Evidence from studies in rodents and in isolated adipocytes that agonists of the chemerin receptor CMKLR1 may be beneficial in the treatment of type 2 diabetes
- Published
- Accepted
- Subject Areas
- Diabetes and Endocrinology, Pharmacology, Metabolic Sciences
- Keywords
- Type 2 diabetes, Glucose uptake, Chemerin receptor, Adipocyte, Obesity, Chemerin, Rosiglitazone
- Copyright
- © 2014 Wargent et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ PrePrints) and either DOI or URL of the article must be cited.
- Cite this article
- 2014. Evidence from studies in rodents and in isolated adipocytes that agonists of the chemerin receptor CMKLR1 may be beneficial in the treatment of type 2 diabetes. PeerJ PrePrints 2:e643v1 https://doi.org/10.7287/peerj.preprints.643v1
Abstract
It is unclear whether the adipokine chemerin has pro- or anti-inflammatory properties, plays any role in the aetiology of obesity or type 2 diabetes, or whether agonists or antagonists of the chemerin receptor CMKLR1 have potential in the treatment of these diseases. To address these questions, we investigatedthe metabolic phenotypes of both male and female, CMKLR1 knockout and heterozygote mice; the effects of murine chemerin and some C-terminal peptides on glucose uptake in wild-type and CMKLR1 knockout adipocytes; and plasma chemerin levels and chemerin gene mRNA content in adipose tissue in models ofobesity and diabetes, and in response to fasting or administration of the insulin sensitizing drug rosiglitazone, which also has anti-inflammatory properties. Both male and female, CMKLR1 knockout and heterozygote mice displayed a mild tendency to obesity and impaired glucose homeostasis, but only when they were fed on a high fat, rather than a standard low fat diet. Obesity and impaired glucose homeostasis did not occur concurrently, suggesting that obesity was not the sole cause of impaired glucose homeostasis. Picomolar concentrations of chemerin and its C15- and C19-terminal peptides stimulatedglucose uptake in the presence of insulin by rat and mouse wild-type epididymal adipocytes, but not by murine CMKLR1 knockout adipocytes. The insulin concentration-response curve was shifted to the left in the presence of 40 pM chemerin or its C-15 terminal peptide. The plasma chemerin level was raised in diet-induced obesity and ob/ob but not db/db mice, and reduced by fasting and, in ob/ob mice, by treatment with rosiglitazone. These findings suggest that an agonist of CMKLR1 is more likely than an antagonist to be of value in the treatment of type 2 diabetes and have associated anti-obesity andanti-inflammatory activities. One mechanism by which an agonist of CMKLR1 might improve glucose homeostasis is by increasing insulin-stimulated glucose uptake by adipocytes.
Author Comment
This is a submission for PeerJ to review.
Supplemental Information
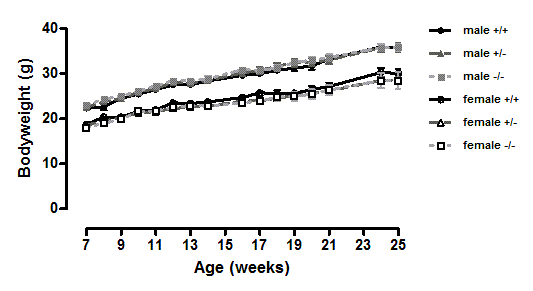
Figure 1. Body weight of wild-type, CMKLR1 knockout and CMKLR1 heterozygote mice fed on a standard chow diet.
n = 15 for wild-type and heterozygote mice, and 11 and 7 for male and female knockout mice respectively. The upper three lines are for males and the lower three for females.
