
A new species of Cenopalpus Pritchard & Baker (Acari: Tenuipalpidae) from Japan, with ontogeny of chaetotaxy and a key to the world species
- Published
- Accepted
- Received
- Academic Editor
- Marcio Pie
- Subject Areas
- Agricultural Science, Entomology, Taxonomy, Zoology
- Keywords
- Acarology, Systematics, Acari, Trombidiformes, Prostigmata, Phytophagous, Classification, Pest
- Copyright
- © 2020 Negm et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. A new species of Cenopalpus Pritchard & Baker (Acari: Tenuipalpidae) from Japan, with ontogeny of chaetotaxy and a key to the world species. PeerJ 8:e9081 https://doi.org/10.7717/peerj.9081
Abstract
A new species of flat mite, Cenopalpus umbellatus sp. nov. (Acari: Trombidiformes: Tenuipalpidae) is described and illustrated based on females, males, deutonymphs, protonymphs and larvae. The morphological ontogeny in idiosomal and leg chaetotaxy is briefly described for all stages. Mite specimens were collected from the leaves of Rhaphiolepis indica var. umbellata Makino (Rosaceae), an evergreen shrub native to Japan. An identification key to the world species of Cenopalpus is also provided.
Introduction
Mites of the family Tenuipalpidae Berlese, 1913 (Acari: Trombidiformes) are harmful pests to a wide range of plants (Jeppson, Keifer & Baker, 1975; Mesa et al., 2009). The genus Cenopalpus Pritchard & Baker, 1958, currently contains 70 species (including the present new species), mostly described from Palearctic and Afrotropical ecozones (Table 1). Mesa et al. (2009) listed the genus Cenopalpus with 60 species, assigning the two species, salignae (Meyer, 1979) and thelycraniae (Livschitz & Mitrofanov, 1967), under Brevipalpus. Later, Saccaggi et al. (2017) cited B. salignae in the genus Cenopalpus, however, the Russian species (B. thelycraniae) was already transferred to Cenopalpus by Mitrofanov & Strunkova (1979). Also, C. iqbali Iqbal, Akbar & Ali, 2007, was not included in Mesa et al. (2009).
| Species | Country | |
|---|---|---|
| 1 | abaii Khosrowshahi & Arbabi, 1997 | Iran |
| 2 | adventicius Ueckermann & Ripka, 2015 | Hungary |
| 3 | aratus Chaudhri, 1971 | Pakistan |
| 4 | arbuti Hatzinikolis & Emmanouel, 1987 | Greece |
| 5 | bagdasariani (Livschitz & Mitrofanov, 1970) | Tajikistan |
| 6 | bakeri Düzgünes, 1967 | Turkey |
| 7 | brachypalpus Hatzinikolis, Panou & Papadoulis, 1999b | Greece |
| 8 | capacis Chaudhri, 1971 | Pakistan |
| 9 | capensis (Meyer, 1979) | South Africa |
| 10 | carpini (Livschitz & Mitrofanov, 1967) | Ukraine |
| 11 | chitraliensis Akbar & Chaudhri, 1985 | Pakistan |
| 12 | crataegi Dosse, 1971 | Iran |
| 13 | creticus Hatzinilkolis, Papadoulis & Panou, 1999a | Greece |
| 14 | cumanicus Ueckermann & Ripka, 2015 | Hungary |
| 15 | dignus Akbar & Chaudhri, 1985 | Pakistan |
| 16 | eriobotryi Hatzinikolis, 1969 | Greece |
| 17 | evini Khosrowshahi, 1991 | Iran |
| 18 | favosus Chaudhri, 1971 | Pakistan |
| 19 | halperini Castagnoli, 1987 | Israel |
| 20 | haqii Akbar & Chaudhri, 1985 | Pakistan |
| 21 | hederae Papaioannou-Souliotis, 1986 | Greece |
| 22 | homalos Akbar & Chaudhri, 1985 | Pakistan |
| 23 | iqbali Iqbal, Akbar & Ali, 2007 | Pakistan |
| 24 | irani Dosse, 1971 | Iran |
| 25 | japonicus Hasan, Akbar & Khalid, 2001 | Pakistan |
| 26 | khosrowshahii Khanjani et al., 2012 | Iran |
| 27 | kritos Hasan et al., 2004 | Pakistan |
| 28 | lanceolatisetae (Attiah, 1956) | Egypt |
| 29 | limbatus Akbar & Chaudhri, 1985 | Pakistan |
| 30 | lineola (Canestrini & Fanzago, 1876) | Italy |
| 31 | longirostris (Livschitz & Mitrofanov, 1967) | Ukraine |
| 32 | mespili (Livschitz & Mitrofanov, 1967) | Ukraine |
| 33 | meyerae Khosrowshahi, 1991 | Iran |
| 34 | mughalii Akbar & Aheer, 1990 | Pakistan |
| 35 | musai Dosse, 1975 | Lebanon |
| 36 | natalensis (Lawrence, 1943) | South Africa |
| 37 | naupakticus Hatzinikolis, Panou & Papadoulis, 1999b | Greece |
| 38 | officinalis Papaioannou-Souliotis, 1986 | Greece |
| 39 | oleunus (Meyer, 1979) | South Africa |
| 40 | orakiensis Akbar & Chaudhri, 1985 | Pakistan |
| 41 | pegazzanoae Castagnoli, 1987 | Italy |
| 42 | pennatisetis (Wainstein, 1958) | Kazakhistan |
| 43 | picitilis Chaudhri, 1971 | Pakistan |
| 44 | piger Wainstein, 1960 | Kazakhistan |
| 45 | pistaciae Hatzinilkolis, Papadoulis & Panou, 1999a | Greece |
| 46 | platani (Livschitz & Mitrofanov, 1967) | Georgia |
| 47 | populi (Livschitz & Mitrofanov, 1967) | Georgia |
| 48 | pritchardi Düzgünes, 1967 | Turkey |
| 49 | prunusi Khanjani et al., 2012 | Iran |
| 50 | pseudospinosus (Livschitz & Mitrofanov, 1967) | Ukranie |
| 51 | pterinus Pritchard & Baker, 1958 | Spain |
| 52 | pulcher (Canestrini & Fanzago, 1876) | Italy |
| 53 | quadricornis (Livschitz & Mitrofanov, 1967) | Armenia |
| 54 | quercusi Khanjani et al., 2012 | Iran |
| 55 | ramus Manson, 1963 | Pakistan |
| 56 | ruber Wainstein, 1960 | Tajikistan |
| 57 | rubusi Khanjani et al., 2012 | Iran |
| 58 | salignae (Meyer, 1979) | South Africa |
| 59 | saryabiensis Akbar & Chaudhri, 1985 | Pakistan |
| 60 | scoopsetus Hatzinikolis & Papadoulis, 1999 | Greece |
| 61 | spinosus (Donnadieu, 1875) | France |
| 62 | sunniensis Hasan et al., 2004 | Pakistan |
| 63 | tamarixi (Nassar & Kandeel)—Zaher (1984) | Egypt |
| 64 | taygeticus Hatzinikolis, Panou & Papadoulis, 1999b | Greece |
| 65 | thelycraniae (Livschitz & Mitrofanov, 1967) | Ukraine |
| 66 | umbellatus sp. nov. Negm, Ueckermann & Gotoh | Japan |
| 67 | viniferus Hatzinikolis, Papadoulis & Kapaxidi, 2001 | Greece |
| 68 | virgulatus Akbar & Chaudhri, 1985 | Pakistan |
| 69 | wainsteini (Livschitz & Mitrofanov, 1967) | Ukraine |
| 70 | xini Ma & Li, 1984 | China |
Note:
In Japan, comparing to spider mites (Tetranychidae), few studies have been done on the taxonomy of tenuipalpid mites. It is expected that several localities are most likely to hold undiscovered species. Ehara & Gotoh (2009) listed 14 species of flat mites from Japan, belonging to the genera Aegyptobia Sayed, Brevipalpus Donnadieu, Cenopalpus, Dolichotetranychus Sayed, Pentamerismus McGregor and Tenuipalpus Donnadieu, with only one species of Cenopalpus (C. lineola; Table 2). Therefore, the present work aimed to increase our knowledge about the tenuipalpid mite fauna in Japan through describing a new species of Cenopalpus. Since immature stages of mites can provide valuable information for better mite systematics, we have described all stages of the new species, with remarks on their ontogenetic changes. Also, an identification key to the world species of Cenopalpus is provided.
| Species | Reference |
|---|---|
| Aegyptobia arenaria Ehara, 1982 | Ehara (1982) |
| Brevipalpus californicus (Banks, 1904) | Ehara (1962) |
| B. lewisi McGregor, 1949 | Ehara (1956b) |
| B. obovatus Donnadieu, 1875a | Ehara (1956a) |
| B. phoenicis (Geijskes, 1939) | Ehara (1966) |
| B. russulus (Boisduval, 1867) | Ehara (1968) |
| Cenopalpus lineola (Canestrini & Fanzago, 1876) | Ehara (1966) |
| C. umbellatus sp. nov. Negm, Ueckermann & Gotoh | Present study |
| Dolichotetranychus floridanus (Banks, 1900) | Baker & Pritchard (1956) |
| D. zoysiae Ehara, 2004 | Ehara (2004) |
| Pentamerismus oregonensis McGregor, 1949 | Ehara (1962) |
| P. taxi (Haller, 1877) | Ehara (1962) |
| Tenuipalpus boninensis Ehara, 1982 | Ehara (1982) |
| T. pacificus Baker, 1945 | Ehara & Ohkubo (1992) |
| T. zhizhilashviliae Reck, 1953b | Ehara (1956b) |
Notes:
Materials and Methods
Mite collection, examination and slide preparations were conducted as previously described in Negm & Gotoh (2019). Measurements (in micrometres) were done using the imaging software Sensiv Measure® ver. 2.6.0 and were presented for the holotype specimen then followed by the range for paratypes in parentheses. The terminology and abbreviations used in the description of the new species follows that of Lindquist (1985) and Mesa et al. (2009). Leg chaetotaxy is adapted from Lindquist (1985) and Seeman & Beard (2011). Several taxonomic keys to Cenopalpus species have been used in the present study, mostly regional (Wainstein, 1960 (Kazakhstan); Livschitz & Mitrofanov, 1967 (USSR); Zaher & Yousef, 1969, Zaher, 1984 (Egypt); Meyer, 1979 (World); Akbar & Chaudhri, 1985 (Pakistan); Hatzinikolis & Emmanouel, 1987; Hatzinilkolis, Papadoulis & Panou, 1999a; Hatzinikolis, Panou & Papadoulis, 1999b (Greece); Khosrowshahi & Arbabi, 1997; Khanjani et al., 2012 (Iran); Çobanoğlu, Ueckermann & Sağlam, 2016; Çobanoğlu, Erdoğan & Kılıç, 2019 (Turkey)).
Nomenclatural Acts. The electronic version of this article in Portable Document Format will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank the online registration system for the ICZN. The ZooBank Life Science Identifiers (LSIDs) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:268B04C7-028B-4C03-8F6C-930035941B89, and the LSID for the new species, Cenopalpus umbellatus is urn:lsid:zoobank.org:act:957E754C-A7F0-4081-A814-48115D276F76. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Results
Family Tenuipalpidae Berlese, 1913
Cenopalpus Pritchard & Baker, 1958
Cenopalpus umbellatus sp. nov.
