
Genome-wide identification, molecular characterization, and gene expression analyses of honeysuckle NHX antiporters suggest their involvement in salt stress adaptation
- Published
- Accepted
- Received
- Academic Editor
- Vladimir Uversky
- Subject Areas
- Agricultural Science, Bioinformatics, Genomics, Plant Science
- Keywords
- Honeysuckle, Genome-wide, Na+/H+ antiporter (NHX), Salt stress
- Copyright
- © 2022 Huang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2022. Genome-wide identification, molecular characterization, and gene expression analyses of honeysuckle NHX antiporters suggest their involvement in salt stress adaptation. PeerJ 10:e13214 https://doi.org/10.7717/peerj.13214
Abstract
Background
Ion homeostasis is an essential process for the survival of plants under salt stress. Na+/H+ antiporters (NHXs) are secondary ion transporters that regulate Na+ compartmentalization or efflux reduce Na+ toxicity and play a critical role during plant development and stress responses.
Methods and Results
To gain insight into the functional divergence of NHX genes in honeysuckle, a total of seven LjNHX genes were identified on the whole genome level and were renamed according to their chromosomal positions. All LjNHXs possessed the Na+/H+ exchanger domain and the amiloride-binding site was presented in all NHX proteins except LjNHX4. The phylogenetic analysis divided the seven NHX genes into Vac-clade (LjNHX1/2/3/4/5/7) and PM-clade (LjNHX6) based on their subcellular localization and validated by the distribution of conserved protein motifs and exon/intron organization analysis. The protein-protein interaction network showed that LjNHX4/5/6/7 shared the same putatively interactive proteins, including SOS2, SOS3, HKT1, and AVP1. Cis-acting elements and gene ontology (GO) analysis suggested that most LjNHXs involve in the response to salt stress through ion transmembrane transport. The expression profile analysis revealed that the expression levels of LjNHX3/7 were remarkably affected by salinity. These results suggested that LjNHXs play significant roles in honeysuckle development and response to salt stresses.
Conclusions
The theoretical foundation was established in the present study for the further functional characterization of the NHX gene family in honeysuckle.
Introduction
Soil salinization is one of the major environmental stress that reduces plant growth and productivity throughout the world (Huang et al., 2019a; Nedjimi, 2016; Rengasamy, 2017). When plants are exposed to a high-salt environment, Na+ will enter the cells through the plasma-membrane non-selective ion channels (NSCCs) and high-affinity K+ transporter-1 (HKT1) protein, resulting in ionic toxicity and osmotic stress (Chen et al., 2015; Deinlein et al., 2014; Dong et al., 2021). The regulation of osmosis and ion homeostasis of cells under salt stress, especially the compartmentalizing and excluding ability of Na+, mainly depends on the activity of ion transporters and channels. In particular, the monovalent cation/proton antiporters (CPA) superfamily is one of the most important families in plant responses to salt stress, including Na+/H+ antiporters (NHXs), K+-efflux antiporters (KEAs), and cation/H+ exchangers (CHXs) and has been well characterized (Jia et al., 2018). The NHX family which belongs to the CPA1 family is expected to have 10–12 transmembrane domains while KEAs and CHXs are predicted to have 8–14 transmembrane domains, all of which contain the Na+/H+ exchanger domain (PF00999) (Wang et al., 2020b). These proteins function in regulating cation and pH homeostasis by exchanging Na+, K+, Li+ for H+ in plants, animals, fungi, and bacteria, and are mainly localized within the vacuole, plasma, and organelle membranes (Bassil et al., 2019; Sharma, Taneja & Upadhyay, 2020).
It was reported that the NHX family is conserved across various evolutionary lineages, which indicates that they play a vital role in the development of organisms (Sharma, Taneja & Upadhyay, 2020). NHXs participate in various biological processes such as salt tolerance, pH and ion balance regulation, cell expansion, stomatal function, cellular vesicle trafficking, protein processing, and flower development (Tian et al., 2017; Zhou et al., 2016). The eight NHX genes from Arabidopsis were divided into vacuole (Vac) -clade (AtNHX1, AtNHX2, AtNHX3, and AtNHX4), endosomal (Endo) -clade (AtNHX5, AtNHX6), and plasma membrane (PM) -clade (AtNHX7/ SOS1, AtNHX8), respectively, according to their subcellular location (Cui et al., 2020). Phylogenetic analysis showed that BvNHXs (Beta vulgaris) and GbNHXs (Gossypium barbadense) also clustered into three subclades, among which Vac-clade NHXs are the most abundant from all the studied species (Akram et al., 2020; Wu, Wang & Li, 2019). The function of NHX transporters can be preliminarily defined by their subcellular localization (Akram et al., 2020). For example, Vac-binding AtNHX1 and AtNHX2 function in controlling vacuolar K+ and pH homeostasis to regulate cell expansion, stomatal conductance, and floral organ development (Bassil et al., 2011b). Endomembrane-bounded AtNHX5 and AtNHX6 are involved in maintaining organelle pH and ion homeostasis with implications in endosomal sorting and cellular stress responses (Bassil et al., 2011a). AtNHX8 is located on the PM and has been proved to be a Li+/H+ antiporter (Meng & Wu, 2018; Wang et al., 2013).
Many studies have provided convincing evidence for the involvement of the NHX gene in salt tolerance. In Arabidopsis, AtNHX1-6 transport either Na+ or K+ into the vacuole or endosome in exchange for H+ efflux to the cytoplasm to maintain the cellular ion homeostasis (Barragan et al., 2012; Bassil & Blumwald, 2014; Bassil et al., 2011a). Different studies have shown that plants have stronger salt stress tolerance when AtNHX1, AtNHX2, AtNHX3, AtNHX5, or AtNHX6 were overexpressed, or AtNHX4 was deficient (Jia et al., 2018; Li et al., 2009; Liu et al., 2008; Wu et al., 2016); the nhx5 nhx6 double-knockout mutant aborted the transport through the tonoplast, increasing the sensitivity to salt stress (Bassil et al., 2011a). PM-binding AtNHX7/ SOS1 endowed plants with salt tolerance through the Ca2+-dependent SOS (Salt overly sensitivity) pathway, which is the core mechanism of plant salt tolerance (Dong et al., 2021; Ji et al., 2013). The SOS signaling pathway consists of SOS3, SOS2, and SOS1. Salt stress triggers cytosolic Ca2+ elevation that activates SOS3. SOS3 recruit SOS2 to the PM and activate its kinase activity. The SOS3-SOS2 complex targets the PM-localized Na+/H+ exchanger SOS1 to regulate ion transport processes at the PM (Deinlein et al., 2014; Ma et al., 2019). AtNHX7 also plays a role in the long-term transport of Na+ (Hamam et al., 2016; Ji et al., 2013). In Arabidopsis, Qiu et al. found that plasma membrane Na+/H+ exchange activity was reduced by 80% in the Atsos1 mutants compared to the control (Jia et al., 2018; Qiu et al., 2002). Overexpression of AtSOS1 increased salt tolerance in transgenic Arabidopsis by reducing Na+ content in xylem and shoot (Yang et al., 2009).
Honeysuckle (Lonicera japonica Thunb.) belongs to the Caprifoliaceae family, is native to eastern Asia, and is now grown in many countries, such as Australia and the United States (Cai et al., 2021b; Pu et al., 2020). Its dried flower buds, leaves, and stems have been prescribed in traditional Chinese medicine (TCM) to treat fever, influenza, sores, and swelling for more than 1,500 years. Moreover, the benefits of honeysuckle have been demonstrated in the treatment of many emerging diseases such as influenza A viruses (H1N1, H5N1, H7N9), SARS coronavirus, hand-foot-and-mouth disease, and the early-stage novel coronavirus infection (Liu et al., 2005; Pu et al., 2020; Shang et al., 2011). It is believed to be an ecologically invasive species in several countries including New Zealand, Australia, Argentina, Mexico, and much of the USA because of its high environmental adaptability (Uddin et al., 2021). Previous studies have found that honeysuckle is highly resistant to salt stress, but the physiological and molecular mechanisms remain unclear (Cai et al., 2021b; Huang et al., 2019b). The analysis of LjNHXs in honeysuckle would enable a more comprehensive understanding of molecular mechanisms underlying Na+ homeostasis and plant salt stress resistance.
In this study, seven NHX genes were identified from the genome of Lonicera japonica and classified into Vac-clade and PM-clade based on their subcellular localization. The physicochemical properties, phylogenetic relationship, architecture of conserved motifs, gene structures, cis-acting elements of LjNHX genes, the distribution of LjNHX genes on chromosomes, protein tertiary structure, and putative protein-protein interaction (PPI) were comprehensively analyzed. Furthermore, we investigated the expression pattern of LjNHX family genes in the Lonicera japonica cultivar “Huajin 6” in different tissues and gradient salt stress. Our results could serve as a theoretical reference for an in-depth analysis of the response mechanisms of LjNHX family genes to salt stress and their mediation of plant resistance/tolerance to high salinity.
Materials and Methods
Identification of NHX genes in honeysuckle genome
The published NHX gene sequences of Arabidopsis and Oryza sativa were downloaded from the Arabidopsis Information Resource database (http://www.arabidopsis.org) and the Rice Genome Annotation Project (http://rice.plantbiology.msu.edu//), respectively. Further, the NHX genes were used as queries to search against Lonicera japonica genome databases to identify NHX genes from honeysuckle (Dong et al., 2021). Then, the Na+/H+ exchanger domain (PF00999) of honeysuckle NHX was confirmed using NCBI Conserved Domain Database (CDD, https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) and InterProscan (http://www.ebi.ac.uk/Tools/pfa/iprscan/) databases (Liu et al., 2019). Finally, Expert Protein Analysis System (ExPASy, http://web.expasy.org/compute_pi/) was used to predicate the isoelectric point (pI) and molecular weight (MW) (Bassil et al., 2019 ; Bjellqvist et al., 1993). TMHMM Server v2.0 (https://services.healthtech.dtu.dk/service.php?TMHMM-2.0) was used to predicate the transmembrane helices (TMHs) (Moller, Croning & Apweiler, 2001) and Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/) was used to predicate subcellular localization of NHX protein sequences (Chou & Shen, 2010). The phosphorylation sites in the LjNHX proteins were predicted using NetPhos 3.1 Server. (https://services.healthtech.dtu.dk/service.php?NetPhos-3.1) (Blom, Gammeltoft & Brunak, 1999; Joshi et al., 2021).
Phylogenetic analysis
The protein sequences of all the identified NHXs from seventeen angiosperms were aligned using the Clustal-Omega (https://www.ebi.ac.uk/services) (Wu, Wang & Li, 2019), including five monocots: O. sativa (Os, seven sequences), Sorghum bicolor (Sb, six sequences), Triticum aestivum (Ta, three sequences), Panicum virgatum (Pv, one sequence), Hordeum vulgare (Hv, four sequences), and twelve eudicots: Lonicera japonica (Lj, seven sequences), Arabidopsis thaliana (At, eight sequences), Populus trichocarpa (Pt, eight sequences), Vitis vinifera (Vv, eight sequences), Solanum lycopersicum (Sl, seven sequences), Cucurbita maxima (Cm, five sequences), Gossypium hirsutum (Gh, fifteen sequences), Salicornia europaea (Se, one sequence), Beta vulgaris (Bv, five sequences), Eutrema halophilum (Eh, five sequences), Spinacia oleracea (So, five sequences), Solanum tuberosum (St, five sequences). The software MEGA 6 was used to construct a phylogenetic tree of 100 NHXs using the Maximum likelihood (ML) method. The bootstrap value was 1,000 replicates (Kumar, Stecher & Tamura, 2016). All the sequences of NHX proteins were presented in Table S1. The EMBOSS needle (https://www.ebi.ac.uk/Tools/psa/) was used to calculate the pairwise identity and similarity of proteins (Tian et al., 2017).
