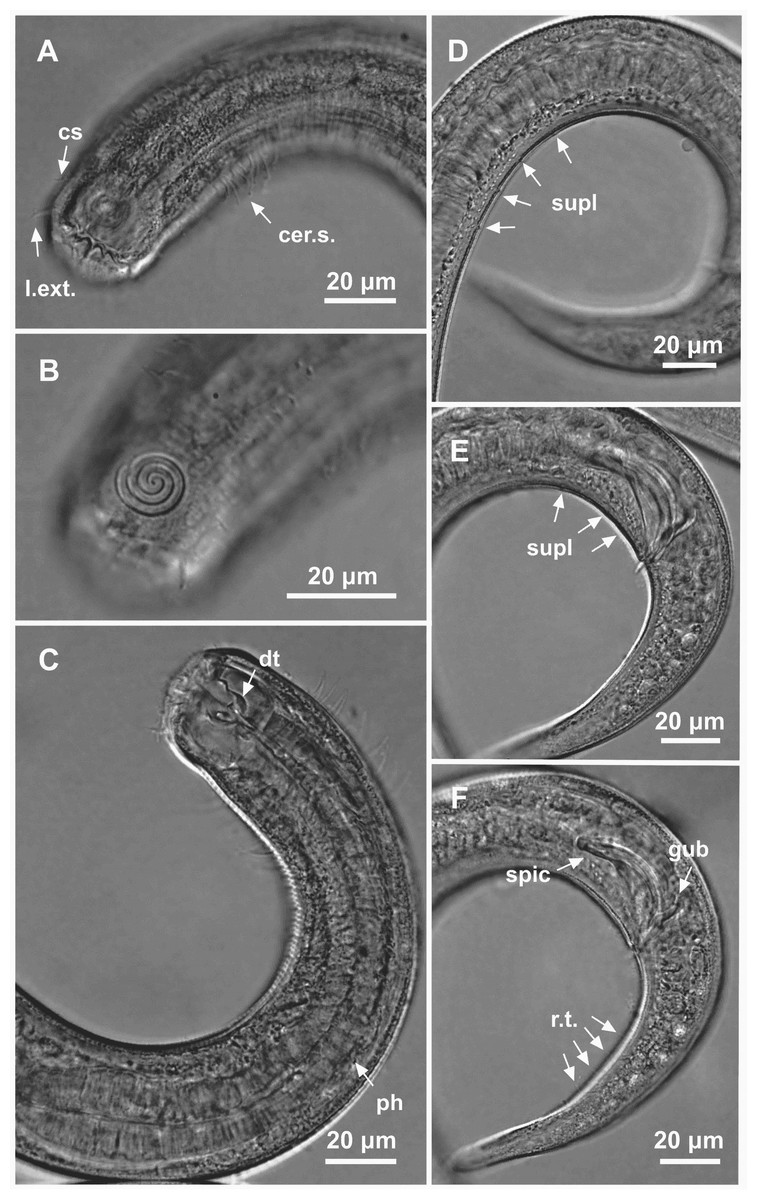
Figure 3: Spirobolbolaimus pernambucanus sp. nov.: holotype male.
Holotype male: (A) anterior region: arrows indicating outer labial setae (l.ext.), cephalic setae (cs) and cervical setae (cer. s.); (B) anterior region (amphidial fovea); (C) anterior end: arrows indicating the dorsal tooth (dt) and pharynx (ph); (D) posterior end: arrows indicating pore-like precloacal supplements farthest from the cloaca; (E) posterior end: arrows indicating pore-like precloacal supplements closer to the cloaca; (F) posterior end: arrow indicating the spicule (spic), gubernaculum (gub) and rows of setae on the tail (r.t.).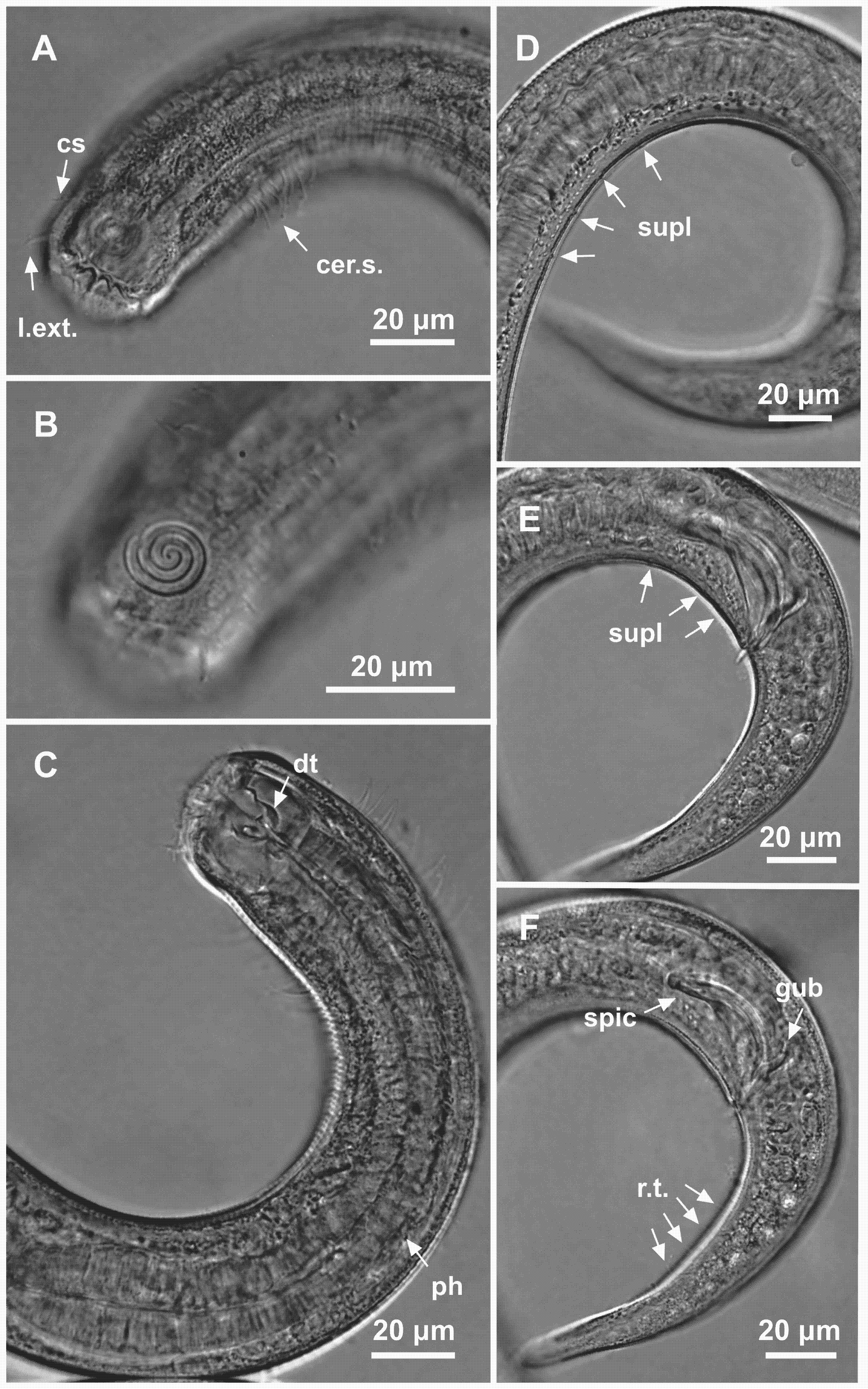{kind=link}
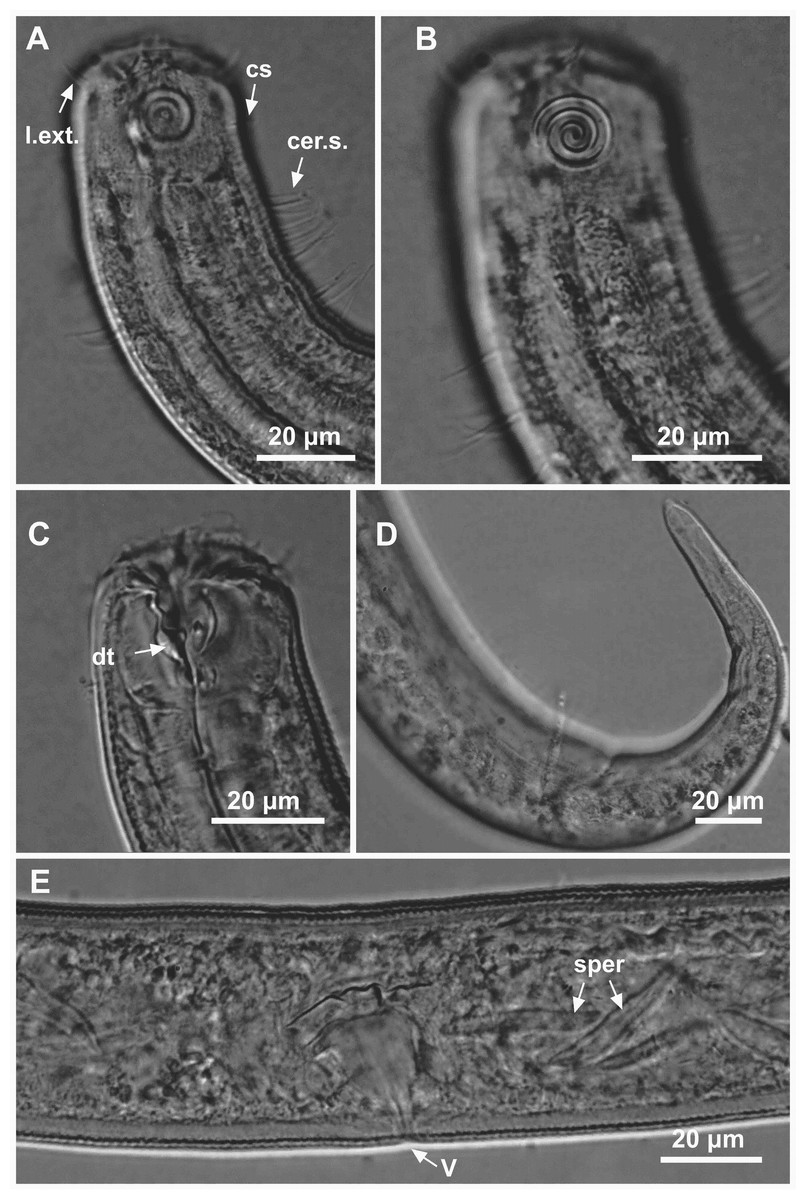
Figure 4: Spirobolbolaimus pernambucanus sp. nov. paratype female.