[Japanese name: Sharimbai-himehadani]
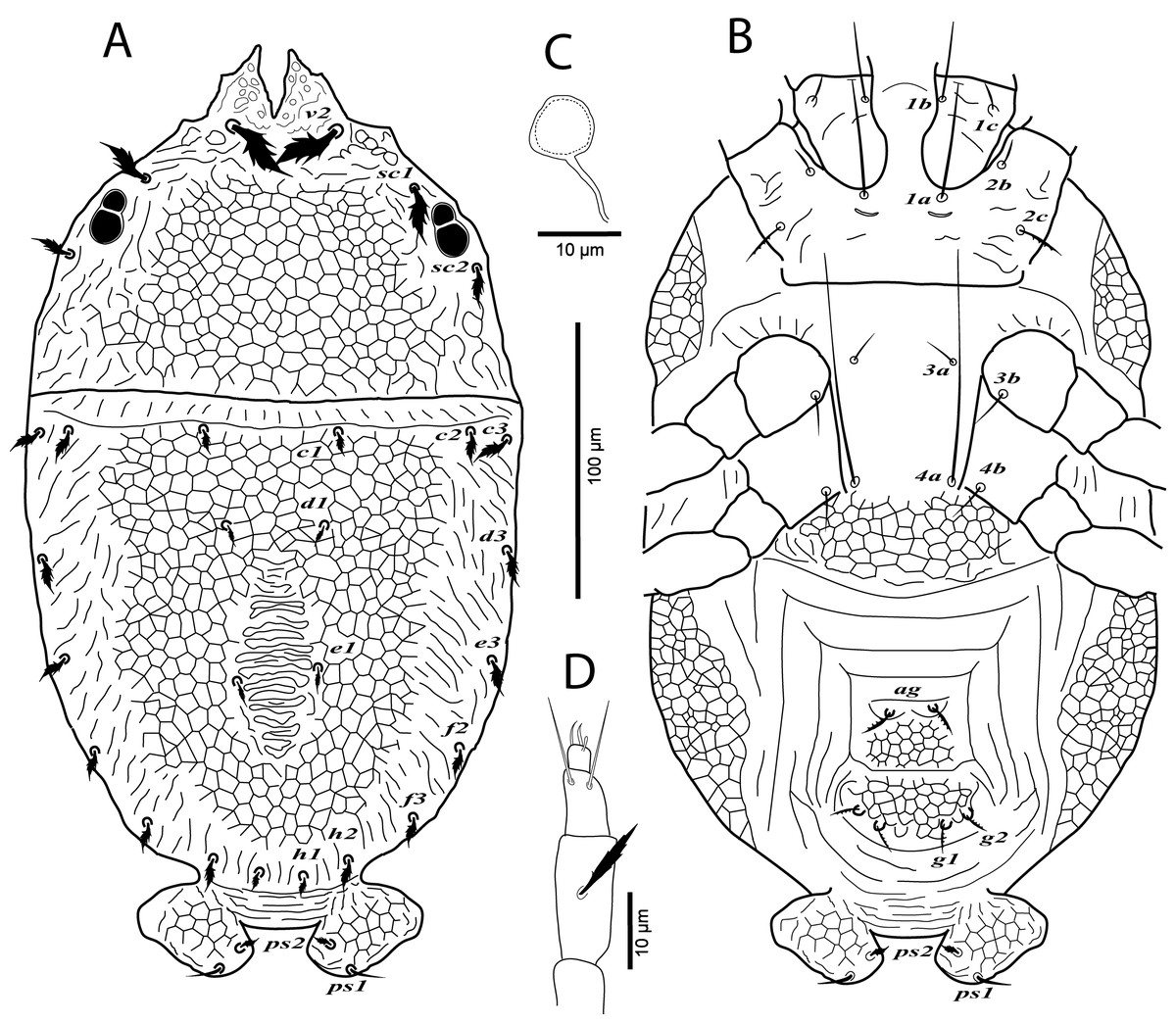
Figure 1: Cenopalpus umbellatus sp. nov.
Female, (A) dorsum, (B) venter, (C) spermatheca, (D) palp. (Image credit: Mohamed Waleed Negm).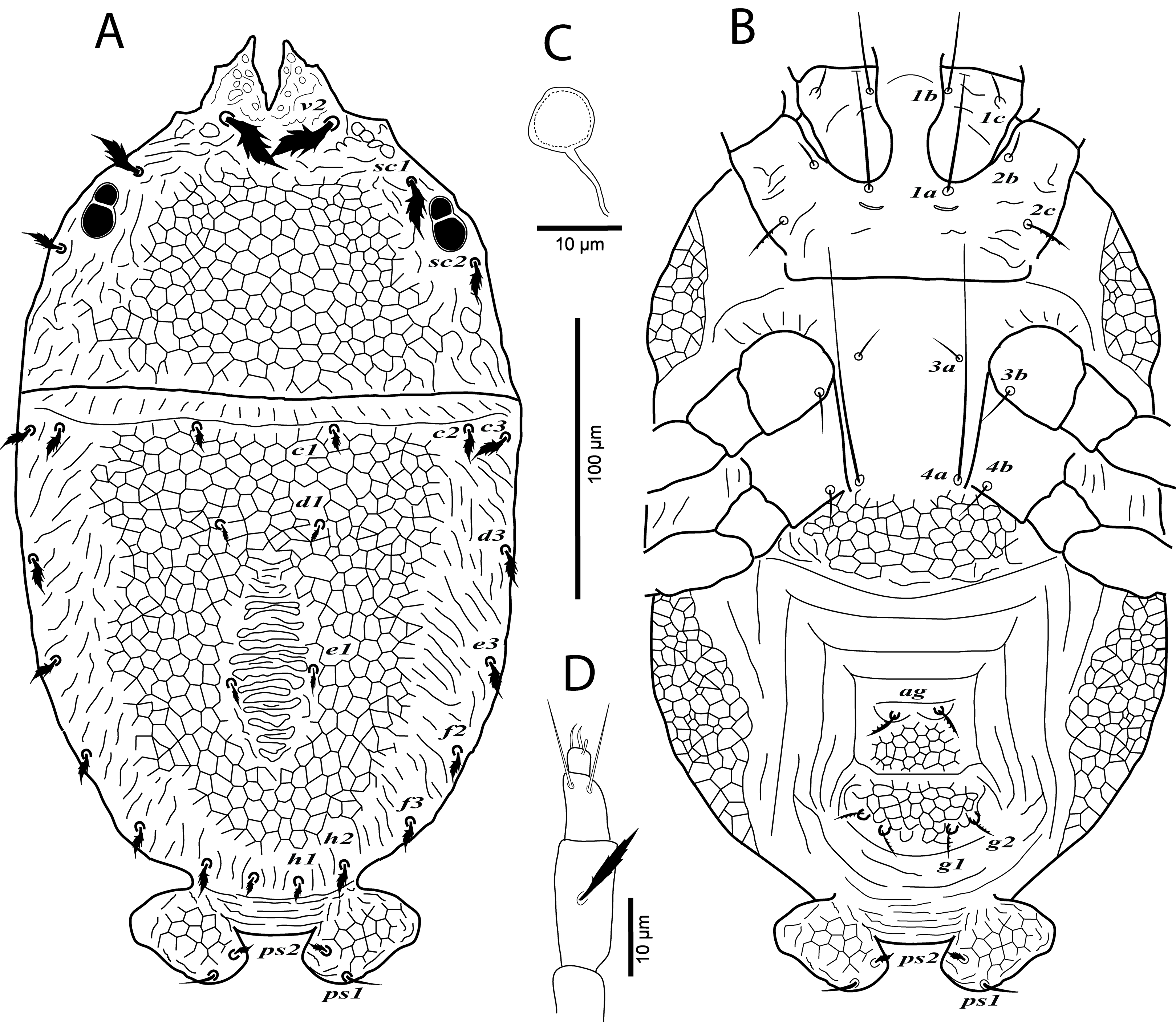{kind=link}
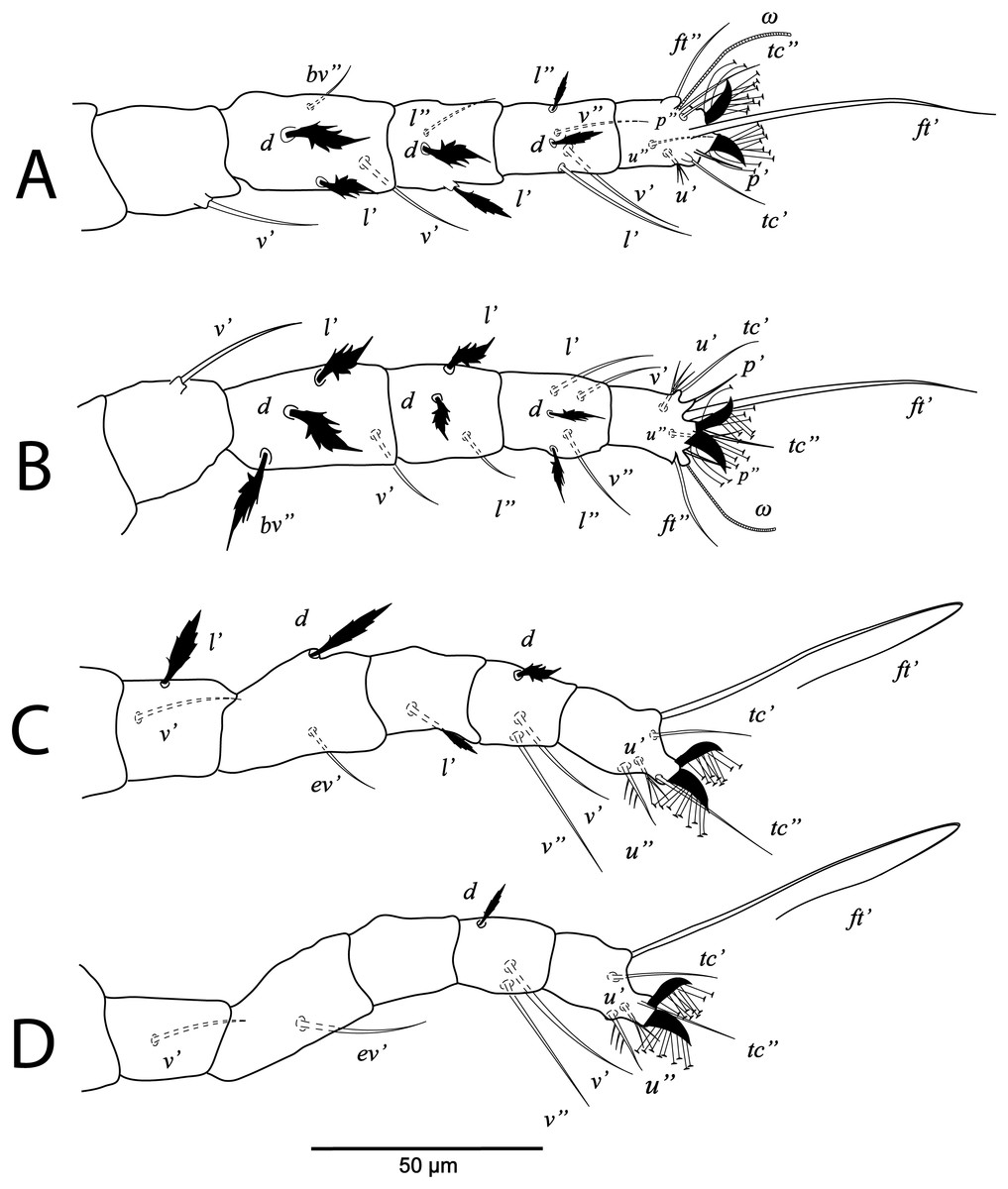
Figure 2: Cenopalpus umbellatus sp. nov.