{kind=link}
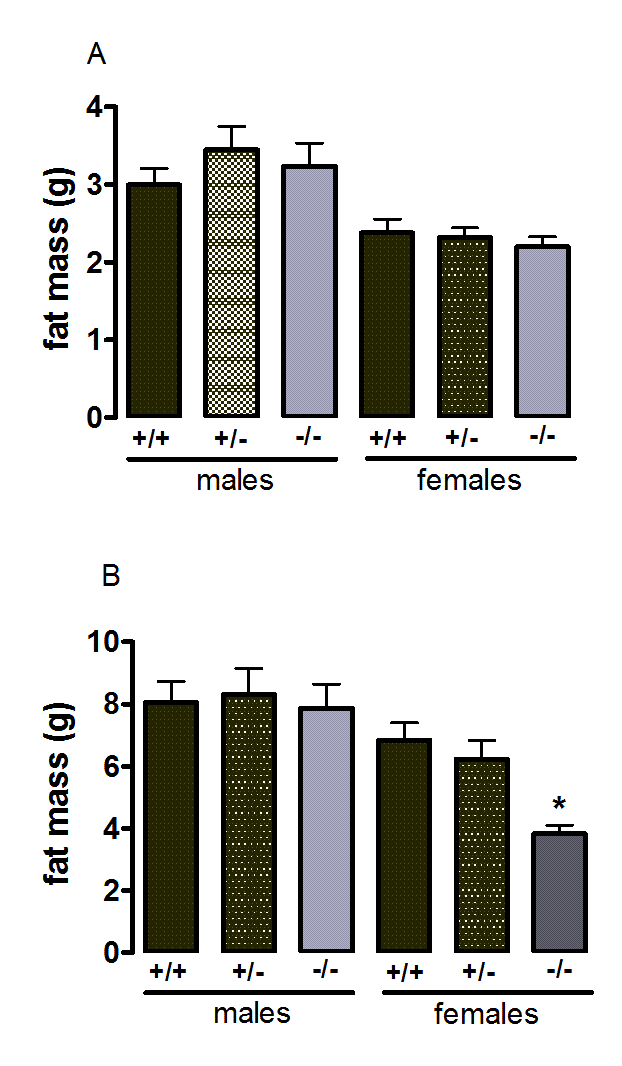
Suppl. Figure 2. Body fat content of wild-type, CMKLR1 knockout and CMKLR1 heterozygotemice fed on a standard chow diet at 12 weeks (A) and six months (B) of age
n values are as for supplementary Figure 1.
{kind=link}
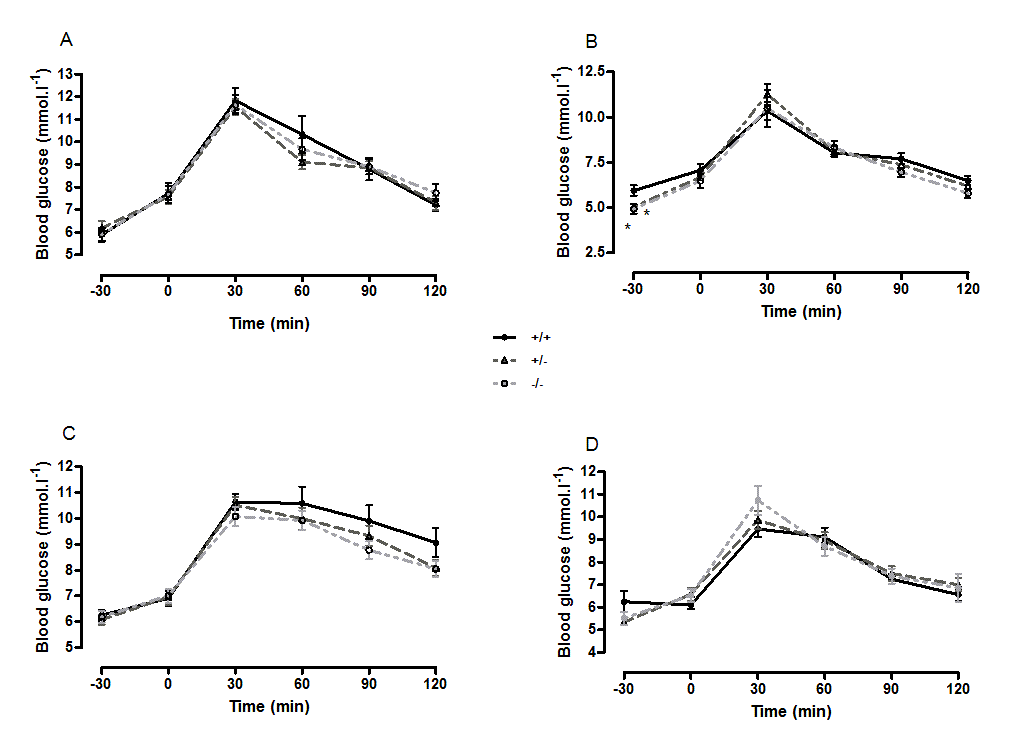
Suppl. Figure 3. Oral glucose tolerance andintraperitoneal insulin tolerance in wild-type, CMKLR1knockout and CMKLR1 heterozygote mice fed on a standard chow diet at 12 weeks (A) and six months (B) of age