Paratype female: (A) anterior region: arrows indicating outer labial setae (l.ext.), cephalic setae (cs) and cervical setae (cer. s.); (B) anterior region (amphidial fovea); (C) anterior end: arrow indicating the dorsal tooth (dt); (D) tail; (E) vulva region: arrows indicating the vulva (V) and sperm (sper). Image source credit: Alex Manoel.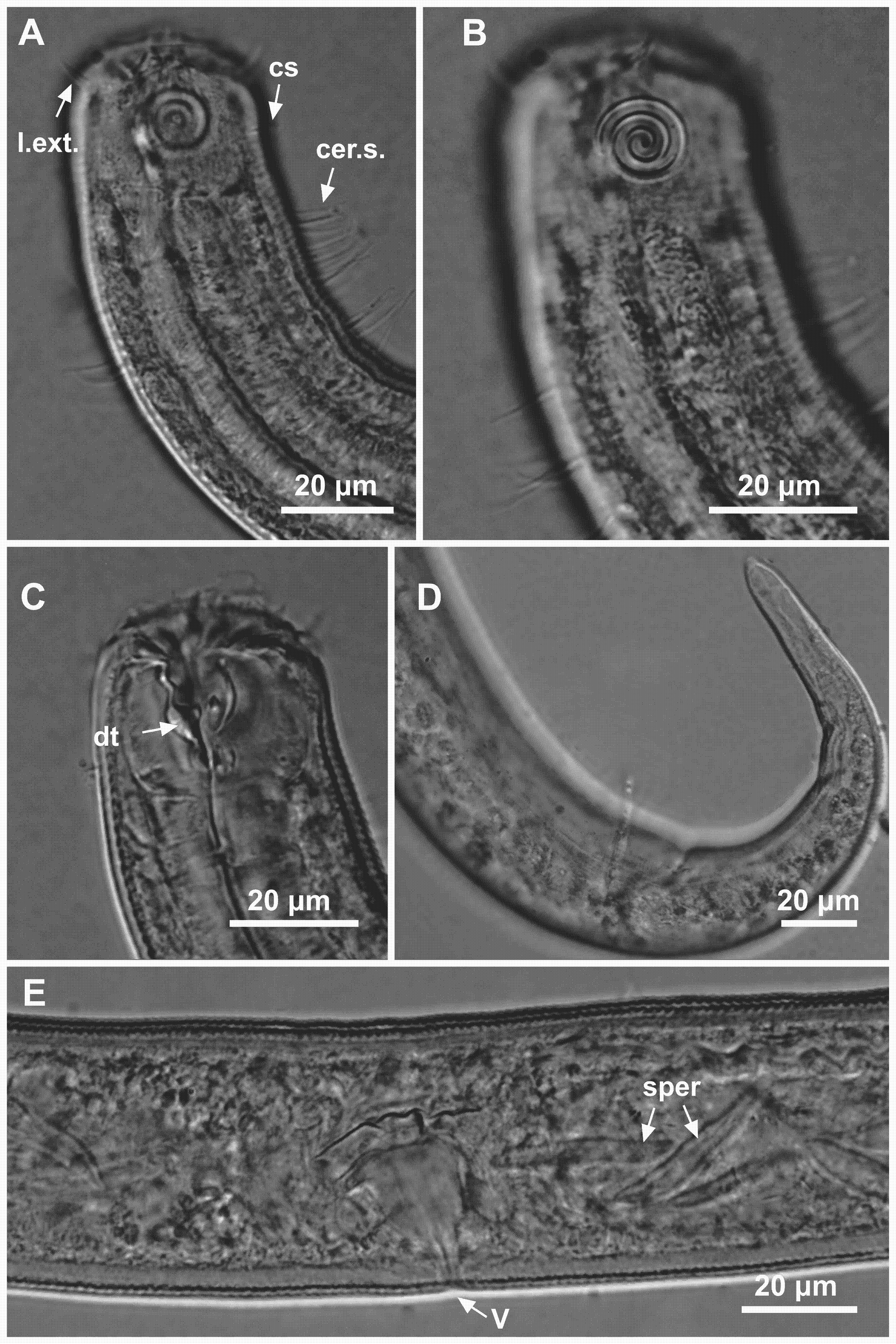{kind=link}
Material studied. Holotype male (MOUFPE 0022), paratype female (MOUFPE 0023), 1 male paratype (485 NM LMZOO-UFPE) and 2 female paratypes (486–487 NM LMZOO-UFPE).
Type locality. South Atlantic Ocean, Continental shelf off the State of Pernambuco, Brazil, (S08°16′17.10″ W34°39′34.80″), November 26, 2019, 52 m.
Locality of paratypes. Female paratypes: South Atlantic Ocean, Continental shelf off the State of Paraíba, Brazil (S07°25′15.48″ W34°29′16.56″), November 27, 2019, 47 m. Paratype male 1: South Atlantic Ocean, Continental shelf of the State of Pernambuco, Brazil (S08°16′17.10″ W34°39′34.80″), November 26, 2019, 52 m.
Etymology. Due to the location where the holotype was collected. Pernambucanus is the Latinized form of the term “pernambucano”. In Brazil, “pernambucano” refers to something or someone originating from the state of Pernambuco.
Holotype male. Body cylindrical (1,716 µm long), slightly narrow anteriorly. Maximum body diameter corresponding to 1.2 times the head diameter. Head blunt, slightly set-off. Cuticle striated from the posterior edge of the amphidial fovea. Cuticular pores not observed. Anterior sensilla arranged in the 6 + 6 + 4 pattern: six inner labial papillae (2 µm long), six outer labial setae with a broad base (7 µm long) and four slender cephalic setae about the same length as the outer labial setae. Cephalic setae corresponding to 21% of head diameter. Amphidial fovea distinctly sclerotized, multispiral, ventrally wound, about 3.5 turns, occupying 36% of corresponding body diameter and located 0.4 times the head diameter of the anterior end. Buccal cavity with a strong dorsal tooth and two ventrosublateral teeth located at the same level. Cheilostoma possesses 12 rugae. Eight rows of cervical setae starting 11 µm behind the amphid fovea. Pharynx (220 µm) with anterior bulb surrounding the buccal cavity. Basal bulb is oval and not very prominent, occupying 67% of corresponding body diameter. Cardia embedded in intestine. Nerve ring situated at 55% of the pharynx length, from anterior end. Ventral gland and secretory-excretory pore not observed. Reproductive system with two opposed and outstretched testes, both to the left of the intestine. Elongated sperm cells. Spicules sclerotized with a capitulum. Gubernaculum plate-like with dorsal apophysis. Twelve pore-like precloacal supplements present and arranged at irregular intervals. The closest supplement to the cloaca is located at 24.5 µm and the farthest at 258.5 µm. A pair of precloacal ventrosublateral setae located between the cloaca and the closest supplement to it. Three caudal glands. Tail conical (four times the cloacal body diameter) with two rows of short ventrosublateral setae.
Paratype female. Similar to male. Body length 1,752 µm, maximum diameter 46.5 µm (1.3 times the head diameter). Cuticle striated behind the posterior edge of the amphidial fovea. Anterior sensilla arrangement consisting of six inner labial papillae (2 µm long), six outer labial setae with a broad base (7 µm long) and four slender cephalic setae (6 µm long, corresponding to 17% of head diameter). Amphidial fovea sclerotized, multispiral, ventrally wound, about three turns, occupying 35% of corresponding body diameter and located 0.3 times the head diameter of the anterior end. Buccal cavity with a strong dorsal tooth and two ventrosublateral teeth. Pharynx similar to that of males. Basal bulb is oval and not very prominent (66% of corresponding body diameter). Nerve ring located at 56% of the pharynx length. Vulva situated at 55% of body length. Reproductive system didelphic-amphidelphic, with outstretched ovaries. Anterior gonad situated to right side of intestine and the posterior gonad to left side of intestine. Tail conical (131 µm long), about four times anal body diameter.
Diagnosis. Spirobolbolaimus pernambucanus sp. nov. it is characterized by its body length (1,716–1,848 µm). Cuticle striated. Head blunt, slightly set-off. Six outer labial setae with a broad base and four slender cephalic setae about the same length as the outer labial setae. Amphidial fovea sclerotized, multispiral, ventrally wound, about 3–3.5 turns, occupying 31–36% of the corresponding body diameter, located at about 0.2–0.4 times the head diameter. Eight rows of cervical setae. Buccal cavity with a strong dorsal tooth and two ventrosublateral teeth. Spicules sclerotized with a capitulum (1.5–1.7 times the cloacal body diameter) and gubernaculum plate-like with dorsal apophysis. Twelve pore-like precloacal supplements. Tail conical which corresponds to 3–4 cloacal or anal body diameter.