Female, (A) leg I (left), (B) leg II (right), (C) leg III (right), (D) leg IV (right). (Image credit: Mohamed Waleed Negm).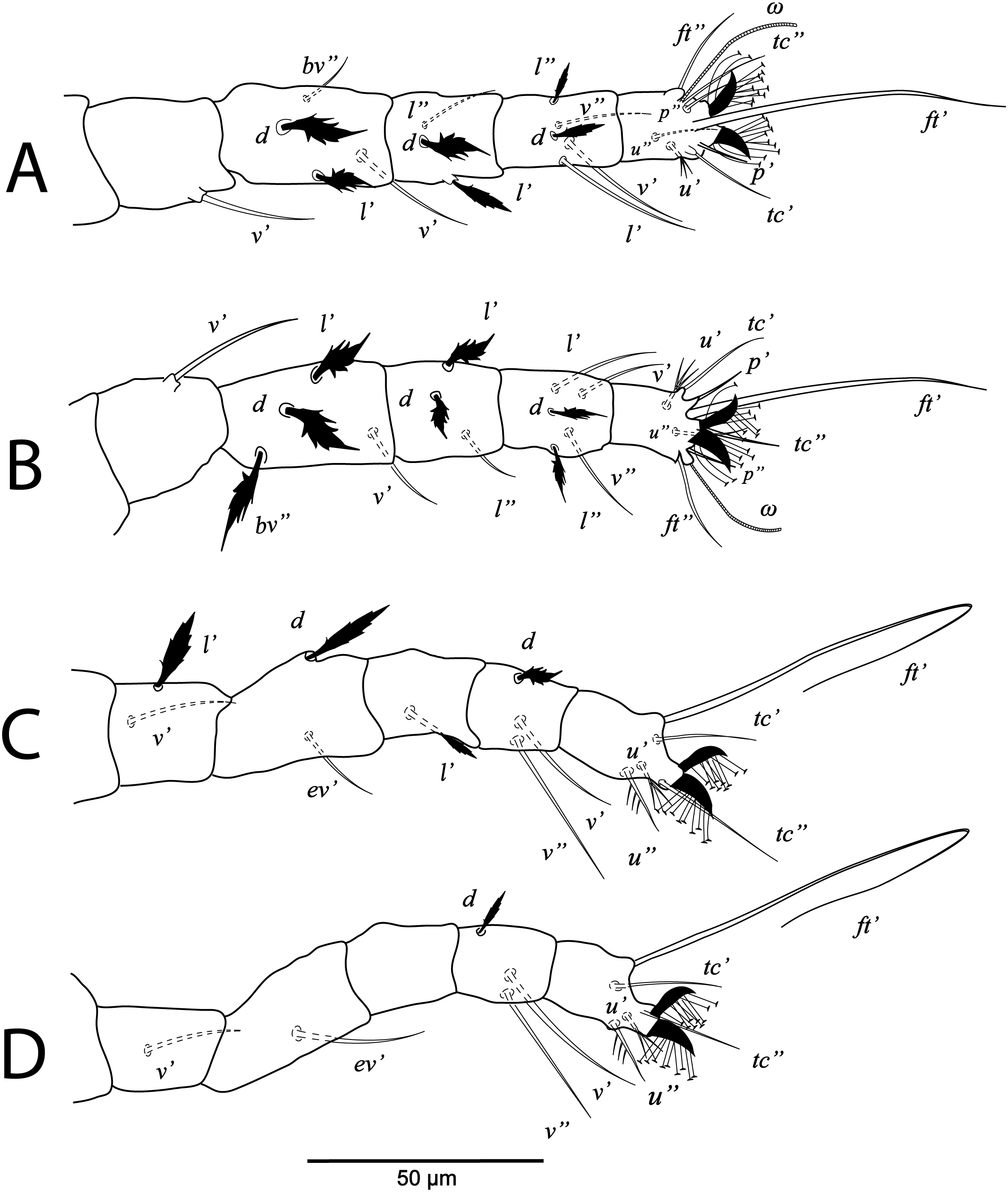{kind=link}
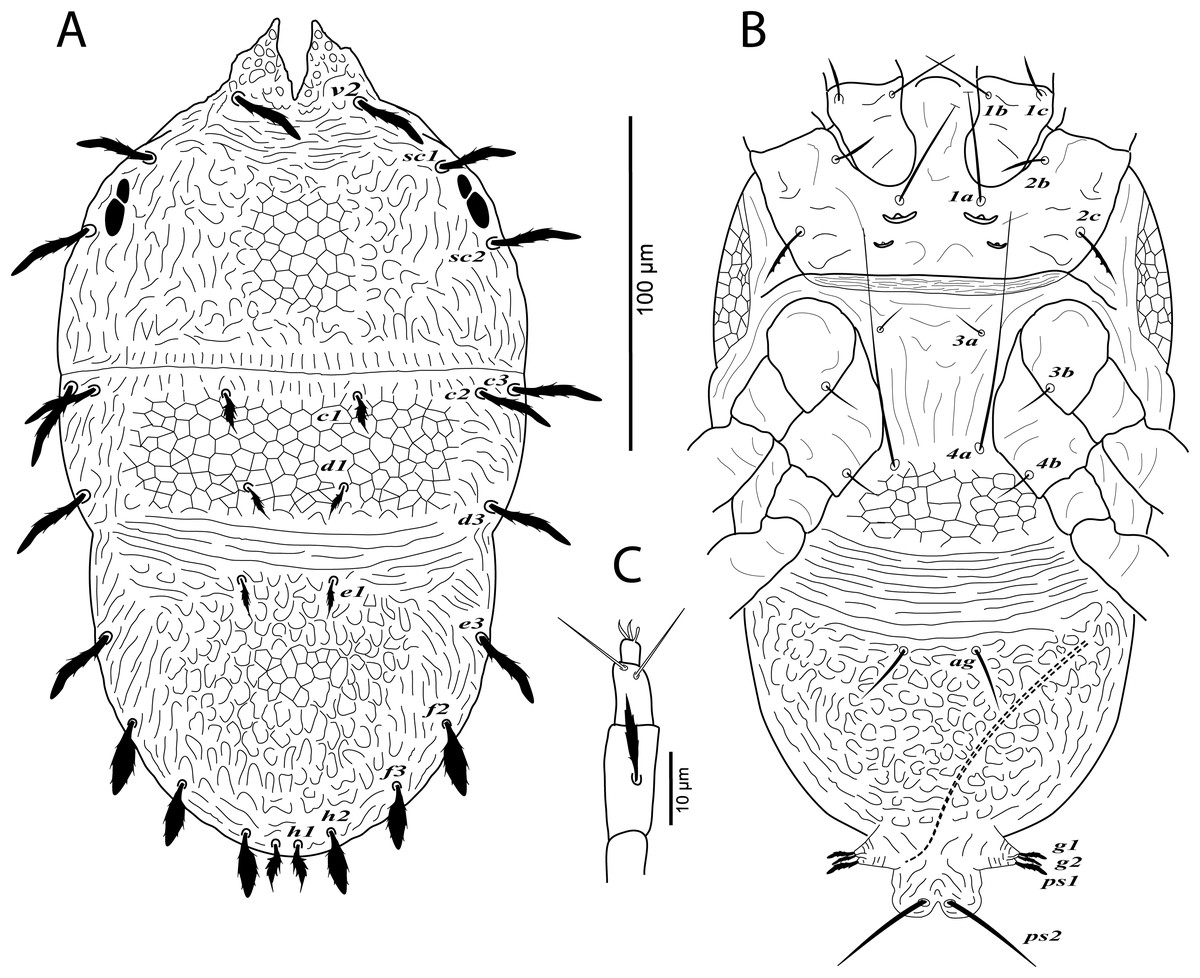
Figure 3: Cenopalpus umbellatus sp. nov.
Male, (A) dorsum, (B) venter, (C) palp. (Image credit: Mohamed Waleed Negm).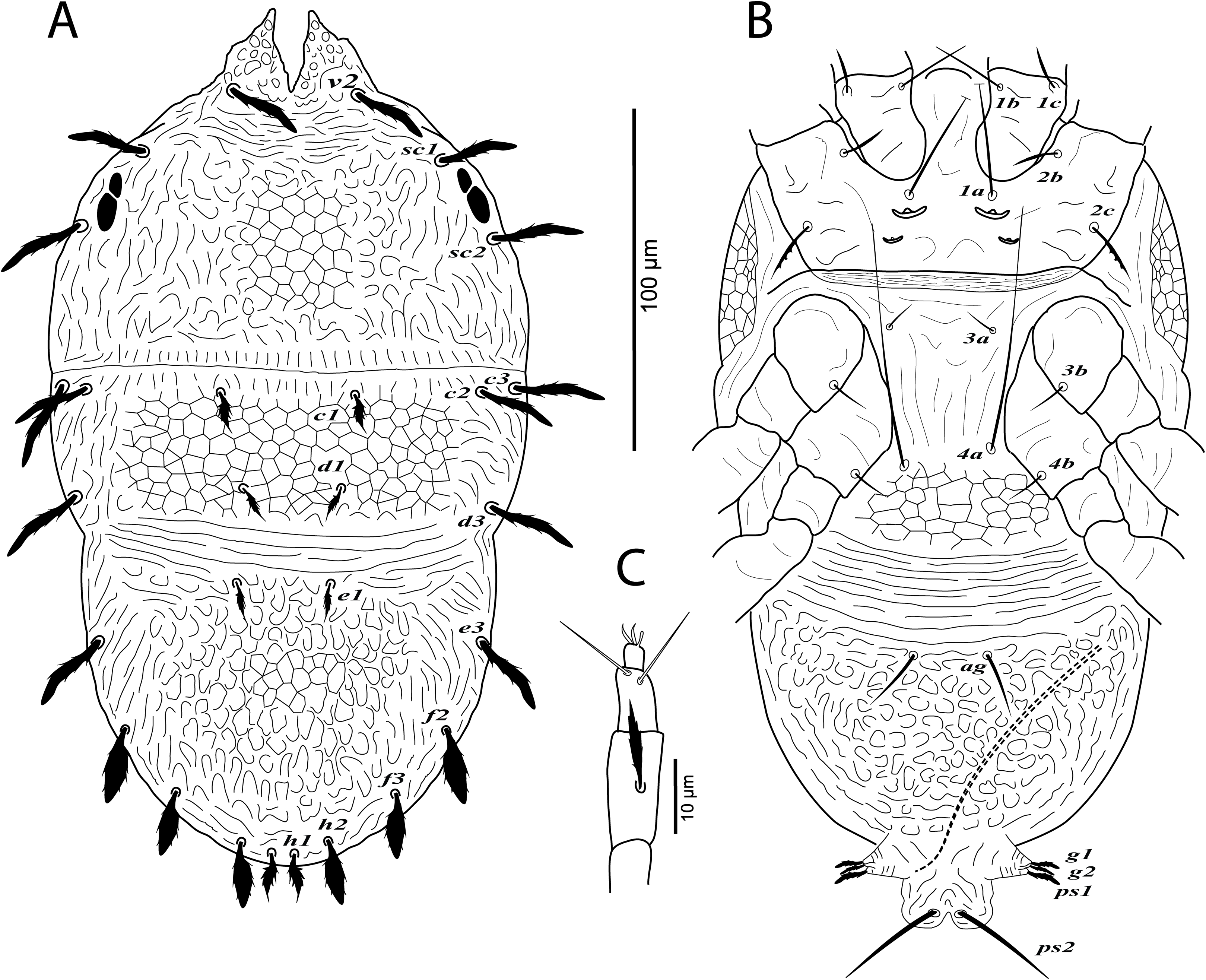{kind=link}

Figure 4: Cenopalpus umbellatus sp. nov.