Conserved motifs, gene structures, and cis-acting elements analysis
The conserved motifs of the NHX protein sequence from honeysuckle were predicted using the Multiple Expectation Maximization for Motif Elicitation program (MEME version 5.0.5, http://meme-suite.org/tools/meme) (Bailey et al., 2009). The exon/intron structure of NHX proteins was graphically displayed by the Gene Structure Display Serve (GSDS, http://gsds.cbi.pku.edu.cn/) based on the genomic sequences (Hu et al., 2015). For cis-acting regulatory elements, the 2,000 nucleotide sequences upstream of the transcription initiation site were predicted and analyzed using PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) (Lescot et al., 2002; Wu, Wang & Li, 2019).
Chromosome location and Ka/Ks ratio analysis
The chromosome distribution of the NHX genes was mined from the annotation information on the honeysuckle genome database and gene distribution visualized with TBtools (Chen et al., 2020; Wu, Wang & Li, 2019). The MCScanX program (https://github.com/tanghaibao/mcscan) was used to identify the gene duplication events (Wang et al., 2012). The synonymous (Ks) and non-synonymous (Ka) substitution of each duplicated gene pair were calculated using the PAL2NAL program (http://www.bork.embl.de/pal2nal/) (Goldman & Yang, 1994).
Protein tertiary structure prediction
The tertiary structure of NHX proteins was predicted using the I-TASSER program (https://zhanglab.ccmb.med.umich.edu/I-TASSER/) (Yang et al., 2015). LOMETS was a multiple threading approach that could be used to identify the best structural templates from the Protein Data Bank (PDB) database (Berman et al., 2003; Wu & Zhang, 2007).
Protein-protein interaction network analysis
The protein-protein interaction (PPI) network of LjNHX proteins was predicted using a model plant Arabidopsis on STRING protein interaction database (STRING, http://string-db.org) (Szklarczyk et al., 2019).
Plant material, treatment, and qRT-PCR analysis
The salt-tolerant honeysuckle cultivar ‘Huajin 6’ was used as the material for pot cultivation in the greenhouse of Shandong University of Traditional Chinese Medicine Medicinal Botanical Garden (Huang et al., 2021; Huang et al., 2019b). Five tissues including mature leaf, young leaf, flower, stem, and root of 2-year-old honeysuckle were collected for tissue-specific expression analysis in June 2021. To verify the genes regulated by salt stress, the annual seedling of honeysuckle was transplanted to plastic containers filled with quartz sand/vermiculite (1/3) in April 2021. After three months, the seedlings were treated with the mixed solution containing 1/2 Hoagland’s nutrient solution and NaCl (0, 100, 200, or 300 mM) solution. Roots were collected from the seedlings at 0, 3, 6, 12, 24, 48, and 72 h after treatments for RT-qPCR analysis.
Total RNA was extracted from the samples using a FastPure Plant Total RNA Isolation Kit (Vazyme, Beijing, China). RNA was reverse transcribed to cDNA using PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa, Shiga, Japan). Primers for RT-qPCR were designed using Primer Premier 6 based on the CDS of genes with melting temperature of 58–62 °C. RT-qPCR analysis was performed using a CFX96 Real-Time System (BIO-RAD, Hercules, CA, USA) with TB Green Premix Ex Taq II (TaKaRa, Shiga, Japan). The relative expression level of LjNHXs in different tissues and gradient salt stress was calculated by the 2–ΔCt and 2–ΔΔCt methods, respectively (Wang et al., 2020a; Zhu et al., 2016). The data were subjected to analysis of variance with Tukey’s multiple range tests means at a significant level of P < 0.05 using the SPSS. Three replicates were performed for each sample. The primers used in this study are listed in Table S1.
Results
Identification of LjNHX genes
A total of eight NHX genes were finally obtained from the honeysuckle genome. These honeysuckle genes were named LjNHX1-LjNHX7 according to their chromosomal positions. As shown in Table 1a, the physicochemical properties showed that the deduced amino acid lengths of LjNHXs were exhibited from 380 aa (LjNHX4) to 1,148 aa (LjNHX6), with an average length of 591 aa. The pI of the LjNHX proteins ranged from 5.88 to 8.15, with an average pI of 7.12. The predicted M.W of the LjNHX proteins ranged from 41.71 to 127.03 kDa, with an average M.W of 75.29 kDa. All the LjNHXs were typical transmembrane transporters, possessed the Na+/H+ exchanger domain. LjNHX1/6/7 contained 12 transmembrane helices. LjNHX2 contained 11 transmembrane helices. LjNHX3, LjNHX5, LjNHX4 contained 10, 9, 5 transmembrane helices, respectively. The prediction of subcellular localization showed that all LjNHX proteins might be located in the vacuoles except LjNHX6, which could be found in the plasma membrane. LjNHXs phosphorylation sites vary in number (Table 1b), serine sites ranged from 25 (LjNHX5) to 87 (LjNHX6), threonine sites ranged from 15 (LjNHX4) to 39 (LjNHX6), while tyrosine sites ranged from zero (LjNHX1) to 9 (LjNHX6). LjNHXs were mostly phosphorylated with PKC, CdC2, and PKA, respectively, and very little with ATM.
| (a) Characteristics of NHX genes identified from honeysuckle. | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene Name | Gene ID | Chr | No. Amino acid | pI | Protein M.W (kDa) | Exon/Intron | Arabidopsis ortholog | TM domains | Subcellular localization | Na+/H+ exchanger domain |
| LjNHX1 | Ljap00034208 | 2 | 512 | 7.72 | 56.05 | 13/13 | At3g05030 | 12 | Vac | 84–407 |
| LjNHX2 | Ljap00014455 | 3 | 536 | 8.15 | 59.21 | 14/14 | At3g05030 | 11 | Vac | 40–427 |
| LjNHX3 | Ljap00006626 | 4 | 536 | 6.78 | 58.94 | 14/13 | At3g05030 | 10 | Vac | 52–439 |
| LjNHX4 | Ljap00011905 | 7 | 380 | 5.88 | 41.71 | 9/9 | At3g05030 | 5 | Vac | 10–274 |
| LjNHX5 | Ljap00015571 | 8 | 503 | 7.71 | 55.48 | 14/15 | At3g05030 | 9 | Vac | 51–398 |
| LjNHX6 | Ljap00035581 | 8 | 1148 | 5.93 | 127.03 | 23/22 | At2g01980 | 12 | PM | 29–451 |
| LjNHX7 | Ljap00009930 | 9 | 522 | 7.65 | 58.32 | 13/12 | At5g55470 | 12 | Vac | 53–439 |
| (b) Phosphorylation sites of LjNHX | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein name | Phosphorylation sites | PKC | CKII | RSK | PKA | UNSP | EGFR | INSR | PKG | CKI | DNAPK | CdC2 | p38MAPK | CDK5 | GSK3 | ATM | SRC | ||
| S | Th | Ty | |||||||||||||||||
| LjNHX1 | 35 | 16 | 0 | 13 | 2 | 0 | 7 | 15 | 0 | 0 | 0 | 5 | 1 | 6 | 2 | 1 | 1 | 1 | 0 |
| LjNHX2 | 31 | 17 | 4 | 14 | 1 | 0 | 5 | 29 | 1 | 2 | 0 | 1 | 1 | 6 | 1 | 1 | 0 | 0 | 0 |
| LjNHX3 | 32 | 21 | 1 | 16 | 1 | 0 | 4 | 23 | 0 | 0 | 0 | 0 | 2 | 5 | 1 | 1 | 0 | 0 | 0 |
| LjNHX4 | 26 | 15 | 2 | 10 | 2 | 0 | 2 | 24 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 1 |
| LjNHX5 | 25 | 19 | 2 | 15 | 2 | 0 | 4 | 15 | 1 | 0 | 0 | 0 | 1 | 5 | 1 | 2 | 0 | 0 | 0 |
| LjNHX6 | 87 | 39 | 9 | 24 | 11 | 1 | 11 | 58 | 0 | 3 | 3 | 7 | 3 | 12 | 1 | 1 | 0 | 0 | 0 |
| LjNHX7 | 30 | 13 | 4 | 11 | 0 | 1 | 5 | 23 | 0 | 0 | 0 | 0 | 1 | 5 | 1 | 0 | 0 | 0 | 0 |
Note:
PKC, protein kinase C; CKII, Casein kinase 2; RSK, Ribosomal S6 kinase; PKA, Protein kinase A; UNSP, Un secified phosphorylation; EGFR, Epidermal growth factor receptor; INSR, Insulin receptor precursor; PKG, Protein kinase G; CKI, Casein kinase 1; DNAPK, DNA dependent protein kinase; CDC2, Cell Division cycle protein 2; P38MAPK, P38 Mitogen activated protein kinase; CDK5, Cyclin dependant kinase 5; GSK3, Glycogen synthase kinase 3; ATM, Ataxia telangiectasia mutated; S, Serine; Th, Threonine; Ty, Tyrosine.
Phylogenetic analysis
The phylogenetic tree (Fig. 1) divided all the 100 NHX proteins into three clades based on their predicted subcellular localization, Vac (vacuolar membrane), Endo (endomembrane), and PM (plasma membrane). There were 85 proteins in the Vac-clade, indicating that most types of NHX proteins from different species are vacuolar membrane-bound, while the PM-clade contained 13 proteins and Endo-clade contained 6 proteins. Among the honeysuckle, the Vac-clade had the largest number of members, with seven LjNHX (LjNHX1/2/3/4/5/7) proteins, PM-clade had only one protein (LjNHX6). Putatively, no NHX protein belongs to Endo-clade in honeysuckle.
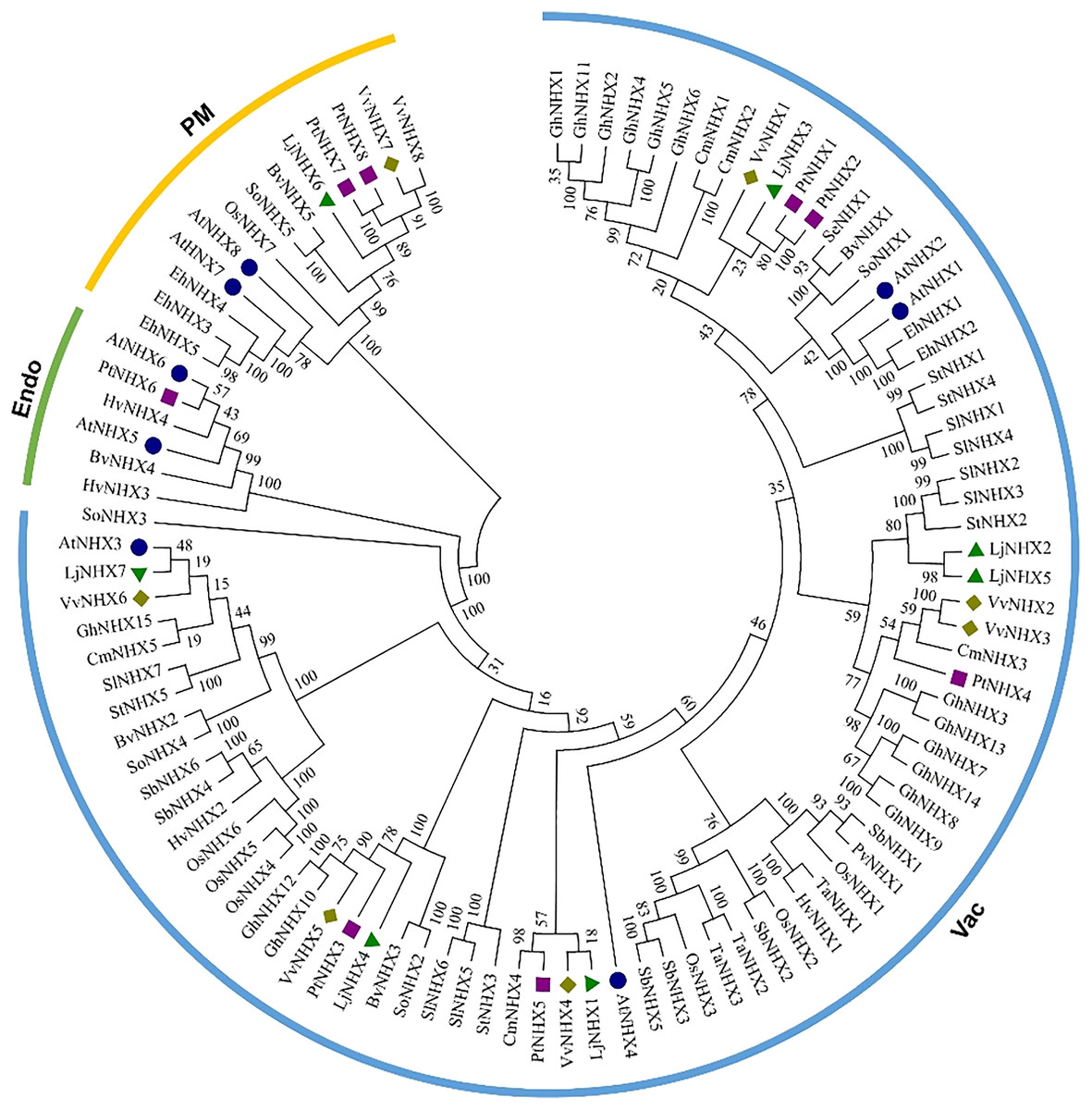
Figure 1: Phylogenetic tree of the NHX proteins.