Differential diagnosis. Spirobolbolaimus pernambucanus sp. nov. shares the following features with S. undulatus: number of longitudinal rows of cervical setae (eight in both species), spicules length (53–55 µm in the new species and 51–55 µm in S. undulatus) and the ratio between spicules length and the cloacal body diameter (1.5–1.7 in S. pernambucanus sp. nov. and 1.3–1.5 in S. undulatus). Nevertheless, S. pernambucanus sp. nov. differs from S. undulatus in terms of body length (1,686–1,848 µm in the new species vs 2,035–2,558 µm in S. undulatus), the absence of jointed outer labial setae (vs outer labial setae jointed in S. undulatus), the presence of outer labial setae about the same length as the cephalic setae (vs outer labial setae longer than cephalic setae in S. undulatus) and the number of precloacal supplements (12 pore-like precloacal supplements in the new species vs 18–19 precloacal supplements in a series of mid-ventral elevations with pores on tops in S. undulatus). Furthermore, additional teeth are absent in S. pernambucanus sp. nov. (vs present in S. undulatus).
Spirobolbolaimus pernambucanus sp. nov. shares some features with adult specimens of S. boucherorum, such as: body length (1,686–1,848 µm in the new species vs 1,460–1,870 µm in S. boucherorum), amphidial fovea diameter (11–12 µm in S. pernambucanus sp. nov. and 10–12 µm in S. boucherorum) and de Man’s ratio c (13–16 in the new species and 12–17 in S. boucherorum). However, S. pernambucanus sp. nov. differs from S. boucherorum in terms of the number of precloacal supplements (12 in the new species vs seven in S. boucherorum), the length and shape of the gubernaculum (19–19.5 µm in S. pernambucanus vs 23–30 µm in S. boucherorum) and the presence of outer labial setae that are about the same length as the cephalic setae (vs outer labial setae longer than cephalic setae in S. boucherorum). A comparison of the main characters of all valid species of Spirobolbolaimus is presented in Table 2.
| Spirobolbolaimus bathyalis | Spirobolbolaimus boucherorum | Spirobolbolaimus pernambucanussp. nov. | Spirobolbolaimus undulatus | |
|---|---|---|---|---|
| L | 595–755 | 1,460–1,870 | 1,686–1,848 | 2,035–2,558 |
| a | 22.2–30.2 | 38.4–50 | 35–43 | 47–58 |
| b | 5–6.2 | 7.3–9.8 | 7–8 | 9–10.2 |
| c | 7.7–10.2 | 12.2–17.4 | 13–16 | 14–16.8 |
| c’ | 3.5–4.5 | 2.9–3.6 | 3–4 | 3.8–4.4 |
| Outer labial setae | non-jointed | non-jointed | non-jointed | jointed |
| cs. length/ols. length % | 25–30% | 75% | 79–100% | 53–80% |
| Amph/hd | 0.3 | 0.25–0.3 | 0.2–0.4 | 0.3 |
| Number of turns of amphids | 4 turns | 2 turns | 3–3.5 turns | 3 turns |
| amph% | 55–65% | 29–35% | 31–36% | 39–43% |
| Rows of cervical setae | 6 | 8 | 8 | 8 |
| Precloacal supplements | – | 7 pore-like | 12 pore-like | 18–19 elevations with pores on tops |
| spic/c.b.d. | 1.5 | 1.7 | 1.5–1.7 | 1.3–1.5 |
| gub/spic % | 36–42% | 43% | 35–37% | 48–56% |
Dichotomous identification key for valid species of SpirobolbolaimusSoetaert & Vincx, 1988
| 1. Six rows of cervical setae and precloacal supplements absent.……..…..…S. bathyalis |
| –Eight rows of cervical setae and precloacal supplements present……………….….. 2 |
| 2. Outer labial setae jointed and additional lateral teeth present…………... S. undulatus |
| –Outer labial setae non-jointed and additional lateral teeth absent.………………..... 3 |
| 3. Seven pore-like precloacal supplements and outer labial setae longer than cephalic setae………………..………………….............…………………….. S. boucherorum |
| –Twelve pore-like precloacal supplements and outer labial setae about the same length as the cephalic setae…………….................……….. S. pernambucanussp. nov. |
Genus IxonemaLorenzen, 1971
Diagnosis. (Emended from Tchesunov, 2014) Microlaimidae. Cuticle finely striated but can appear smooth and may be covered with sediment particles. Anterior sensilla arranged according to 6 + 6 + 4 pattern: six inner labial papillae; six outer labial papillae; and four cephalic setiform sensilla. Anterior end narrowed and elongated. Amphid fovea small circular or pocket-shaped, far posterior to the anterior end, with protruding rod-shaped corpus gelatum (not seen in some species, lost/broken or absent?). Buccal cavity small, armed with a dorsal tooth and a pair of ventrosublateral teeth (additional small teeth posterior to ventrosublateral may be present). Somatic setae stout or on peduncles, sometimes jointed, arranged in rows along the body. Males with two opposite and outstretched testicles. Precloacal supplements in the form of jointed or non-jointed setae may be present. Ventral supplements in the form of jointed setae located just behind the pharynx may be present. Gubernaculum present or absent, when present without apophyses. Female didelphic-amphidelphic, with outstretched ovaries. Tail conical. Each caudal gland with its own outlet on the tail tip.
Type species: Ixonema sordidum Lorenzen, 1971
List of valid species of IxonemaLorenzen, 1971
| Ixonema deleyiMuthumbi & Vincx, 1999 |
| Ixonema gracieleae sp. nov. |
| Ixonema powelliJensen, 1985 |
| Ixonema sordidumLorenzen, 1971 |
Description of new species
Ixonema gracieleae sp. nov.
| Ixonema gracieleae sp. nov. | Holotype male | Males paratypes (n = 3) | Females paratypes (n = 3) |
|---|---|---|---|
| Body length | 586.5 | 562.5–630 | 553.5–667.5 |
| Outer labial setae length | <2 | <2 | <2 |
| Cephalic setae length | 6 | 5–7 | 6–7 |
| Head diameter at level of the cephalic setae | 8 | 8–9 | 8–9 |
| Cephalic setae in relation to head diameter (%) | 74% | 67–82% | 71–88% |
| Distance from anterior end to amphidial fovea | 31 | 31.5–34 | 29–32.5 |
| Distance from anterior end to amphidial fovea in relation to head diameter | 4 | 3.7–4.1 | 3.2–3.9 |
| Amphidial fovea diameter (maximum width) | 3 | 3 | 2.5–3 |
| Body diameter at level of the amphidial fovea | 13 | 14 | 13–14 |
| % of the amphidial fovea diameter in relation to corresponding body diameter | 25% | 21–22% | 19%–23% |
| Pharynx length | 104 | 99–104 | 96–102 |
| Position of nerve ring from anterior end | 63 | 68–71 | 64–67 |
| Nerve ring position in relation to pharynx length (%) | 60% | 68% | 65–67% |
| Pharyngeal bulb diameter | 18 | 17.5–19 | 19–19.5 |
| Body diameter at level of the pharyngeal bulb | 26 | 24–26 | 25–26 |
| % of basal bulb diameter in relation to corresponding body diameter | 68% | 70–78% | 74–76% |
| Maximum body diameter | 32 | 31–32 | 35–36 |
| Anal or cloacal body diameter | 21 | 20 | 20–20.5 |
| Tail length | 73 | 65.5–69 | 64–67 |
| Length of spicules along arc | 39 | 33–41 | * |
| Length of spicules along cord | 33 | 30–32 | * |
| Length of gubernaculum | 11.5 | 13–14.5 | * |
| Length of gubernaculum in relation to length of spicules along arc | 29% | 35–38% | * |
| Length of spicules along arc in relation to cloacal body diameter | 1.9 | 1.7–2 | * |
| Distance from anterior end to vulva | * | * | 261–411 |
| Position of vulva from anterior end (%) | * | * | 47%–62% |
| Body diameter in vulva region | * | * | 35–36 |
| Anterior ovary length | * | * | 52–69 |
| Posterior ovary length | * | * | 42–69 |
| Reproductive system length | 431 | 425–479.5 | 105–138 |
| % of reproductive system in relation to body length | 73% | 67%–82% | 16–25% |
| a | 18 | 18–20 | 15–19 |
| b | 6 | 6 | 5–7 |
| c | 8 | 9 | 8–10 |
| c’ | 3.5 | 3–3.5 | 3 |
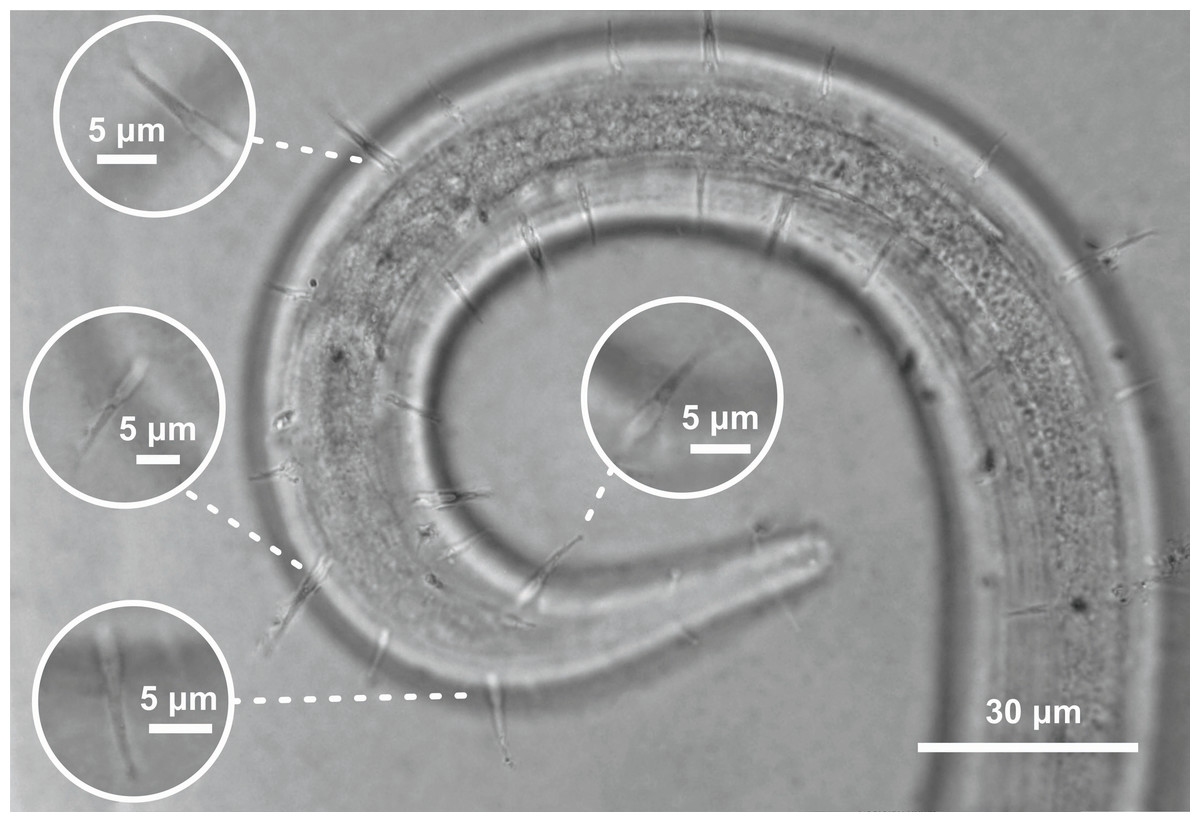
Figure 5: Ixonema gracieleae sp. nov. holotype male.