Male, (A) leg I (left), (B) leg II (right), (C) leg III (right), (D) leg IV (right). (Image credit: Mohamed Waleed Negm).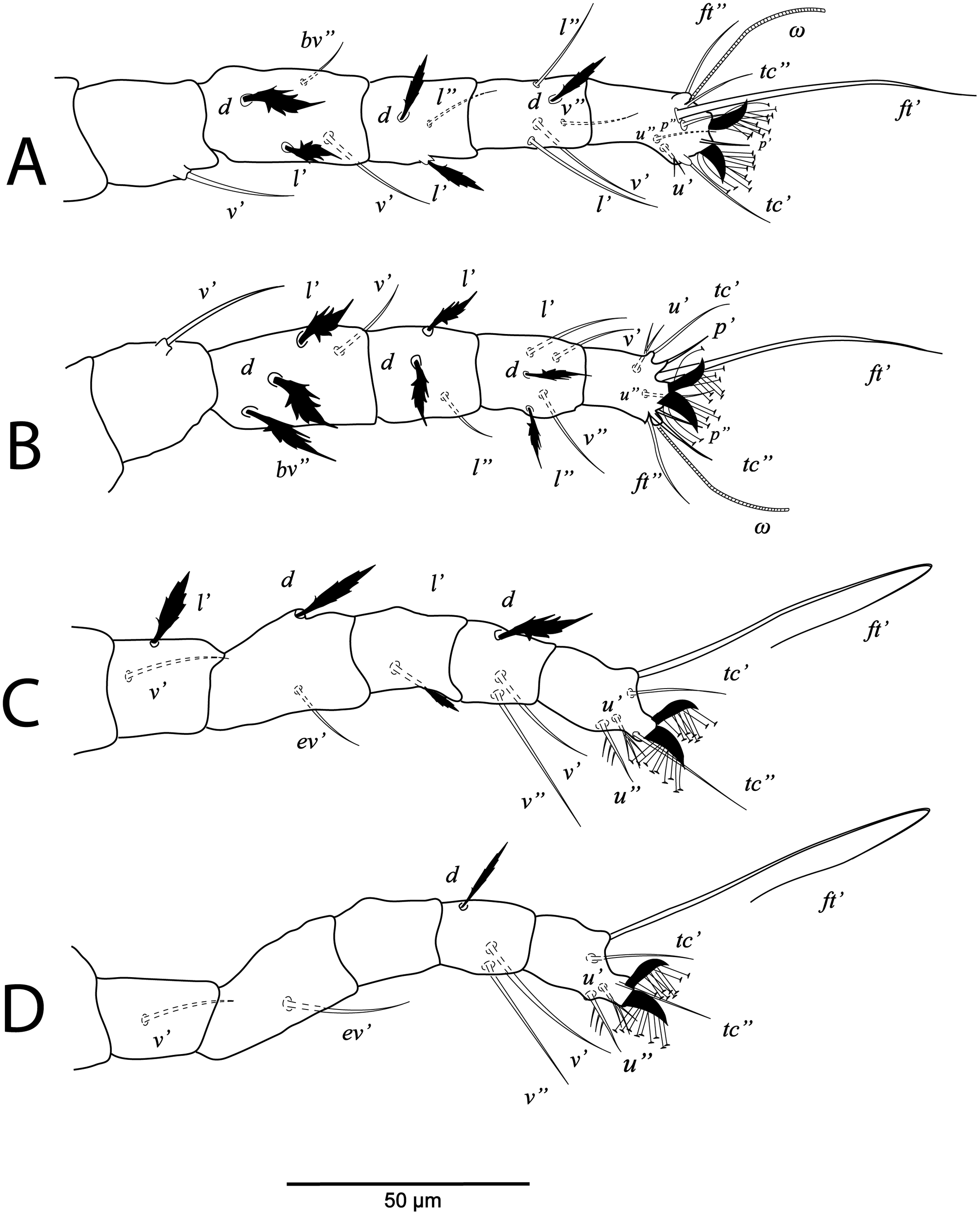{kind=link}
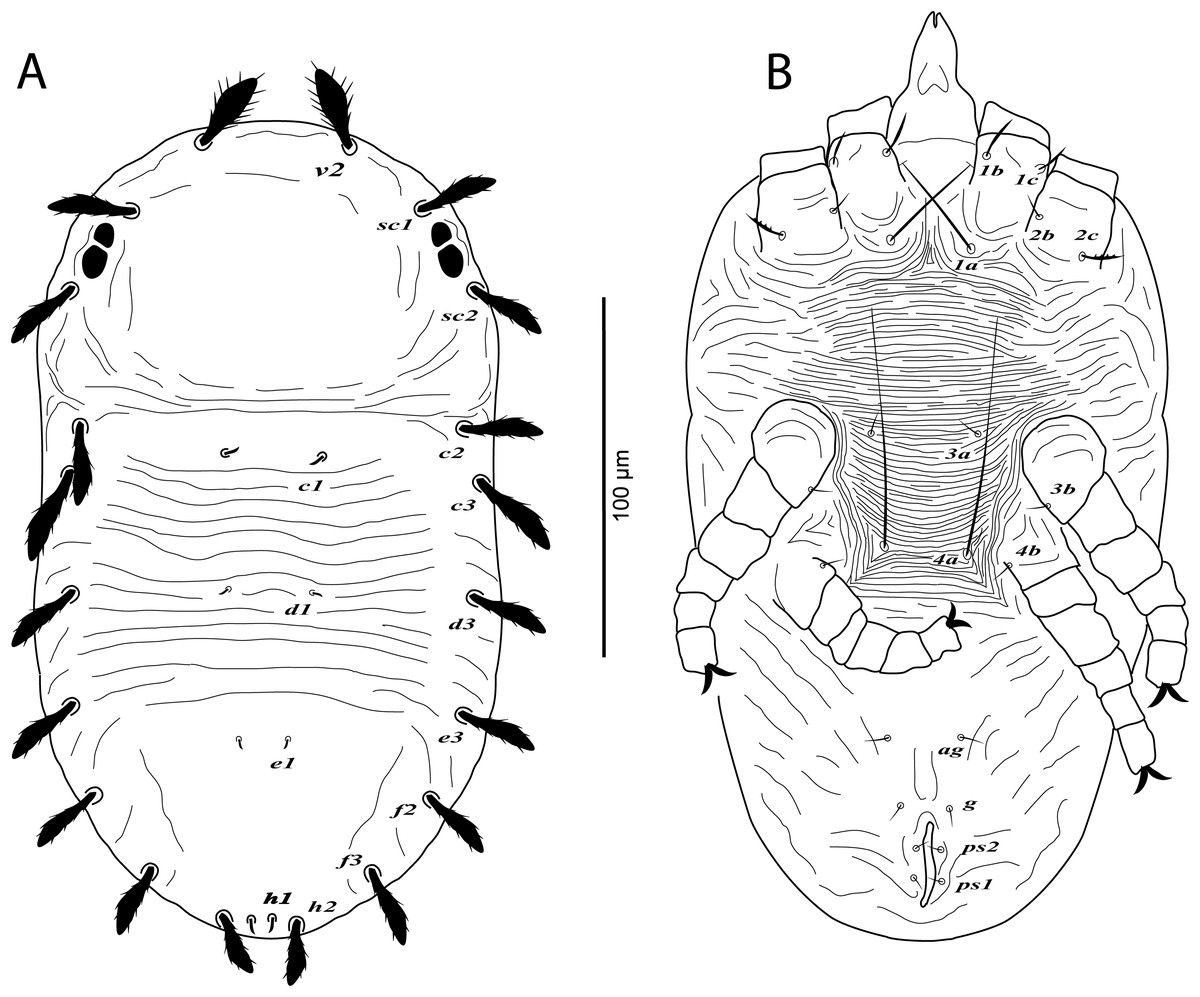
Figure 5: Cenopalpus umbellatus sp. nov.
Deutonymph, (A) dorsum, (B) venter. (Image credit: Mohamed Waleed Negm).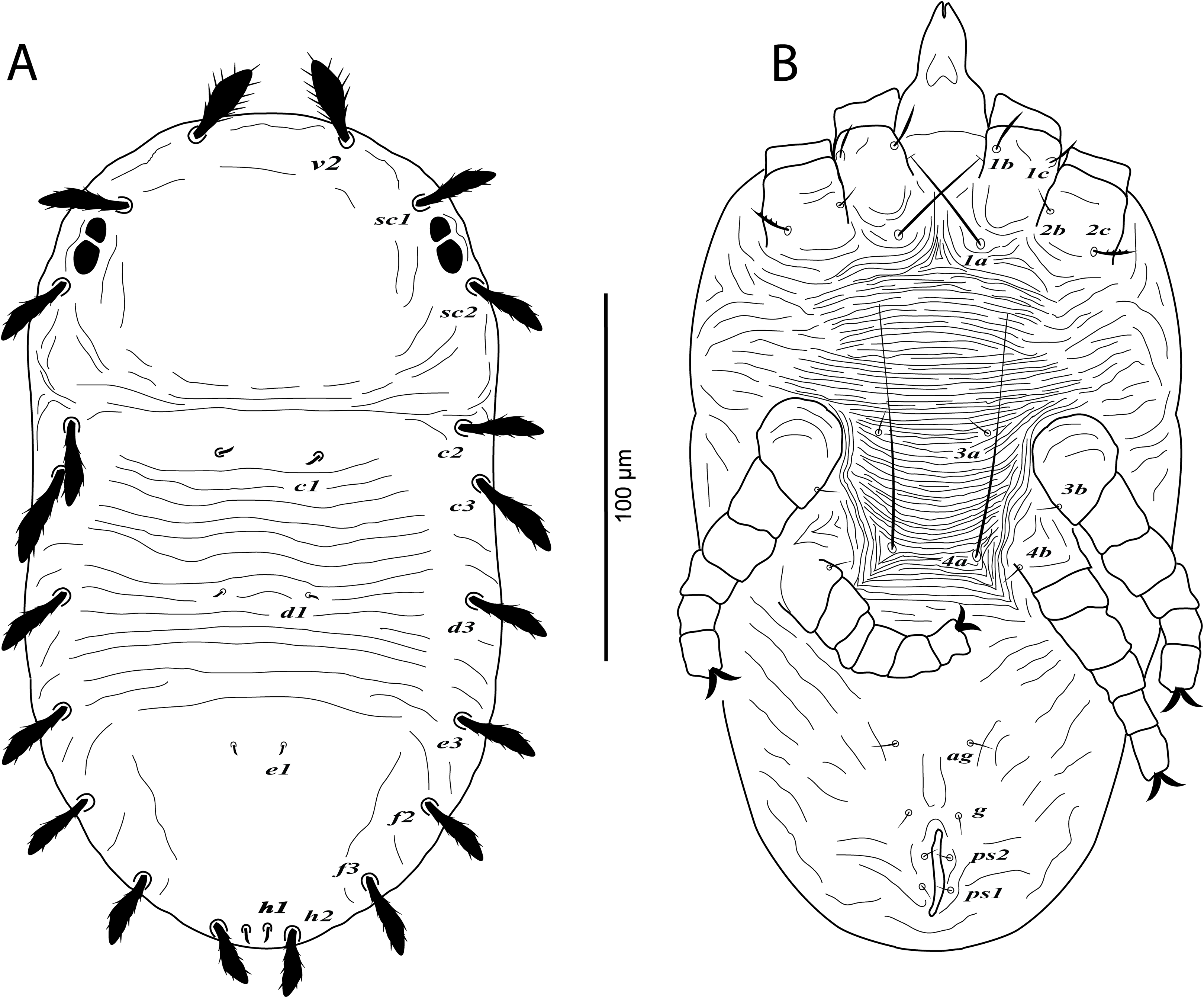{kind=link}
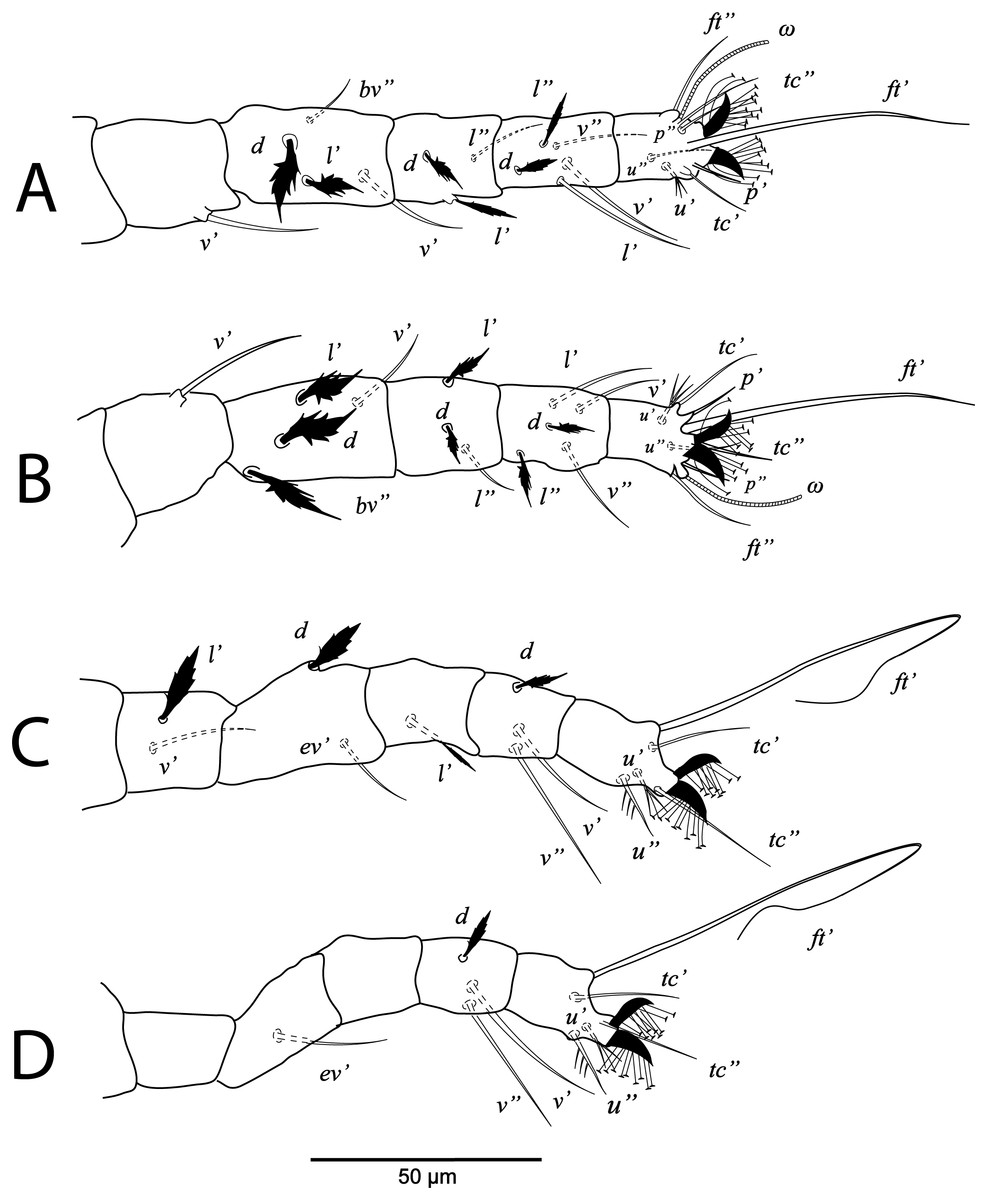
Figure 6: Cenopalpus umbellatus sp. nov.
