
Rock sponges (lithistid Demospongiae) of the Northeast Atlantic seamounts, with description of ten new species
- Published
- Accepted
- Received
- Academic Editor
- Joseph Pawlik
- Subject Areas
- Biodiversity, Biogeography, Marine Biology, Taxonomy, Zoology
- Keywords
- Porifera, Deep-sea, Lithistids, Biodiversity, Tetractinellida, Bubarida, New species, Biogeography
- Copyright
- © 2020 Carvalho et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Rock sponges (lithistid Demospongiae) of the Northeast Atlantic seamounts, with description of ten new species. PeerJ 8:e8703 https://doi.org/10.7717/peerj.8703
Abstract
Background
Lithistid demosponges, also known as rock sponges, are a polyphyletic group of sponges which are widely distributed. In the Northeast Atlantic (NEA), 17 species are known and the current knowledge on their distribution is mainly restricted to the Macaronesian islands. In the Mediterranean Sea, 14 species are recorded and generally found in marine caves.
Methods
Lithistids were sampled in nine NEA seamounts during the scientific expeditions Seamount 1 (1987) and Seamount 2 (1993) organized by the MNHN of Paris. Collected specimens were identified through the analyses of external and internal morphological characters using light and scanning electron microscopy, and compared with material from various museum collections as well as literature records.
Results
A total of 68 specimens were analysed and attributed to 17 species across two orders, seven families, and seven genera, representing new records of distribution. Ten of these species are new to science, viz. Neoschrammeniella inaequalis sp. nov., N. piserai sp. nov., N. pomponiae sp. nov., Discodermia arbor sp. nov., D. kellyae sp. nov., Macandrewia schusterae sp. nov., M. minima sp. nov., Exsuperantia levii sp. nov., Leiodermatium tuba sp. nov. and Siphonidium elongatus sp. nov., and are here described and illustrated. New bathymetric records were also found for D. ramifera, D. verrucosa and M. robusta. The Meteor seamount group has a higher species richness (15 species) compared to the Lusitanian seamount group (six species). The majority of the species had their distribution restricted to one seamount, and ten are only known from a single locality, but this can be a result of sample bias.
Discussion
The number of species shared between the seamounts and the Macaronesian islands is very reduced. The same pattern repeats between the NEA and Mediterranean Sea. This study demonstrates that NEA seamounts are ecosystems with a higher diversity of lithistids than previously thought, increasing the number of lithistids known to occur in the NEA and Mediterranean Sea from 26 to 36 species.
Introduction
The class Demospongiae Sollas (1885) contains several groups of sponges artificially unified under the name ‘lithistid demosponges’ or ‘rock sponges.’ Lithistids produce hypersilicified spicules (desmas) (Pisera & Lévi, 2002a) that usually creates a very rigid skeleton. For a very long time, they were classified into an order, Lithistida (Schmidt, 1870), but more recently, several studies have shown the polyphyletic nature of this group (Cárdenas et al., 2011; Kelly & Pomponi, 1994; Pisera & Lévi, 2002a; Schuster et al., 2015). It is now acknowledge that this trait, i.e., is the desmas, has evolved independently multiple times (Schuster et al., 2015) and the 211 valid species currently recognized worldwide are distributed in three orders-Tetractinellida Marshall (1876), Sphaerocladina Schrammen (1924) and Bubarida Morrow & Cárdenas (2015), with the large majority belonging to the former order (Morrow & Cárdenas, 2015; Pisera & Lévi, 2002a; Schuster et al., 2015; Van Soest et al., 2019, WPD).
In the Northeast Atlantic (NEA), the current state of knowledge on lithistid sponges is mainly restricted to the Macaronesian islands. So far, 17 species have been described and recorded from the Azores (Carvalho & Pisera, 2019; Gray, 1859; Topsent, 1928, 1904, 1898, 1892), Madeira and Selvagens (Bowerbank, 1869; Carter, 1873; Carvalho & Pisera, 2019; Johnson, 1863), Canary Islands (Carvalho & Pisera, 2019; Cruz, 2002; Topsent, 1892), Portugal mainland (Schmidt, 1870) and Morocco (Lendenfeld, 1907), whereas in the Mediterranean Sea, 15 species have been reported (Maldonado et al., 2015; Manconi, Serusi & Pisera, 2006; Manconi & Serusi, 2008; Perez et al., 2004; Pisera & Vacelet, 2011; Pulitzer-Finali, 1972; Vacelet, 1969). They are commonly found on hard substrate at 110–1,700 m depth (Carter, 1873; Carvalho, Pomponi & Xavier, 2015; Topsent, 1928), whereas in the Mediterranean Sea they usually occur in shallower waters or in cave systems (Manconi & Serusi, 2008; Pisera & Vacelet, 2011). Although the knowledge on distribution for lithistids in the NEA has been increasing, there is no data regarding their occurrence on seamounts in the area.
These topographic features, which provide important habitats for both benthic and pelagic organisms, are very numerous and worldwide distributed (Yesson, 2011). In the NEA, examples include the Lusitanian Seamounts (Coral Patch, Ampere, Gorringe Bank, Hirondelle II, Josephine, Lion, Dragon, Unicorn and Seine), located near the Euro-African continental shelf, approximately 250 km from the Portuguese coast and the Meteor Seamounts (Great Meteor, Hyères, Irving, Cruiser, Plato, Tyro and Atlantis), situated in the central part of the North Atlantic, close to the Mid-Atlantic Ridge (MAR) and south of the Azores archipelago. These seamounts have evoked interest for research in the late 19th and early 20th Century, and several scientific expeditions took place, such as Josephine (1869), Challenger (1873) and numerous Prince Albert I of Monaco expeditions. Late in the 20th and early 21st Centuries, new efforts aiming to explore the benthic fauna of these seamounts were undertaken. Two of these expeditions—Seamount 1 and Seamount 2—organized by the Natural History Museum of Paris (MNHN), surveyed various of the Lusitanian and Meteor seamounts at depths above 1,000 m (Bouchet & Métivier, 1988; Gofas, 1993). These expeditions resulted in the discovery and description of several species of various taxonomic groups, such as brachiopods (Logan, 1998), bryozoans (Berning, Harmelin & Bader, 2017; Souto, Berning & Ostrovsky, 2016), bivalves (Dijkstra & Gofas, 2004), corals (Molodtsova & Shirshov, 2011), cirripeds (Young, 2001), hydrozoans (Ramil, Vervoort & Ansín, 1998), polychaetes (Gillet & Dauvin, 2003; Paxton & Gillet, 2004) and gastropods (Gofas, 2007) greatly advancing the understanding of the biogeographic patterns and the biodiversity of these ecosystems. However, several taxonomic groups, including sponges, remain scarcely documented in the literature for these ecosystems (Cárdenas et al., 2018; Cristobo et al., 2015; Lévi & Vacelet, 1958; Topsent, 1928; Xavier & Van Soest, 2007).
In this study, we describe the lithistid demosponges collected during the French expeditions Seamount 1 and Seamount 2. New records of geographic distribution are reported, ten new species for science are described and illustrated, and the diversity and biogeographic patterns discussed. An identification key of all lithistid species reported for the NEA and Mediterranean is also provided.
Materials and Methods
The material examined in this study was collected during Seamount 1 and Seamount 2 scientific expeditions undertaken by the MNHN of Paris to several NEA seamounts (Fig. 1; Supplemental Material S1). The main aims of these campaigns were to study the patterns of faunal diversity and endemism found on isolated seamounts in comparison to continental areas and the relation with the dispersal capacity of the various taxonomic groups. The Seamount 1 campaign, coordinated by Dr. Philippe Bouchet, took place in 1987 onboard of the research vessel L. Noroît, and explored the Galicia Banks and the Lusitanian Seamounts (Gorringe, Josephine, Ampère, Lion and Seine) (Bouchet & Métivier, 1988). The second campaign, Seamount 2, this time lead by Dr. Serge Gofas, explored the Meteor Seamounts group (Great Meteor, Hyères, Irving, Cruiser, Plato, Atlantis and Tyro) and the Antialtair Seamount on board of the RV L. Suroît, sampling 165 stations also at depths above 1,000 m (Gofas, 1993). Lithistids were collected in 10 stations on Seamount 1 (11%) and in 42 stations on Seamount 2 (32%) between 280 and 1,035 m depth using various sampling gears (beam trawl (CP), epibenthic dredge (DE) and Warén dredge (DW)), and preserved in formalin onboard. The specimens examined are deposited in the ‘zoothèque’ of the MNHN in Paris, and stored at room temperature in ethanol 70%. Detailed information regarding the collection of the specimens studied here, is deposited in PANGAEA® Data Publisher (www.pangaea.de) under the digital object identifier (DOI): https://doi.pangaea.de/10.1594/PANGAEA.896492.
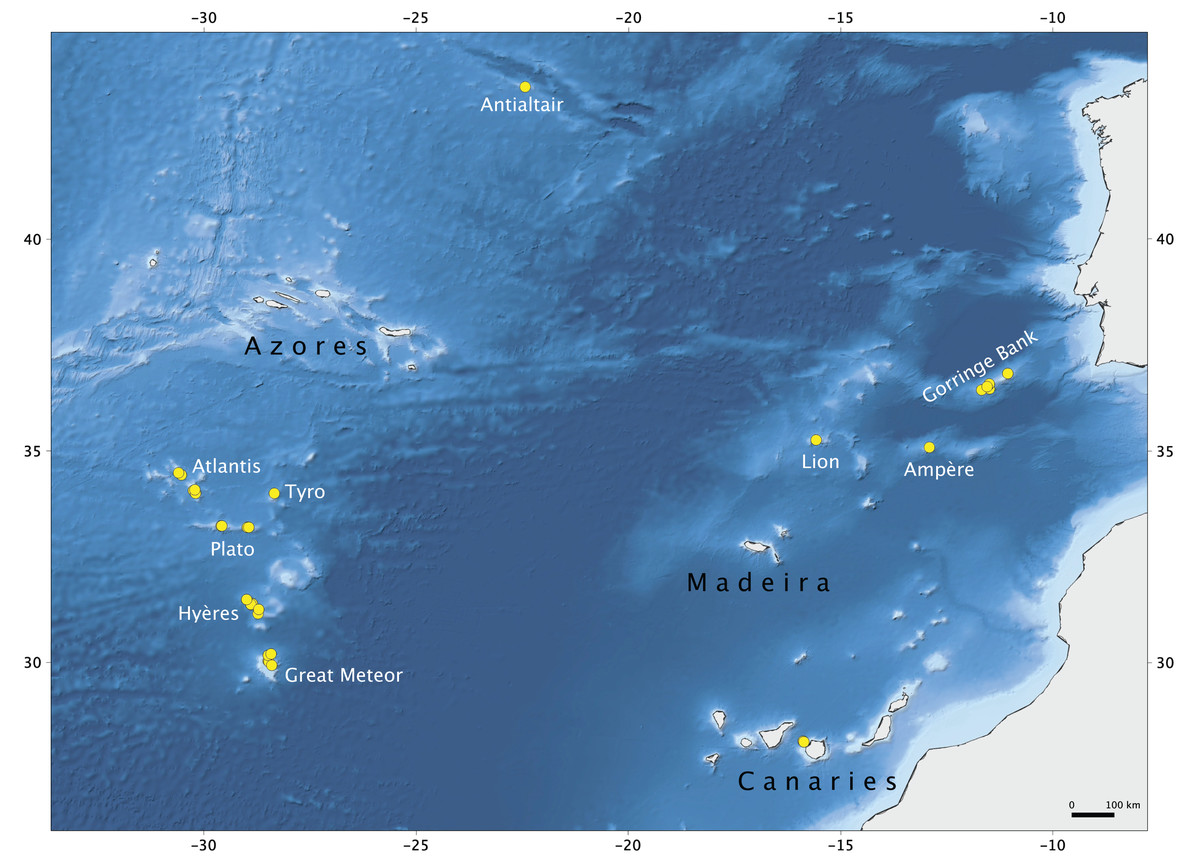
Figure 1: Map of the study area.