Setae on peduncles distributed along the body. Image source credit: Alex Manoel.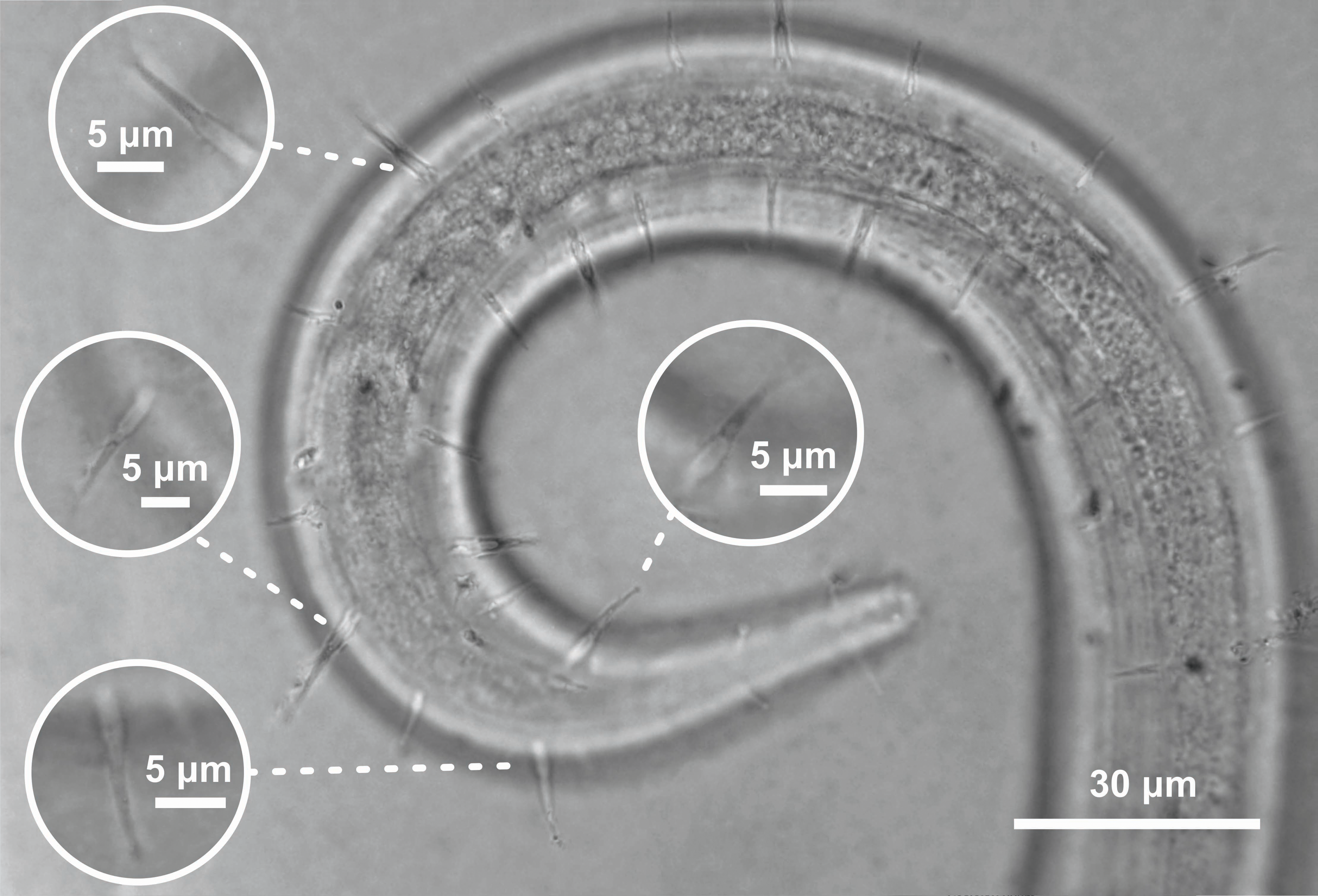{kind=link}
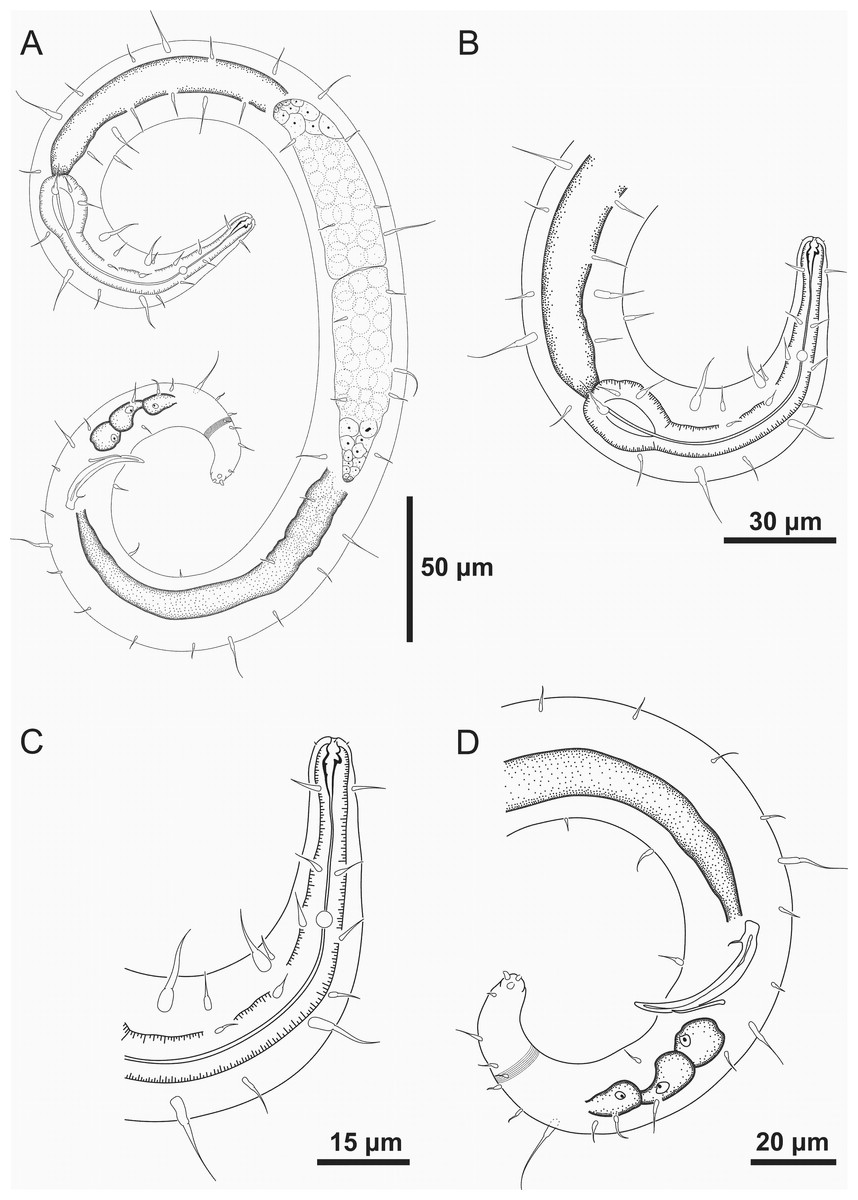
Figure 6: Ixonema gracieleae sp. nov. holotype male.
