Almost all known sauropod necks are incomplete and distorted
- Published
- Accepted
- Subject Areas
- Evolutionary Studies, Paleontology
- Keywords
- Sauropod, Dinosaur, Preservation, Neck, Cervical vertebrae
- Copyright
- © 2015 Taylor
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ PrePrints) and either DOI or URL of the article must be cited.
- Cite this article
- 2015. Almost all known sauropod necks are incomplete and distorted. PeerJ PrePrints 3:e1418v1 https://doi.org/10.7287/peerj.preprints.1418v1
Abstract
Sauropods are familiar dinosaurs, immediately recognisable by their great size and long necks. However, their necks are much less well known than is usually assumed. Very few complete necks have been described in the literature, and even important specimens such as the Carnegie Diplodocus and Apatosaurus, and the giant Berlin brachiosaur, in fact have imperfectly known necks. In older specimens, missing bone is often difficult to spot due to over-enthusiastic restoration. Worse still, even those vertebrae that are complete are often badly distorted – for example, in consecutive cervicals of the Carnegie Diplodocus CM 84, the aspect ratio of the posterior articular facet of the centrum varies so dramatically that C14 appears 35% broader proportionally than C13. Widespread incompleteness and distortion are both inevitable due to sauropod anatomy: large size made it almost impossible for whole individuals to be preserved because sediment cannot be deposited quickly enough to cover a giant carcass; and distortion of presacral vertebrae is common due their lightweight pneumatic construction. This ubiquitous incompleteness and unpredictable distortion compromise attempts to determine habitual neck posture and range of motion by modelling articulations between vertebrae.
Author Comment
Although I am submitting this article for formal peer-review at the same time as publishing it as a preprint, I also solicit comments from readers. In particular I am very keen to know if I have missed any complete sauropod necks that have been described in the literature. In the final version of the manuscript, I will acknowledge those who have offered helpful comments.
Supplemental Information
Spinophorosaurus nigerensis holotype
Spinophorosaurus nigerensis holotype GCP-CV-4229 in situ during excavation in the region of Aderbissinat, Thirozerine Dept., Agadez Region, Republic of Niger. Reproduced from Remes et al. (2009: figure 1).
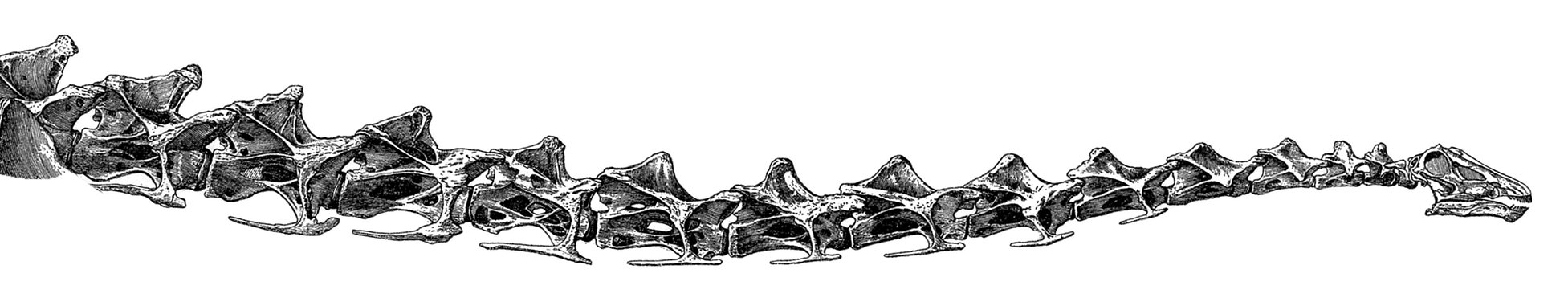
Neck of Diplodocus carnegii
Neck of Diplodocus carnegii holotype CM 84, as reconstructed by Hatcher (1901: plate XIII), with fifteen undamaged cervical vertebrae.
Carnegie Diplodocus quarry
W. H. Reed's diagram of Quarry C near Camp Carnegie on Sheep Creek, in Albany County, Wyoming. The coloured bones belong to CM 84, the holotype of Diplodocus carnegii; other bones belong to other individuals, chiefly of Brontosaurus, Camarasaurus and Stegosaurus. Modified (cropped and coloured) from Hatcher (1901: plate I). Cervical vertebrae are purple (and greatly simplified in outline), dorsals are red, the sacrum is orange, caudals are yellow, limb girdle elements are blue, and limb bones are green.