n values are as for Supplementary Figure 1. *P<0.05 for knockout and heterozygote compared to wild-type mice of the same sex.
{kind=link}
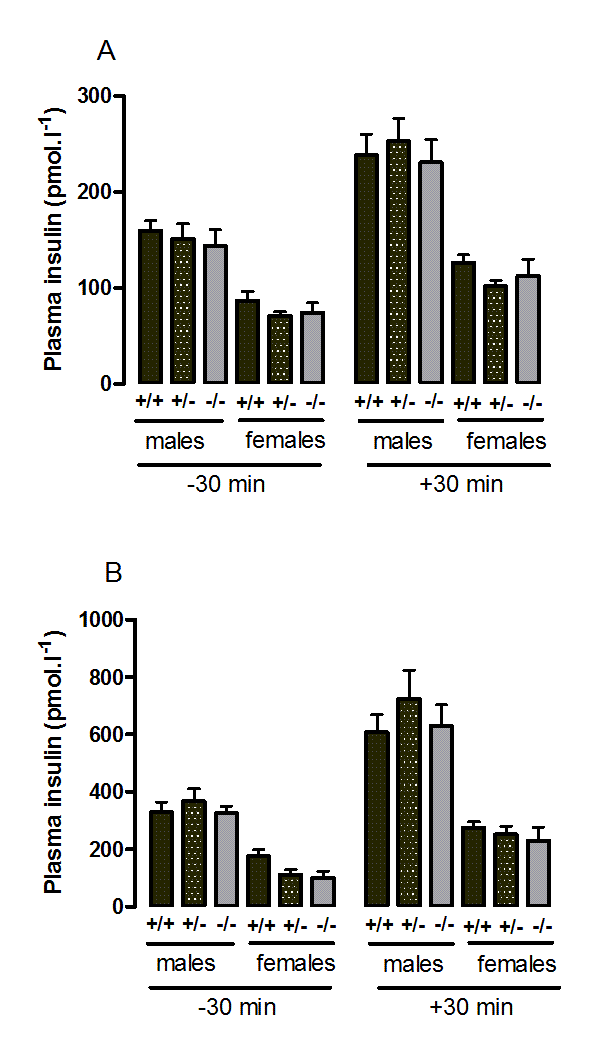
Figure 4. Plasma insulin during the oral glucose tolerance test in wild-type, CMKLR1 knockoutand CMKLR1 heterozygote mice fed on a standard chow diet at 12 weeks (A) and six months (B) of age.
n values are as for Supplementary Figure 1.
{kind=link}
Suppl. Figure 5. Insulin tolerance test in6-month-old wild-type, CMKLR1 knockout and CMKLR1 heterozygote mice fed on a standard chowdiet
n values are as for Supplementary Figure 1.