Holotype male: (A) overview; (B) anterior region; (C) anterior end; (D) posterior region. Image source credit: Alex Manoel.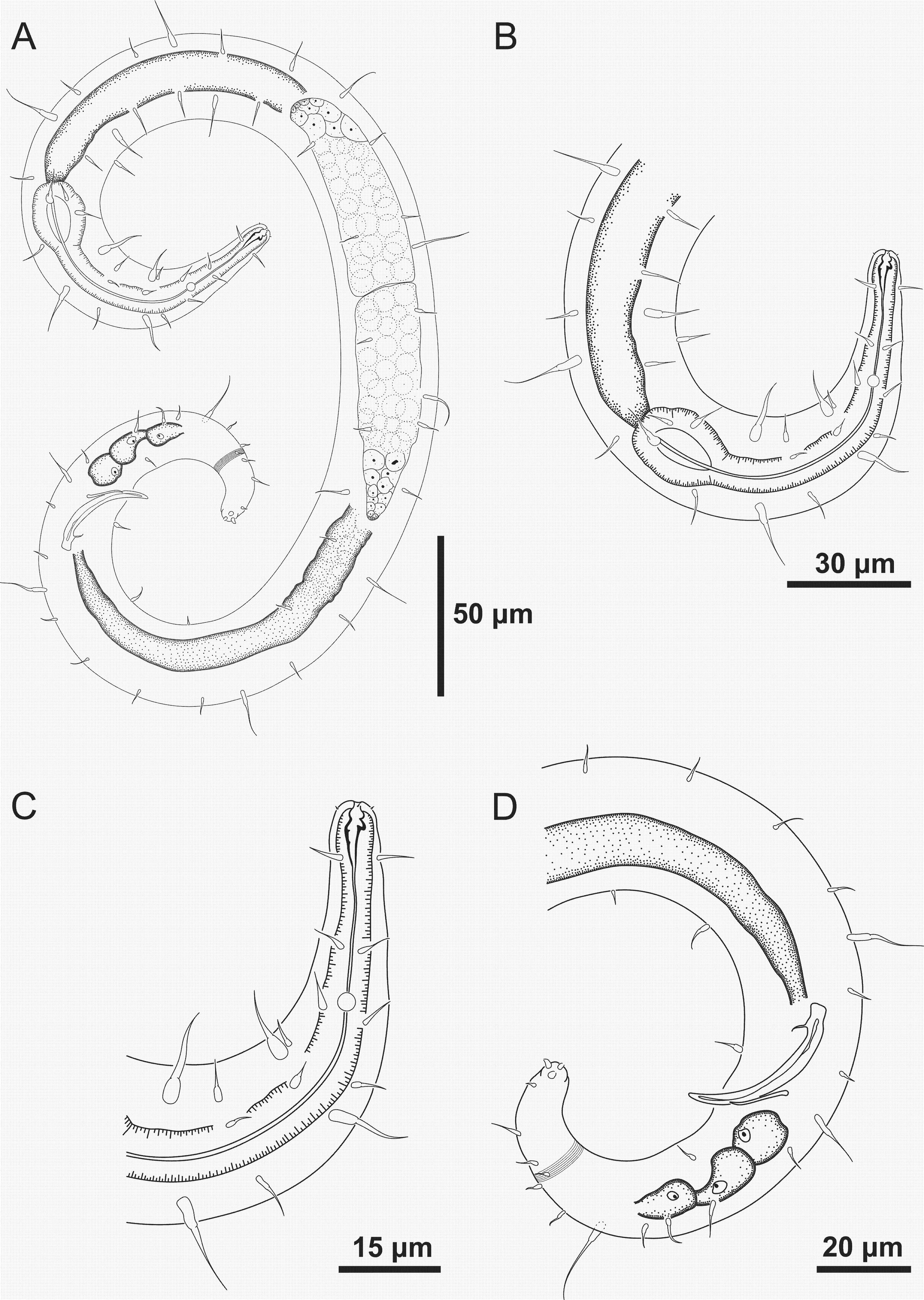{kind=link}
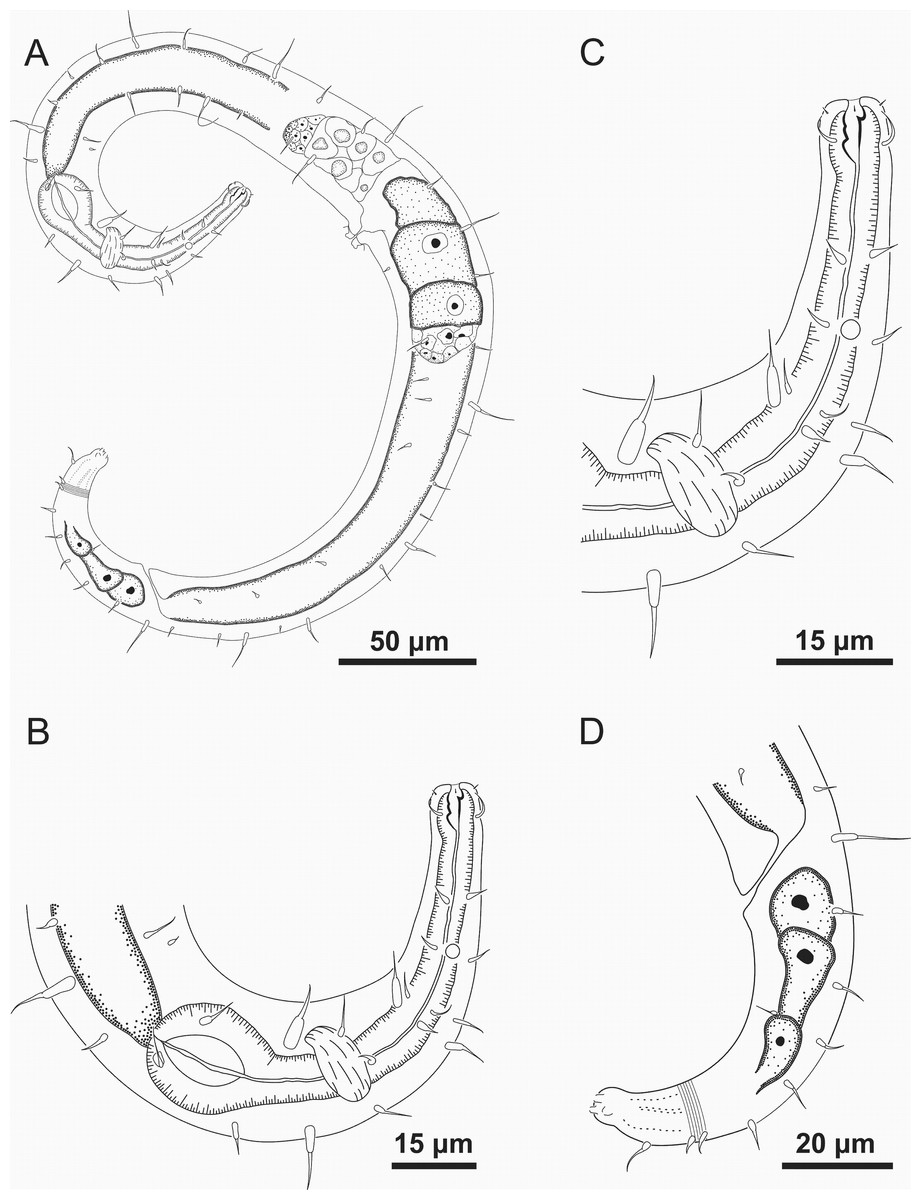
Figure 7: Ixonema gracieleae sp. nov.: paratype female.
Paratype female: (A) overview; (B) anterior region; (C) anterior end; (D) tail. Image source credit: Alex Manoel.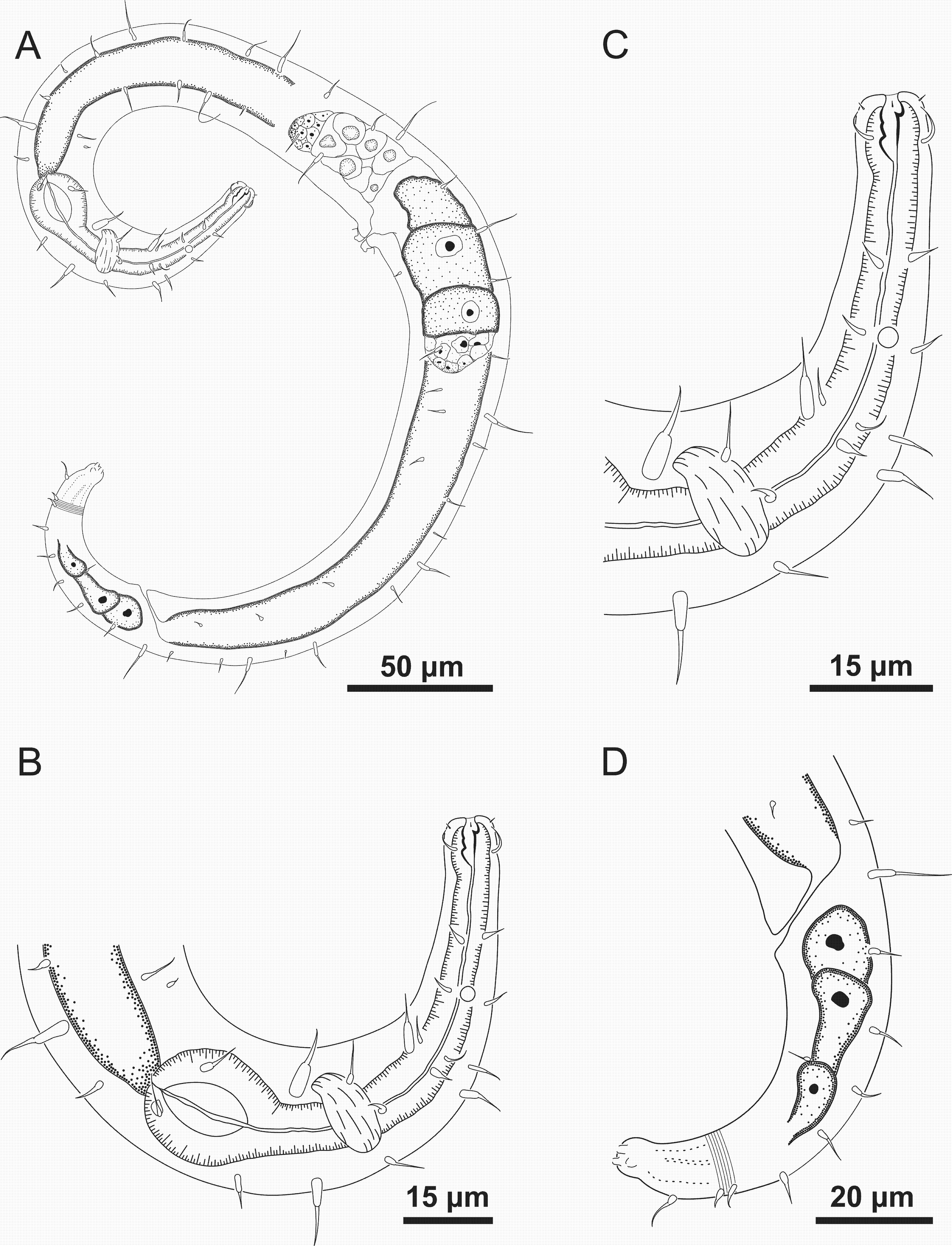{kind=link}
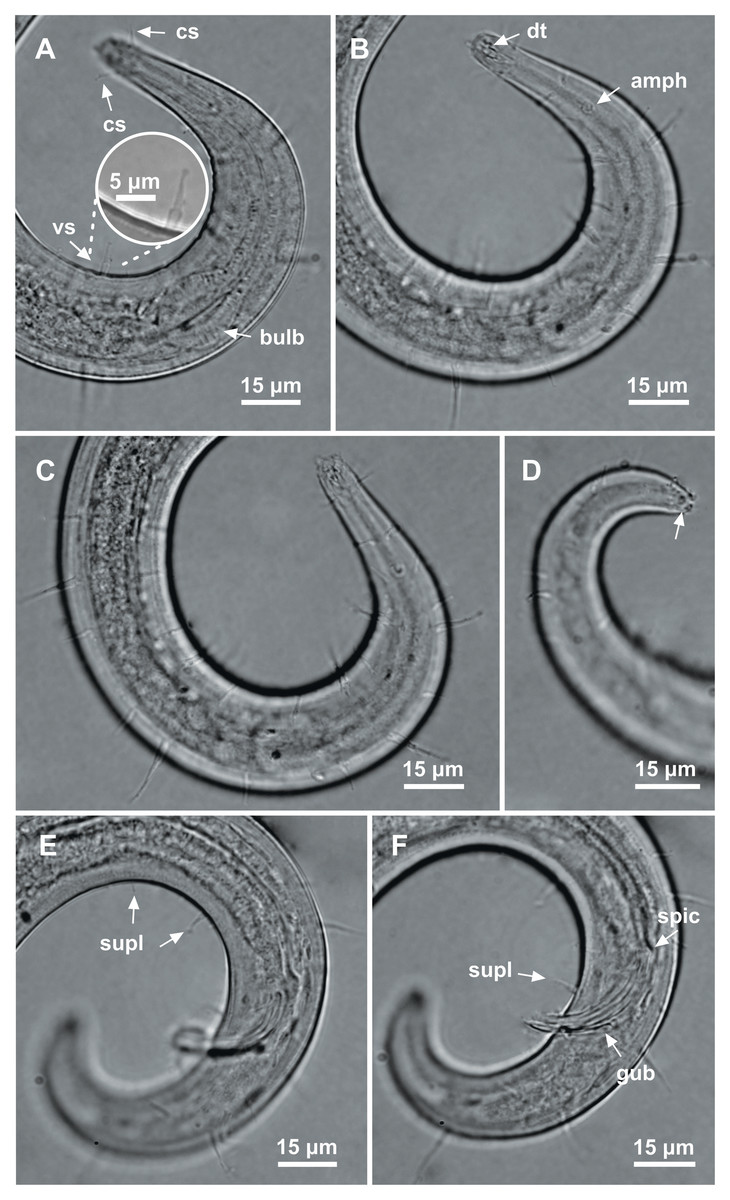
Figure 8: Ixonema gracieleae sp. nov. holotype male.