Seamounts of the Northeast Atlantic and stations of the Seamount 1 and Seamount 2 campaigns where lithistid demosponges were collected. Map produced with the software QGIS Development Team (2019); bathymetry obtained from GEBCO Compilation Group (2019).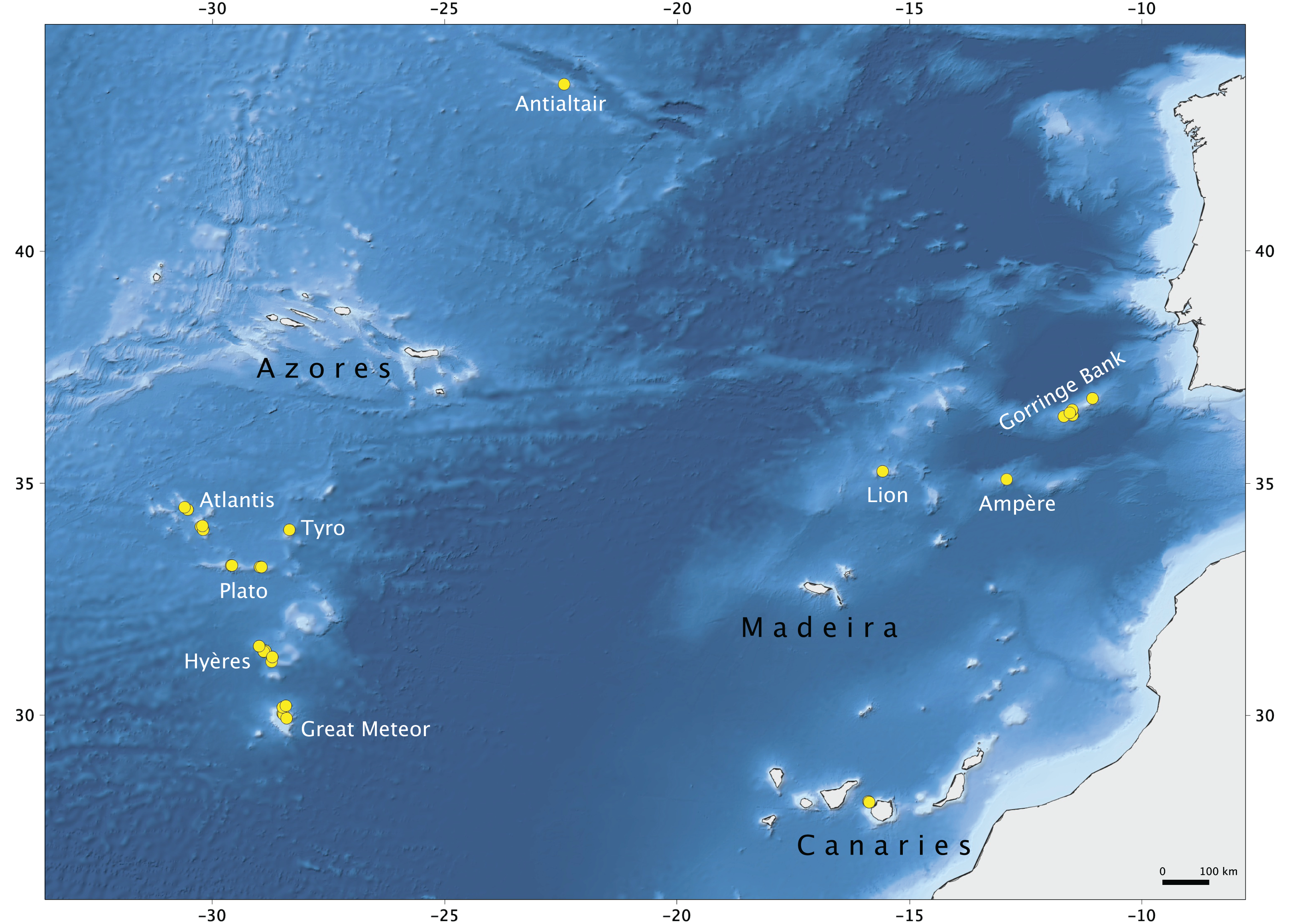{kind=link}
The specimens were analysed through the use of Light Microscopy (LM) and Scanning Electron Microscopy (SEM). For light microscopy, cross sections and slides of loose spicules were mounted in Canada Balsam® Sigma–Aldrich or Eukit® Sigma–Aldrich following standards procedures (Boury-Esnault & Rutzler, 1997). In addition, a few specimens, representative of each species, were selected and prepared for SEM. For this purpose, pieces of both the ectosome and choanosome of the sponge were excised and then either directly mounted or digested in nitric acid, washed several times with distilled water and then fixed in ethanol. The spicules were then placed on a stub and covered with gold-paladium. Thirty spicules of each spicule type were measured using the Leica Application Suite (LAS v. 4.5), for individual specimens. Minimum, mean and maximum values are presented for the measurements obtained for each analysed specimen. For the higher taxa classification, we followed the revised Demospongiae classification (Morrow & Cárdenas, 2015).
Due to the formalin fixation, we were not able to extract DNA for molecular analysis, and any attempts to barcode the mitochondrial COI gene, including the mini-barcode protocol used in other tetractinellids (Cárdenas & Moore, 2017) were unsuccessful.
The electronic version of this article in PorTable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank Life Science Identifiers (LSIDs) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:A0DA0236-4579-47A4-8BE4-E68803C2EC8F. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Results
In this study we analysed 68 specimens, collected between 280 and 1,035 m depth on eight NEA seamounts, and assigned them to 17 species distributed across two orders, seven families, and seven genera (Figs. 2–3). Of these, ten species are new for science—Neoschrammeniella inaequalis sp. nov., N. piserai sp. nov., N. pomponiae sp. nov., Discodermia arbor sp. nov., D. kellyae sp. nov., Macandrewia schusterae sp. nov., M. minima sp. nov., Exsuperantia levii sp. nov., Leiodermatium tuba sp. nov. and Siphonidium elongatus sp. nov (see below descriptions and illustrations). All analysed material is described and illustrated below and compared with additional specimens from various museum collections (MNHN, HBOI, RMNH and DOP). An identification key for all lithistid species recorded to date for the NEA and MED is also provided. All new species described here have the taxonomic authority restricted to the first and last author.
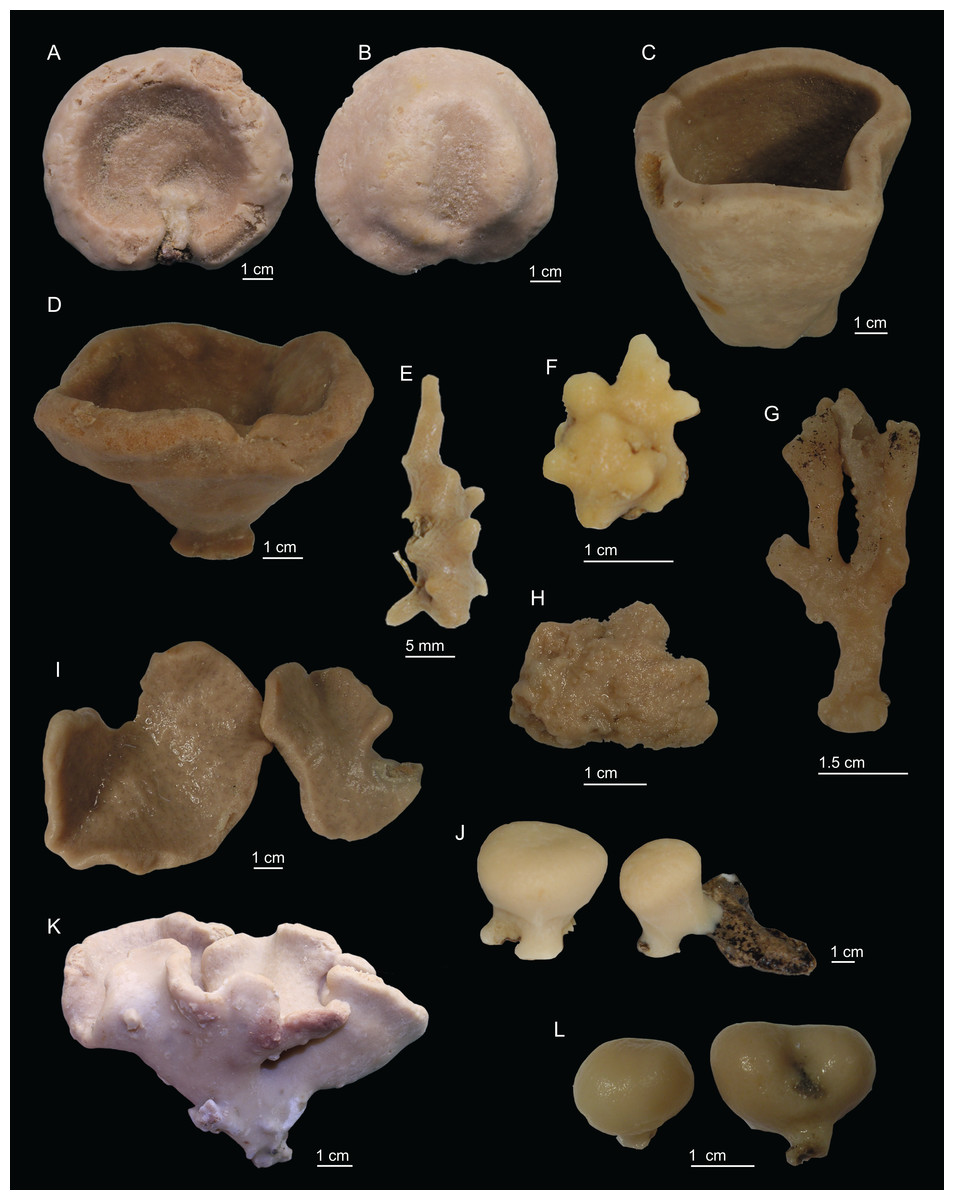
Figure 2: Specimens collected during Seamount 1 and Seamount 2 expeditions.
(A) Top view of Neoschrammeniella inaequalis sp. nov., holotype MNHN-IP-2018-84, (B) bottom view of N. inaequalis sp. nov., holotype MNHN-IP-2018-84. (C) N. piserai sp. nov., holotype MNHN-IP-2008-234. (D) N. pomponiae sp. nov., holotype MNHN-IP-2008-233. (E) Discodermia ramifera Topsent, 1892, specimen MNHN-IP-2008-213. (F) D. verrucosa Topsent, 1928, specimen MNHN-IP-2008-205. (G) D. arbor sp. nov., holotype MNHN-IP-2008-211. (H) D. kellyae sp. nov., holotype MNHN-IP-2008-208. (I) Macandrewia cf. azorica, specimen MNHN-IP-2008-220. (J) M. robusta Topsent, 1904, specimens MNHN-IP-2008-216. (K) M. schusterae sp. nov., holotype MNHN-IP-2018-87. (L) M. minima sp. nov., holotype MNHN-IP-2008-222.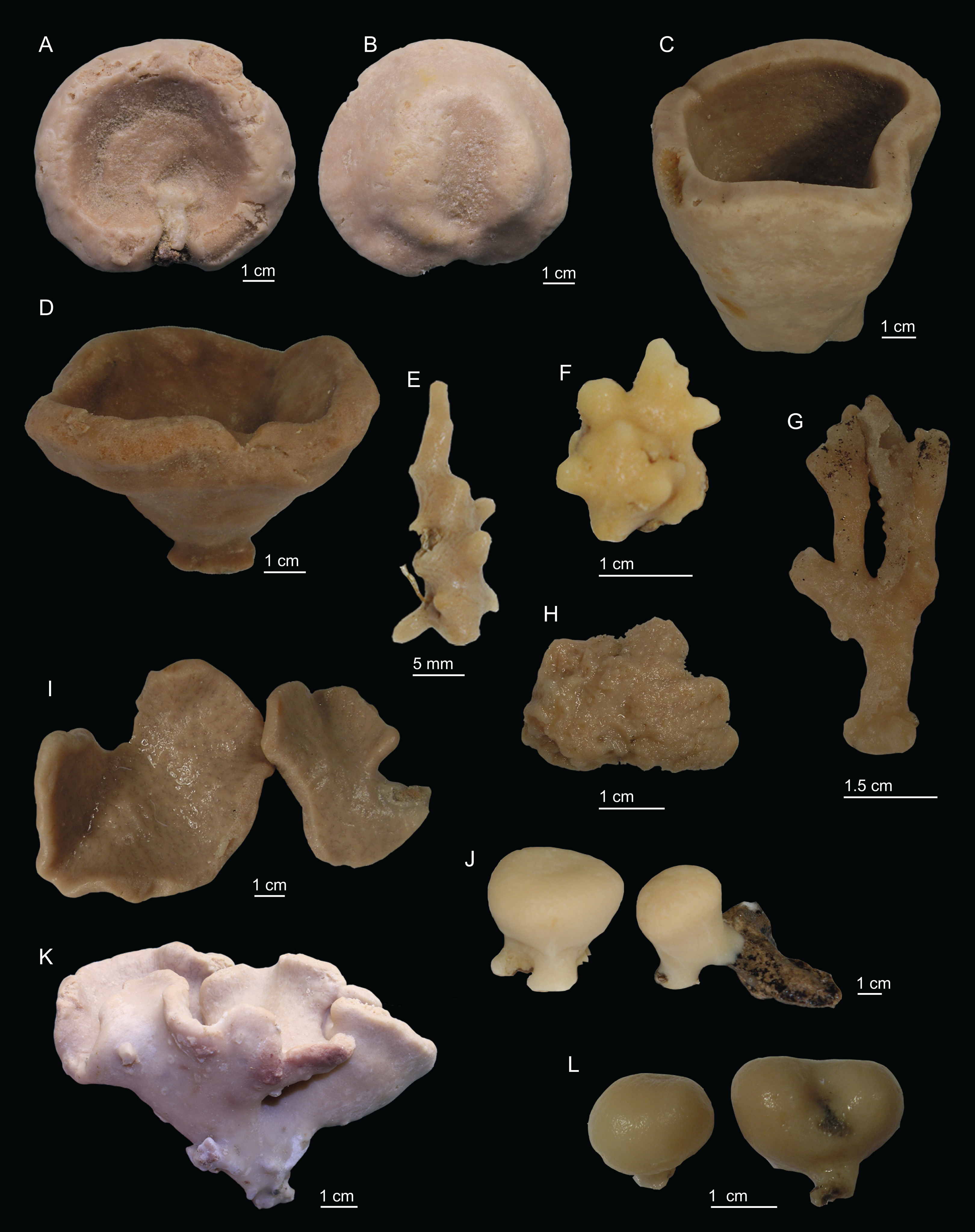{kind=link}