{kind=link}
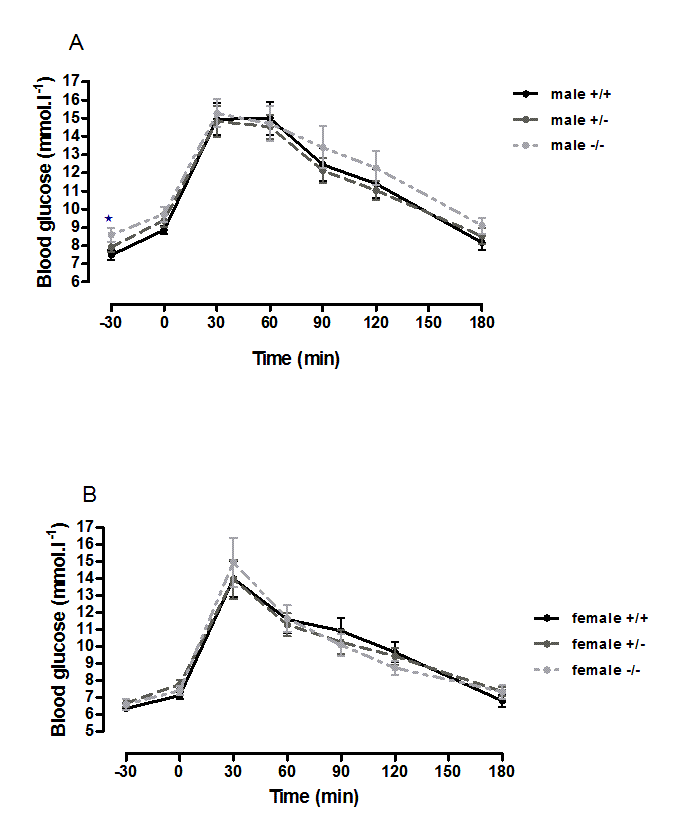
Suppl. Figure 6. Oral glucose tolerance andintraperitoneal insulin tolerance in 6-month-old wild-type, CMKLR1knockout and CMKLR1 heterozygote mice fed on a high fat diet .
n = 17 to 21, except for female knockout mice, where n = 10.
{kind=link}
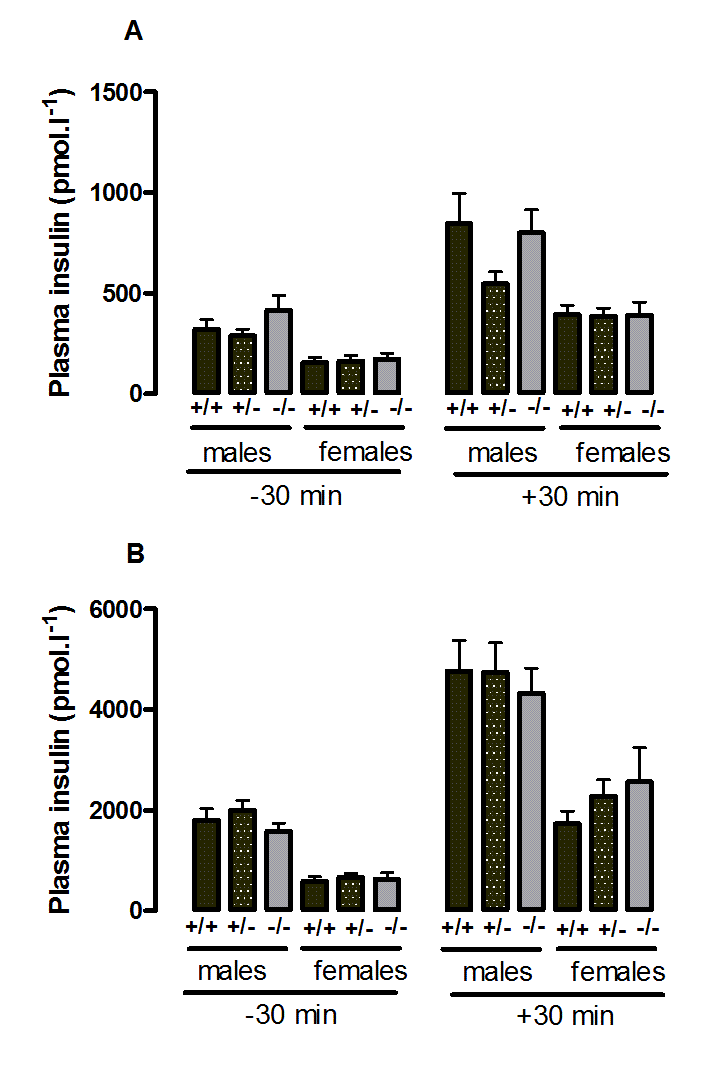
Suppl. Figure 7. Plasma insulin during the oral glucose tolerance test in wild-type, CMKLR1 knockout and CMKLR1 heterozygote mice fed on a high fat diet at 12 weeks (A) and six months (B) ofage.