Holotype male: (A) anterior region: arrows indicating cephalic setae (cs), a single ventral seta in the anterior region (vs) and pharyngeal bulb (bulb); (B) anterior region: arrows indicating the dorsal tooth (dt) and amphidial fovea (amph); (C) anterior end (somatic setae); (D) tail (arrow indicating three separate tail tip outlets); (E) posterior end: arrows indicating precloacal supplements (supl); (F) posterior end: arrows indicating precloacal supplement (supl), spicule (spic) and gubernaculum (gub). Image source credit: Alex Manoel.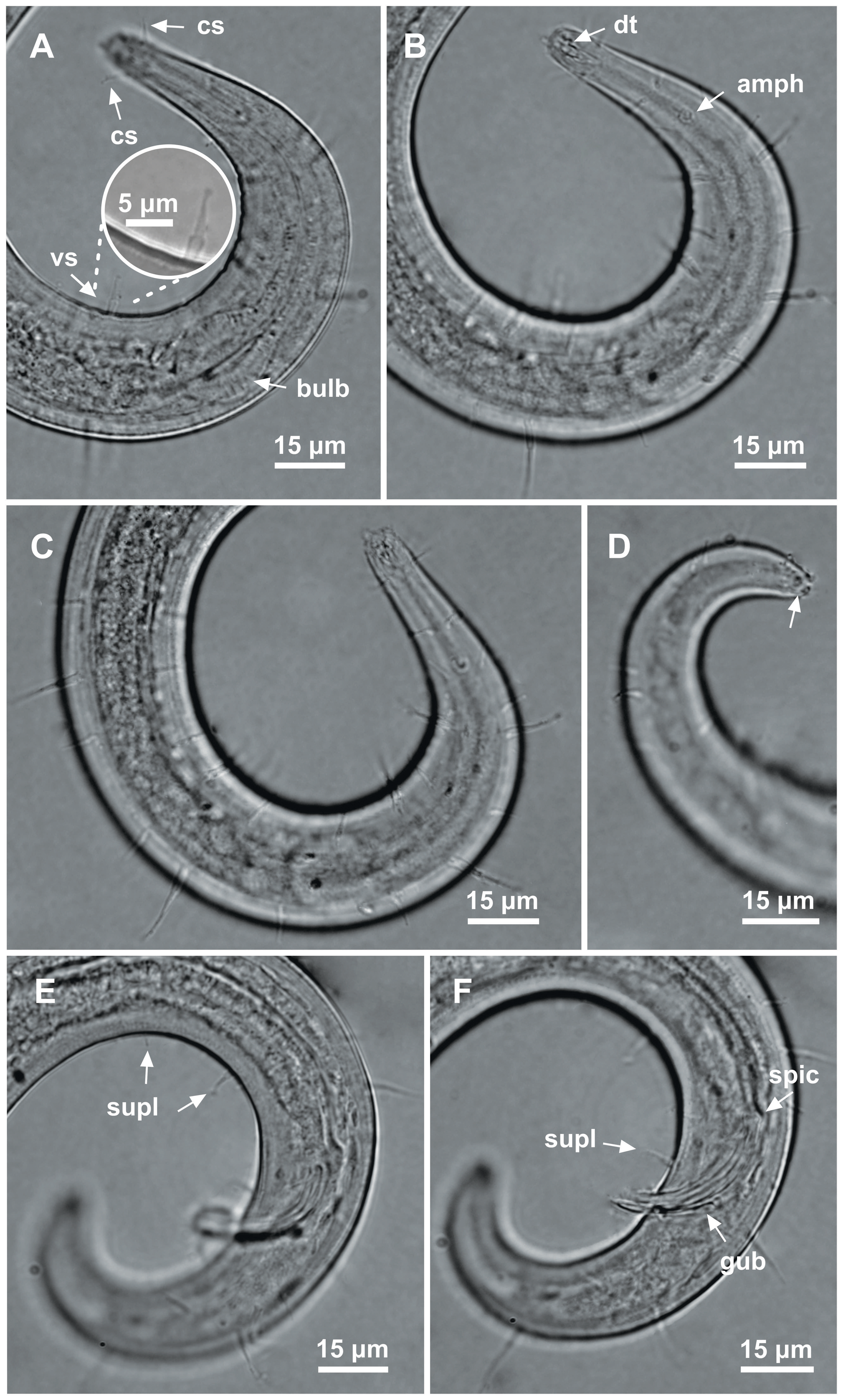{kind=link}
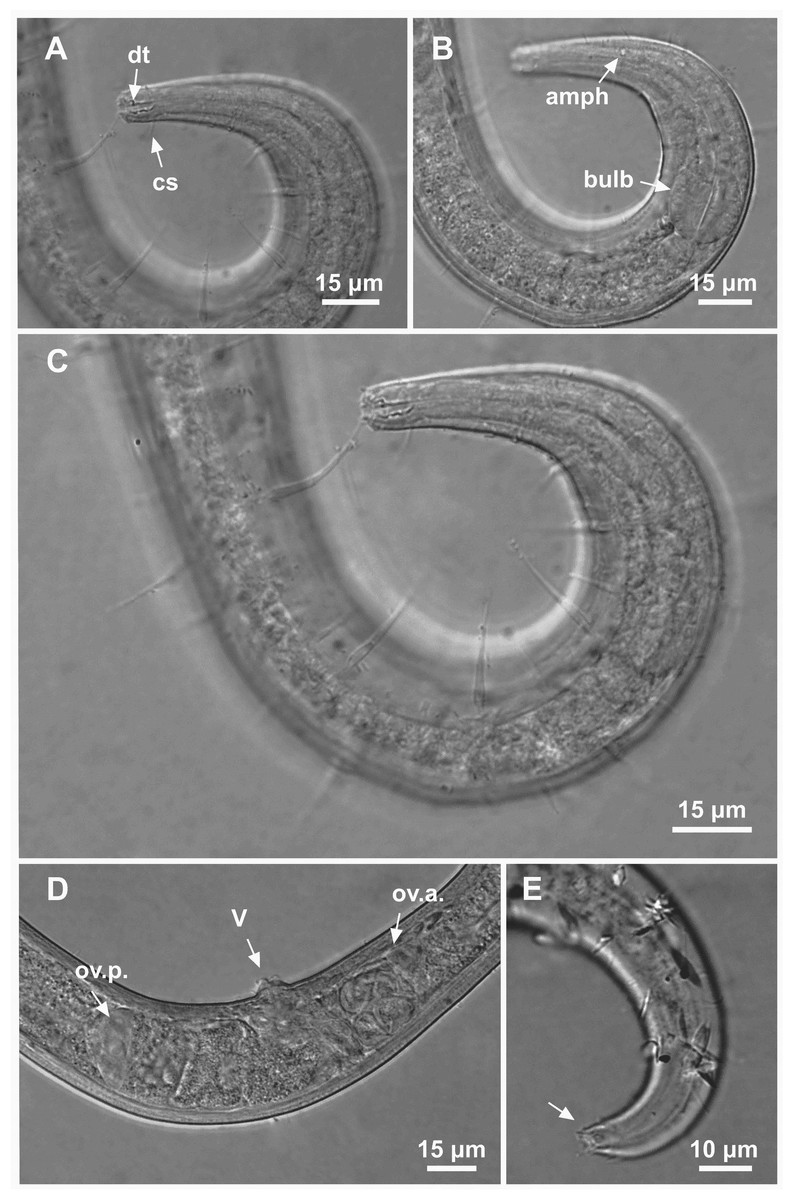
Figure 9: Ixonema gracieleae sp. nov. paratype female 1 and paratype female 2.