Unhelpful restoration of a Brachiosaurus altithorax vertebra
Three images of presacral vertebra 6 (probably dorsal 7) of Brachiosaurus altithorax holotype FMNH P25107, in right lateral view, showing misleading restoration. Left: Field Museum photograph CSGEO16166, photographer Charles Carpenter, taken in 1905, the year after Riggs's (1904) descriptive monograph. Note the “crazy-paving” effect of the many cracks and missing areas of bone surface. Middle: Illustration of the same vertebra in Riggs (1904: plate LXXII). Note that the damage to the vertebral surface is not depicted. Right: photograph of the same vertebra, taken by the author in 2005. Note that the damage apparent in the 1905 photograph is no longer visible: the vertebra seems to have been painted to conceal its incompleteness.
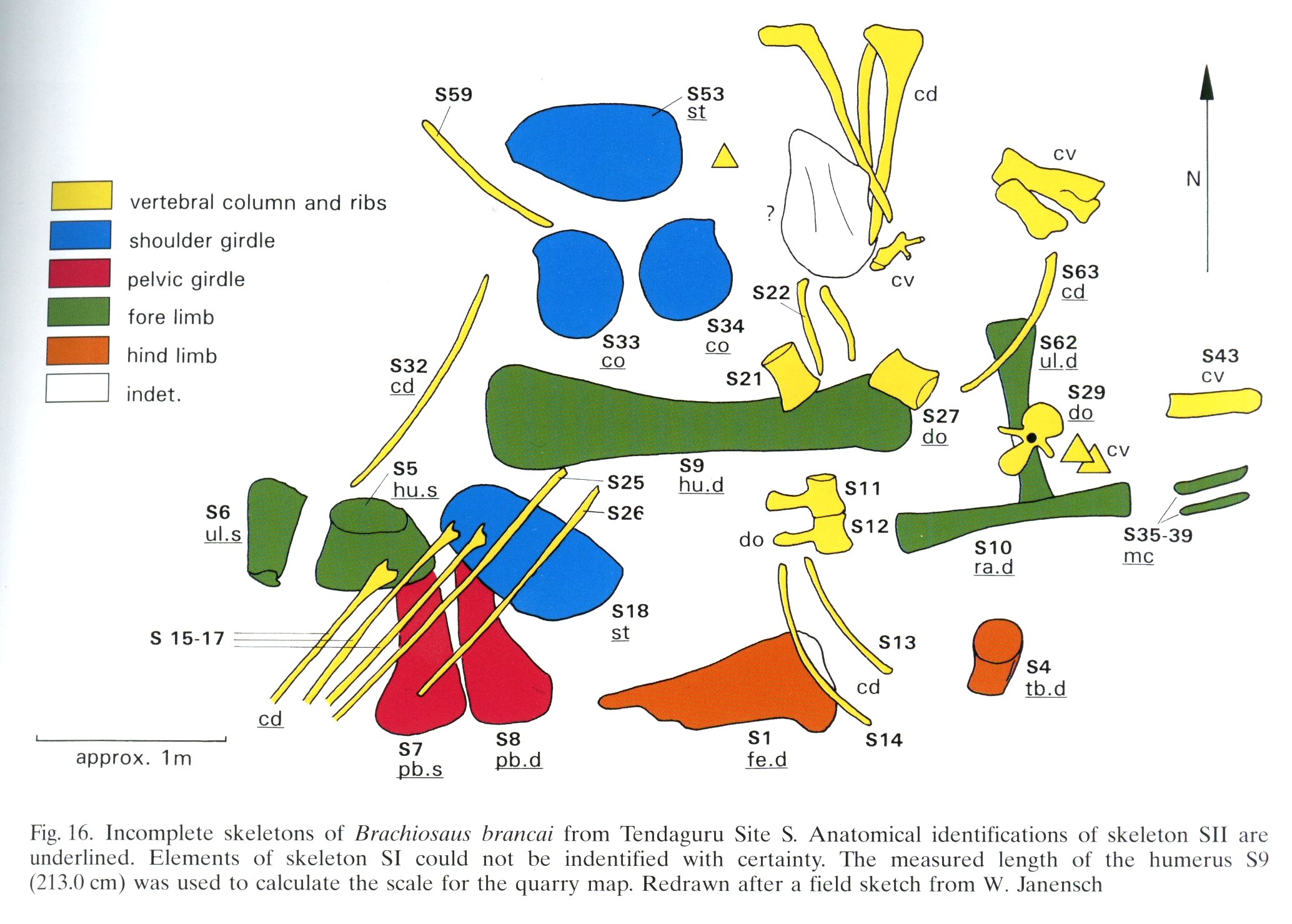
Quarry map of Tendaguru Site S
Quarry map of Tendaguru Site S, Tanzania, showing incomplete and jumbled skeletons of Giraffatitan brancai specimens MB.R.2180 (the lectotype, formerly HMN SI) and MB.R.2181 (the paralectotype, formerly HMN SII). Anatomical identifications of SII are underlined. Elements of SI could not be identified with certainty. From Heinrich (1999: figure 16), redrawn from an original field sketch by Werner Janensch.