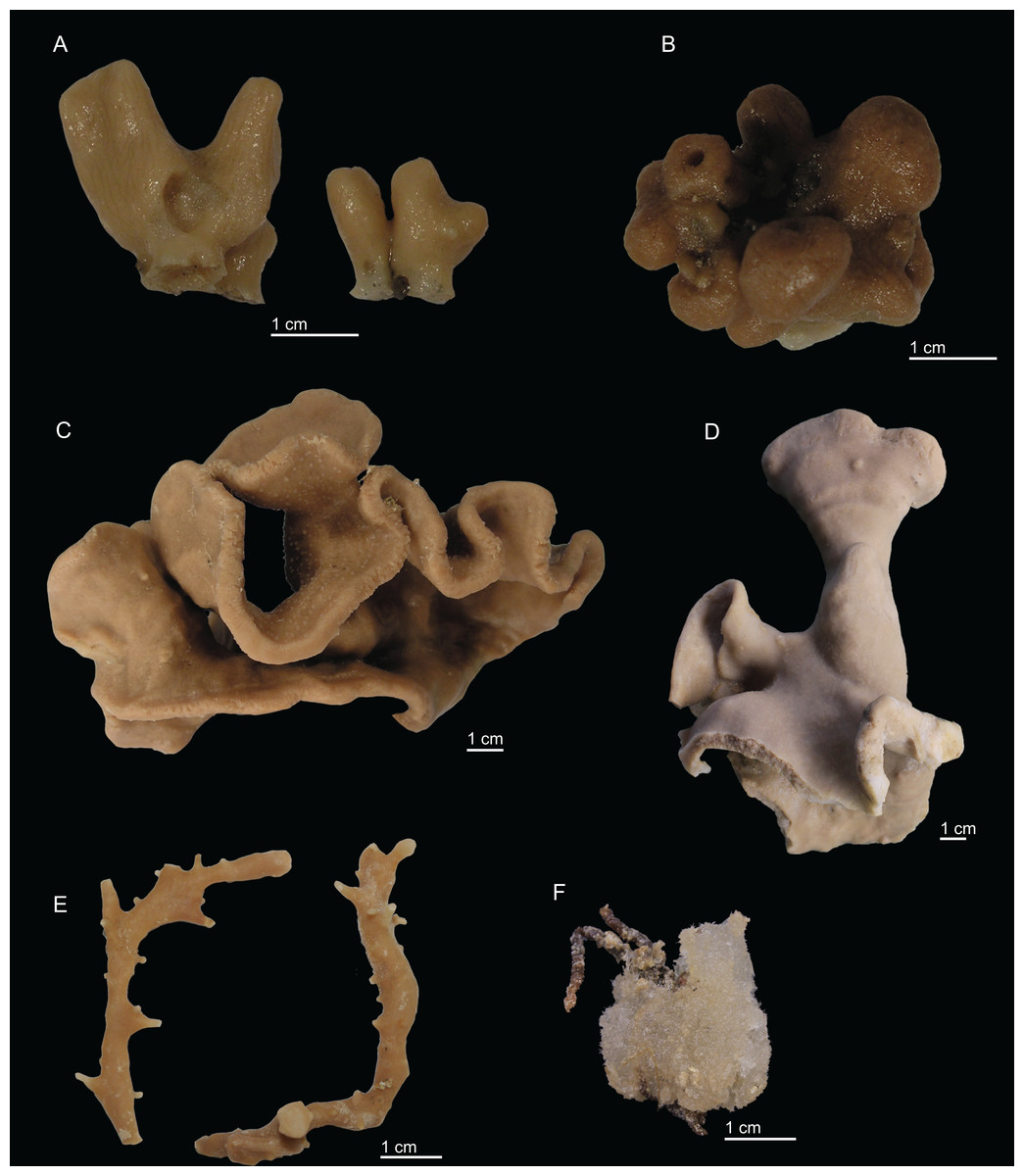
Figure 3: Specimens collected during Seamount 1 and Seamount 2 expeditions.
(A) Exsuperantia archipelagus Carvalho & Pisera (2019), specimen MNHN-IP-2008-196. (B) E. levii sp. nov., holotype MNHN-IP-2008-201. (C) Leiodermatium lynceus Schmidt (1870), specimen MNHN-IP-2008-239. (D) L. tuba sp. nov., holotype MNHN-IP-2018-72. (E) Siphonidium elongatus sp. nov., holotype MNHN-IP-2008-236. (F) Petromica (Petromica) grimaldii Topsent, 1898, MNHN-IP-2018-92.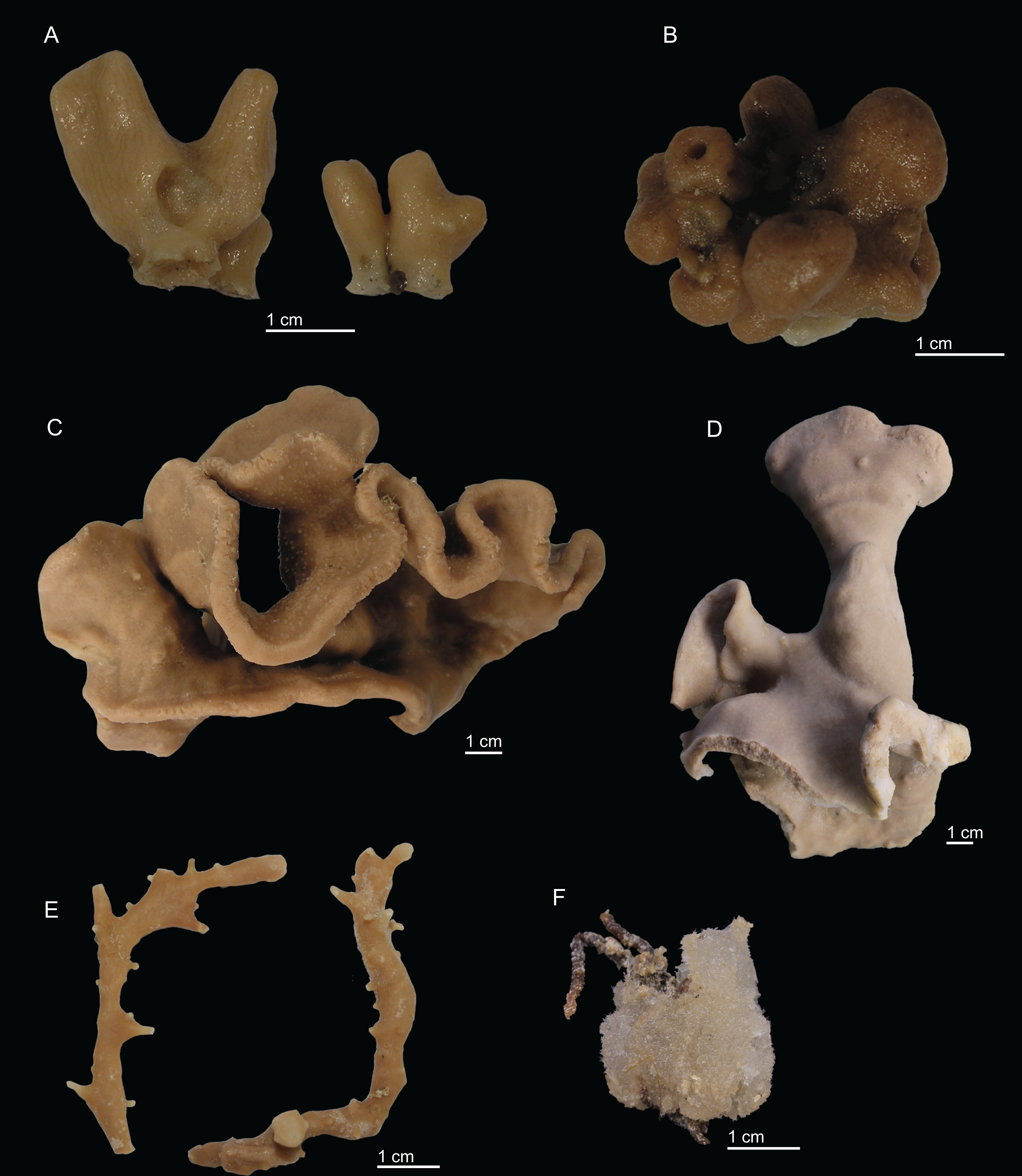{kind=link}
Systematic Index
Phylum Porifera Grant, 1836
Class Demospongiae Sollas, 1885
Subclass Heteroscleromorpha Cárdenas, Pérez & Boury-Esnault, 2012
Order Tetractinellida Marshall, 1876
Suborder Astrophorina Sollas, 1887
Family Corallistidae Sollas, 1888
Genus Neoschrammeniella Pisera & Lévi, 2002b
Species Neoschrammeniella inaequalis sp. nov.
Species Neoschrammeniella piserai sp. nov.
Species Neoschrammeniella pomponiae sp. nov.
Family Theonellidae Lendenfeld, 1903
Genus Discodermia du Bocage, 1869
Species Discodermia ramifera Topsent, 1892
Species Discodermia cf. ramifera Topsent, 1892
Species Discodermia verrucosa Topsent, 1928
Species Discodermia arbor sp. nov.
Species Discodermia kellyae sp. nov.
Family Macandrewiidae Schrammen, 1924
Genus Macandrewia Gray, 1859
Species Macandrewia cf. azorica Gray, 1859
Species Macandrewia robusta Topsent, 1904
Species Macandrewia schusterae sp. nov.
Species Macandrewia minima sp. nov.
Family Phymaraphiniidae Schrammen, 1924
Genus Exsuperantia Özdikmen, 2009
Species Exsuperantia archipelagus Carvalho & Pisera, 2019
Species Exsuperantia levii sp. nov.
Suborder Spirophorina Bergquist & Hogg, 1969
Family Azoricidae Sollas, 1888
Genus Leiodermatium Schmidt, 1870
Species Leiodermatium lynceus Schmidt, 1870
Species Leiodermatium tuba sp. nov.
Family Siphonidiidae Lendenfeld, 1903
Genus Siphonidium Schmidt, 1879
Species Siphonidium elongatus sp. nov.
Order Bubarida Morrow & Cárdenas, 2015
Family Desmanthidae Topsent, 1893
Genus Petromica Topsent, 1898
Subgenus Petromica (Petromica) Topsent, 1898
Species Petromica (Petromica) grimaldii Topsent, 1898
Species descriptions
Order TETRACTINELLIDA Marshall, 1876
Suborder ASTROPHORINA Sollas, 1887
Family CORALLISTIDAE Sollas, 1888
Genus Neoschrammeniella Pisera & Lévi, 2002b
Synonymy. Iouea sensu Lévi & Lévi, 1988: 248.
Diagnosis. Corallistidae with smooth dichotriaenes and two to three types of microscleres: metasters, amphiasters/streptasters and/or spirasters (emended after Kelly, 2007; Pisera & Lévi, 2002b; Pisera & Vacelet, 2011; Schlacher-Hoenlinger, Pisera & Hooper, 2005).
Definition. Polymorphic Corallistidae, shallow cup-shaped or deep vase-shaped; surface can be smooth or rugose; ectosomal megascleres are smooth dichotriaenes; choanosomal megascleres are dicranoclone desmas with different types of ornamentation, varying from poorly to extremely tuberculated in different species; diactines are frequently present in the ectosome and triaenes are rare; microscleres are metasters, amphiaster/streptaster and/or acanthose spirasters (type I covered by short blunt rays, and type II irregular with short blunt rays only on the edges), but the number and type of microscleres varies between species (emended after Kelly, 2007; Pisera & Lévi, 2002b; Pisera & Vacelet, 2011; Schlacher-Hoenlinger, Pisera & Hooper, 2005).
Type species. Neoschrammeniella moreti Lévi & Lévi, 1988 (type by monotypy).
Neoschrammeniella inaequalis sp. nov.