Paratype female 1: (A) anterior region: arrows indicating cephalic setae (cs) and the dorsal tooth (dt); (B) anterior region: arrows indicating amphidial fovea (amph) and pharyngeal bulb (bulb); (C) anterior end (somatic setae); (D) reproductive system: arrows indicating the anterior ovary (ov.a.), posterior ovary (ov.p.) and the vulva (V). Paratype female 2: (E) tail (arrow indicating three separate tail tip outlets). Image source credit: Alex Manoel.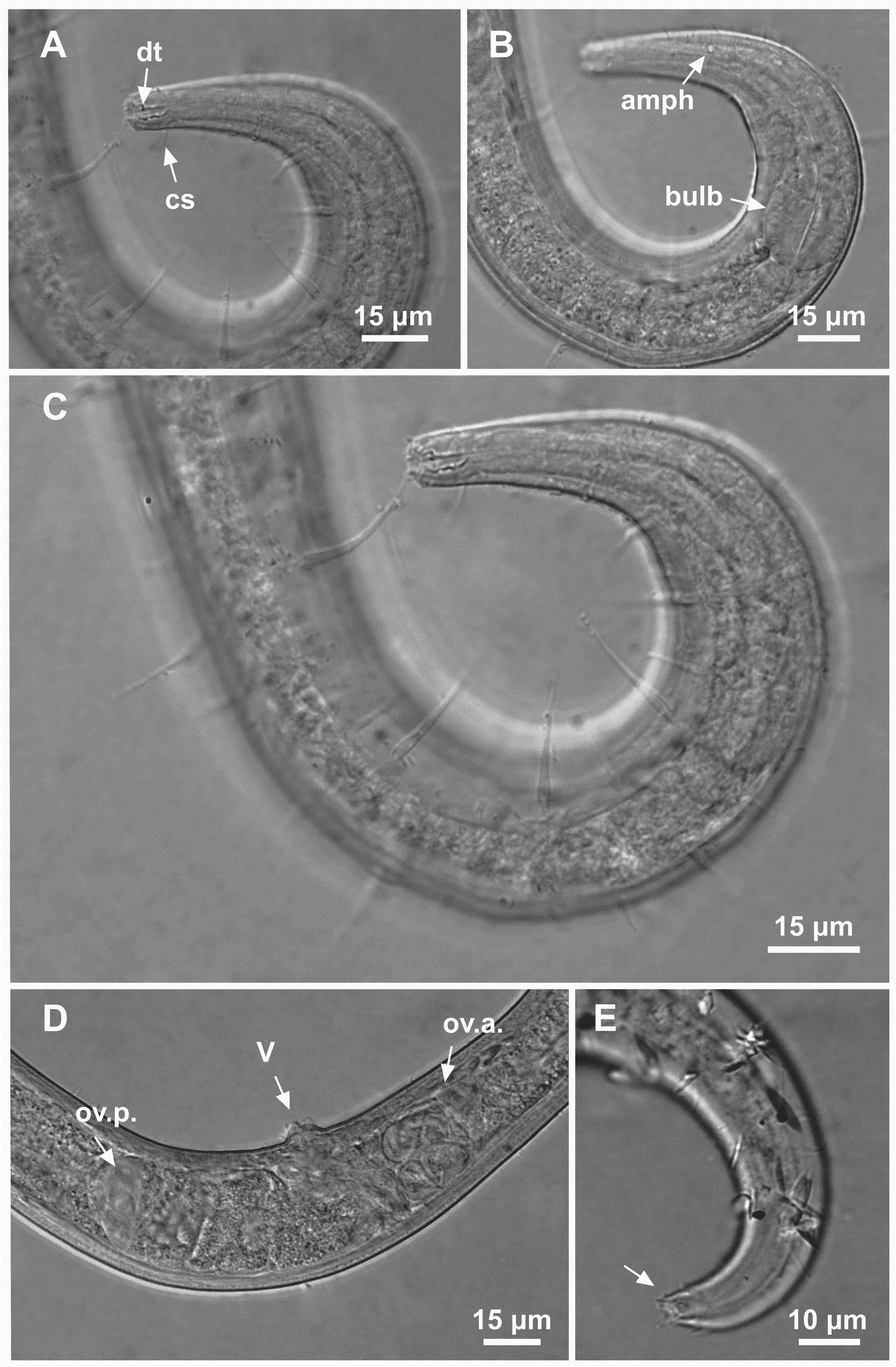{kind=link}
Material studied. Holotype male (MOUFPE 0024), paratype female (MOUFPE 0025), 3 male paratypes (488–490 NM LMZOO-UFPE) and 2 female paratypes (491–492 NM LMZOO-UFPE).
Type locality. South Atlantic Ocean, Continental shelf off the State of Alagoas (S09°39′14.52″ W35°15′21.66″), November 25, 2019, 50 m. The paratypes were found in the same locality.
Etymology. The specific epithet is a tribute to Graciele Mariza dos Santos Alves, wife of the first author.
Holotype male. Body cylindrical, short and plump, attenuating on both ends and thick in the middle (586.5 µm long). Maximum body diameter corresponding to 4.1 times the head diameter. Cuticle finely striated, the striations are so delicate that they are difficult to visualize (striations are most visible on the tail). Anterior sensilla arranged in the 6 + 6 + 4 pattern: six inner labial papillae, six outer labial papillae and four cephalic setae (6 µm long). Cephalic setae corresponding to 74% of head diameter. Amphidial fovea small and circular, occupying 25% of corresponding body diameter and located far posterior to the anterior end (four times the head diameter of the anterior end). Protruding Corpus gelatum not observed. Buccal cavity with a dorsal tooth and two small ventrosublateral teeth. Cheilostoma rugae indiscernible under a light microscope. Four sublateral rows of stout cervical setae located between the posterior region of the cephalic setae and the base of the amphidial fovea. The first row is 24 µm from the anterior end and the second is at the same height of the amphidial fovea base, 31 µm from the anterior end. Six rows of cervical setae on peduncles located between the posterior region of the amphidial fovea and close to the bulb base: four sublateral rows where relatively smaller setae (4–7 µm) and long setae (10–16 µm) alternate; two lateral rows (about 4 µm). The first row is about 11 µm from the base of the amphidial fovea. After the cervical region, the somatic setae are distributed in two rows where the alternation between two smaller setae (about 5–9 µm) and a larger one (12–19 µm) usually occurs. Pharynx (104 µm) with prominent terminal bulb, occupying 73% of corresponding body diameter. Cardia embedded in intestine. Nerve ring situated at 60% of the pharynx length, from anterior end. Ventral gland and secretory-excretory pore not observed. Reproductive system with two opposed and outstretched testes on right side of intestine. Spicules curved (39 µm), about two times the cloacal body diameter. Gubernaculum slender and without apophysis. Four setiform ventral supplements: three precloacal supplements plus one located just behind the pharynx. The two setae closest to the cloaca (10.5 µm and 40 µm anterior to the cloaca, respectively) are jointed, measuring 7 µm and 6 µm respectively; the third precloacal seta is smaller (about 2 µm length), apparently non-jointed in light microscopy and far from the cloaca (62 µm). The setae located just behind the pharynx (about 7 µm length) are jointed (morphologically similar to the two setae closest to the cloaca) and are located 84 µm from the anterior end. Tail conical (73 µm long), with a blunt tip where the three caudal glands open through separate outlets into papilla-like extensions.
Paratype female. Similar to male. Body measuring 667.5 µm in length, maximum diameter 36 µm (4.3 times the head diameter). Cuticle finely striated, the striations are so delicate that they are difficult to visualize. Anterior sensilla arranged in the 6 + 6 + 4 pattern: six inner labial papillae, six outer labial papillae and four cephalic setae (7 µm long, corresponding to 88% of head diameter). Amphidial fovea small and circular, occupying 21% of corresponding body diameter and located far posterior to the anterior end (3.9 times the head diameter of the anterior end). Protruding corpus gelatum not observed. Morphologies and distribution patterns of cervical and somatic setae are similar to the male. Jointed ventral seta located just behind the pharynx absent. Buccal cavity, teeth and pharynx similar to that of male. Bulb occupying 74% of corresponding body diameter. Nerve ring situated at 67% of the pharynx length, from anterior end. Vulva located 411 µm from anterior end, at 62% of body length. A pair of ventral papillae surround the vulva. Reproductive system didelphic-amphidelphic, with outstretched ovaries. Anterior and posterior ovary to right side of intestine. Tail conical (67 µm long), similar to the male.