Articular facets of vertebrae of extant animals
Sequences of cervical vertebrae of extant animals, showing that articular facet shape remains similar along the column. Top. Cervical vertebrae 3–7 of a mature savannah monitor lizard, Varanus exanthematicus, in anterior view. (The cervicals of monitor lizards, unlike those of sauropods and most mammals, are procoelous, with the anterior facet being concave and the posterior convex.) Bottom. cervical vertebrae 2–5 of a mature house-cat, Felis catus, in posterior view. All photographs by the author, of specimens in his personal collection.
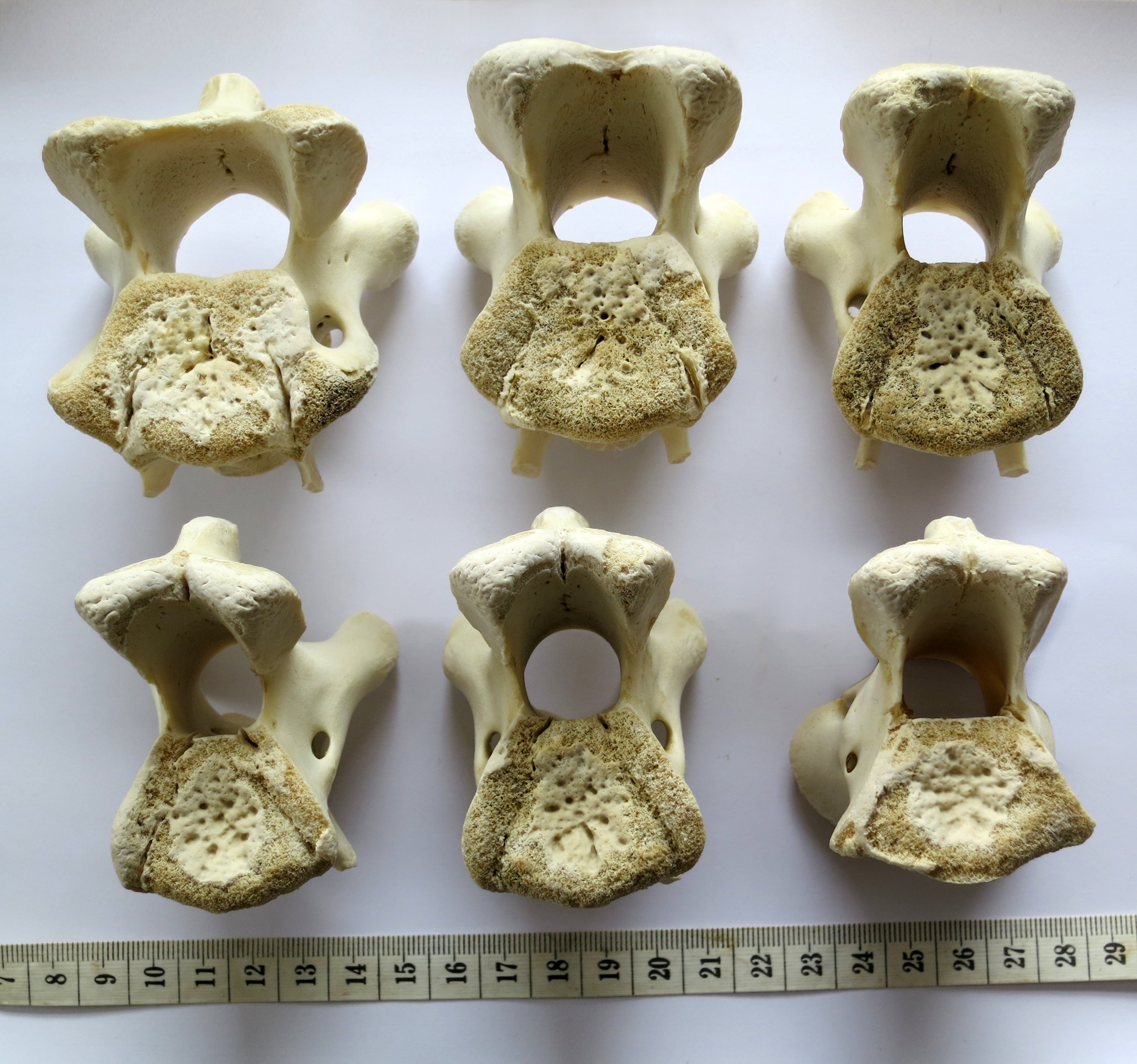
Articular facets of vertebrae of a baby giraffe
Cervical vertebrae of a baby giraffe, Giraffa camelopardalis, in posterior view. Top row, left to right: cervicals 7, 6 and 5; bottom row, left to right: cervicals 4, 3 and 2. Despite changes in the vertebrae along the column, the flattened pentagon shape of the articular facets remains similar along the sequence. (Note that extensive cartilage caps existed on the articular facets of this very young specimen, but were lost in preparation.) Photograph by the author, of a specimen in his personal collection.