Deutonymph, (A) leg I (left), (B) leg II (right), (C) leg III (right), (D) leg IV (right). (Image credit: Mohamed Waleed Negm).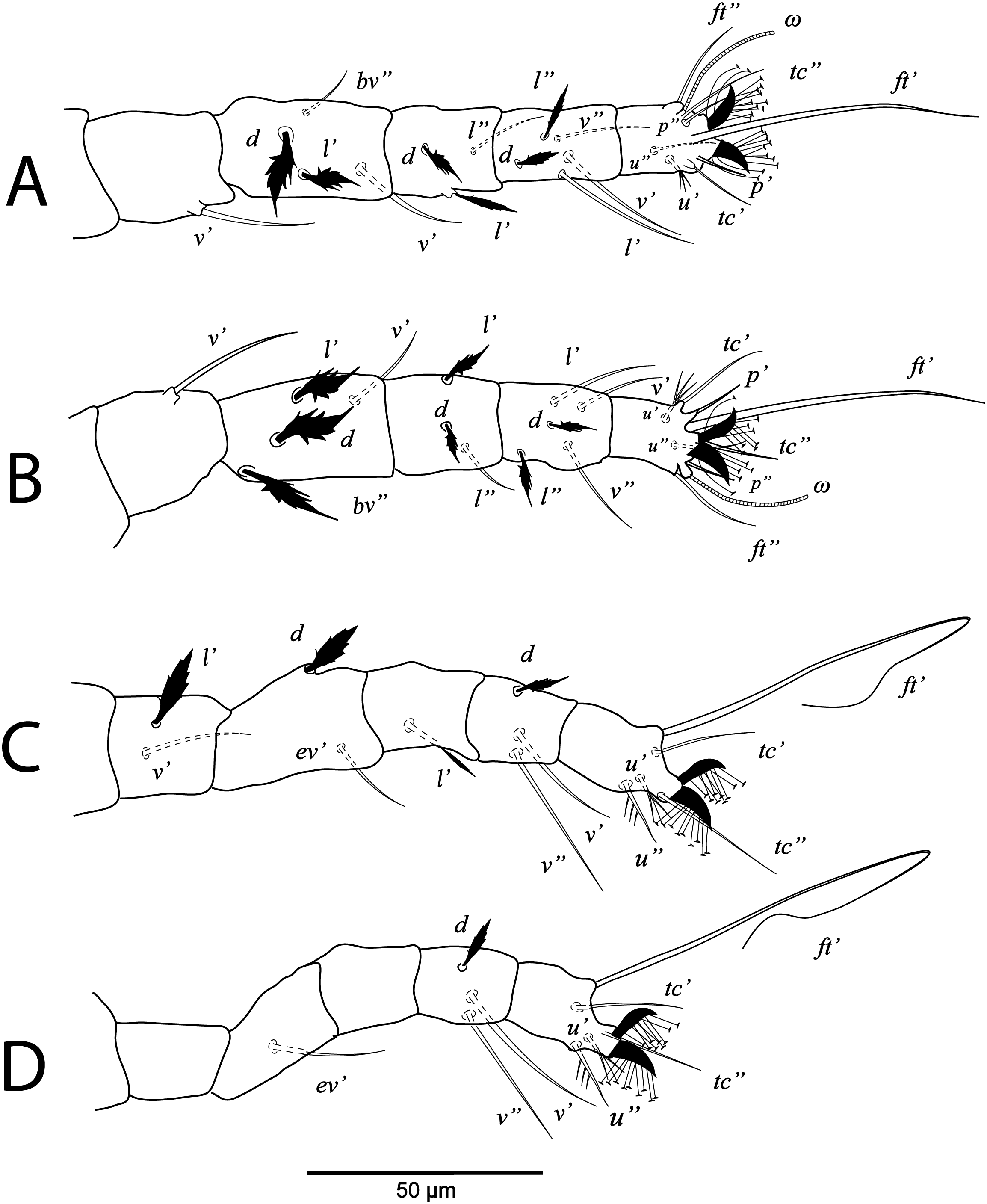{kind=link}
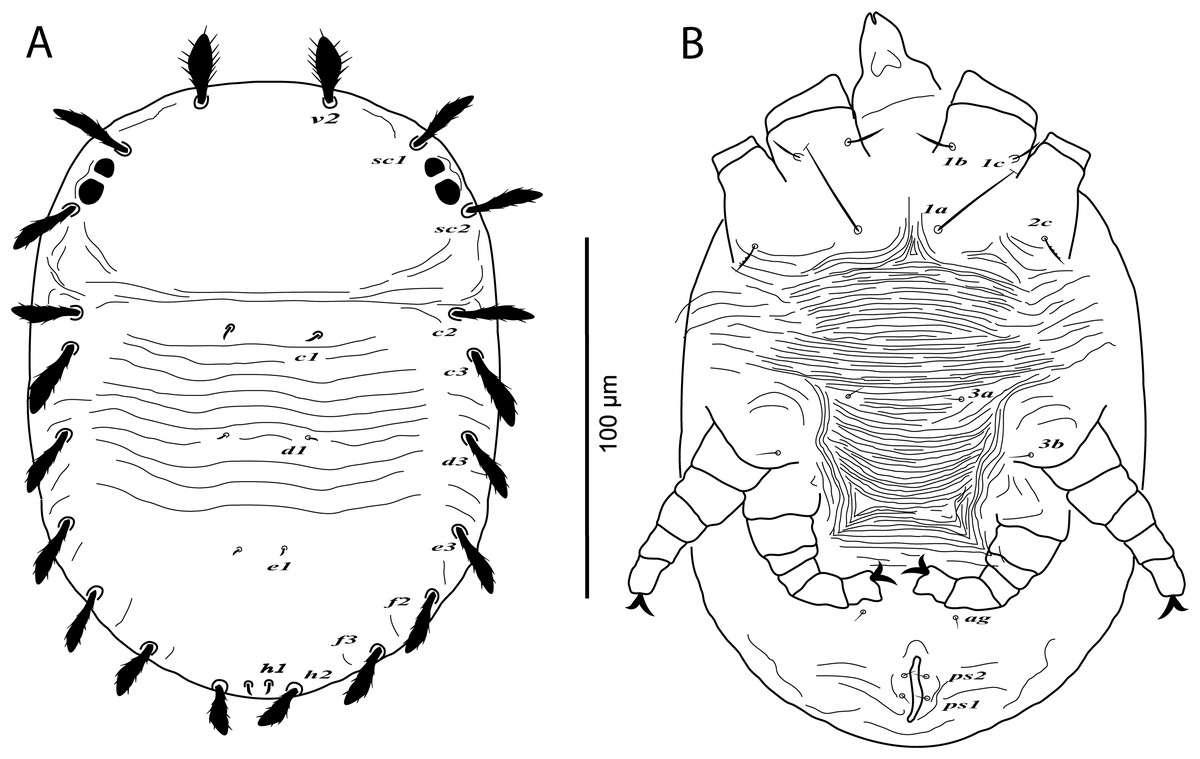
Figure 7: Cenopalpus umbellatus sp. nov.
Protonymph, (A) dorsum, (B) venter. (Image credit: Mohamed Waleed Negm).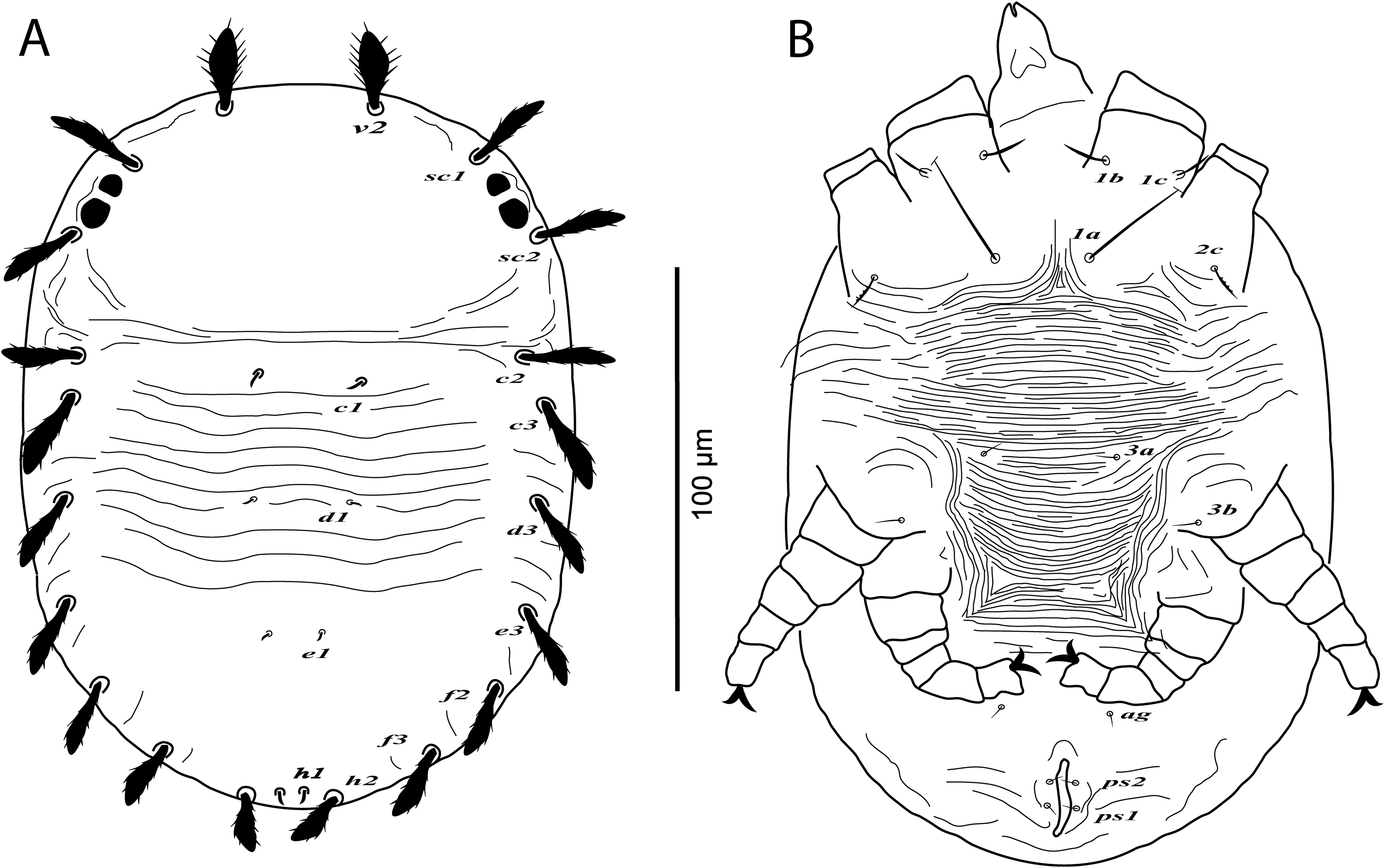{kind=link}
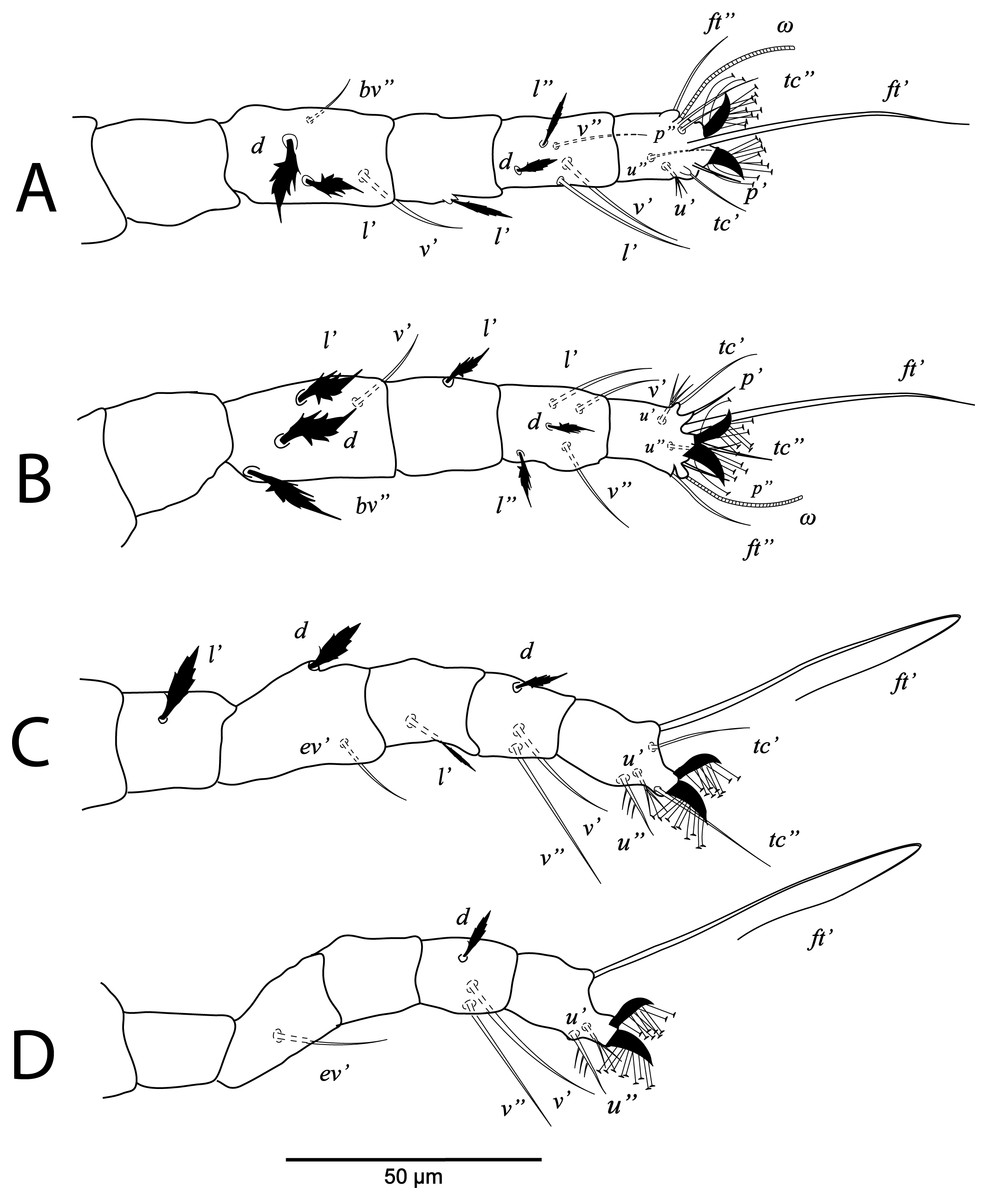
Figure 8: Cenopalpus umbellatus sp. nov.