Phylogenetic tree of the NHX proteins from O. sativa (Os), Sorghum bicolor (Sb), Triticum aestivum (Ta), Panicum virgatum (Pv), Hordeum vulgare (Hv), and twelve eudicots Lonicera japonica (Lj), Arabidopsis thaliana (At), Populus trichocarpa (Pt), Vitis vinifera (Vv), Solanum lycopersicum (Sl), Cucurbita maxima (Cm), Gossypium hirsutum (Gh), Salicornia europaea (Se), Beta vulgaris (Bv), Eutrema halophilum (Eh), Spinacia oleracea (So), Solanum tuberosum (St). The different colored arcs indicate different clades. Proteins from honeysuckle, Arabidopsis, grape, and poplar are denoted by green triangles, blue circles, yellow diamonds, and purple squares, respectively. Details of sequences from seventeen species are listed in Table S1.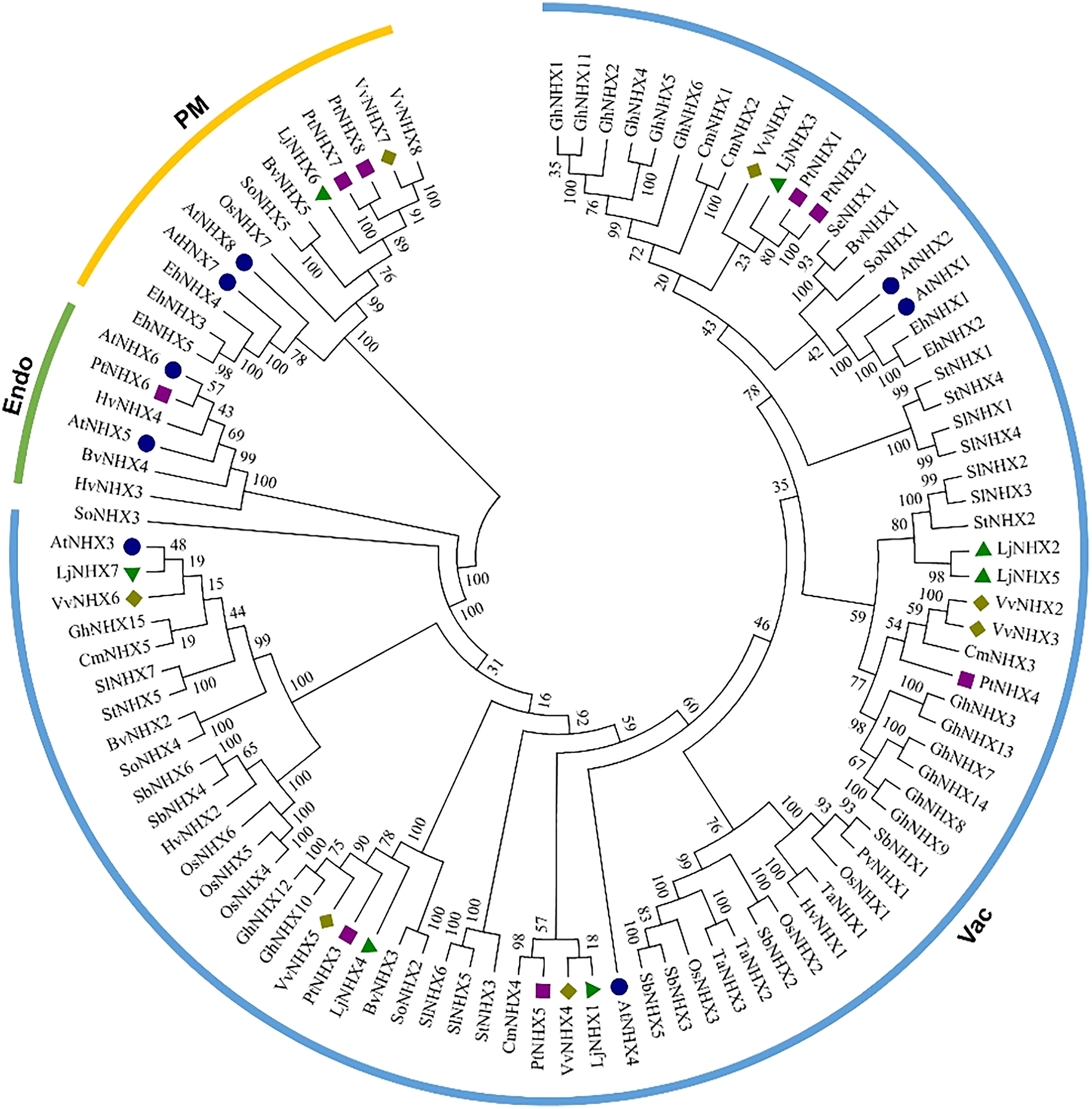{kind=link}
The conservation of the sequence of NHX genes was also confirmed by the identities and similarities of amino acid sequences (Table 2). The results showed that the amino acid sequence identity of different LjNHXs ranged from 8.7% to 78.9%, while the amino acid sequences similarity ranged from 13.8% to 83.6%. The sequences of LjNHX2/ LjNHX5 have higher identities (78.9%), which were members of a close evolutionary relationship. Overall, the genes that belong to the same clades exhibit higher identities.
| Identity (%) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein names | LjNHX1 | LjNHX2 | LjNHX3 | LjNHX4 | LjNHX5 | LjNHX6 | LjNHX7 | AtNHX1 | AtNHX2 | AtNHX3 | AtNHX4 | AtNHX5 | AtNHX6 | AtNHX7 | AtNHX8 | |
| Similarity (%) | LjNHX1 | 68.2 | 66.2 | 36.4 | 63.3 | 10.5 | 49.6 | 63.0 | 64.2 | 53.4 | 59.3 | 26.8 | 25.0 | 11.4 | 14.7 | |
| LjNHX2 | 76.3 | 76.6 | 44.9 | 78.9 | 10.0 | 53.2 | 73.4 | 73.9 | 56.7 | 66.2 | 30.0 | 27.3 | 12.5 | 15.4 | ||
| LjNHX3 | 76.3 | 84.7 | 42.7 | 72.6 | 10.6 | 54.9 | 74.8 | 76.1 | 56.7 | 66.2 | 30.1 | 26.7 | 12.4 | 16.9 | ||
| LjNHX4 | 46.1 | 53.2 | 51.5 | 41.4 | 8.7 | 36.8 | 42.2 | 41.4 | 37.7 | 36.0 | 20.8 | 19.7 | 8.6 | 12.8 | ||
| LjNHX5 | 72.4 | 83.6 | 80.5 | 48.3 | 9.8 | 50.7 | 69.9 | 69.7 | 52.3 | 62.2 | 26.2 | 25.4 | 11.9 | 16.4 | ||
| LjNHX6 | 17.8 | 17.9 | 18.1 | 13.8 | 18.4 | 10.8 | 10.6 | 11.1 | 10.5 | 10.5 | 10.7 | 12.3 | 60.1 | 42.4 | ||
| LjNHX7 | 62.8 | 66.4 | 69.6 | 49.5 | 61.8 | 18.6 | 53.9 | 53.8 | 68.8 | 51.9 | 27.8 | 26.0 | 11.6 | 15.7 | ||
| AtNHX1 | 74.5 | 84.0 | 86.4 | 51.1 | 79.8 | 18.3 | 68.7 | 87.4 | 56.5 | 65.2 | 29.3 | 28.8 | 12.2 | 16.5 | ||
| AtNHX2 | 75.8 | 83.3 | 86.1 | 51.1 | 79.3 | 18.7 | 67.4 | 92.9 | 56.3 | 65.3 | 29.7 | 28.9 | 12.0 | 16.9 | ||
| AtNHX3 | 66.7 | 70.7 | 72.6 | 48.3 | 66.5 | 18.3 | 78.6 | 71.6 | 71.2 | 54.4 | 28.9 | 27.3 | 11.3 | 16.5 | ||
| AtNHX4 | 68.5 | 75.5 | 76.5 | 44.2 | 69.6 | 17.5 | 64.7 | 74.4 | 75.9 | 68.0 | 30.1 | 26.7 | 11.3 | 14.9 | ||
| AtNHX5 | 42.1 | 45.5 | 46.8 | 33.0 | 41.2 | 18.8 | 45.4 | 45.0 | 45.4 | 46.5 | 46.4 | 76.4 | 11.7 | 14.6 | ||
| AtNHX6 | 41.0 | 44.0 | 43.7 | 32.6 | 42.4 | 21.2 | 42.5 | 46.6 | 45.1 | 44.3 | 44.2 | 84.9 | 11.9 | 17.9 | ||
| AtNHX7 | 20.5 | 21.2 | 20.9 | 14.7 | 19.8 | 73.5 | 20.8 | 21.2 | 21.1 | 20.8 | 20.1 | 19.5 | 19.7 | 48.3 | ||
| AtNHX8 | 27.9 | 26.5 | 28.9 | 22.5 | 28.6 | 51.1 | 29.0 | 28.9 | 30.3 | 29.0 | 25.6 | 26.1 | 31.2 | 56.1 | ||
Chromosomal location, Ka/Ks ratio calculation of LjNHX genes
As shown in Fig. 2, seven LjNHX genes were mapped onto six of the total nine honeysuckle chromosomes, indicating a diverse distribution. Chromosome 8 contained two members of LjNHXs, while chromosomes 2, 3, 4, 7 and 9 each contained only one LjNHXs gene. In this study, however, only one duplicated gene pair (LjNHX2/LjNHX5) was identified, and the gene pair was located on different chromosomes (Table S1). The genes had undergone strong purifying selection pressure because the Ka/Ks ratio between LjNHX2 and LjNHX5 is 0.0979 (Wu, Wang & Li, 2019).