{kind=link}
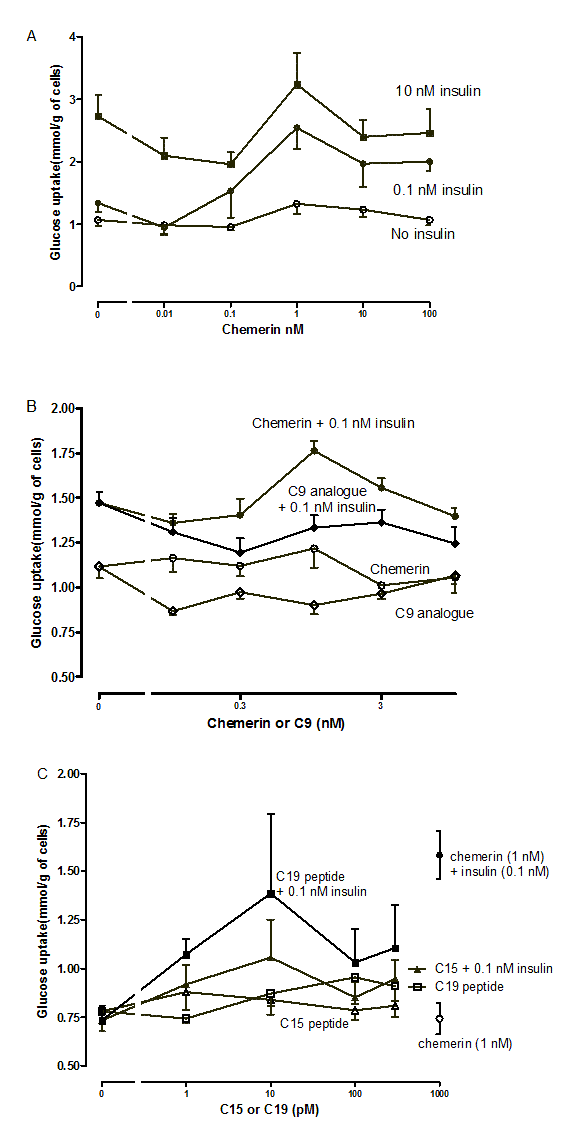
Suppl. Figure 8. Preliminary experiments on the effects of murine chemerin, its C-15 and C-19 terminal peptides and a stable analogue of its C9-terminal peptide on glucose uptake inmurine mesenteric adipocytes.
Panel (A) shows consolidated results for two preparations of adipocytes which gave very similar basal values and where concentrations of chemerin were identical between experiments; n = 4 to 10. Panels(B) and (C) show means for 4 replicates from single preparations of adipocytes.