Protonymph, (A) leg I (left), (B) leg II (right), (C) leg III (right), (D) leg IV (right). (Image credit: Mohamed Waleed Negm).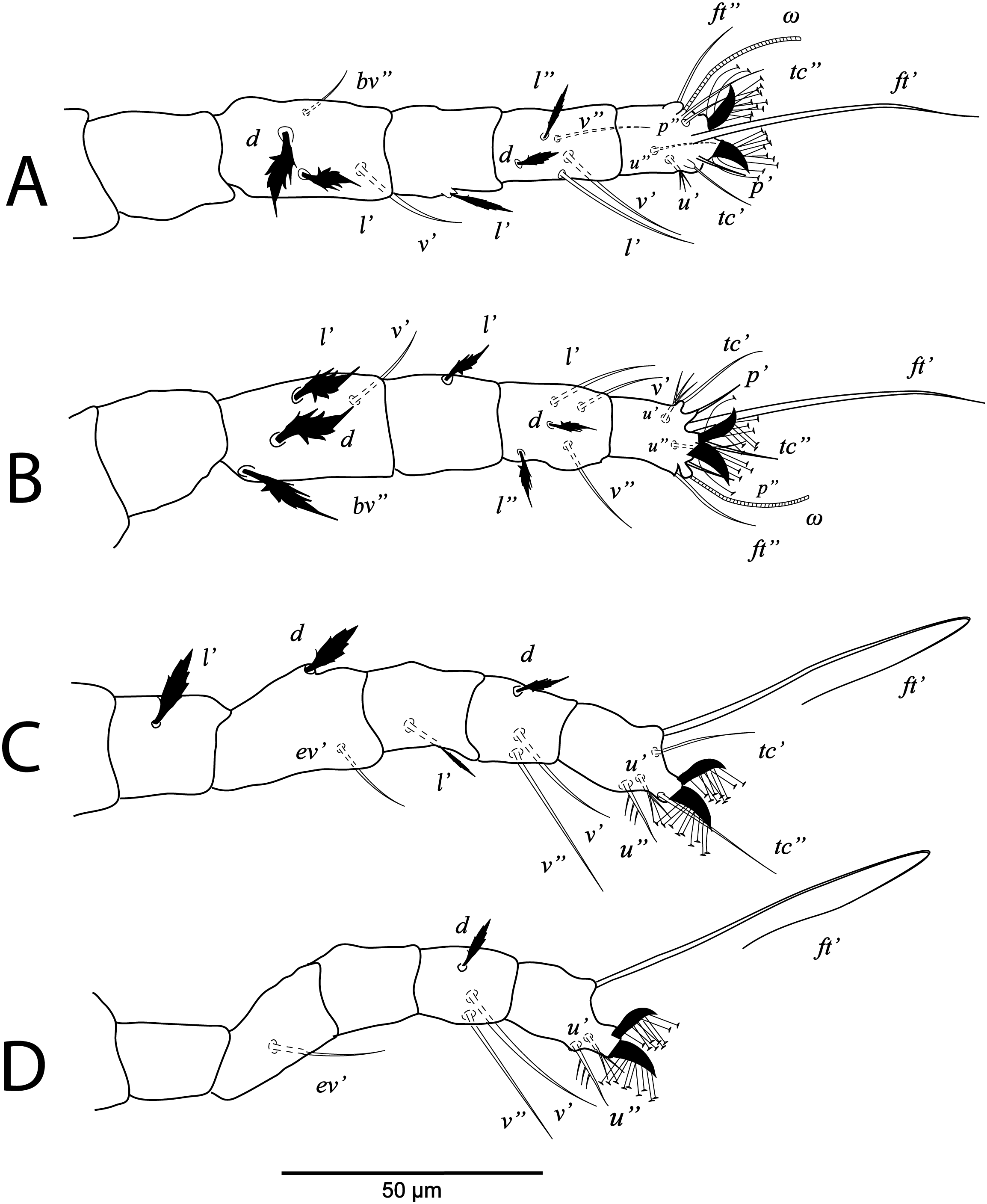{kind=link}

Figure 9: Cenopalpus umbellatus sp. nov.
Larva, (A) dorsum, (B) venter. (Image credit: Mohamed Waleed Negm).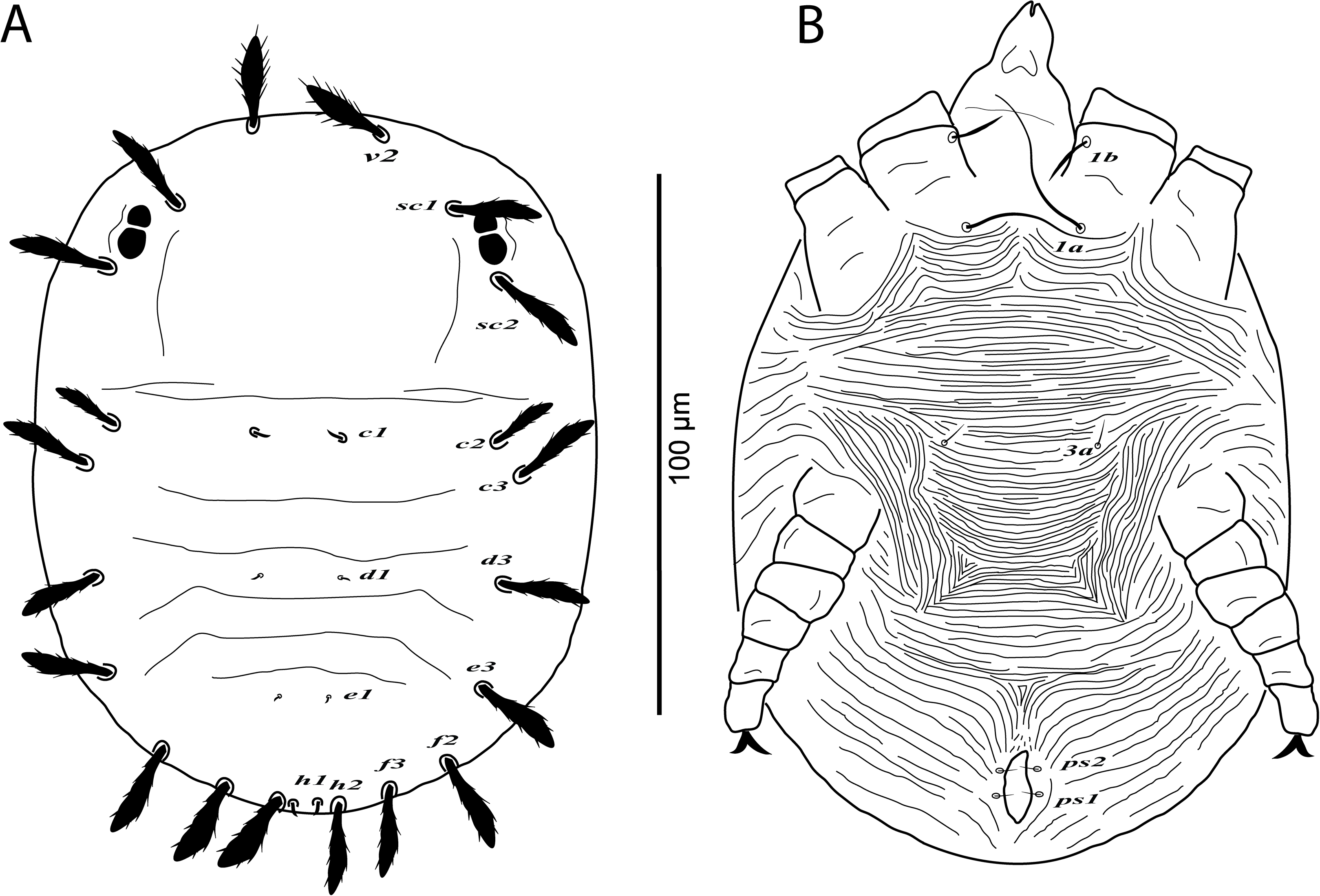{kind=link}
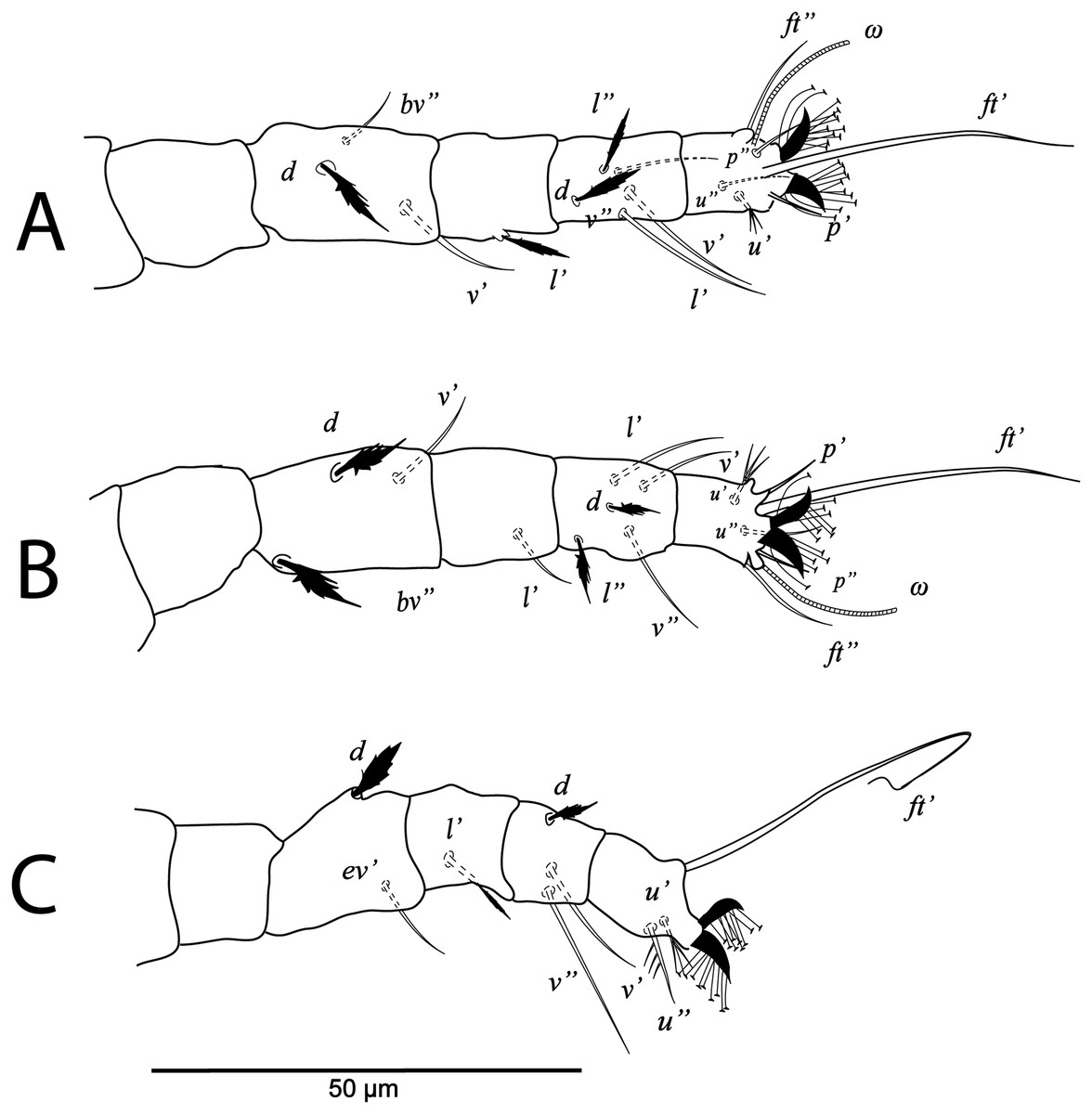
Figure 10: Cenopalpus umbellatus sp. nov.