Figures 2A–2B, 4–5 and Table 1
| Habitus | Size | Dicranoclones | Dichotriaenes | Oxeas | Spirasters | Metasters | Locality | |
|---|---|---|---|---|---|---|---|---|
| 1N. bowerbankii Holotype BMNH 69.11.60.1 (PZS 1862) | – | – | – | Cladome: 319–397 µm in diameter; rhabdome 487–939 µm length | – | 20.2–23.7 × 7.0–11.7 µm (as spiraster type I) | 28.3–39.3 × 19.6–32.7 µm (as spiraster type II) | Madeira (depth unknown) |
| 2N.bowerbankii (Johnson, 1863) | Cup-shaped to contornated lamellate masses with thick walls; colour white | 80 × 60 × 60 mm in size | 290–402 µm in size | Cladome: 176–323 µm; rhabdome: 223–513 µm |
340–820 × 1.5–2.5 µm | Short arms, 17–24 × 7.06–11.1 µm in size (as spiraster type I) | Long arms, 26.2–39.2 × 18.5–23.9 µm (as spiraster type II) | Mediterranean Sea (20–22 m) |
| N. inaequalis sp. nov. (Holotype MNHN-2018-84) | Flattened cup-shape, with a concave center; both surfaces are smooth; colour light brown | 73 × 64 × 29 mm in size; walls, 14–17 mm thick | 354–576–975 × 12–25–39 µm (n = 12) | Cladome: smooth, very irregular, 118–233–406 µm; rhabdome: long with a round tip, 136–432–1211 × 9–18–31 µm |
Large, thin, curved, 670–1144 × 5.2–7.8–13.4 µm (n = 5) | Short with thick arms, very abundant, 12.1–18.5–26.6 µm | Long and thin arms, 14.6–31.6–47.9 µm | Gorringe Seamount (605–675 m depth) |
| N. inaequalis sp. nov. (Paratype MNHN-2018-85) | Small, ball shaped with a concave top; both surfaces are smooth; colour light brown | 34 mm diameter, 20 mm height | 308–431–575 × 21–34–49 µm (n = 15) | Cladome: 158–298–463 µm; rhabdome: 221–550–1228 × 13–23–38 µm |
Large, thin, curved 449–1034 × 5–7–10 µm (n = 8) | 10.4–20.3–26.1 µm | 15.1–32.7–47.6 µm (n = 17) | Gorringe Seamount (605–675 m depth) |
|
N. piserai sp. nov. (Holotype MNHN-IP-2008-234) |
Large cup-rectangular sponge attached to the substrate by the entire lower base; both surfaces smooth; colour beige | 69 mm in diameter at the top, and 43 mm at the base, 98 mm height; walls, 11 mm thick | 280–428–522 × 16–25–37 µm (n = 6) | Cladome: smooth, 153–244–389 µm; rhabdome: long with a round tip, 198–366–535 × 10–19–33 µm |
Not present | Short with thick arms, very abundant, 14.7–18.7–23.7 µm; some very irregular, rhab-like, 13.5–17.8–23.1 µm | Long and thin arms, 18.9–30.7–41.5 µm | Plato Seamount (695 m depth) |
|
N. pomponiae sp. nov. (Holotype MNHN-IP-2008-233) |
Cup-rounded shape | 54 × 81 mm in size with a small pedicel, 23 mm in size; walls, 11 mm thick | 185–427–666 × 18–39–88 µm (n = 13) | Cladome: 157–274–374 µm; rhabdome: 239–478–684 × 11–21–37 µm (n = 17) |
Large, thick, 1,455–1,643 × 17–18 µm (n = 2) | Very abundant, 10.7–18.9–35.8 µm | 16.2–27.6–39.3 µm | Hyères Seamount (480 m depth) |
Notes:
‘–’ no information/not mentioned.
Urn:lsid:zoobank.org:act:8A516D9B-5351-47AF-8EC2-7EBC44166D35
Holotype. MNHN-IP-2018-84 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°38′N, 11°29.8′W, 605–675 m, Seamount 1 campaign).
Paratype. MNHN-IP-2018-85 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°38′N, 11°29.8′W, 605–675 m, Seamount 1 campaign).
Other material. MNHN IP-2018-86 (1988-09-24, Gorringe Seamount, beam trawl, DW21, 36°34.9′N, 11°28.4′W, 460–480 m, Seamount 1 campaign).
Comparative material examined. Neoschrammeniella bowerbankii (Johnson, 1863) (HBOM 003:00592, Madeira), N. bowerbankii (HBOM 003:00810, Madeira), N. piserai sp. nov. (MNHN-IP-2008-234, Plato Seamount), N. pomponiae sp. nov. (MNHN-IP-2008-233, Hyères Seamount).
Diagnosis. Cup-shaped Neoschrammeniella with rounded edges and smooth surfaces; dicranoclone desmas of vine-like appearance; irregular dichotriaenes.
Description (holotype MNHN-IP-2018-84). Massive, flattened cup-shaped, with a concave centre, 73 mm length, 29 mm high and 64 mm wide (Fig. 2A); top surface is smooth with some oxeas perforating the surface and several small openings evenly distributed; walls are rounded and thick, 14–17 mm wide; bottom surface is also smooth, full of little openings dispersed throughout the entire surface, 31–56 μm in diameter, and some oxeas (Fig. 2B); colour is light brown in ethanol; the smooth surfaces could indicate that these specimens were not attached to any substrate, and therefore had a free living mode (Fig. 2B).
Skeleton. Ectosomal skeleton composed of smooth dichotriaenes of variable shape and size, along with a dense layer of microscleres (Figs. 4A and 4B); long-shafted triaenes or under-developed dichotriaenes, can also be observed (Fig. 4E); choanosomal skeleton is made of an irregular and loose network of dicranoclone desmas (Figs. 4C and 4D), spirasters and metasters; oxeas can be observed crossing the skeleton and projecting the surface.
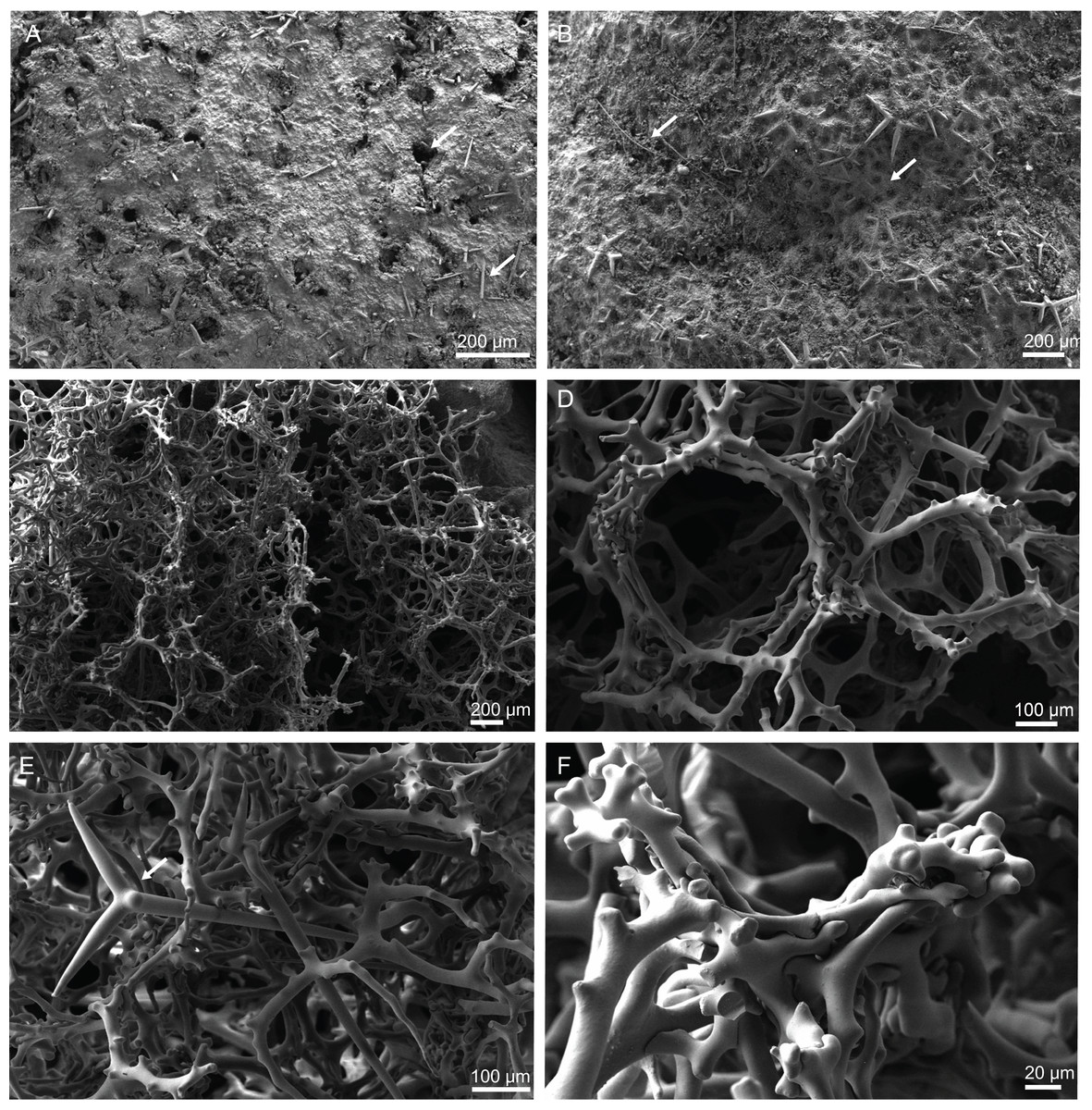
Figure 4: Surface and skeleton of Neoschrammeniella inaequalis sp. nov., holotype MNHN-IP-2018-84.
(A) Upper surface, showing the openings and some oxeas, (B) lower surface, showing oxeas and small openings, (C) overview of choanosomal desmas, (D) dicranoclones desmas, (E) plagiotriaenes crossing the desmas, (F) detail of the ornamentation of the desmas and zygosis.{kind=link}
Spicules (holotype MNHN-IP-2018-84).
Dicranoclones, smooth, irregular, slender, of vine-like appearance, 354–576–975 × 12–25–39 μm in size; clones can have few to several tubercles, that are smooth or slightly rugose (Figs. 4C–4F);
Oxeas, large, thin, curved, 670–1,144 × 5.2–7.8–13.4 μm in size (Figs. 4A and 4B);
Dichotriaenes, have a smooth cladome, that can be very irregular, having rounded or pointed tips, or clades of unequal size, 118–233–406 μm in diameter (Figs. 5A–5D); rhabdome is either short or long, and has a rounded tip, 136–432–1,211 × 9–18–31 μm in size (Fig. 5A); small branches or protuberances can be observed on the rhabdome, but they are uncommon (Fig. 5B);
Spirasters, with short and thick arms, mainly spiny on the arms, 12.1–18.5–26.6 μm in size (Figs. 5E–5H);
Metasters, less abundant, covered by spines, with long and thin arms, 14.6–31.6–47.9 μm in size (Fig. 5I).
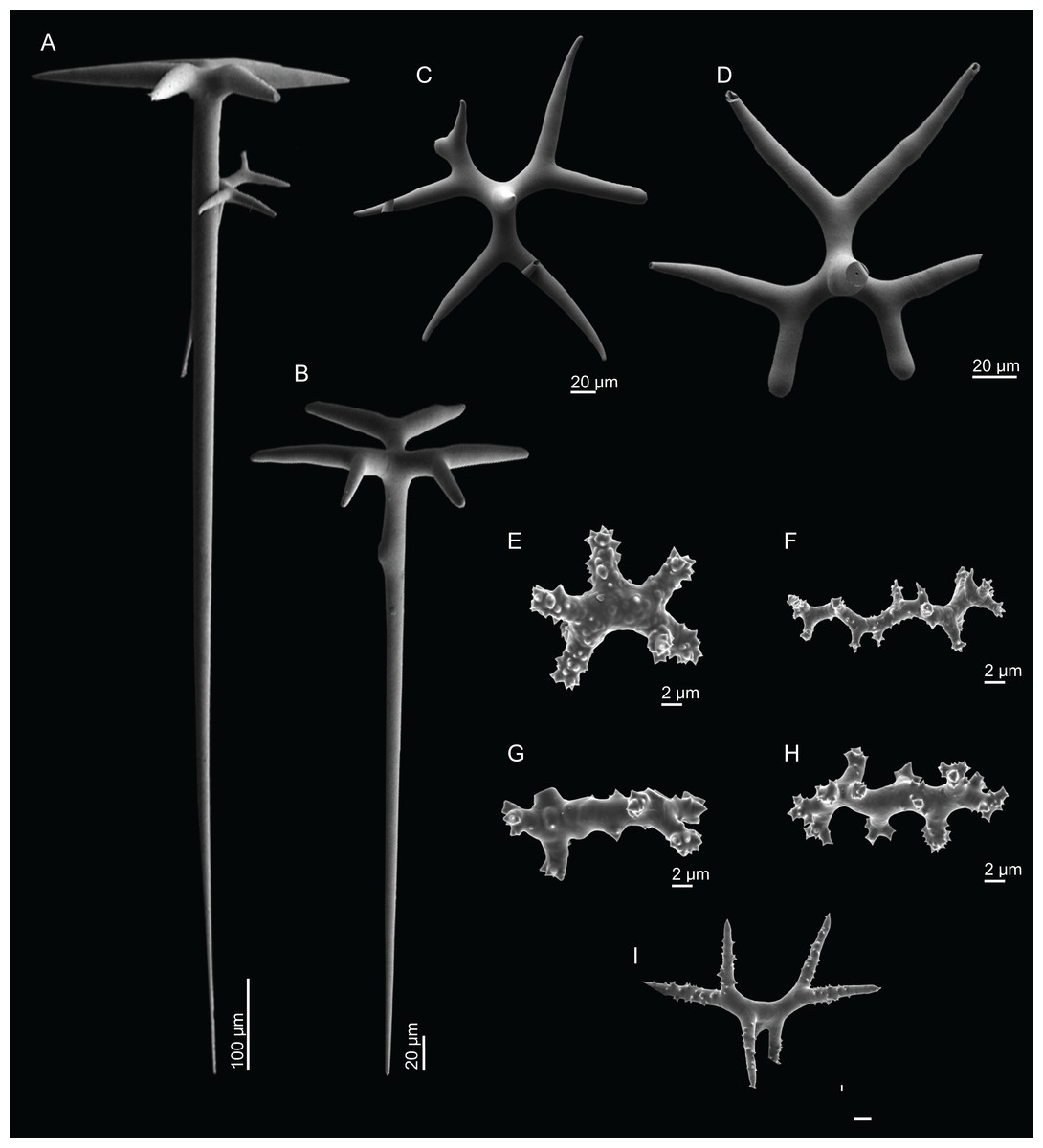
Figure 5: Spicules of Neoschrammeniella inaequalis sp. nov., holotype MNHN-IP-2018-84.