{kind=link}
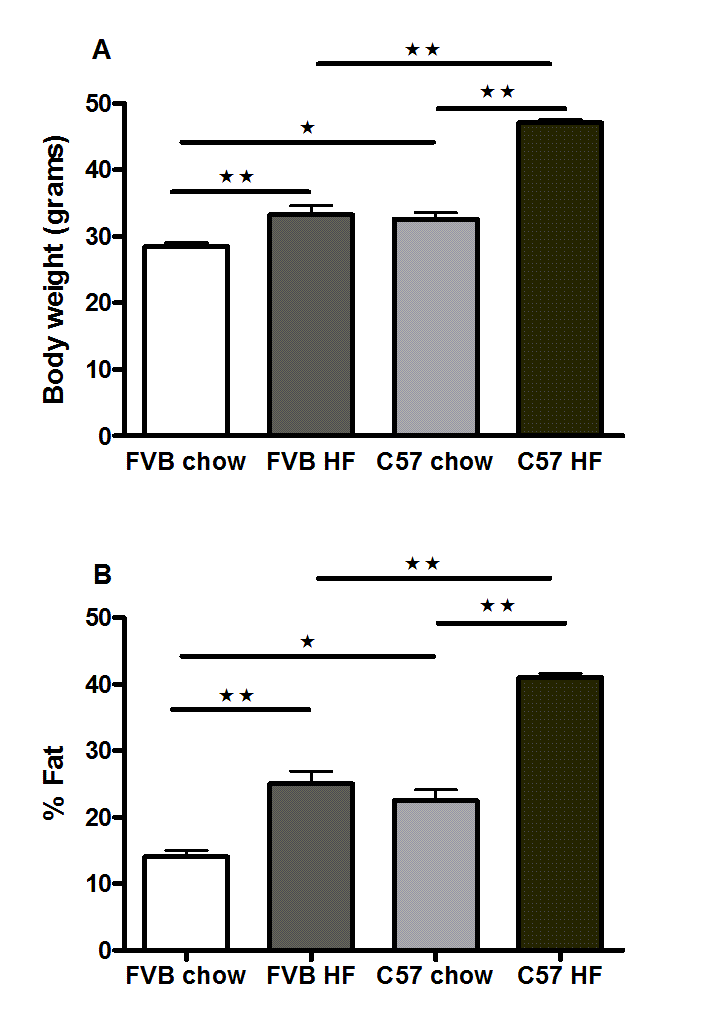
Suppl. Figure 9. Body weights (A) and percentage bodyfat (B) of male FVB and C57Bl/6 mice fed on chow and high fat diets for 6 months.
n = 7 to 9. *P< 0.05; **P<0.01; ***P<0.001 for the comparisons indicated by the bars.
{kind=link}
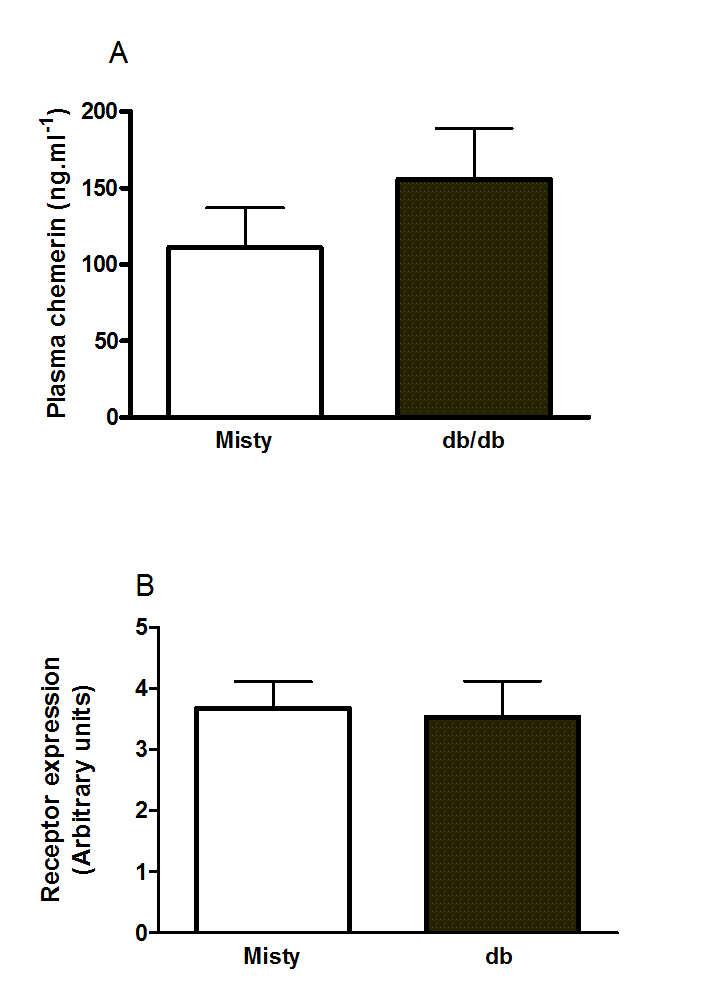
Suppl. Figure10. Plasma chemerin concentration (A)and RARRES2 mRNA levels (B) in inguinal adipose tissue of 11- to 12-week-old male db/db mice and the background C57Bl/Ks strain.