Larva, (A) leg I (left), (B) leg II (right), (C) leg III (right). (Image credit: Mohamed Waleed Negm).{kind=link}
Description
Female (n = 10)
Dorsum (Fig. 1A). Idiosoma oval, length 300 (278–315), excluding gnathosoma; width 170 (157–174), at level of sejugal furrow. Rostral shield with 2 medial, 2 submedial and 2 lateral lobes; propodosoma regularly reticulated, with few irregular areolae sculpturing laterally; sejugal furrow thick and well defined; opisthosoma mostly reticulated, with few irregular transverse reticulations medially and small irregular areolae laterally; opisthosomal pores absent; propodosomal setae v2 and sc1 broadly lanceolate, serrate, setae sc2 narrowly lanceolate; setae v2 shorter than distance between v2–v2; opisthosomal setae narrowly lanceolate. Lengths of dorsal setae: v2 24 (22–26), sc1 16 (15–17), sc2 13 (12–14), c1 9 (9–11), c2 13 (14–15), c3 17 (16–19), d1 8 (7–8), d3 14 (13–14), e1 7 (6–7), e3 13 (12–14), f2 12 (10–11), f3 11 (11–12), h1 6 (6–7), h2 10 (9–10).
Venter (Fig. 1B). Venter of propodosoma and area between setae 3a and 4a smooth; opisthosomal area behind ventral setae 4a entirely reticulated; coxal seta 2c serrate. ventral shield medially with a reticulation consisting of pentagonal cells; genital shields reticulated with pentagonal cells; genital setae g1 posterior to g2. Lengths of ventral setae: 1a 80 (75–82), 3a 9 (8–10), 4a 70 (65–70); aggenital setae ag 13 (12–14); genital setae g1 10 (10–12), g2 9 (9–11); anal setae ps1 10 (9–10), ps2 8 (8–10). Distances between genital area setae: ag–ag 12–18, g1–g1 21–28, g2–g2 34–40. Spermatheca (n = 3) (Fig. 1C). Spermathecal tube narrow and vesicle semi-circular 8 (8–9) in diameter.
Gnathosoma. Rostrum not reaching distal end of femur I. Palp 4-segmented, palp tarsus with a solenidion and 2 eupathidia, palp tibia with 2 setae, palp femur/genu with 1 lanceolate-serrate dorsal seta (Fig. 1D).
Legs (Figs. 1B and 2A–2D). Chaetotaxy of legs as follows: coxae 2-2-1-1; trochanters 1-1-2-1; femora 4-4-2-1; genua 3-3-1-0; tibiae 5-5-3-3; tarsi 8+ω-8+ω-5-5. Setae d on femora I-III and genua I-II, setae l’ on femora I-II and genua I-II broadly lanceolate-serrate. Setae bv” on femur II and l’ on trochanter III also broadly lanceolate-serrate. Tarsus I and II with solenidia Iω 15–25, IIω 12–18.
Male (n = 10)
Dorsum (Fig. 3A). Idiosoma broadly oval, length 223–238; width 130–140. Rostral shield with 2 medial and 2 slightly shorter submedial lobes; propodosoma regularly reticulated medially, with irregular areolae sculpturing laterally; sejugal furrow distinct; metapodosoma and opisthosoma separated by transverse bands of striae, with irregular reticulations and areolae sculpturing; opisthosomal pores indistinct; propodosomal and lateral setae of opisthosoma long and narrowly lanceolate, serrate; setae v2 shorter than distance between v2–v2. Lengths of dorsal setae: v2 27–28, sc1 24–26, sc2 22–24, c1 12–14, c2 16–18, c3 21–23, d1 9–10, d3 23–26, e1 9–11, e3 23–25, f2 21–24, f3 19–22, h1 10–11, h2 19–21.
Venter (Fig. 3B). Venter of propodosoma and area between setae 3a and 4a slightly striated; opisthosomal area behind ventral setae 4a reticulated, followed by transverse striae posteriorly; coxal seta 2c serrate; ventral shield posterior to setae ag areolate. Lengths of ventral setae: 1a 58–68, 3a 10–12, 4a 55–63; ag 18–20; g1 8–9, g2 9–10; ps1 10–12, ps2 26–28.
Gnathosoma. Rostrum short not reaching distal end of trochanter I. Palp 4-segmented, palp tarsus with a solenidion and 2 eupathidia, palp tibia with 2 setae, palp femur/genu with 1 lanceolate-serrate dorsal seta (Fig. 3C).
Legs (Figs. 3B and 4A–4D). Chaetotaxy of legs as in female. Leg setae also similar to that of female. Tarsus I and II with solenidia Iω 25–30, IIω 20–23.
Deutonymph (n = 6)
Dorsum (Fig. 5A). Idiosoma oval, length 257–266; width 144–162. Rostral shield absent; propodosoma rounded anteriorly, smooth; opisthosoma with transverse striae in the area between setae c1 and e1; opisthosomal pores absent. Dorsal body setae long and narrowly lanceolate except dorsocentral setae c1, d1, e1, h1 minute; setae v2 distinctly shorter than distance between v2–v2. Lengths of dorsal setae: v2 28–30, sc1 26–27, sc2 25–27, c1 4–6, c2 23–25, c3 25–27, d1 2–4, d3 23–25, e1 2–3, e3 22–24, f2 21–23, f3 20–22, h1 4–6, h2 16–18.
Venter (Fig. 5B). Venter of propodosoma and area between setae 1a and 4a with transverse striae; seta 2c serrate; posterior opisthosomal area with irregular striae. Lengths of ventral setae: 1a 42–48, 3a 6–8, 4a 38–45; ag 6–7; g1 4–5; ps1 3–4, ps2 3–4.
Gnathosoma. Palp 4-segmented, palp chaetotaxy as in female.
Legs (Figs. 5B and 6A–6D). Chaetotaxy of legs: coxae 2-2-1-1; trochanters 1-1-2-0; femora 4-4-2-1; genua 3-3-1-0; tibiae 5-5-3-3; tarsi 8+ω-8+ω-5-5. Leg setae similar to that of female.
Protonymph (n = 2)
Dorsum (Fig. 7A). Idiosoma broadly oval, length 164–170; width 106–110. Rostral shield absent; propodosoma rounded anteriorly, smooth; opisthosoma with transverse striae in the area between setae c1 and e1; opisthosomal pores absent. Dorsal body setae long and narrowly lanceolate except dorsocentral setae c1, d1, e1, h1 minute; setae v2 distinctly shorter than distance between v2–v2. Lengths of dorsal setae: v2 21–24, sc1 17–18, sc2 19–21, c1 4–5, c2 16–18, c3 19–20, d1 2–3, d3 17–19, e1 2–3, e3 14–15, f2 15–17, f3 15–16, h1 3–5, h2 12–13.
Venter (Fig. 7B). Venter of idiosoma with transverse striae; posterior opisthosomal area with irregular striae; seta 2c smooth or slightly serrate, 2b absent; ventral setae 4a, 4b and genital setae g absent. Lengths of ventral setae: 1a 31–40, 3a 4–5; ag 3–4; ps1 2–3, ps2 2–3.
Gnathosoma. Palp 4-segmented, palp chaetotaxy as in deutonymph.
Legs (Figs. 7B and 8A–8D). Chaetotaxy of legs: coxae 2-1-1-0; trochanters 0-0-1-0; femora 4-4-2-1; genua 1-1-1-0; tibiae 5-5-3-3; tarsi 8+ω-8+ω-5-3. Leg setae similar to that of female.
Larva (n = 4)
Dorsum (Fig. 9A). Idiosoma broadly oval, length 150–162; width 110–118. Rostral shield absent; idiosoma smooth, with few transverse striae posteriorly; opisthosomal pores absent. Dorsal body setae long and narrowly lanceolate except dorsocentral setae c1, d1, e1, h1 minute; setae v2 shorter than distance between v2–v2. Lengths of dorsal setae: v2 16–18, sc1 14–16, sc2 15–17, c1 3–4, c2 12–14, c3 15–16, d1 2–3, d3 15–17, e1 2–3, e3 17–18, f2 16–17, f3 16–17, h1 3–5, h2 17–18.
Venter (Fig. 9B). Venter of idiosoma completely striated; ventral setae 4a, coxal setae 1c, 2b, 2c, 3b, aggenital setae ag and genital setae g absent. Lengths of ventral setae: 1a 28–34, 3a 6–7; ps1 3–4, ps2 2–3.
Gnathosoma. Palp 4-segmented, palp chaetotaxy as in female.
Legs (Figs. 9B and 10A–10C). Chaetotaxy of legs: coxae 1-0-0; trochanters 0-0-0; femora 3-3-2; genua 1-1-1; tibiae 5-5-3; tarsi 6+ω-6+ω-3.
Type material
Female holotype, 24 female paratypes, 10 male paratypes, six deutonymphs, two protonymphs and four larvae; ex. leaves of Rhaphiolepis indica var. umbellata Makino (Rosaceae); Chiba, Japan (35°02′16″N, 139°50′15″E); 14 June 2018; leg. M.W. Negm. Type depository: female holotype, two female paratypes, three male paratypes, two deutonymphs, two protonymphs and two larvae will be deposited in the National Museum of Nature and Science (NMNS), Tsukuba, Ibaraki Prefecture, Japan. The remainder types are deposited in the Laboratory of Applied Entomology and Zoology, Ibaraki University (AEZIU) with the voucher specimen no. 895.
Etymology
The specific name umbellatus is named after the host plant species. The gender is masculine.
Differential diagnosis
Cenopalpus umbellatus sp. nov. closely resembles C. lanceolatisetae (Attiah, 1956) in various aspects including the chaetotaxy of legs; however, female differs in having rostrum not reaching distal end of femur I (vs. rostrum extending to middle of genu I in C. lanceolatisetae), reticulations behind ventral setae 4a medially connected (vs. smooth or slightly striate medially in C. lanceolatisetae) and variation in lengths of some idiosomal setae (Table 3). Male of C. umbellatus sp. nov. also differs in having reticulations behind ventral setae 4a (vs. reticulations absent in C. lanceolatisetae) and in having no opisthosomal pores (vs. one pair of opisthosomal pores present in C. lanceolatisetae). Also, the deutonymph of the new species has propodosoma smooth medially (vs. propodosoma reticulated medially in C. lanceolatisetae).
| Setae |
C. lanceolatisetae (range for 10 females) (Khanjani et al., 2012) |
C. umbellatus sp. nov. holotype (range for paratypes) |
|---|---|---|
| v2 | 18–26 | 24 (22–26) |
| sc1 | 17–23 | 16 (15–17) |
| sc2 | 18–24 | 13 (12–14) |
| c1 | 11–16 | 9 (9–11) |
| c2 | 13–19 | 13 (14–15) |
| c3 | 12–18 | 17 (16–19) |
| d1 | 7–11 | 8 (7–8) |
| d3 | 11–18 | 14 (13–14) |
| e1 | 7–12 | 7 (6–7) |
| e3 | 13–16 | 13 (12–14) |
| f2 | 13–16 | 12 (10–11) |
| f3 | 10–14 | 11 (11–12) |
| h1 | 5–9 | 6 (6–7) |
| h2 | 10–14 | 10 (9–10) |
| 1a | 75–103 | 80 (75–82) |
| 3a | 12–16 | 9 (8–10) |
| 4a | 80–119 | 70 (65–70) |
| ag | 13–18 | 13 (12–14) |
| g1 | 9–12 | 10 (10–12) |
| g2 | 8–13 | 9 (9–11) |
| ps1 | 12–16 | 10 (9–10) |
| ps2 | 5–10 | 8 (8–10) |
Ontogeny
The ontogenetic changes in the idiosomal and leg chaetotaxy of Cenopalpus umbellatus sp. nov. resemble the typical pattern for tenuipalpid mites (Lindquist, 1985). Regarding the setal additions on ventral idiosoma, the ventral (1a, 3a) and anal (ps2, ps1) setae appeared since the larval stage. However, aggenital seta (ag) is added in the protonymph and the ventral seta (4a) is added in the deutonymph. Also, genital setae (g1) appeared in the deutonymph and g2 in the adults. The coxal setae 1c, 2c and 3b are added in the protonymph and the setae 2b and 4b are added in the deutonymph. Setae v’ appeared on trochanters I, II and III in the deutonymph while appeared on trochanters IV in the adults. Seta l’ on trochanter III is added in the protonymph. Also, seta l’ is added to femora I and II in protonymph. Setae l’ is present on genua I and II of the larva. Setae d and l” are added to genua I and II in the deutonymph. The tectal setae (tc’, tc”) are added to tarsus I, II and III in the protonymphal stage.