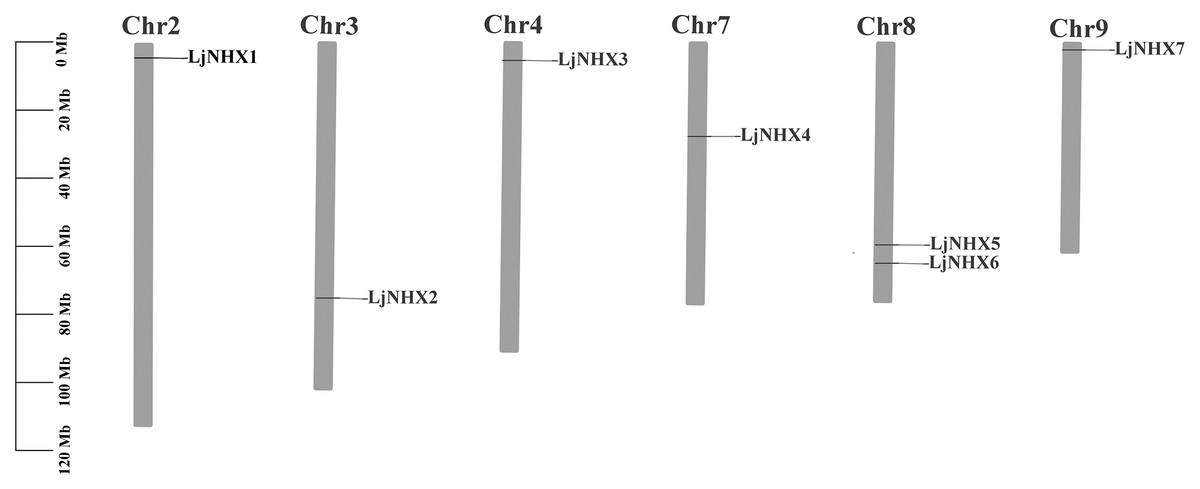
Figure 2: Physical mapping of honeysuckle NHX genes.
The chromosome number is displayed at the top of each chromosome.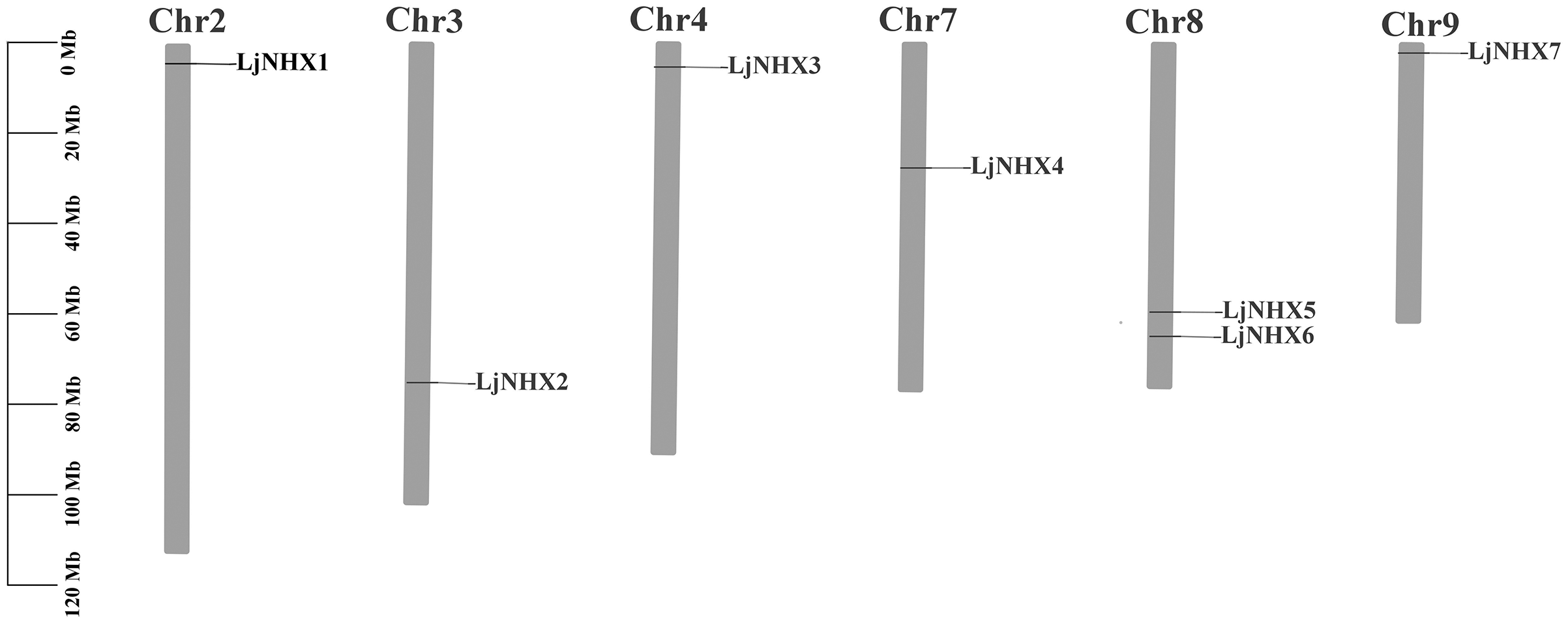{kind=link}
Gene structure and conserved motifs analysis
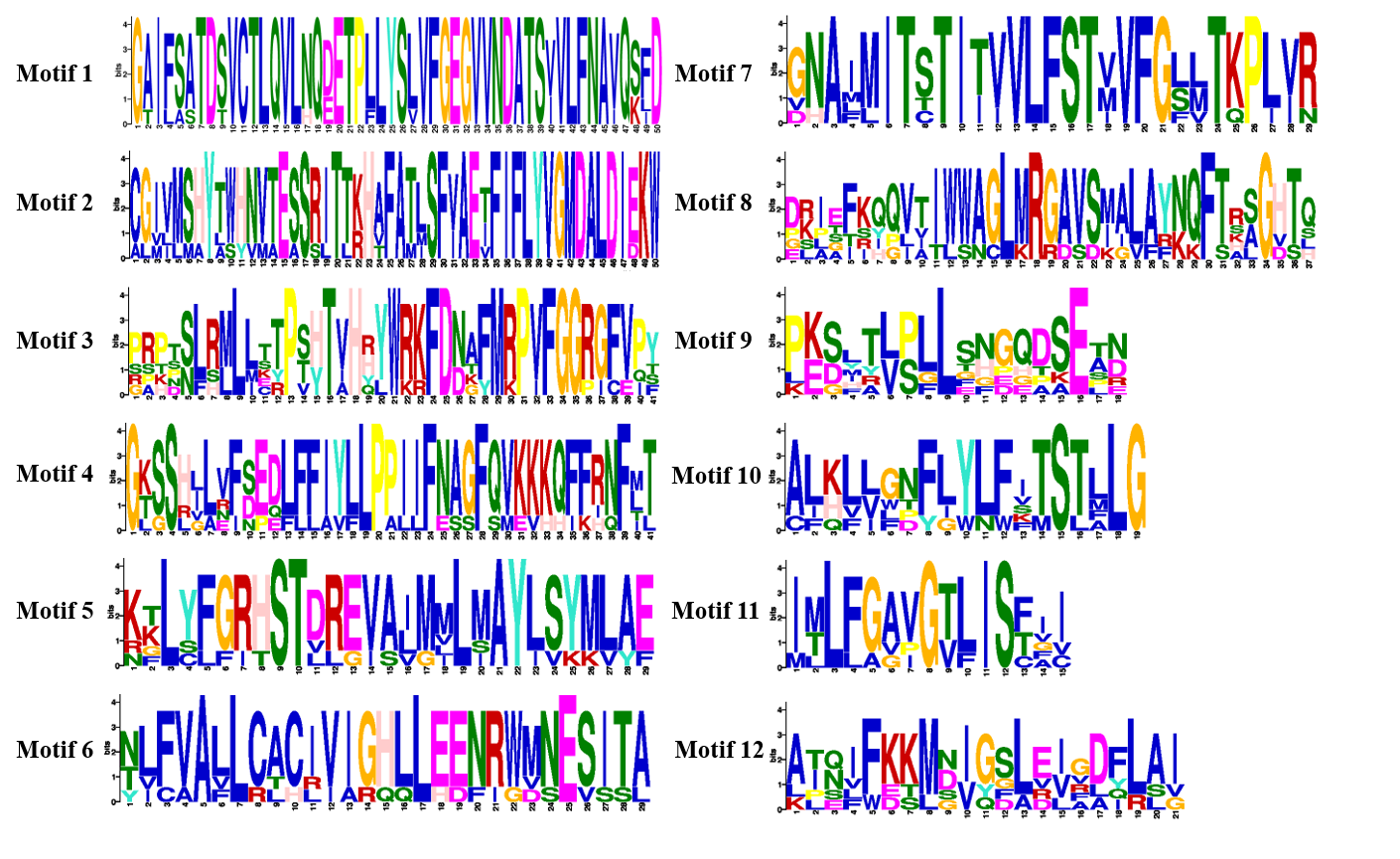
The intron-exon structure of the identified LjNHX genes showed that the coding sequence (CDS) of all the LjNHXs was discontinuous by the presence of introns (Fig. 3A). LjNHX1/2/3/5/7 in Vac-clade contained 13–14 exons and 12–15 introns, but the LjNHX4 contained 9 exons and 9 introns. LjNHX6 in PM-clade had 23 exons and 33 introns. Overall, although gene length differed significantly among the NHX gene family members, the exon length, exons/intron number are moderately conserved among the various subclades, indicating the similar biological functions of members with close evolutionary relationships.
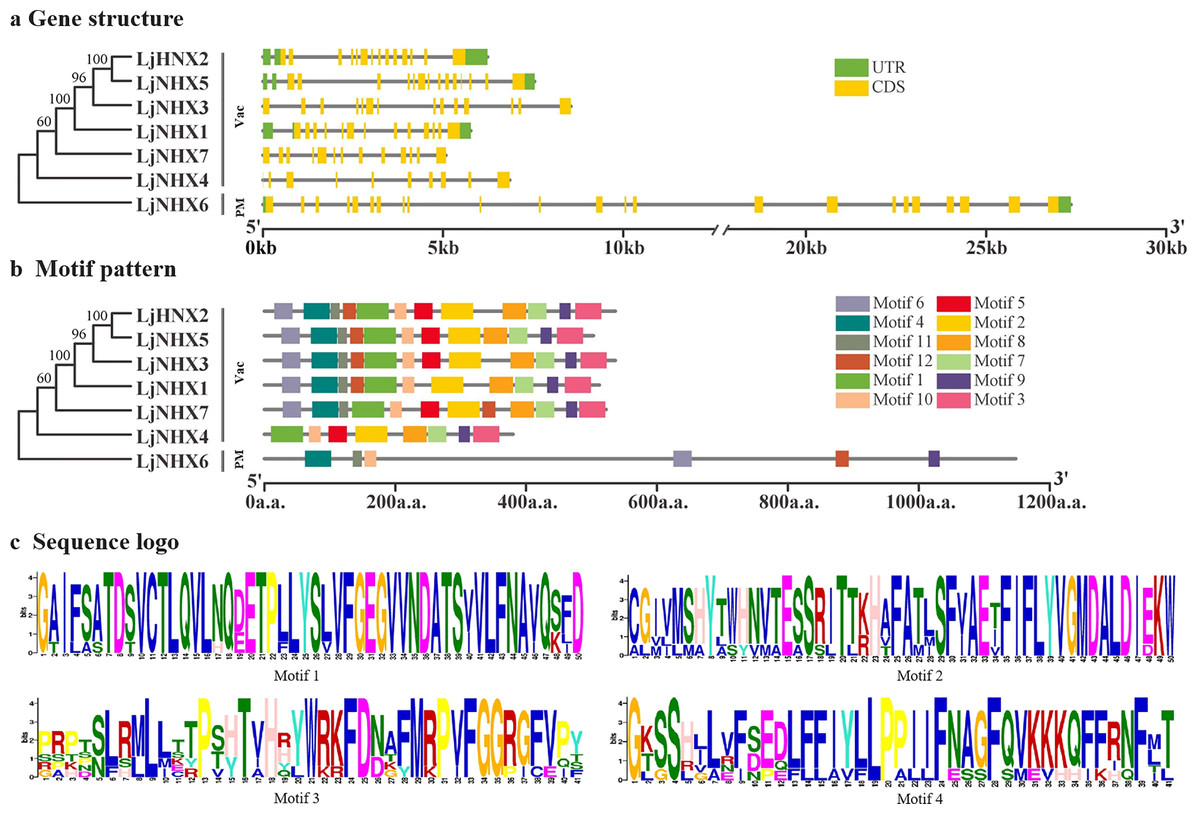
Figure 3: Phylogenetic relationships, gene structure and motif pattern of LjNHXs.