n = 6 in A and 5 in B.
{kind=link}
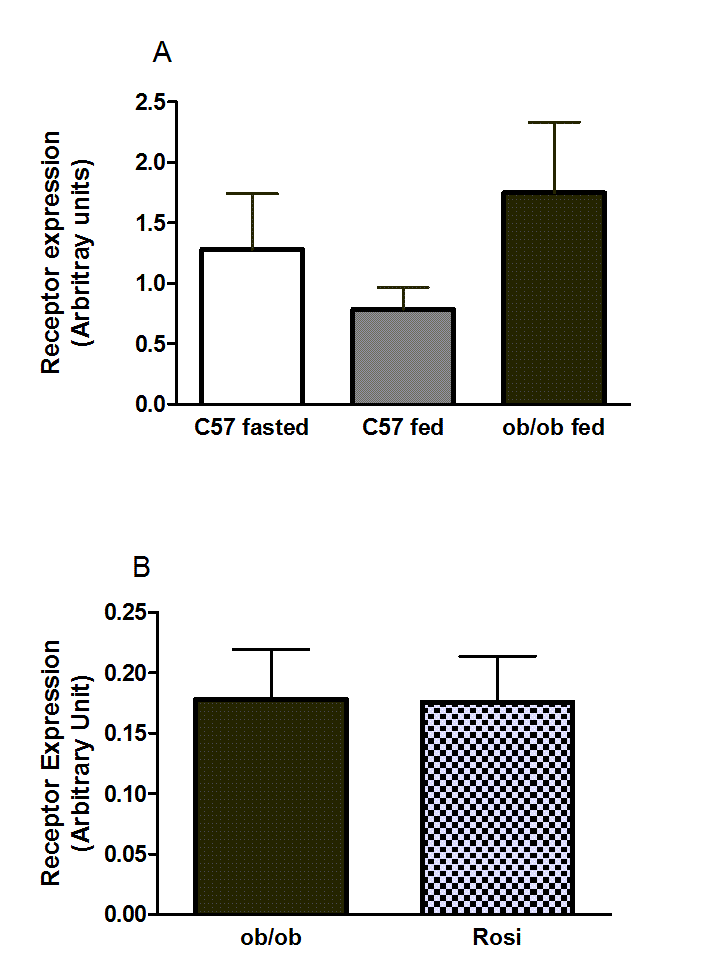
Suppl. Figure11. RARRES2 mRNA in perigenital adipose tissue.
(A) Fed and 5 h-fasted C57BL/6 8-week-old wild-type and 10-week-old ob/ob mice, and (B) female ob/ob mice treated with rosiglitazone for 3 weeks. n values and other details are given in the legends to Figures 7D and 8B of the main paper, which, by contrast, show changes in RARRES2 expression in inguinal fat.
{kind=link}