Key to world species of Cenopalpus (based on females)
1. Opisthosoma with 6 pairs of dorsolateral setae2
—Opisthosoma with 7 pairs of dorsolateral setae7
2. Palp-tibia and palp-tarsus with 2 setae each3
—Palp-tibia with 1 seta and palp-tarsus with 2 setaecreticus
3. Rostrum extending beyond distal end of femur I4
—Rostrum extending to mid-level of femur I, not reaching to distal end5
4. Dorsal setae rod-likepistaciae
—Dorsal setae feather-likepterinus
5. Setal formula of tibiae 5-5-3-36
—Setal formula of tibiae 5-5-5-3arbuti
6. Setal formula of trochanters 1-1-1-1; reticulations behind setae 4a partly separated mediallyofficinalis
—Setal formula of trochanters 1-1-2-1; reticulations behind setae 4a prominent and not separated mediallyadventicius
7. Idiosoma mostly striate or partly striate and partly reticulate8
—Idiosoma mostly reticulate12
8. Dorsum mostly striate but also with reticulations on prodorsum and between c and d series on hysterosoma; setae 3a and 4a very longtamarixi
—Dorsum striate with setae 4a much longer than short 3a9
9. Rostral shield with 2 slightly notched medial lobes10
—Rostral shield with 2 medial and 2 lateral lobes11
10. Setae 4a on venter much longer than distance between setae 3a and 4a, setae 1a very long and whip-like extending considerably pass rostrumwainsteini
—Setae 4a approximately equal to, or little longer than, distance between setae 3a and 4a, setae 1a not extending pass rostrumsaryabiensis
11. Rostrum reach almost to middle of genu I; hysterosoma with transverse striae from prodorsum to behind setae d1 and longitudinal to posterior marginaratus
—Rostrum reach almost to middle of femur I; striae on hysterosoma mainly transverselineola
12. Propodosomal setae broadly lanceolate to spatulate or scoop-like13
—Propodosomal setae narrowly lanceolate to setiform or slender37
13. Propodosomal setae broadly lanceolate to spatulate; opisthosomal pores absent (one pair present in pennatisetis)14
—Propodosomal setae scoop-like; 2 pairs of opisthosomal pores presentscoopsetus
14. Rostrum reaching behind distal end of femur I15
—Rostrum not reaching beyond distal end of femur I30
15. Rostrum extending beyond distal end of genu I16
—Rostrum not extending beyond distal end of genu I18
16. Setae sc1 shorter than distance between bases of setae sc1 and sc217
—Setae sc1 longer than distance between bases of setae sc1 and sc2khosrowshahi
17. Setae sc1 less than half of distance between bases of setae sc1 and sc2prunusi
—Setae sc1 more than half of distance between bases of setae sc1 and sc2longirostris
18. Propodosoma with reticulations regular19
—Propodosoma with reticulations irregular26
19. Setae sc1 shorter than distance between bases of setae sc1 and sc2.20
—Setae sc1 longer than, or equal to, distance between bases of setae sc1 and sc223
20. Dorsal body setae subspatulate, narrowly or broadly lanceolate21
—Dorsal body setae broadly spatulateeriobotryi
21. Setae v2 broadly lanceolate and much longer than half of distance between their bases; rostral shield with 2 medial, 2 submedial and 2 lateral lobes22
—Setae v2 narrowly lanceolate and equal to, or little longer than, half of distance between their bases; rostral shield with 2 medial lobeschitraliensis
22. Metapodosomal venter posterior to setae 4a smooth medially or slightly striate; rostrum extending to middle of genu Ilanceolatisetae
—Metapodosomal reticulations on venter posterior to setae 4a connected medially; rostrum not reaching pass distal end of femur Iumbellatus sp. nov.
23. Dorsal setae subspatulate with long spinesviniferus
—Dorsal setae subspatulate or narrowly lanceolate and serrate24
24. Dorsal setae narrowly lanceolate and setae c1 almost as long as distance between its members25
—Dorsal setae subspatulate with setae c1 clearly shorter than distance between its membersxini
25. Setal formula of trochanters 1-1-2-1, femora 4-4-2-1pennatisetis
—Setal formula of trochanters 1-1-1-1, femora 4-4-2-0virgulatus
26. Setae v2 shorter than distance between their bases27
—Setae v2 longer than, or equal to, distance between their bases28
27. Rostrum at level of distal end of genu I; rostral shield basically with only 2 medial lobeshalperini
—Rostrum not reaching distal end of genu I; rostral shield with 2 medial and 2 lateral lobespegazzanoae
28. Rostrum reaching to middle or to distal margin of genu I; propodosomal setae broadly lanceolate29
—Rostrum reaching beyond distal end of femur I; propodosomal setae spatulateevini
29. Propodosoma with large polygonal reticulations mediallyabaii
—Propodosoma smooth or weakly reticulate mediallybagdasariani
30. Dorsal body setae spatulate or subspatulate31
—Dorsal body setae lanceolatehaqii
31. Dorsal body setae spatulate32
—Dorsal body setae subspatulate34
32. Propodosoma with regular polygonal reticulationscapensis
—Propodosoma with irregular reticulations, especially mediodorsally and mediolaterally33
33. Metapodosomal venter with area posterior to setae 4a completely reticulated, anterior to 4a weakly reticulatesalignae
—Metapodosomal venter with area posterior to setae 4a smooth medially or slightly striate and smooth anterior to 4aoleunus
34. Metapodosomal venter with area posterior to setae 4a smooth medially35
—Metapodosomal venter with area posterior to setae 4a reticulated36
35. Setae v2 equal to, or little shorter than, distance between their basesplatani
—Setae v2 approximately half of distance between their basesramus
36. Setae v2 approximately half of distance between their bases; idiosoma with dorsal reticulations regular; dorsal setae short and serratenatalensis
—Setae v2 equal to distance between their bases; idiosoma with dorsal reticulations irregular; dorsal setae clearly longer and strongly serratepritchardi
37. Setae v2 approximately longer than, or equal to, distance between their bases38
—Setae v2 shorter than distance between their bases51
38. Rostral shield with 2 medial lobes, lateral lobes excluded39
—Rostral shield with 4 medial lobes, one pair can be reduced or obsolete, lateral lobes also excluded42
39. Rostrum reaching up to distal end of femur I; metapodosomal venter with area posterior to setae 4a smooth medially40
—Rostrum reaching to middle of genu I; metapodosomal venter with area posterior to setae 4a reticulated41
40. Setal formula of tibiae 4-4-3-3mughalii
—Setal formula of tibiae 5-5-3-3orakiensis
41. Propodosoma with small, rounded crenulate elementsspinosus
—Propodosoma with large polygonal reticulationspulcher
42. Dorsal body setae mostly lanceolate43
—Dorsal body setae mostly setiform47
43. Opisthosoma with pores44
—Opisthosoma without pores45
44. Rostrum not extending beyond distal end of femur I, rostral shield with 4 distinct lobes mediallyquadricornis
—Rostrum extending beyond distal end of femur I, second pair of medial lobes obsoleteirani
45. Setae c1 and d1 long, almost as long as distances between their membersquercusi
—Setae c1 and d1 much shorter, half or less than half the distances between their members.46
46. Setal formula of genua 3-3-3-1, trochanters 1-1-2-1taygeticus
—Setal formula of genua 3-3-1-0, trochanters 1-1-1-1naupakticus
47. Setae sc1 approximately equal to, or longer than, distance between bases of setae sc1 and sc248
—Setae sc1 distinctly shorter than distance between bases of setae sc1 and sc2meyerae
48. Setae sc1 approximately equal to distance between bases of setae sc1 and sc249
—Setae sc1 distinctly longer than distance between bases of setae sc1 and sc2brachypalpus
49. Setae sc2 long, almost reaching to sejugal furrowmusai
—Setae sc2 short, distinctly far from sejugal furrow50
50. Venter between setae 3a and 4a striaterubusi
—Venter between setae 3a and 4a smoothpseudospinosus
51. Rostrum extending to middle of femur I or somewhat beyond middle52
—Rostrum extending to distal end of femur I or beyond56
52. Opisthosoma with dorsolateral setae c3 about a fifth as long as distance to bases of setae d353
—Opisthosoma with dorsolateral setae c3 about a third as long as distance to bases of setae d354
53. Setae v2 shorter than half of distance between their bases; reticulations ventrally behind setae 4a continuouscumanicus
—Setae v2 longer than half of distance between their bases; reticulations behind setae 4a smooth mediallythelycraniae
54. Metapodosomal venter at area posterior to setae 4a with smaller polygonal to rounded crenulate elements medially55
—Metapodosomal venter at area posterior to setae 4a with medial reticulation elements polygonal and broader than longcarpini
55. Setae v2 shorter than half of distance between their baseshederae
—Setae v2 longer than half of distance between their basesmespili
56. Rostrum reaching not beyond distal end of genu I; palp-tarsus with at least a solenidion and seta or eupathium57
—Rostrum reaching to distal end of tibia I; palp-tarsus with 1 solenidion onlypicitilis
57. Rostrum reaching to mid-level or distal end of genu I58
—Rostrum reaching not beyond distal end of femur I63
58. Dorsal setae narrowly lanceolate59
—Dorsal setae setiform60
59. Body almost round; rostrum reaching distal end of genu I; setal formula of tibiae 4-4-3-3sunniensis
—Body oval; rostrum reaching to mid-level of genu I, not reaching distal end; setal formula of tibiae 5-5-3-3ruber
60. All dorsal setae serrate61
—All dorsal setae simpledignus
61. Rostrum reaching distal end of femur I62
—Rostrum reaching distal end of genu Ifavosus
62. Setae v2 more than 15 µm length; setal formula of tibiae 5-4-3-3, coxae 2-2-1-1kritos
—Setae v2 less than 10 µm; setal formula of tibiae 5-5-3-3, coxae 3-2-1-1homalos
63. Rostral shield with 2 medial lobes64
—Rostral shield with more than 2 lobes65
64. Metapodosoma with large polygonal reticulation medioventrally; setae 4a much longer than distance between bases of setae 3a and 4a; setal formula of coxae 2-2-1-1, trochanters 1-1-2-2piger
—Metapodosoma with irregular reticulation medioventrally; setae 4a shorter than distance between bases of setae 3a and 4a; setal formula of coxae 2-2-2-1, trochanters 1-1-1-0japonicus
65. Reticulations almost absent or medially smooth behind ventral setae 4a66
—Area behind setae 4a completely reticulated68
66. Area behind setae 4a almost smooth with only a few reticulations behind coxae IV; dorsal setae narrowly lanceolate and serrate or short setiform, serrate67
—Reticulations behind setae 4a with a narrow smooth band medially; dorsal setae short, setiform and serrate or some smoothiqbali
67. Dorsal setae narrowly lanceolate, serratecapacis
—Dorsal setae short, setiform, serratelimbatus
68. Rostral shield with 2 medial and 2 lateral lobes69
—Rostral shield with 2 medial, 2 submedial and 2 lateral lobescrataegi
69. Propodosomal setae narrowly lanceolate; some setae on opisthosoma also lanceolatepopuli
—All dorsal setae setiformbakeri
Discussion
The present study provides morphological description of a new species of flat mites belonging to the genus Cenopalpus, with a key to the world species. This genus is mainly reported from the Mediterranean and East Asia regions. Only 14 tenuipalpid species were previously known from Japan, with only one Cenopalpus species. Though members of the Tenuipalpidae are currently not posing a serious threat to agriculture in the country, we must be prepared for the consequences of global trafficking of people and goods. Therefore, this study will for sure act as a very useful early intervention tool. Examination of all known species of Cenopalpus was toilsome especially with some species which are poorly described, and we had to rely on what was available.
Conclusions
Faunistic information about flat mites in Japan is scarce. The new mite species described with the world key to species increases the available information about the taxonomy of tenuipalpid mites in this country. We hope that this study will serve as the departure point for future research on Cenopalpus mites and encourage for more comprehensive surveys in Japan since a large number of undiscovered species is expected.
{kind=link}