(A) Two dichotriaenes with different size classes, (B) small dichotriaene with a protuberance in the rhabdome, (C) and (D) irregular cladomes, (E)–(H) variation of spirasters, (I) metaster.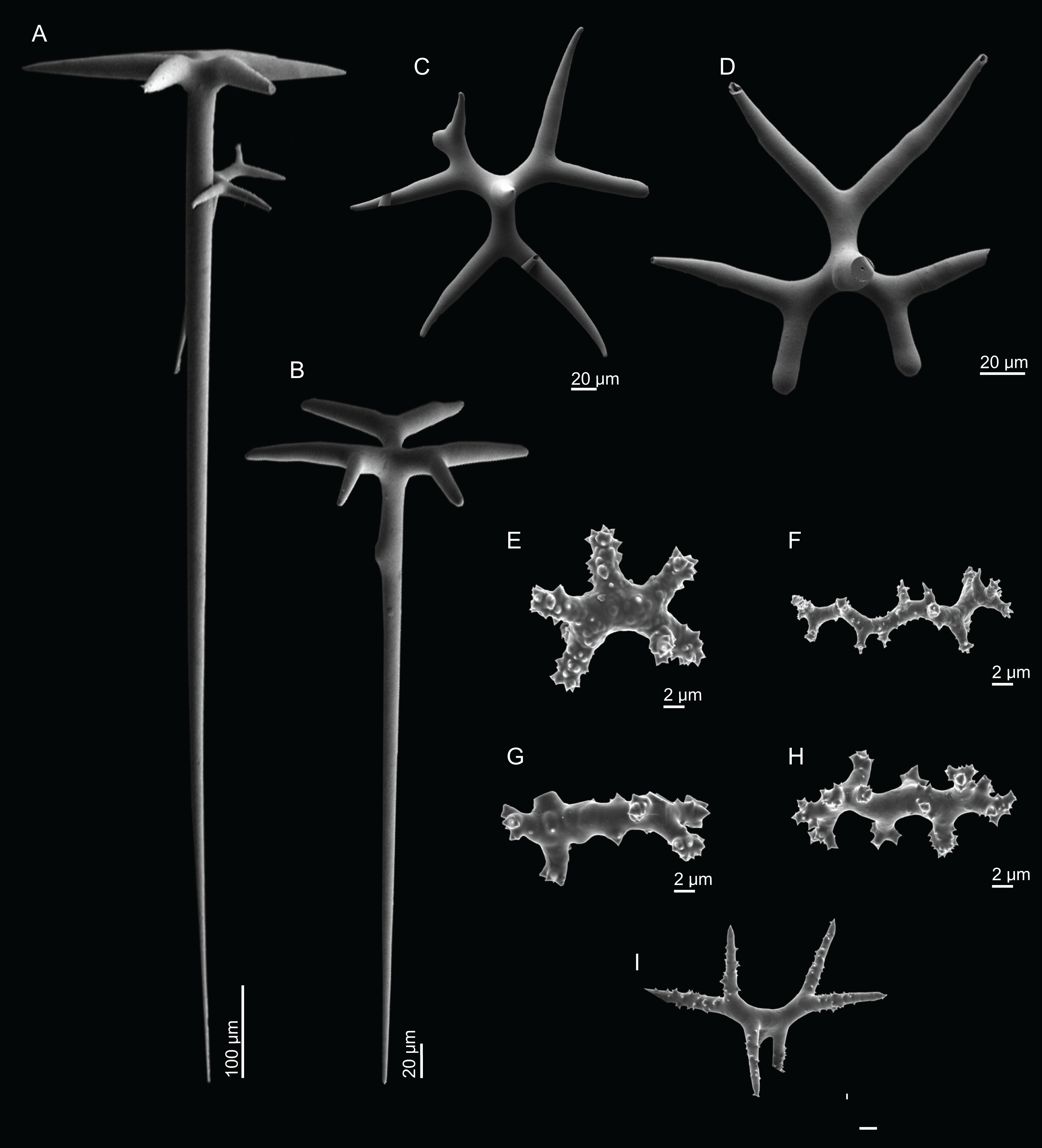{kind=link}
Distribution. N. inaequalis sp. nov. was found in the Gorringe Seamount between 460 and 675 m depth.
Etymology. From the latin inaequalis = unequal, due to the uneven and irregular cladomes of the dichotriaenes.
Remarks. N. inaequalis sp. nov. is a distinct species due to (1) the growth form, being flattened cup-shaped with a concave center; (2) the fact that both surfaces were completely smooth may indicate that the sponge is free-living, i.e., not attached to the substrate; (3) triaenes can be present, although rare, being the second time this kind of spicule is reported for the genus (see illustration of the redescription of N. moreti (Lévi & Lévi, 1988)) in Systema Porifera (Pisera & Lévi, 2002b); (4) the vine-like desmas also resemble the desmas found in the genus Isabella (Carvalho, Pomponi & Xavier, 2015; Ekins et al., 2016; Schlacher-Hoenlinger, Pisera & Hooper, 2005); (5) the shape and ornamentation of desmas are distinct from the other Neoschrammeniella species (see descriptions below and Remarks under N. pomponiae sp. nov.). It is also important to note that this species presents dichotriaenes very variable in size and shape (cladomes are irregular and unequal, and rhabdomes can present small protuberances or branches), so far only found in Isabella spp. (Carvalho, Pomponi & Xavier, 2015; Schlacher-Hoenlinger, Pisera & Hooper, 2005). These irregularities can be attributed to a pathologic development.
Neoschrammeniella piserai sp. nov.
Figures 2C, 6–7 and Table 1
Urn:lsid:zoobank.org:act:77F1F52E-28C9-43C0-A501-1ADAD03241A5
Holotype. MNHN-IP-2008-234 (1993-01-31, Plato Seamount, epibenthic Warén dredge, DW241, 33°12′N, 28°59′W, 695 m, Seamount 2 campaign).
Comparative material examined. N. bowerbankii (HBOM 003:00592, Madeira), N. bowerbankii (HBOM 003:00810, Madeira), N. inaequalis sp. nov. (holotype MNHN-IP-2018-84 and paratype MNHN-IP-2018-85, Gorringe Seamount), N. pomponiae sp. nov. (holotype MNHN-IP-2008-233, Hyères Seamount).
Diagnosis. Cup rectangular shaped Neoschrammeniella fixed to the substratum by the entire base; oxeas not present.
Description (holotype MNHN-IP-2008-234). Large cup-rectangular sponge, 98 mm height and 69 mm width on top; the sponge was attached to the substratum by the entire base, which has 43 mm in diameter; walls are 11 mm thick (Fig. 2C); surfaces are smooth with visible subdermal water canals and openings evenly distributed on both surfaces, 20–44 μm in diameter (Figs. 6A and 6B), colour beige in ethanol.

Figure 6: Surface and skeleton of Neoschrammeniella piserai sp. nov., holotype MNHN-IP-2008-234.
(A) Overview of the surface with several openings, (B) close up of the surface where dichotriaenes are surrounded by a large number of microscleres, (C) overview of the skeleton showing the separation of the ectosome, made by a layer of dichotriaenes, and the choanosome composed of desmas, (D) dicranoclone desmas, (E) detail of dicranclone desmas, (F) zygosis and detail on the sculpture of the desmas.{kind=link}
Skeleton. Ectosomal skeleton is made of a layer of dichotriaenes perpendicular to the surface, and a dense layer of numerous microscleres (Fig. 6C); choanosomal skeleton has a net of compact dicranoclone desmas with several metasters and spirasters spread out through the tissue.
Spicules (holotype MNHN-IP-2008-234).
Dicranoclones, irregular, usually smooth, 280–428–522 × 16–25–37 μm in size; the rays of the desmas have several ramifications and some tubercles, that are usually smooth (some can have a rugosity) (Figs. 6C–6F).
Dichotriaenes, with a smooth cladome, 153–244–389 μm in diameter; rhabdome has a rounded tip and 198–366–535 × 10–19–33 μm in size (Fig. 7A).
Metasters, covered by spines, with long and thin arms, 18.9–30.7–41.5 μm in size (Figs. 7E–7G).
Spirasters, spiny, with short and thick arms, very abundant, 14.7–18.7–23.7 μm in size (Figs. 7B–7D); some can present an irregular shape, i.e., rhabd-like with spiny tips, scarce, 13.5–17.8–23.1 μm in size (Figs. 7H–7J) (see “Remarks”).
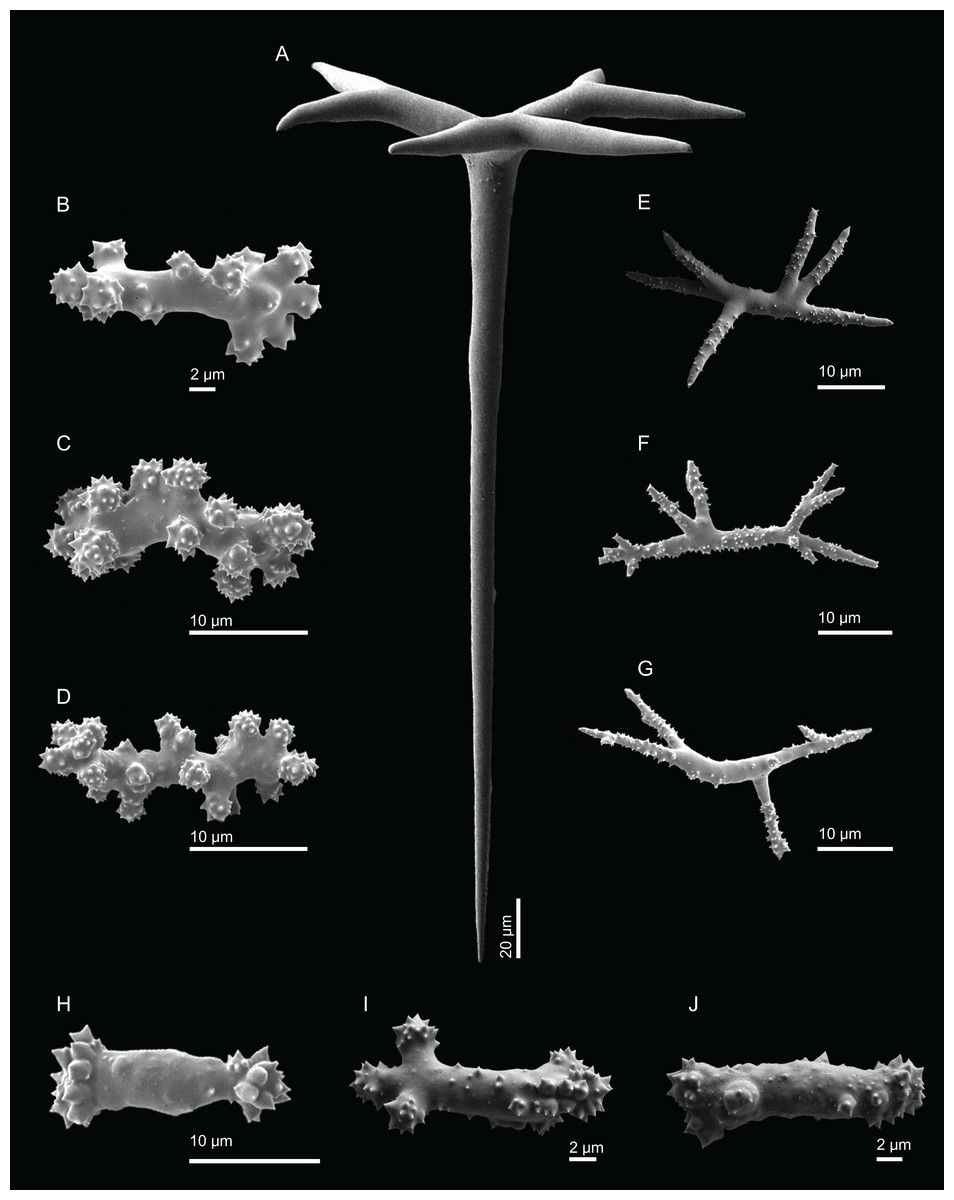
Figure 7: Spicules of Neoschrammeniella piserai sp. nov., holotype MNHN-IP-2008-234.