(A) Exon-intron structure. (B) Motif composition. (C) Sequence logo of motif 1, 2, 3, 4. The sequence information and sequence logo for each motif is provided in Table S1 and Fig. S1, respectively.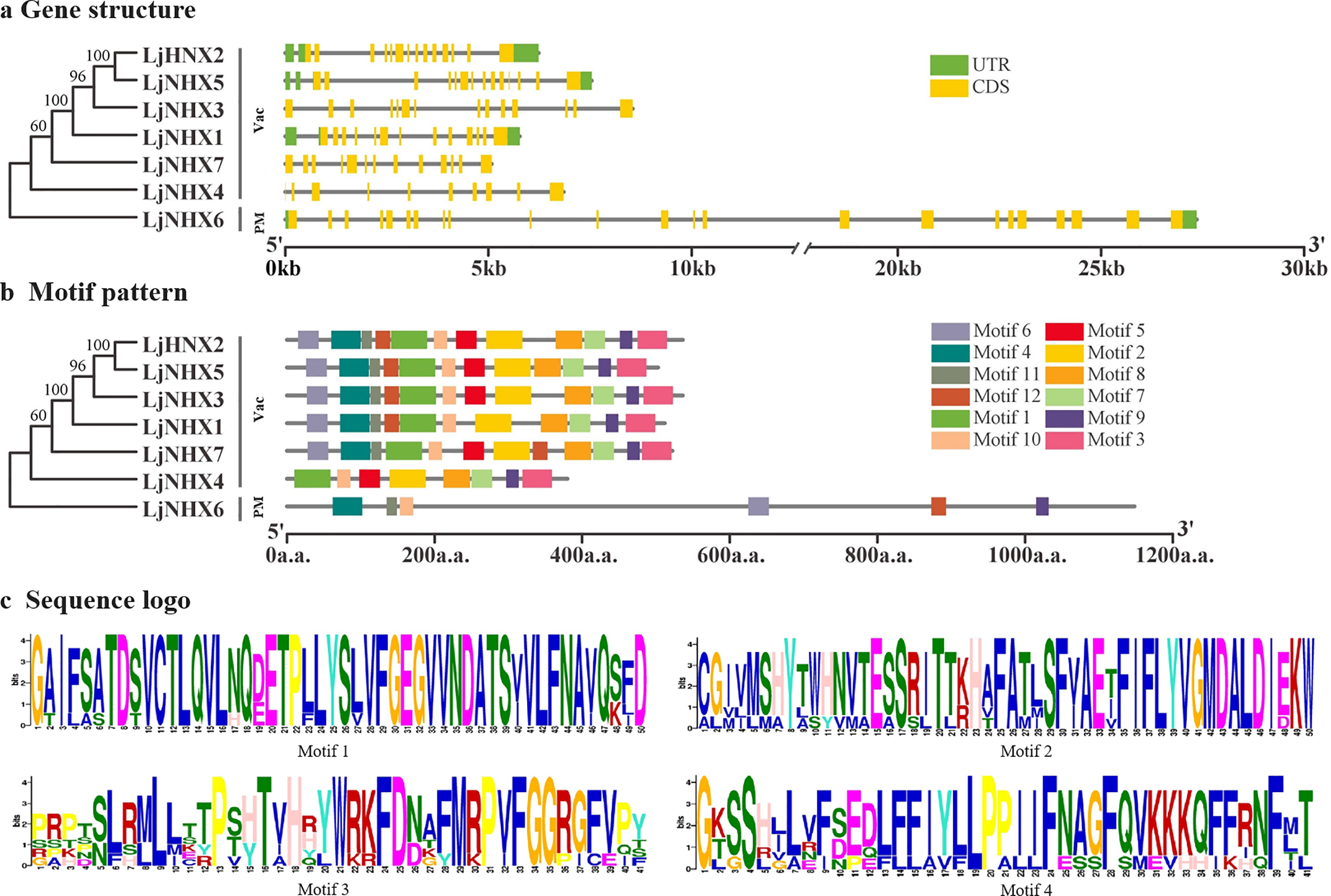{kind=link}
A total of 12 putative motifs were identified in NHX proteins, which represent the characteristic region of proteins (Fig. 3B). Here, motif 9/10 existed in all the members of NHX proteins, whilst motif 1/2/3/7/8 existed in all members of Vac-clade. Amiloride-binding site, a typical feature of NHX protein, was presented in motif 4. Motif 4 was presented in all NHX proteins except LjNHX4. However, there were differences in the composition of amiloride-binding sites. All proteins in Vac-clade were FFIYLLPPI, while LLAVFLPALL was in PM-clade.
Cis-acting elements located in promoters of LjNHXs
In the NHX promoters (Table 3), five hormone-related (e.g., ABRE, TCA-element, CGTCA-motif, TGACG-motif, TGA-motif), seven stress-related (e.g., ARE, LTR, MBS, TC-rich repeats, W box, WUN-motif, STRE), and eight development-related (e.g., G-box, GT1-motif) elements were identified (Wu, Wang & Li, 2019). All of the members contained hormone-related elements, except LjNHX2. LjNHX6 contained four types of hormone-related elements, including ABA (abscisic acid), SA (salicylic acid), MeJA (methyl jasmonate) and IAA (auxin)-responsive element. LjNHX7 contained 14 stress-response elements, which was the largest number of hormone-related elements contained within one gene. Among stress-related cis-acting elements, ARE was found in all LjNHX promoters, while WUN-motif and STRE were found in most LjNHX promoters. LjNHX6 contained six types of stress-related elements, while LjNHX2/3 contained five types of stress-related elements. Additionally, light-responsive elements were found in all LjNHX promoters. These results implied that LjNHX genes could be involved in hormone signal responsiveness and stress adaptation.
| Functional class | Elements | Function | Genes | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LjNHX1 | LjNHX2 | LjNHX3 | LjNHX4 | LjNHX5 | LjNHX6 | LjNHX7 | |||
| Hormone | ABRE | ABA-responsive element | 0 | 0 | 4 | 0 | 2 | 2 | 9 |
| TCA-element | Salicylic acid-responsive element | 1 | 0 | 1 | 2 | 0 | 1 | 0 | |
| CGTCA-motif | MeJA-responsive element | 0 | 0 | 0 | 0 | 0 | 1 | 2 | |
| TGACG-motif | MeJA-responsive element | 0 | 0 | 0 | 0 | 0 | 1 | 2 | |
| TGA-element | Auxin-responsive element | 0 | 0 | 1 | 1 | 0 | 0 | 0 | |
| Stress | ARE | Anaerobic induction | 1 | 2 | 3 | 2 | 1 | 1 | 1 |
| LTR | Low-temperature responsiveness | 2 | 0 | 1 | 0 | 0 | 3 | 0 | |
| MBS | MYB binding site involved in drought-inducibility | 1 | 1 | 1 | 0 | 7 | 0 | 0 | |
| TC-rich repeats | Defense and stress responsive element | 1 | 0 | 0 | 0 | 0 | 1 | 1 | |
| W box | WRKY Transcription factor binding site | 0 | 3 | 0 | 1 | 0 | 2 | 0 | |
| WUN-motif | Wound-responsive element | 0 | 1 | 1 | 1 | 1 | 2 | 1 | |
| STRE | Stress response element | 0 | 3 | 1 | 4 | 2 | 1 | 2 | |
| Others | MYB | Transcription factor | 5 | 4 | 8 | 1 | 9 | 2 | 4 |
| G-box | Light-responsive element | 0 | 0 | 4 | 0 | 1 | 3 | 10 | |
| GT1-motif | Light-responsive element | 1 | 2 | 2 | 0 | 1 | 3 | 3 | |
| TCT-motif | Light-responsive element | 1 | 1 | 1 | 1 | 2 | 0 | 0 | |
| Box 4 | Light responsiveness | 3 | 2 | 1 | 4 | 2 | 1 | 2 | |
| CAT-box | Meristem expression | 1 | 0 | 1 | 1 | 0 | 0 | 0 | |
| GCN4_motif | Endosperm expression | 0 | 1 | 0 | 0 | 0 | 0 | 0 | |
| O2-site | Zein metabolism regulation | 0 | 0 | 0 | 0 | 0 | 1 | 0 | |
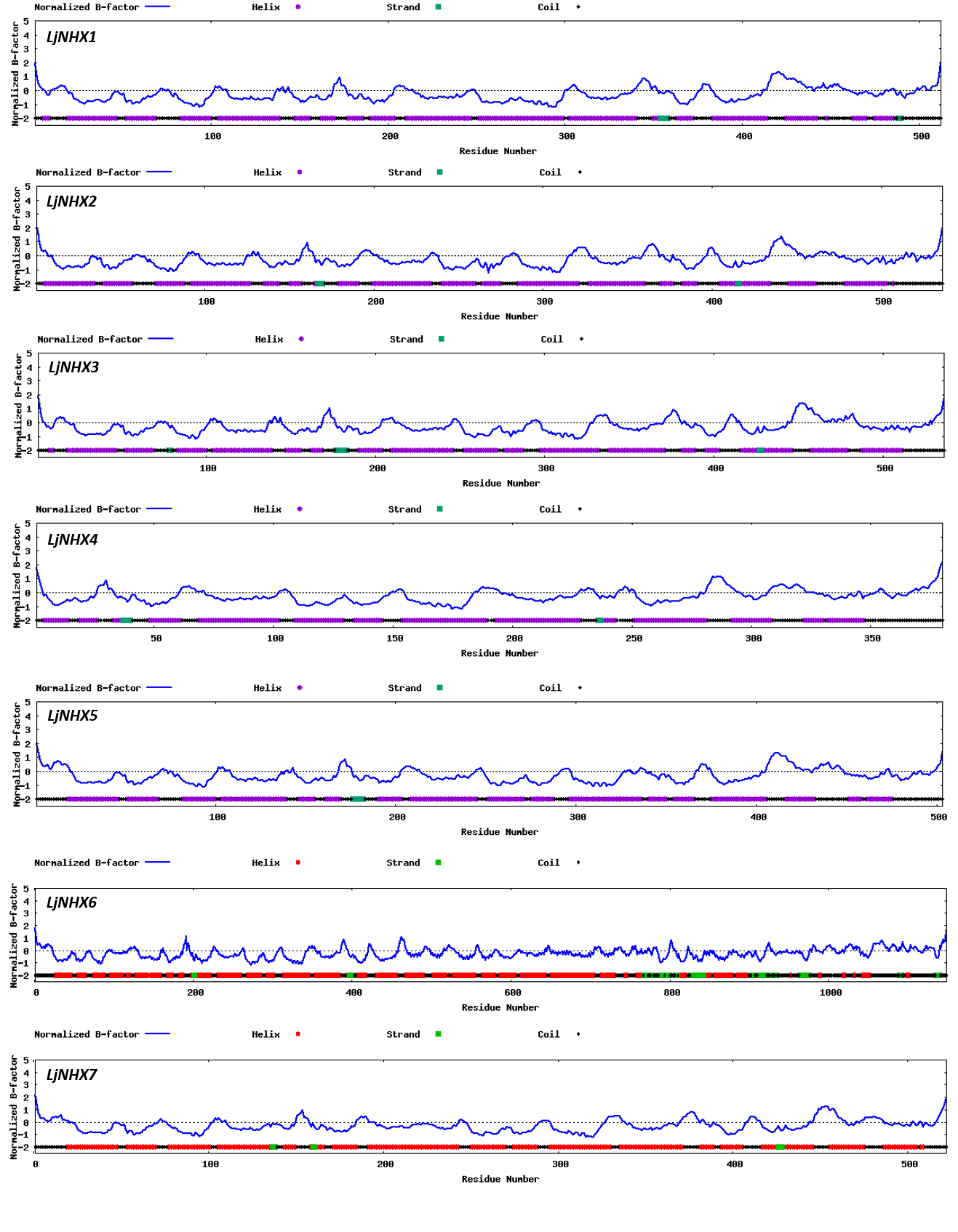
Protein tertiary structure
Tertiary structures of LjNHX protein (Fig. 4) were construed based on the ideal structural templates and crystal structures from Protein Data Bank (PDB) (Tian et al., 2017; Wu, Wang & Li, 2019). All the predicted NHX models in this study had a C-score varied from −1.79 (LjNHX1) to −0.17 (LjNHX7), suggesting the structures of LjNHXs were constructed with high credibility (Table 4). Among them, LjNHX 2/4/5/7 shared the same PDB hit 4cz8A, indicating that their tertiary structures were similar. Therefore, we speculated that LjNHX 2/4/5/7 has similar biological functions.