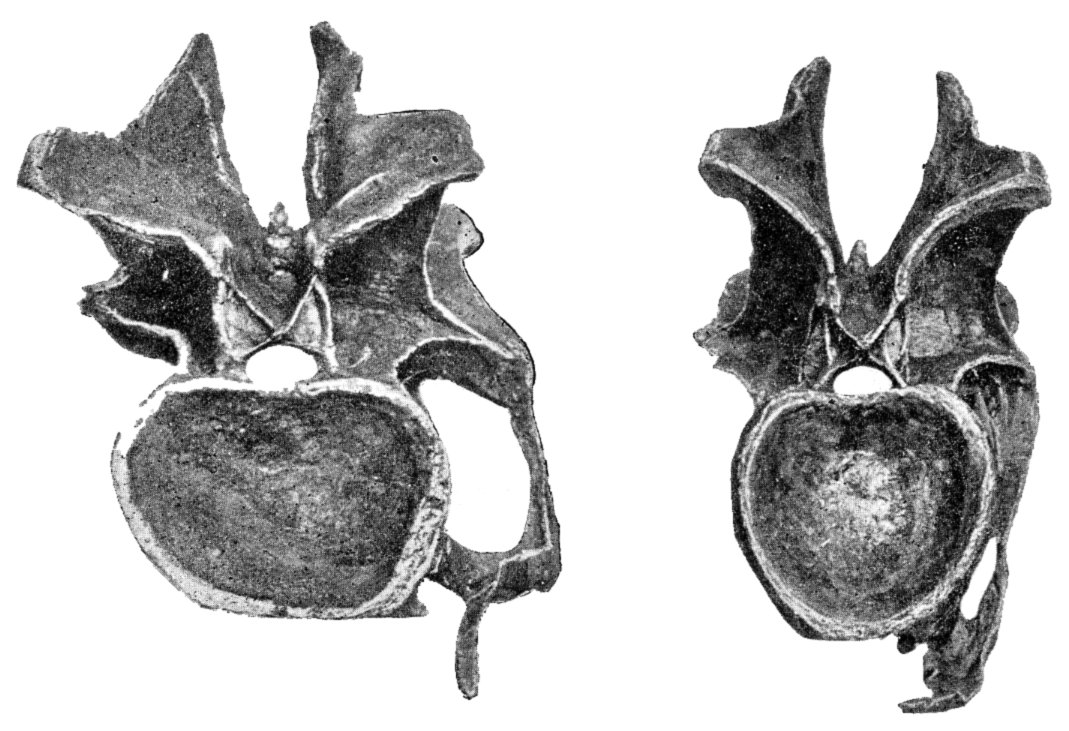
Distorted cotyles of Giraffatitan brancai vertebrae
Cervical vertebrae 4 (left) and 6 (right) of Giraffatitan brancai lectotype MB.R.2180 (previously HMN SI), in posterior view. Note the dramatically different aspect ratios of their cotyles, indicating that extensive and unpredictable crushing has taken place. Photographs by author.
Distorted cotyles of Diplodocus carnegii vertebrae
Cervical vertebrae 14 (left) and 13 (right) of Diplodocus carnegii holotype CM 84, in posterior view. Note the dramatically different aspect ratios of their cotyles, indicating that extensive and unpredictable crushing has taken place.
Manipulation of consecutive sauropod vertebrae
Manipulation of consecutive sauropod vertebrae by hand. Cervicals 2 and 3 of Giraffatitan brancai lectotype MB.R.2181 (formerly HMN SI). I attempted to articulate these two vertebrae, and empirically determine the feasible range of motion. Due to subtle distortion of the zygapophyses of these vertebrae, it was not possible to articulate C2 in a more extended position relative to C3 than shown here. Photograph by Mathew J. Wedel.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}