(A) Dichotriaene, (B)–(D) spirasters, (E)–(G) mestasters, (H)–(J) underdeveloped spirasters.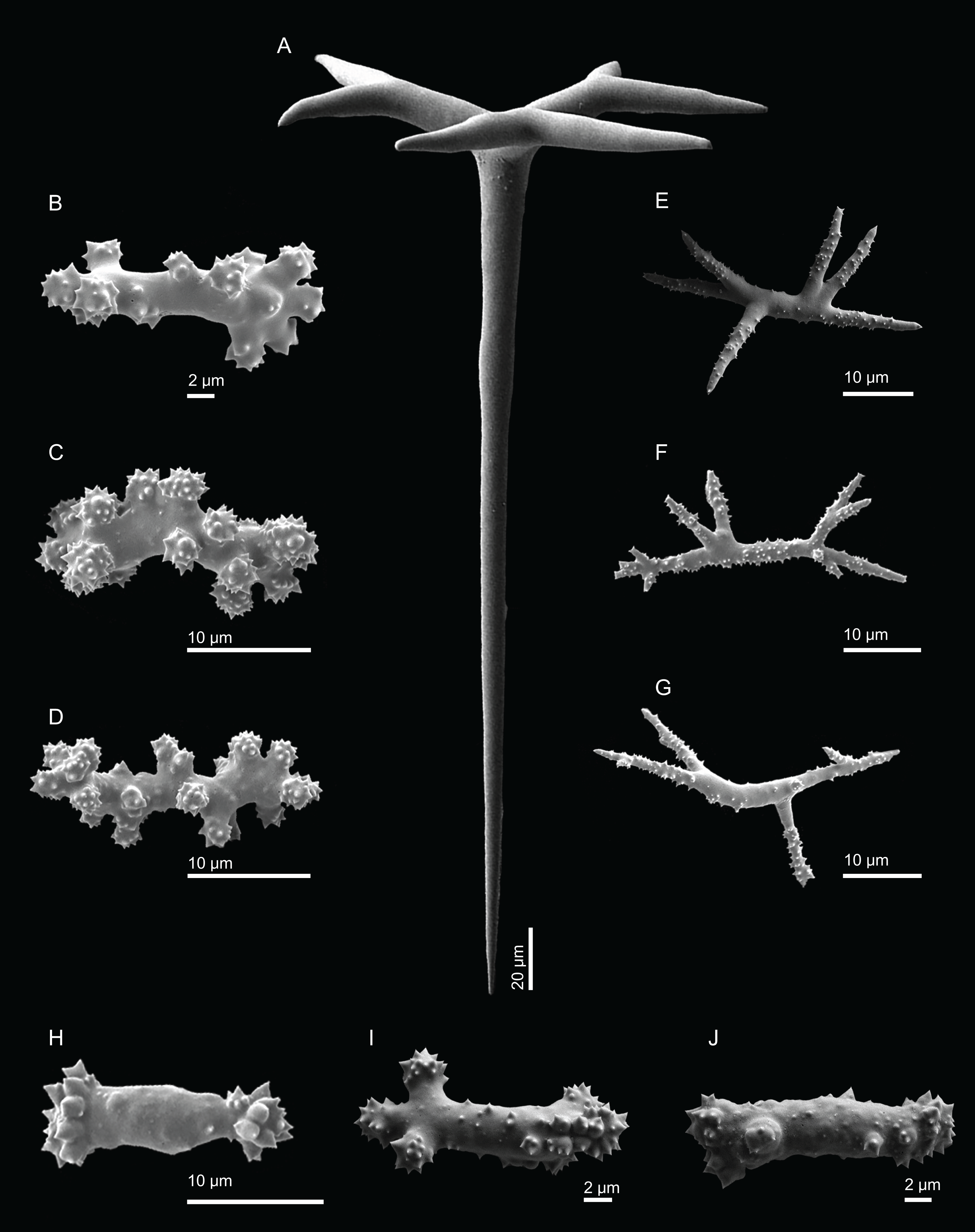{kind=link}
Distribution. N. piserai sp. nov. is only known from its type locality, Plato Seamount (695 m depth).
Etymology. Named after Professor Andrzej Pisera from the Institute of Paleobiology Warszawa (ZPAL), in recognition of his outstanding contributions on the taxonomy of both fossil and extant lithistid sponges.
Remarks. The peculiar external morphology (cup-rectangular shape) of N. piserai sp. nov., together with the smooth surface, the ornamentation of the desmas are the features that differentiate this new species from the other NEA and MED Neoschrammeniella species (Table 2; Remarks under N. pomponiae sp. nov.). One could also not observe oxeas on this species, a spicule type that was found in other Neoschrammeniella spp. from the NEA and MED. Some spirasters presented an irregular shape. They were rhabd-like with spiny tips (Figs. 7H–7J) and they had approximately the same size as the typical spirasters. Since these underdeveloped spirasters were scarce we decided to include them in the same category of spirasters, but analyses of new material may show that they belong to a different category.
| Habitus | Size | Tetraclones | Discotriaenes | Oxeas | Acanthomicroxeas | Acanthorhabds | Locality | |
|---|---|---|---|---|---|---|---|---|
| 1D. polydiscus (Bowerbank, 1869) (Holotype BMNH 40.10.23.12) | Small irregular mushroom shaped, with strongly concave upper side; short stem and slightly expanded attachment base | 25 × 20 mm large, 18 mm high | Regular massive with strongly branched and tuberculated zygomes and smooth rays; 300–450 µm in size and 100–110 µm thick | Cladome: round to oval, 250–350 µm in diameter; rhabdome: short and conical, 87–108 µm | Present | Slender, fusiform and slightly curved or bent acanthoxeas (spines are hook-like), 38–59 µm long, 2.4–4 µm thick | Fusiform, massive, 15– 22 µm long, 2–4.5 µm thick | St. Vincent Island, Caribbean (depth unknown) |
| D. inscripta (Schmidt, 1879) (unknown type) | 2Incertae sedis (type material is deciduous: ectosomal discotriaenes and microscleres were not found) | |||||||
| 3D. dissoluta Schmidt, 1880 (HBOM 003:01093) | Cluster of knobby fingers; colour is purple brown in exterior and cream-coloured in interior when alive | 200 mm diameter, 50 mm tall and 10 mm in diameter | Smooth, regular, with a weak zygosis, 475–525 µm in size | Cladome: round, concave, smooth (except growth lines), 203–294 µm in diameter; rhabdome: short, delicate and conical | Curved oxeas/styles, 500–530 × 9–10 µm | Fusiform, 41.6–68.0 × 5.5–6.1 µm in size | Fusiform with pointed tips, 15.1–18.9 × 4.3–5.2 µm in size | Florida (81 m depth) |
| 4D. ramifera Topsent, 1892 (Holotype) | Sponge more and less elongated with several finger-like extensions; water canals visible under the surface; smooth surface; colour is white in ethanol | 1–15 mm wide; finger-like extensions 2–20 × 2–3 mm | Desmas rays full of tubercles in the extremities | Whole or barely lobed, 300 µm diameter | Present | Numerous, fusiform, spiny, curved, seldom centrotylotes, 40–45 µm long | Very abundant, thorny, often curved, 20–25 µm long | Azores (318 m depth) |
| D. ramifera (specimen MNHN-IP-2008-213) | Small, elongated to branching shape sponge; colour is beige to light yellow | 15–29 mm high and 3–10 mm thick | 182–328–470 × 24–32–48 µm in size (n = 19) | Cladome: very variable in shape, 124–160–213 µm in diameter (n = 16); rhabdome: 23–32–40 × 8–10–14 µm (n = 9) | Present (all broken) | Slightly curved, thorny, 22.8–27.6–32.6 × 1.0–1.5–1.8 µm (n = 15) | Thorny with blunt tips, 3.9–10.3–13.9 × 1.1–1.4–1.9 µm (n = 19) | Great Meteor Seamount (320 m depth) |
| D. cf. ramifera (specimen MNHN-IP-2008-210) | Small, elongated; colour is beige | 20 high and 10 mm thick (fragment) | 400–455–534 × 30–51–82 (n = 20) | Cladome: 195–328–560 µm; rhabdome: 20–42–68 × 9.5–20.3–37.9 µm (n = 16) | Present | 24.6–39.0–59.8 × 1.8–3.3–5.4 µm | 15.2–20.2–24.2 × 2.1–2.9–4.4 µm | Atlantis Seamount (420 m depth) |
| 5D. verrucosa Topsent, 1928 (Holotype MNHN DT 1199) | Cup-shaped with rounded edges and numerous warts; irregular contour and a depressed center; short pedicel laterally compressed; colour is grey-yellow in ethanol | 35–38 mm high and 58 mm wide | Skeleton is very solid and regular, desmas are robust and have a complex zygosis; Protoclad with tubercles and 60 µm; deuteroclad has several cylindrical nodules intended for zygosis |
Cladome: flat, variable shapes, 360–400 µm on average (can vary between 200 and 560 µm); Rhabdome: conical shape, simple, 100 µm long |
Slightly curved, bigger than 1 mm, rarely exceeding 7 µm width | Numerous, fusiform, spiny, slightly sharp, 43–52 × 3–3.5 µm | More abundant than microxeas, 15–17 × 2–2.8 µm | Gran Canaria (400 m depth) |
| D. verrucosa (specimen MNHN-IP-2008-205) | Spherical polymorphic with several rounded protuberances; colour varies from whitish to light brown | 15–20 high and 12–13 mm wide | 106–170–278 (n = 19) × 19–34–46 µm in size | Cladome: 102–153–222 µm in diameter (n = 17); rhabdome: 15–25–47 × 5–8–13 µm (n = 9) | Broken | 22.8–35.2–53.5 × 1.3–2.2–3.9 µm | 7.5–12.9–19.0 × 1.2–1.6–3.0 µm | Atlantis Seamount (338 m depth) |
| 6D. polymorpha Pisera & Vacelet, 2011 (Holotype ZPAL Pf.21/1) | Small and polymorphic, nearly spherical to irregular masses with protuberances; can be attached to the by a short pedicel or the entire surface | Up to 57 mm in diameter | Irregular skeleton; desmas are smooth with poorly branches tips, 370–718 µm in diameter | Cladome: very variable in shape, 174–366 µm in diameter; rhabdome: 60–65 µm long |
Not present | Spinous, very variable, 24.8–68.3 × 1.66–3.78 µm | Very variable, cylindrical to fusiform, 13.20–37.20 × 1.85–4.25 µm | 3PPs Cave, Marseille area, France (3–20 m depth) |
| 7D. adhaerens Van Soest, Meesters & Becking, 2014 (Holotype RMNH Por. 9241) | Thinly to massively encrusting limestone rockwalls with a smooth surface; colour is bright orange | Several dm2 in lateral expansion, 2–3 mm thick | Large, robust, with arms heavily tuberculated, 320–428–520 µm long and 40–66 µm thick | Discs: 130–202–350 × 100–155–280 µm; Rhabds 24–34–41 µm | Thin, curved, with wispy endings, 670–795–910 × 5–6.3–7 µm | Not present | 15–20–25 µm (as acanthomicrorhabds) | Bonaire (146 m depth) |
| D. arbor sp. nov. (Holotype MNHN-IP-2008-211) | Massive discodermia of tree like appearance, with a long stem and three branches; surface is smooth; colour is beige in ethanol | Full sponge length is 58 mm; stem is 15 mm high and 7.5–12 mm wide and branches are 13–28 mm long | Usually with the arms tuberculated, but can be smooth; very strong zygoses;181–392–567 × 15–36–56 µm in size | Cladome: 148–256–396 µm in diameter; rhabdome: 34–53–71 × 15–21–24 µm (n = 9) | Not present | Slightly curved, spinous, with sharp tips, 24.1–35.1–50.1 × 1.4–2.3–3.5 µm | Covered by numerous spines, with unequal tips (blunt or sharp) 6.7–16.1–25.9 × 1.1–2.2–4.3 µm | Great Meteor Seamount (330 m depth) |
| D. kellyae sp. nov. (Holotype MNHN-IP-2008-208) | Massive sponge, polymorphic of bulb appearance, with large protuberances of round shape; surface is irregular with a crumble/rugose appearance; colour is beige to light brown | 53 mm high and 31 mm wide | Large, compact, thick, 112–338–589 × 20–42–76 µm (n = 20) | Cladome: very variable in shape and size, 121–289–425 µm in diameter; rhabdome 36–81–142 × 13–31–44 µm | Strongyles, one tip rounded and the other one sharp, 418–444 × 6.0–7.9 µm in size (n = 2) | Straight or curved, with sharp tips, spinous, 16.7–43.2–66.5 × 1.5–2.5–3.7 µm | Spinous, with blunt tips, 5.3–13.3–24.9 × 1.2–2.1–3.7 µm | Plato Seamount (580 m depth) |
Notes:
Neoschrammeniella pomponiae sp. nov.
Figures 2D, 8–9 and Table 1
urn:lsid:zoobank.org:act:2AA76193-B27E-491E-8E50-FE591786FA26
Holotype. MNHN-IP-2008-233 (1993-01-16, Hyères Seamount, epibenthic Warén dredge, DW182, 31°23′N, 28°54′W, 480 m, Seamount 2 campaign).
Comparative material examined. N. bowerbankii (HBOM 003:00592, Madeira), N. bowerbankii (HBOM 003:00810, Madeira), N. inaequalis sp. nov. (holotype MNHN-IP-2008-84 and paratype MNHN-IP-2018-85, Gorringe Seamount), N. piserai sp. nov. (holotype MNHN-IP-2008-234, Plato Seamount).
Diagnosis. Neoschrammeniella with a cup-rounded shape and a rugose surface, fixed to the substratum by a small pedicel; dicranoclones are densely covered by numerous and ornamented tubercles with a rugose appearance.