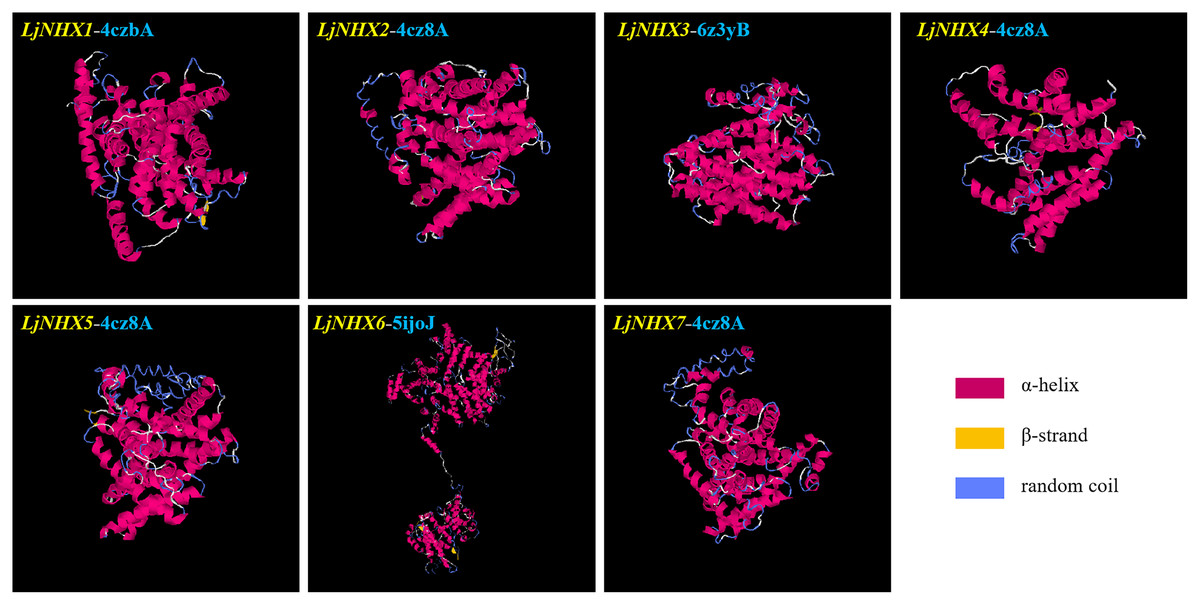
Figure 4: The tertiary structure of seven LjNHX proteins.
Details of the secondary structure of LjNHXs are shown in Fig. S2.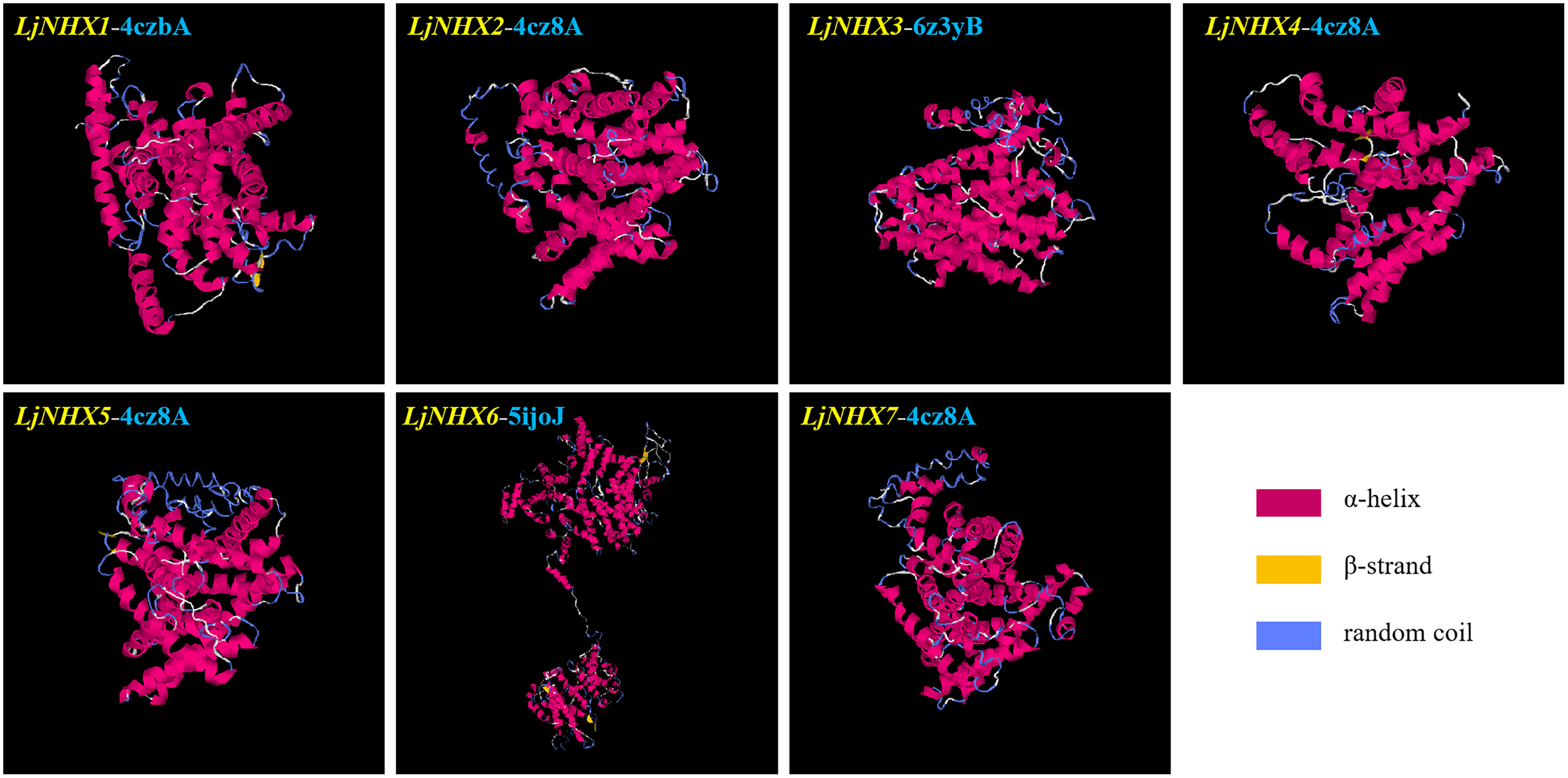{kind=link}
| Protein | C-score | TM-score | RMSD (Å) | Best identified structural analogs in PDB | ||||
|---|---|---|---|---|---|---|---|---|
| PDB hit | TM-score | RMSD | IDEN | Cov | ||||
| LjNHX1 | −1.79 | 0.50 ± 0.15 | 11.6 ± 4.5 Å | 4czbA | 0.722 | 0.68 | 0.177 | 0.727 |
| LjNHX2 | −0.25 | 0.68 ± 0.12 | 8.0 ± 4.4 Å | 4cz8A | 0.719 | 1.19 | 0.207 | 0.731 |
| LjNHX3 | −1.38 | 0.54 ± 0.15 | 10.7 ± 4.6 Å | 6z3yB | 0.708 | 1.1 | 0.312 | 0.718 |
| LjNHX4 | −0.92 | 0.60 ± 0.14 | 8.8 ± 4.6 Å | 4cz8A | 0.697 | 1.31 | 0.21 | 0.716 |
| LjNHX5 | −1.7 | 0.51 ± 0.15 | 11.4 ± 4.5 Å | 4cz8A | 0.717 | 1.69 | 0.201 | 0.742 |
| LjNHX6 | −0.90 | 0.60 ± 0.14 | 11.4 ± 4.5 Å | 5ijoJ | 0.922 | 1.41 | 0.076 | 0.931 |
| LjNHX7 | −0.17 | 0.69 ± 0.12 | 7.8 ± 4.4 Å | 4cz8A | 0.727 | 1.26 | 0.194 | 0.741 |
Note:
C-score [−5, 2] is the confidence of each model, a higher value indicates a model with higher confidence and vice-versa. TM-score and RMSD are determined based on the C-score value and the protein length following the correlation observed between these qualities. TM-score indicates a measure of global similarity between query structure and known structure in PDB. RMSD represents the RMSD between residues that are structurally aligned by TM-align. IDEN is the percentage sequence identity in the structurally aligned region. Cov is the coverage of the alignment by TM-align and is equal to the number of structurally aligned residues divided by length of the query protein.
Protein-protein interaction network analysis
Protein-protein interaction (PPI) network was constructed using STRING database to further explore the potential functions of LjNHX during their interactions with other cellular proteins based on either known experimental or predicted interactions. On the STRING, Lonicera japonica PPI network is not available until now, therefore, we used the homolog gene between Arabidopsis thaliana and Lonicera japonica to predict the LjNHX PPI network (Bassil et al., 2011a). As shown in Fig. 5A, LjNHX4/5/6/7 shared the same putatively interactive proteins, including SOS2, SOS3, HKT1, and AVP1. LjNHX6 interacts with five proteins, such as HKT1, conferring salinity tolerance and RCD1, which supports chloroplasts against high ROS (Reactive oxygen species). SOS2 and SOS3 were involved in the regulatory pathway of salt stress by controlling intracellular Na+ and Ca2+ homeostasis and directly interacted with LjNHX4/5/6/7 proteins. All predicted LjNHX proteins worked together to respond to salt stress.