Diagnosis. Ixonema gracieleae sp. nov. it is characterized by its finely striated cuticle. Amphidial fovea small and circular located far posterior to the anterior end. Protruding corpus gelatum not observed. Rows of cervical and somatic setae on peduncles along the body. Reproductive system with two opposed and outstretched testes in the males and didelphic-amphidelphic, with outstretched ovaries in the females. Spicules curved and gubernaculum without apophysis. Four setiform ventral supplements: three precloacal setae (the two closest to the cloaca are jointed and the furthest apparently non-jointed) and a jointed ventral seta located just behind the pharynx. Tail conical with a blunt tip where the three caudal glands open through separate outlets into papilla-like extensions.
Differential diagnosis. Ixonema gracieleae sp. nov. resembles the monospecific genus Bathynox (Bussau, 1993) Bussau & Vopel, 1999 due to the presence of the setae on peduncles distributed along the whole body. However, I. gracieleae sp. nov. differs from Bathynox in having three caudal glands opening through separate outlets in papilla-like extensions (vs three glands opening in a single outlet in Bathynox). Furthermore, the male reproductive system is different between the genera: male diorchic in I. gracieleae sp. nov. vs male monorchic in Bathynox and the gubernaculum lacks apophysis in I. gracieleae sp. nov. vs the presence of dorsal apophysis in Bathynox.
The presence of setae on peduncles in I. gracieleae sp. nov. differentiates it from other species of the genus. The new species shares the presence of a single jointed ventral seta located just behind the pharynx with I. powelli, as well as jointed precloacal setae in the males. Nonetheless, I. gracieleae sp. nov. differs from I. powelli in terms of the shape of the amphidial fovea (circular in I. gracieleae sp. nov. vs pocket-like in I. powelli), the distribution pattern and number of rows of cervical setae (6 rows of cervical setae in I. gracieleae sp. nov. vs 4 rows of cervical setae I. powelli) and gubernaculum length (11.5–14.5 µm in I. gracieleae sp. nov. vs about 6 µm in I. powelli).
Ixonema gracieleae sp. nov. differs from I. sordidum in having long somatic setae alternating with short somatic setae along the body and due to the presence of precloacal supplements, these characteristics are absent in the mentioned species. I. gracieleae sp. nov. differs from I. deleyi in terms of the presence of the gubernaculum (vs absent in I. deleyi) and the number and morphology of supplements (jointed ventral seta located just behind the pharynx + two jointed setae closer to the cloaca and a smaller non-jointed seta further away in I. gracieleae sp. nov. vs a single non-jointed ventral precloacal seta in I. deleyi). A comparison of the main characters of all valid species of Ixonema is presented in Table 4.
| Ixonema deleyi | Ixonema gracieleae sp. nov. | Ixonema powelli | Ixonema sordidum | |
|---|---|---|---|---|
| L | 245–405 | 553.5–667.5 | 640–900 | 530 |
| a | 12.3–22.5 | 15–19 | 13–23 | 16–20 |
| b | 3.6–4.9 | 5–7 | 5.9 –7 | 5.1–5.3 |
| c | 7.3–9.2 | 8–10 | 7.3–10.6 | 8.2–8.8 |
| c’ | 2.5–4.5 | 3–3.5 | 3.5 | 3 |
| Amphidial fovea | circular (rod-like cg.) |
circular (cg. not seen or absent) |
pocket-shaped (cg. not seen or absent) |
circular (rod-like cg.) |
| Amph/hd | 4.8–6.2 | 3.2–4 | 4–5 | 3.6–3.8 |
| Somatic setae | stout | on peduncles | jointed | stout |
| Supplements | 1 non-jointed precloacal setae |
4 jointed setae* (1 just behind pharynx +3 precloacal) |
3 jointed setae (1 just behind pharynx +2 precloacal) |
- |
| spic/c.b.d. | 2 | 1.7–2 | 1.4–1.8 | 1.4 |
| Gubernaculum | - | + | + | + |
| gub/spic % | x | 29–38% | 14% | 44% |
Notes:
Dichotomous identification key for valid species of IxonemaLorenzen, 1971
| 1. Body length less than 500 µm and gubernaculum absent…………………..…I. deleyi |
| –Body length longer than 500 µm and gubernaculum present…………………………2 |
| 2. Somatic setae stout and precloacal supplements absent…………………... I. sordidum |
| –Somatic setae on peduncles or jointed and precloacal supplements present………... 3 |
| 3. Amphidial fovea circular and somatic setae on peduncles ... I. gracieleaesp. nov. |
| –Amphidial fovea pocket-shaped and somatic setae jointed …………...…..... I. powelli |
Discussion
Although there are no records of species originally described for the South Atlantic, the occurrence of Spirobolbolaimus and Ixonema was previously reported for this region. Spirobolbolaimus was found in the sublittoral of Pedra do Xaréu Beach, Pernambuco, Northeastern Brazil (Rocha et al., 2006). This taxon was also found in sediment samples from the Grussaí canyon and a point adjacent to it in the Campos Basin, Southeastern Brazil (Silva, 2012). Ixonema was identified in samples from the Campos Basin Slope, Southeastern Brazil (Moura, 2013).
Spirobolbolaimus pernambucanus sp. nov. is the first species of the genus described for the South Atlantic. In all previously described species, the outer labial setae are longer than the cephalic setae. However, the outer labial setae of S. pernambucanus sp. nov. are similar in length to the cephalic setae. We included this feature in the diagnosis of the genus. Based on the described species, we added the following characteristics to the diagnosis of the genus: the morphology of the buccal cavity and teeth that usually occur in species; the types of precloacal supplements; sperm cells and tail shape; occurrence of jointed outer labial setae. Jointed labial setae, as seen in S. undulatus, may be present in more genera or species than mentioned in the descriptions, since in some cases this characteristic may have been overlooked (Lorenzen, 1994).
Ixonema gracieleae sp. nov. can be easily confused and classified as belonging to Bathynox, mainly due to the presence of rows setae on peduncles distributed along the body. However, I. gracieleae sp. nov. presents a combination of differential characteristics that typically only occur in representatives of Ixonema: three caudal glands open through separate outlets into papilla-like extensions, two opposite and outstretched testes and gubernaculum without apophysis (Lorenzen, 1971; Jensen, 1985; Muthumbi & Vincx, 1999). Therefore, the presence of such characteristics invalidates the hypothesis of including this species in another genus and reinforces its taxonomic position. In the Microlaimidae family, the presence of caudal glands with independent outlets is unique for Ixonema, and this feature is considered phylogenetically primitive (Lorenzen, 1971). Ixonema gracieleae sp. nov. is the first species of the genus described from the South Atlantic. The description of this new species strongly contributes to the knowledge and the variability of this genus.
Among the three valid species of Ixonema, it was possible to observe variations in some important characteristics for the identification of the genus that were absent in the last diagnosis of the genus provided by Tchesunov (2014). The cuticle of the species I. sordidum and I. powelli was described as smooth (Lorenzen, 1971; Jensen, 1985). In both species, the cuticle was covered by a thin layer of particles, except in a I. powelli male, where it was possible to visualize subcuticular striae in the most anterior region of the pharynx. Electron microscopy analysis allowed the description of a very finely striated cuticle for I. deleyi (Muthumbi & Vincx, 1999). Tchesunov, Jeong & Lee (2021) included this variation in a comparative table between the genera of Microlaimidae but did not provide a complete diagnosis of Ixonema. In the new species, the cuticle is similar to that described for I. deleyi, and it is possible to visualize very fine striations on the tail of the analyzed specimens, even using optical microscopy. Therefore, the cuticle of Ixonema is very finely striated, but can also appear smooth, especially when using light microscopy.
The amphidial fovea can vary from circular (I. sordidum, I. deleyi and I. gracieleae sp. nov.) to pocket-like (I. powelli). A protruding rod-shaped corpus gelatum is present in I. sordidum and I. deleyi. This characteristic was not observed in I. powelli and I. gracieleae sp. nov. When establishing the genus, Lorenzen (1971) argued that the walls of the gelatinous rods do not appear to be delicate structures. However, although I. deleyi presents a protruding corpus gelatum, in the electron microscopy analyzes provided in the original description of this species, it is possible to observe that this structure seems to have been lost during specimen preparation (see in Muthumbi & Vincx, 1999-figure 7). This observation is not in line with Lorenzen’s assumption (non-delicate structures). Therefore, we do not know for sure if this structure is absent in I. powelli and I. gracieleae sp. nov. or if it was lost/broken during sample processing and organism preparation, which commonly occurs with other structures such as the tail and setae.
The gubernaculum is present and lacks apophyses in the species I. gracieleae sp. nov., I. sordidum and I. powelli. However, this structure is absent in I. deleyi. Supplements in the form of jointed setae in a ventral position are present just behind the pharynx and in front of the cloaca in I. powelli and I. gracieleae sp. nov. A non-jointed ventral seta located anterior to the cloacal opening is present in I. deleyi. Due to the similarity with I. powelli and I. garacielea sp. nov. in relation to the position at which the seta is located, we will consider it as a precloacal supplement. Jointed somatic setae may be present (I. powelli). The occurrence of somatic setae on peduncles described for the new species is unprecedented for the genus. The variation of the characters discussed above, in addition to the new features found in the new species, were included in the diagnosis of the genus.
The present study increases the number of Microlaimidae species originally described from sediment samples collected in the Brazilian coast. These results demonstrate that a great effort is still required in order to fully understand the real richness of the Microlaimidae assemblage present in marine sediments of continental margins, such as the Brazilian coast.