Description (holotype MNHN-IP-2008-233). Large sponge, 54 mm height and 81 mm in diameter, with a small pedicel 23 mm wide; its external morphology resembles a bowl; walls are about 11 mm thick; the surfaces of the sponge are rugose, and hispid due to oxeas protruding the surface; openings are small and evenly spread on both surfaces, 40–87 μm in diameter; colour is brown in ethanol (Fig. 2D).
Skeleton. Ectosome is composed of a layer of dichotriaenes perpendicular to the surface that is covered by various microscleres (Figs. 8A and 8B); choanosome composed of a dense mesh of dicranoclone desmas, oxeas crossing the choanosome protruding the surface (Fig. 8A), and several microscleres spread through the skeleton.

Figure 8: Surface and skeleton of Neoschrammeniella pomponiae sp. nov., holotype MNHN-IP-2008-233.
(A) Surface showing several openings, dichotriaenes and some oxeas protruding the surface, (B) detail of the surface with a dichotriaene surrounded by numerous microscleres, (C) overview of the dicranoclone desmas, (D) choanosomal dicranoclone desmas, (E) detail of the sculpture of the desmas, (F) zygosis.{kind=link}
Spicules (holotype MNHN-IP-2008-233).
Dicranoclones, compact, irregular and with the clones very tuberculated, 185–427–666 × 18–39–88 μm in size; rays of desmas are covered by numerous and ornamented tubercles that have a rugose appearance (Figs. 8C–8E); clones articulated into complex and intricate zygoses (Fig. 8F);
Oxeas, long, with sharp tips, 1455–1643 × 17–18 μm in size (Fig. 8A);
Dichotriaenes, with a smooth cladome, 157–274–374 μm in diameter and a long rhabdome with a blunt tip, 239–478–684 × 11–21– 37 μm in size (Fig. 9A);
Spirasters, very abundant, irregular, spiny, with short and thick arms, 10.7–18.9–35.8 μm in size (Figs. 9B–9E).
Metasters, less abundant, spiky, with long and thin arms, 16.2–27.6–39.3 μm in size (Figs. 9F–9I).
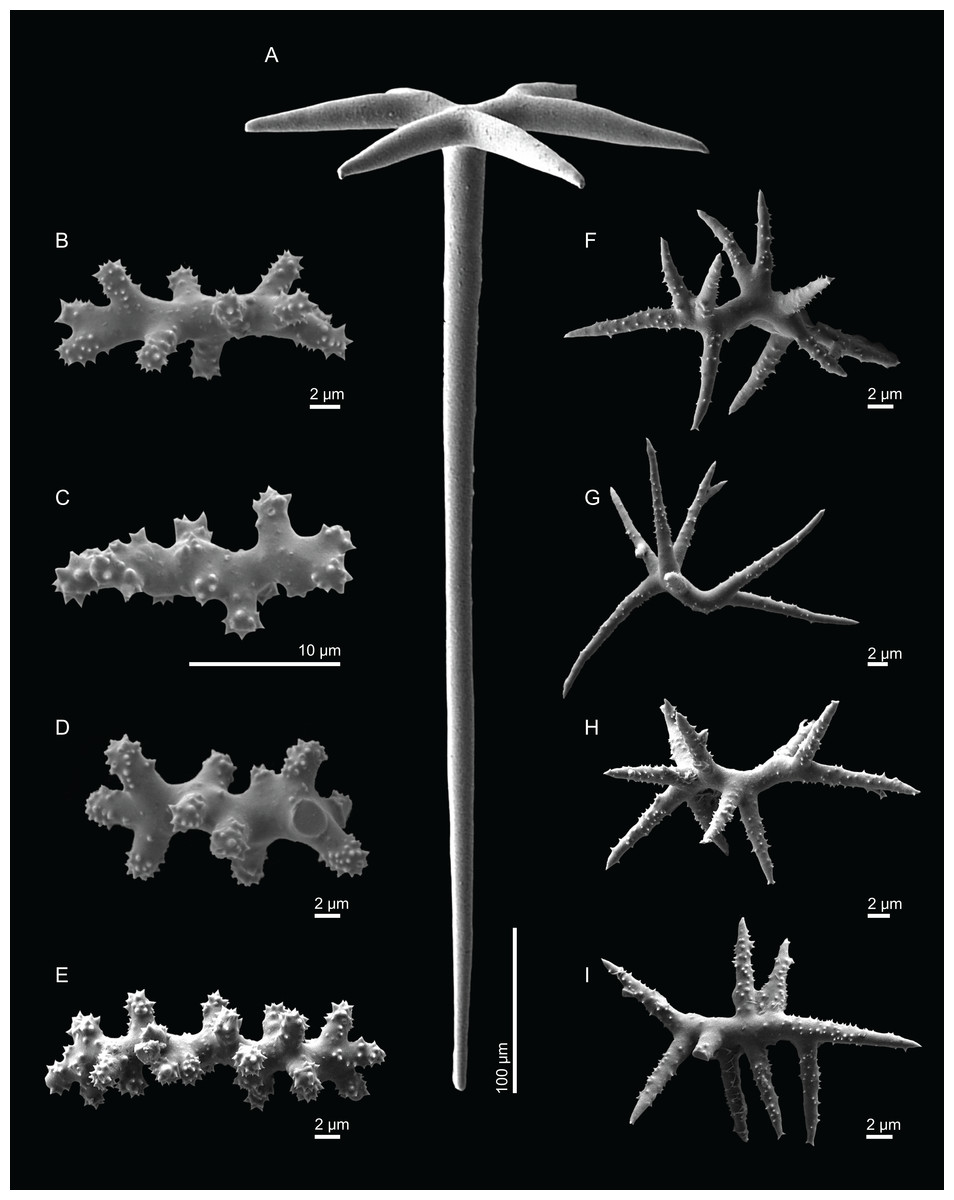
Figure 9: Spicules of Neoschrammeniella pomponiae sp. nov., holotype MNHN-IP-2008-233.
(A) Smooth dichotriaene, (B)–(E) spirasters, (F)–(I) metaster.{kind=link}
Etymology. Named after Dr. Shirley Pomponi from the Harbour Branch Oceanographic Institute (HBOI) in recognition of her valuable contributions to the knowledge of deep-sea sponges (including lithistids) of the North-western Atlantic Ocean and Caribbean.
Distribution. N. pomponiae sp. nov. is known from its type locality, Hyères Seamount, where it was collected at 480 m depth.
Remarks. The genus Neoschrammeniella was erected by Pisera & Lévi (2002b) to accommodate Corallistidae with smooth dichotriaenes and two to three types of microscleres. This genus is widely distributed, with records spanning the Southern Ocean, SW Pacific, Mediterranean Sea and NEA. Until now, six species were described and only one, N. bowerbankii (Johnson, 1863), was known to occur in the Mediterranean Sea (Pisera & Vacelet, 2011) and the NEA in the Madeira archipelago (Carvalho & Pisera, 2019; Johnson, 1863). In the present work, we described and illustrate three new species of Neoschrammeniella, that can mainly be distinguished by their habitus, sculpture of the desmas, presence or absence of oxeas, and, shape and size of the dichotriaenes. The external morphology of N. pomponiae sp. nov. resembling a bowl, contrasts with the cup-shaped to contorted lamellate masses with thick walls in N. bowerbankii, the flattened cup-shaped with a concave centre in N. inaequalis sp. nov. and the large cup-rectangular shape in N. piserai sp. nov. The sculpture of the desmas is also very distinct among all these species, while N. bowerbankii has very tuberculated dicranoclones divided into smaller and irregular lobes/tubercles (redescription in Pisera & Vacelet, 2011), N. inaequalis sp. nov. presents a distinct shape of desmas with vine-like appearance and few to several tubercles, N. piserai sp. nov. has irregular and compact dicranoclones that are usually smooth, and N. pomponiae sp. nov. has desmas densely covered by numerous and ornamented tubercles with a rugose appearance. Finally, N. inaequalis sp. nov. is the only one with very variable dichotriaenes either in size and shape, while N. piserai sp. nov. does not have oxeas, a type of megasclere present in the other three species.
Family Theonellidae Lendenfeld, 1903
Genus Discodermia du Bocage, 1869
Synonymy. Collinella Schmidt, 1879 (junior synonym); Desmahabana Alcolado & Gotera, 1986 (junior synonym).
Diagnosis. Theonellidae with discotriaenes exclusively as ectosomal megascleres and choanosomal tetraclone desmas; microscleres are acanthoxeas and acanthorhabds.
Definition. Polymorphic sponges, from massive irregular to cup-shaped, branched or cylindrical; ectosomal megascleres are smooth discotriaenes; choanosomal megascleres are tetraclone desmas (regular or irregular) that can be smooth or tuberculated, and oxeotes or stylotes; microscleres are acanthoxeas and acanthorhabds (Kelly, 2007; Pisera & Lévi, 2002c; Pisera & Vacelet, 2011).
Type species. Dactylocalyx polydiscus Bowerbank, 1869.
Discodermia ramifera Topsent, 1892
Figures 2E, 10–11 and Table 2
Material examined. MNHN-IP-2008-204 (1993-01-09, Meteor Seamount, beam trawl, CP138, 30°02′N, 28°29′W, 300 m), MNHN-IP-2008-207 (1993-01-10, Great Meteor Seamount, epibenthic dredge, DE140, 30°01′N, 28°28′W, 308 m), MNHN-IP-2008-213 (1993-01-11, Great Meteor Seamount, beam trawl, CP156, 29°56′N, 28°24′W, 320 m), MNHN-IP-2008-214 (1993-01-10, Great Meteor Seamount, beam trawl, CP144, 30°10′N, 28°29′W, 335 m). All from the Seamount 2 campaign.
Comparative material examined. Discodermia verrucosa Topsent, 1928 (MNHN-IP-2008-205, Atlantis Seamount; MNHN-IP-2008-206, Plato Seamount; HBOM 003:00869, Madeira; HBOM 003:00870, Madeira; HBOM 003:00868, Selvagens; HBOM 003:00640, Canary Islands; RMNH6237, Selvagens), D. kellyae sp. nov. (holotype MNHN-IP-2008-208, Plato Seamount), D. arbor sp. nov. (holotype MNHN-IP-2008-211, Great Meteor Seamount).
Diagnosis. Small Discodermia, elongated to branching in shape, with smooth tetraclone desmas.
Description (MNHN-IP-2008-213). Elongated and branched, small sponge, 15–29 mm high and 3–10 mm thick (Fig. 2E); surface is smooth and transparent, where it is possible to see the subdermal water canals, that gives a striated appearance to the sponge when observed under a magnifier; openings form a small elevation on the sponges’ surface; colour is beige to light yellow in ethanol.
Skeleton. Ectosome is composed of a layer of overlapping discotriaenes and abundant microscleres such as acanthomicroxeas and acanthorhabds, spread through this part of the skeleton; choanosomal skeleton has tetraclone desmas (Fig. 10), smooth oxeas and some microscleres spread through the entire sponge; desmas form an irregular and compact net on the choanosome but a loose mesh near the ectosome with big spaces between them; oxeas can be observed crossing the interior of the skeleton.
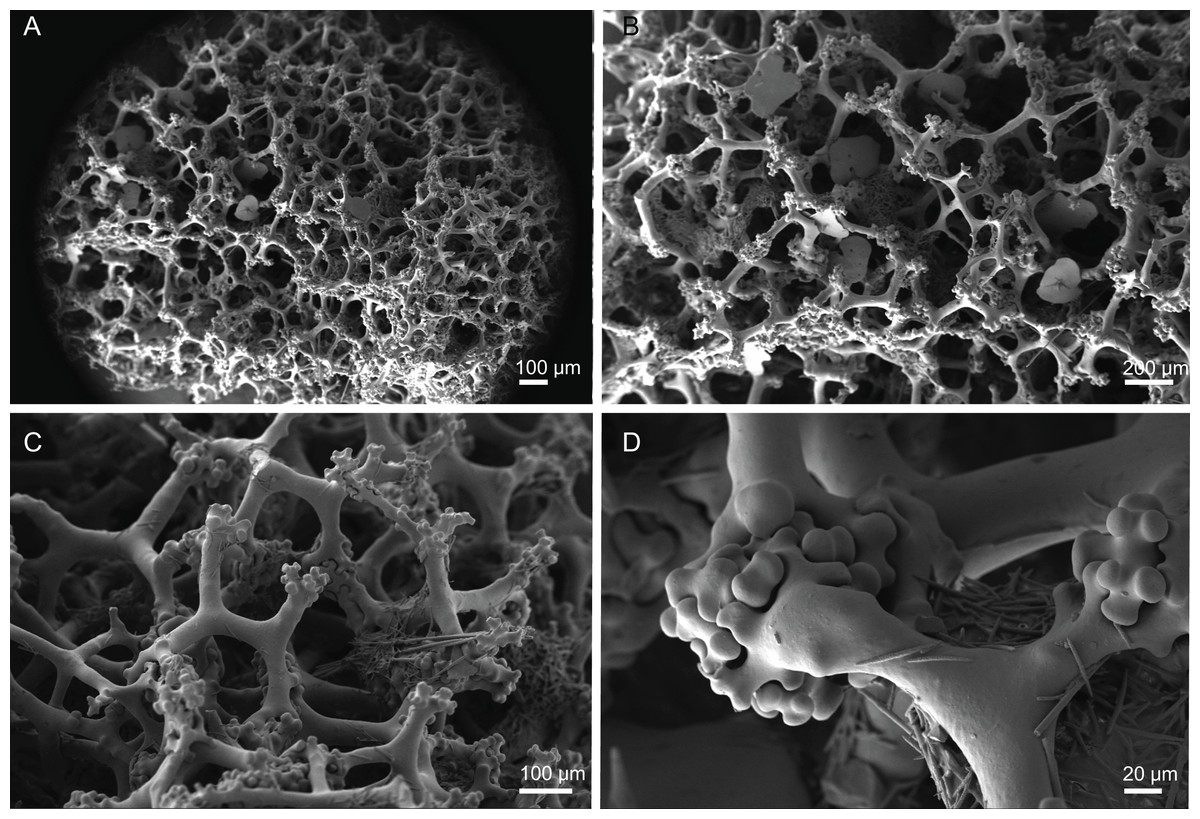
Figure 10: Skeleton of Discodermia ramifera Topsent, 1892, specimen MNHN-IP-2008-213.