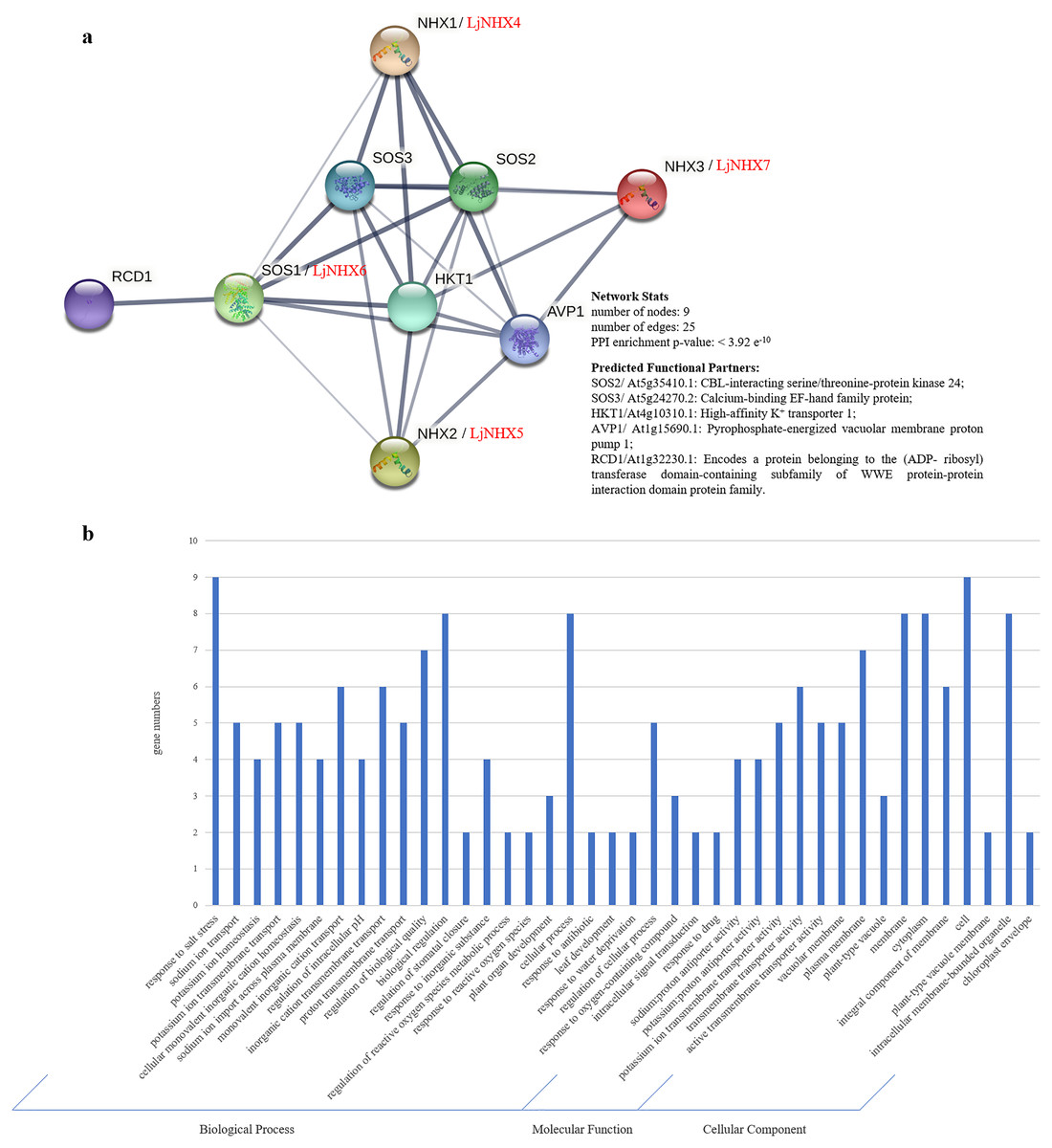
Figure 5: Protein-protein interaction (PPI) prediction of LjNHXs.
(A) PPI network. Line thickness indicates the strength of data support. (B) Gene Ontology (GO) analysis of the genes from the PPI network.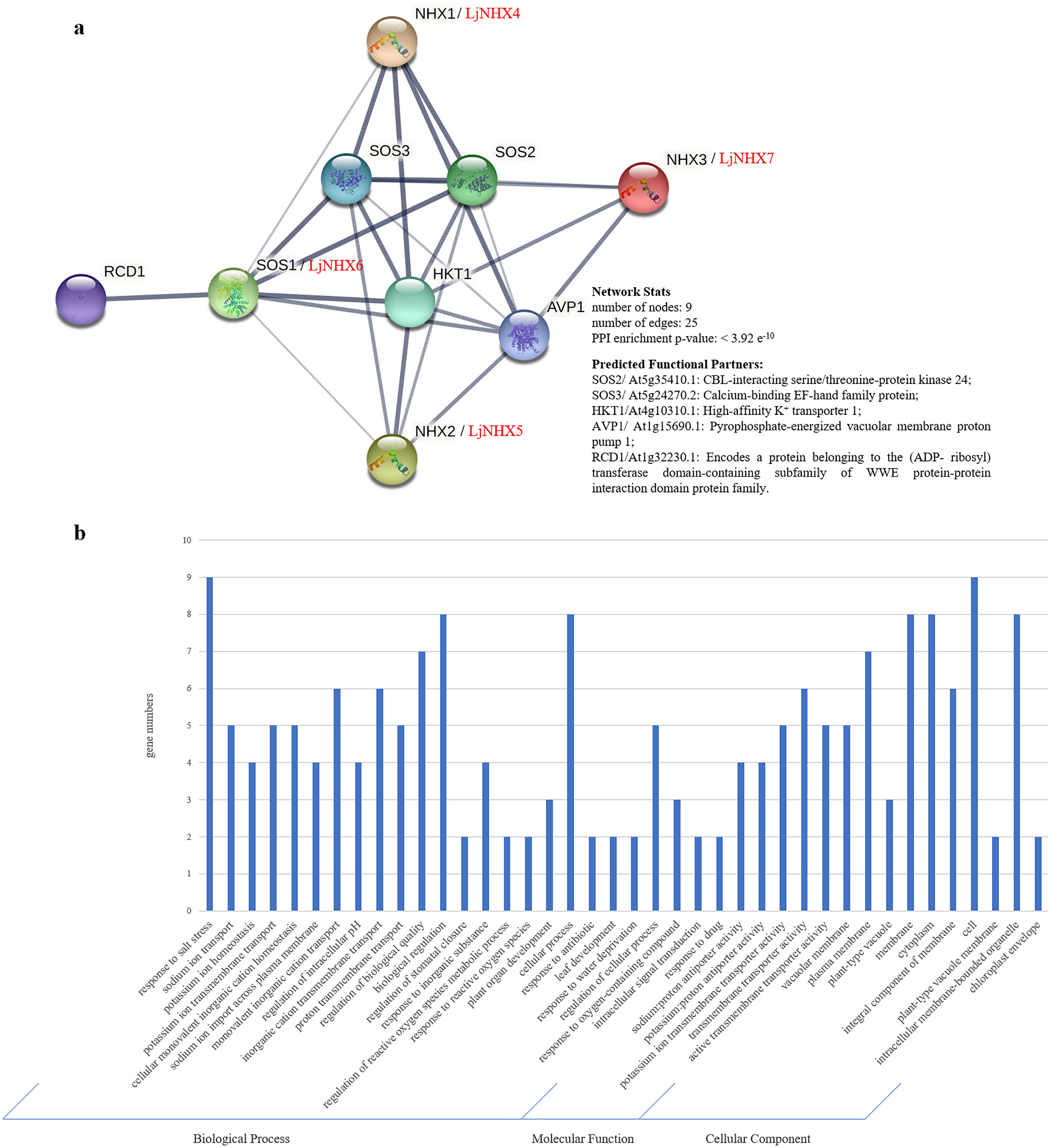{kind=link}
Gene Ontology (GO) analysis (Fig. 5B) was used to describe the proteins from the interaction network, including molecular function (MF), biological processes (BP), and cellular components (CC). It has been shown that the proteins were mainly localized to the plasma membrane, vacuolar membrane, and cytoplasm. Regarding MF, most of the proteins possess transmembrane transporter activity, including sodium: proton transporter activity and potassium: proton transporter activity. In the BP process, most of the proteins mainly participate in the response to salt stress and play a role in ion transmembrane transport and ion homeostasis.
Expression patterns of LjNHXs
For tissue-specific expression analysis (Fig. 6A), the examined genes were expressed in all selected tissue under normal conditions, although their expression levels differed among tissues. LjNHX3/7 had a high level of expression in the mature leaf, while LjNHX6 exhibited a high level of expression in the mature leaf and flower. LjNHX2 had a high level of expression in root and flower. Our results showed that LjNHX genes may play important roles in the growth and development of honeysuckle, and for which functional variations are probable.
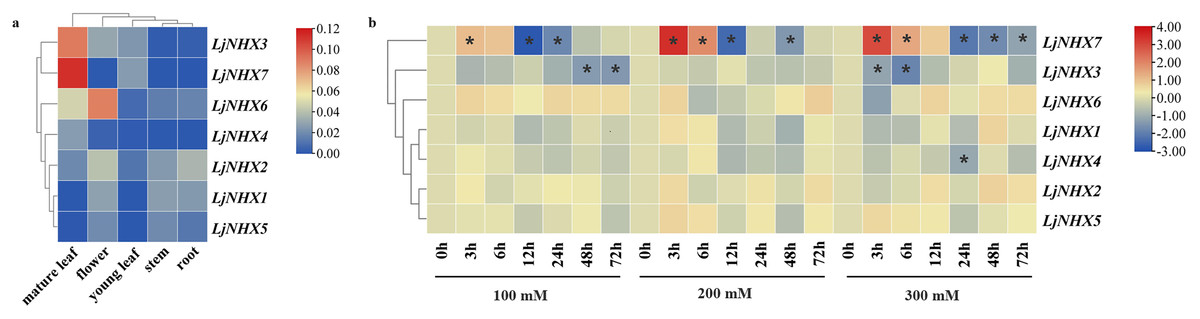
Figure 6: Expression patterns of LjNHX genes.
(A) Expression patterns of LjNHX in five tissues. All values were expressed relative to the expression levels of reference genes using formula 2−ΔCt (Wang et al., 2020a). (B) Expression patterns of the LjNHX at 0, 3, 6, 12, 24, 48, and 72 h after treated with NaCl (100, 200, and 300 mM NaCl). All values were expressed relative to the expression levels of reference genes using formula 2−ΔΔCt. Ljactin was used as a marker gene. Different colors indicate different levels of gene expression based on the log2 value of the fold change by RT-qPCR. The asterisk indicates significant (P < 0.05) up/down-regulated expression (>2-fold).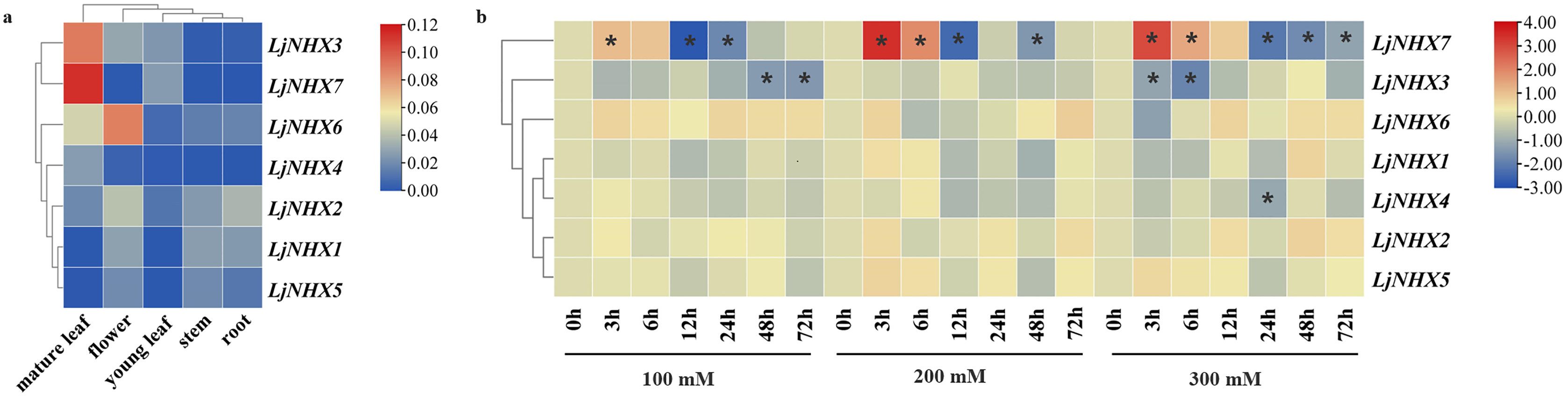{kind=link}
The expression pattern of NHX genes under salinity stress was illustrated in Fig. 6B, the expression of LjNHX7 under all NaCl treatments rapidly increased at the first time and then decreased. The highest expression of LjNHX7 appeared at 3 h, while the lowest expression appeared at 12 h (100 and 200 mM NaCl) and 24 h (300 mM NaCl), respectively. LjNHX3 decreased its expression level at 3 h and maintained a lower expression level over time. Notably, when exposed to low salt stresses (100 mM NaCl), the expression of LjNHX3 reached the lowest level at 72 h, but its expression reached the lowest level at 6 h under high salt concentration (300 mM NaCl).
Discussion
Transmembrane ion transport is a critical process in the cellular response to salt stress. NHXs are secondary ion transporters to exchange H+ and transfer the Na+ or K+ across the membrane, playing an essential regulatory role in maintaining intracellular pH and ion balance (Farooq et al., 2015; Rubio, Gassmann & Schroeder, 1995). Honeysuckle plays an irreplaceable role in the development of TCM (Traditional Chinses Medicine) (Li et al., 2019b; Tang et al., 2021) and has the potential to grow in saline-alkaline soil (Cai et al., 2021a). The whole-genome sequence of the honeysuckle was completed, which made it possible to analyze the NHX gene families of honeysuckle using bioinformatics.
Identification and characterization of the NHX family have been widely reported in some plant species, but the number of members is varied. A total of seven NHX genes were identified in honeysuckle based on the Na+/H+ exchanger domains. There are eight NHX genes in Arabidopsis thaliana (Maser et al., 2001), six in Vitis vinifera (Ma et al., 2015), eight in Populus trichocarpa (Tian et al., 2017), and 25 in Gossypium barbadense (Akram et al., 2020). The difference in the number of NHX genes among different species is due to gene duplication and loss events during evolution (Wu, Wang & Li, 2019). Duplication events provide opportunities for the generation of new genes and their functional divergence in the process of gene family expansion and evolution. As a consequence, the paralogous genes were generated (Tian et al., 2017). The paralogous pair (LjNHX2/LjNHX5) in honeysuckle was generated by segmental duplication, as the genes are present on different chromosomes (Akram et al., 2020). The Ka/Ks ratio gives an insight into the selection pressure on amino acid substitutions. A Ka/Ks ratio > 1 suggests positive selection, Ka/Ks ratio = 1 shows neutral selection, while the ratio of Ka/Ks < 1 suggests purifying selection (Bassil et al., 2011a; Nekrutenko, Makova & Li, 2002). The result from LjNHX2/LjNHX5 pair revealed that LjNHX gene family was strongly purified and selected during long-term evolution and is functionally conserved (Akram et al., 2020).
Most members of the NHX family contain 10–12 transmembrane structures, about 550 amino acid residues, and a putative amiloride-binding domain (FFI/LY/FLLPPI) in the third transmembrane region (Joshi et al., 2021; Ma et al., 2015). But, not all of the members have these characteristics. LjNHX5 and AtNHX5 have nine transmembrane structures, while LjNHX4 only has five transmembrane structures; LjNHX6 has 1,148 aa residues and AtNHX7/SOS1 has 1,146 aa residues. All except LjNHX4 possess an amiloride-binding site in the N-terminal. It has been shown that the presence of amiloride even in a micro concentration in the Na+/H+ antiporters inhibits the transport of Na+ (Bassil et al., 2011a; Carmen & Larry, 1999; Counillon, Franchi & Pouyssegur, 1993). The site was found in most Na+/H+ antiporters of plants, such as Vitis vinifera and Arabidopsis, implying that the site on the transmembrane region of LjNHXs is sensitive to the Na+ of the substrate (Ma et al., 2015; Qiu et al., 2004). However, Ma et al. (2015) showed that the position of this domain was not conserved, and many NHXs genes don’t contain the amiloride-binding domain, including the VIT_15s0024g 00280 in grapevine (Ma et al., 2015).
The phylogenetic analysis serves as an excellent method to determine evolutionary relationships and functional associations among genes. Phylogenetic analysis of NHX genes in honeysuckle, together with five monocots and twelve eudicots, classified in total 100 NHXs into three different clades according to their vacuolar, endosomal, and plasma membrane localization. Similar clustering has also been found in poplar (Populus trichocarpa) (Tian et al., 2017) and Sugar Beet (Beta vulgaris) (Wu, Wang & Li, 2019). In general, subcellular localization is an effective reference for defining the function of the NXH transporter. Consistent with the results from the prediction of TMHMM Server, the phylogenetic analysis of honeysuckle divided the LjNHXs into Vac-clade (LjNHX1/2/3/4/5/7) and PM-clade (LjCIPK6). NHX members located on the PM function in the exclusion and compartmentalization of excess Na+, while the endomembrane-bounded NHXs are essential for cellular cargo trafficking, growth development, and the regulation of protein processing (Akram et al., 2020; Bassil et al., 2011a). However, no gene was identified in the Endo-clade, indicating an uneven distribution of NHXs among subclades and species. Additionally, consistent with the current information of plant evolution, the phylogenetic tree of LjNHXs was more closely related to eudicots, especially grape (Vitis vinifera) and poplar (Populus trichocarpa), compared to monocots. The results of similar exon/intron and motifs patterns within the same subclade further support the accuracy of the phylogenetic tree (Cui et al., 2020). However, the differences in motif patterns among members of the same subclade cannot be ignored. And the structural and motif differences between different subclades reveal the functional diversity of the NHX gene family in honeysuckle (Dong et al., 2021).
As the binding sites of transcription factors, cis-acting regulatory elements play an important role to determine genes’ expression patterns (Wu, Wang & Li, 2019). Hormones, such as ABA, SA, MeJA, and IAA, are essential for every stage of plant development and response to stresses (Gallego-Giraldo et al., 2008; Li et al., 2019a; Mishra et al., 2014; Zhang & Li, 2019). ABA is a common mediator of plant responses to abiotic stress like high salt, low water and high temperature (Nakashima & Yamaguchi-Shinozaki, 2013). ACGT-containing ABA-responsive elements (ABREs, PyACGTGG/TC) were identified upstream for the most LjNHXs, and similar to that detected for many ABA and abiotic stress-inducible genes (Guiltinan, Marcotte & Quatrano, 1990; Hobo et al., 1999; Skriver et al., 1991), indicating that the LjNHXs might be involved in the ABA signal pathway, which mainly controls stomatal closure and physiological responses to salinity, drought, and cold stress (Mishra et al., 2014). TCA-element associated with biotic and abiotic stress was found in LjNHX1/3/4/6. Seven stress-responsive elements were identified namely W box, ARE, MBS, TC-rich repeats, LTR, WUN-motif, STRE, suggesting that LjNHXs responded to many kinds of stresses. For example, STRE elements are activated by heat shock, osmotic stress, low pH, nutrient starvation, etc. ARE is induced by anaerobiosis, and TC-rich repeats play a role in response to defense and stress (Tong et al., 2021). Among them, W box (TTGACC) was identified in LjNHX 2/4/6. W box is recognized by the family of WRKY transcription factors, which is involved in certain developmental processes and stress response (Wu, Wang & Li, 2019), such as salt stress response in Populus (Jiang et al., 2020) and Arabidopsis (Xu et al., 2018). Consistent with this, the expression of multiple genes of LjNHXs was significantly up/down-regulated under NaCl treatment. In short, NHX genes contains a variety of cis-elements related to stress and hormone response, indicating that the NHX family plays a key role in the process of honeysuckle coping with stress.
It was reported that NHXs are involved in salt tolerance responses of different species, such as Gossypium barbadense (Akram et al., 2020) and Glycine max (Joshi et al., 2021). Our study revealed that in honeysuckle, the NHX genes express differentially at different time intervals under different salt stress intensities. As a key pathway for plants to maintain intracellular ion homeostasis, the SOS pathway is composed of SOS3, SOS2 and SOS1 (Ji et al., 2013; Liu et al., 2020). Pairwise sequence alignment results showed that the sequences of LjNHX6 and AtNHX7 (At2g01980) have higher identities. PM-bounded LjNHX6 was predicted to interact with SOS3 and SOS2 in the PPI network and LjNHX6 possesses several hormone- and stress-related cis-acting elements, which indicated the key role of LjNHX6 proteins in the exclusion of Na+ ions from the cell. Besides, as shown in Fig. 6B, expression profile analysis revealed that LjNHX6 showed a higher expression in roots under almost all treatments. The up-regulation of SOS signaling pathway genes in response to salt stress has been confirmed in a variety of plants including poplar (Populus trichocarpa) (Tang et al., 2010) and spinach (Spinacia oleracea) (Zhao, William & Sandhu, 2020). These results collectively illustrate the conservation of SOS pathway genes in honeysuckle, and at the same time, prove that the SOS pathway is a common but necessary pathway for regulating plant salt stress resistance (Zhao, William & Sandhu, 2020), and the detailed mechanism is still to be explored.
The putative interactions between LjNHX4/5/6/7 and HKT1, AVP1 might play essential roles in the salt tolerance of honeysuckle. The GO analysis showed that these genes are involved in sodium ion transport across the membrane and salinity response activities. LjNHX7 showed a unique pattern of first increase and then decrease under salt stress. HKT1 participates in the recirculation of Na+ from above ground to the root system, thereby preventing Na+ from accumulating to a toxic level in shoots (Deinlein et al., 2014). In Puccinellia tenuiflora, the HKT1;5 is strongly expressed in a high salt environment to increase the salinity tolerance by unloading excess Na+ from the xylem (Bassil et al., 2011a; Zhang et al., 2017). AVP1 participates in the regulation of extracellular pH and auxin transport (Dong et al., 2021). Shen et al. (2014) found that the salt tolerance of transgenic cotton co-overexpressed AVP1 and AtNHX1 was improved. The expression profile analyses revealed that LjNHX4 was downregulated under salt stress. In PPI network, LjNHX4 protein was also hypothesized to interact with RCD1, and RCD1 might play a significant role in response to high salt or oxidative stress. In Arabidopsis, SOS1 interacts with RCD1 to increase the tolerance against oxidative stress caused by ROS (Surekha Katiyar-Agarwal et al., 2006). In addition, a high number of stress-related cis-acting elements was observed in promoters of LjNHX4/5/6/7, so we hypothesized that there is an interaction between LjNHX4/5/6/7 and HKT1, AVP1, RCD1 to participate in the salt stress response of honeysuckle.
Conclusions
In the present study, a total of seven LjNHX genes were identified. The phylogenetic analysis divided LjNHX genes into two subclades based on their subcellular localization and the same clade had similar motif compositions and gene structures. Analysis of cis-acting elements showed that LjNHX members may all be involved in hormone signaling response and stress adaptation. PPI network analysis showed that LjNHX4/5/6/7 shared the same putatively interactive proteins, including SOS2, SOS3, HKT1, and AVP1, and LjNHX6 might be the primary Na+/H+ antiporter involved in the SOS pathway during the salt stress response. The GO analysis showed that these genes mainly participate in the response to salt stress and play a role in ion transmembrane transport and ion homeostasis. The salt-induced expression patterns confirmed that the expression levels of LjNHX3/7 were remarkably affected by salinity. The systematic bioinformatics analysis indicates that the NHX family plays an important role in the response of honeysuckle to salt stress, and the results lay a foundation for gene transformation technology, to obtain highly salt-tolerant medicinal plants in the context of the global reduction of cultivated land.
{kind=link}
{kind=link}