(A) Overview of choanosomal desmas, (B) tetraclone desmas and some discotriaenes, (C) detail of the smooth tetraclone desmas with tubercles in the zygome, (D) zygosis.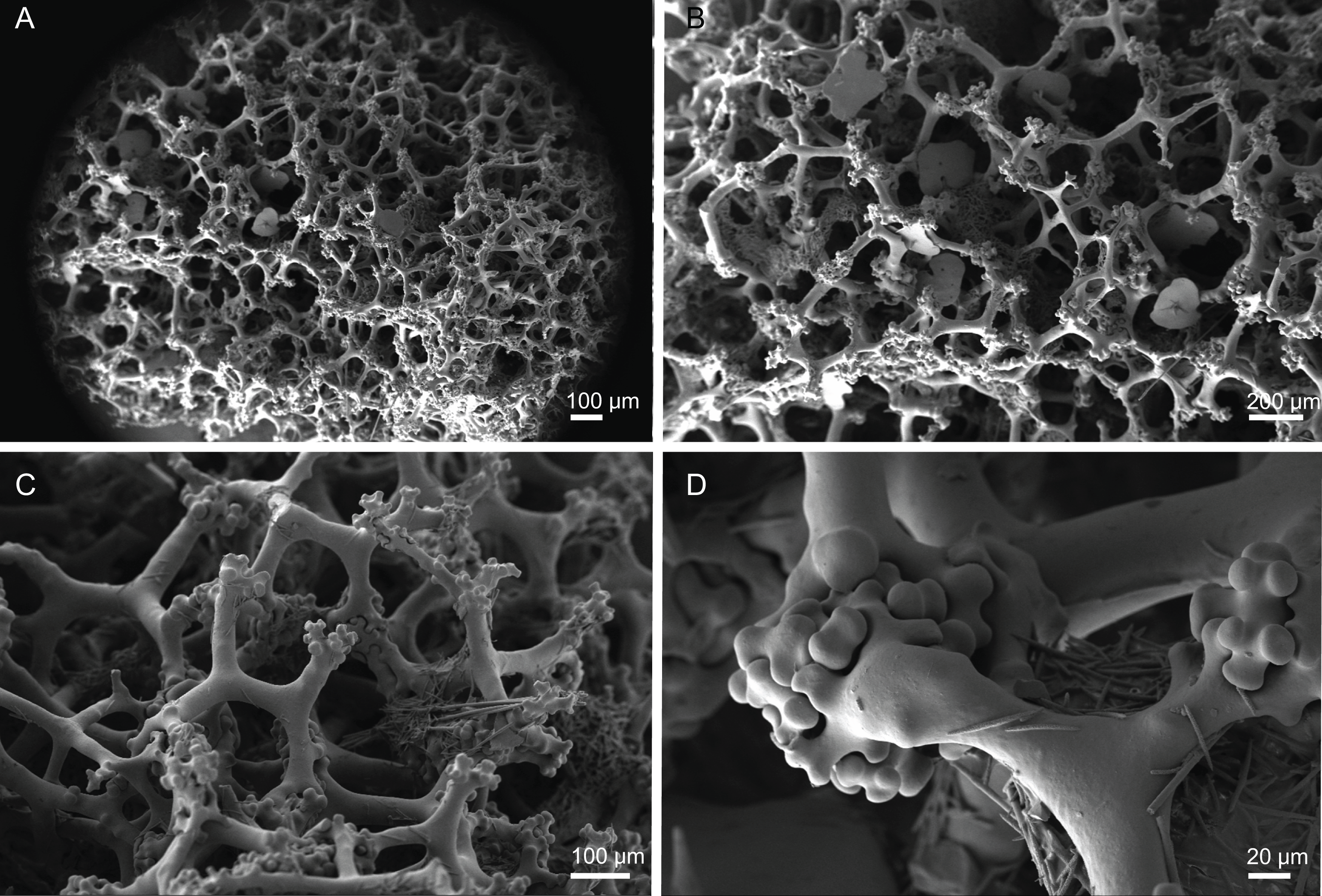{kind=link}
Spicules (MNHN-IP-2008-213).
Tetraclone desmas, with smooth rays (Figs. 10A–10C) and tuberculated zygoses (Fig. 10D); tubercles are generally smooth but in some cases one tubercle may be divided into various smaller tubercles; tetraclones are 182–328–470 × 24–32–48 µm in size;
Discotriaenes, very variable in shape, from round/oval to irregular and indented cladome; cladome can be flat or slightly concave, 124–160–213 µm diameter; rhabdome, short and conical, 23–32–40 µm × 8–10–14 µm in size (Figs. 11A–11D).
Oxeas, long, smooth with rounded extremities (Fig. 10C); the vast majority of oxeas were broken, thus measurements of these megascleres are not presented here.
Acanthomicroxeas, slightly curved with pointed ends, rarely centrotylotes, 23–28–33 × 1.0–1.5–1.8 µm in size (Fig. 11E).
Acanthorhabds, similar to microxeas with the exception they are smaller and have rounded tips, 3.9–10.3–13.9 × 1.1–1.4–1.9 µm in size (Fig. 11F).
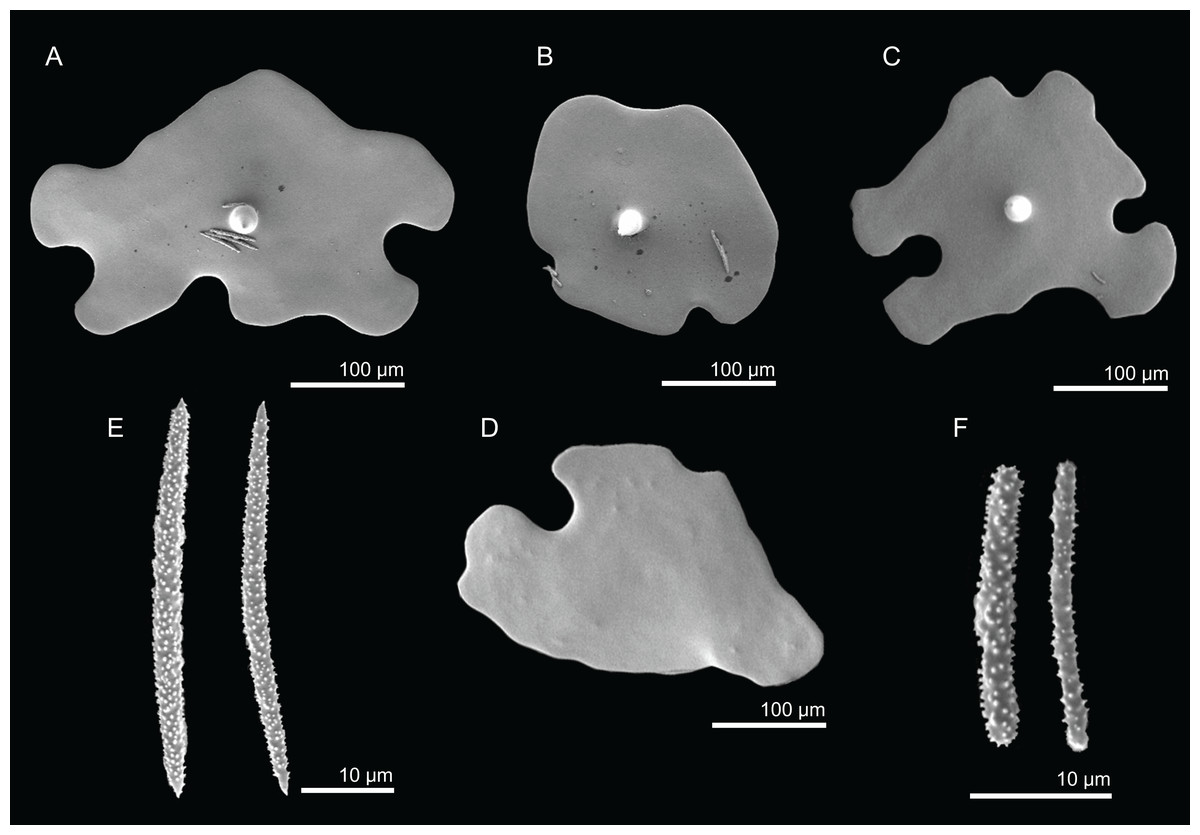
Figure 11: Spicules of Discodermia ramifera Topsent, 1892, MNHN-IP-2008-213.
(A)–(C) Lower view of discotriaenes, (D) top view of discotriaene, (E) acanthomicroxeas, (F) acanthorhabds.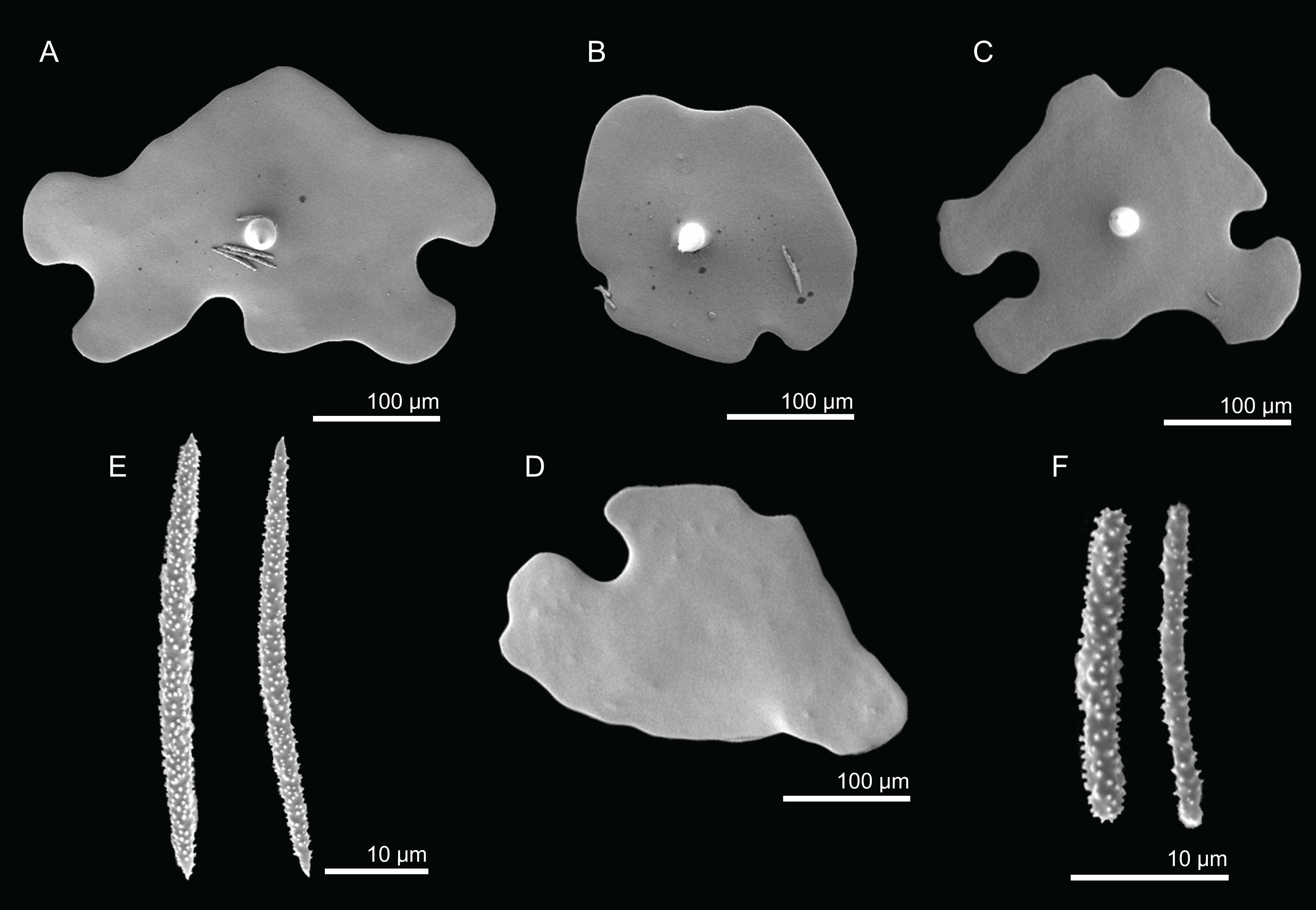{kind=link}
Distribution. Specimens were collected at the Great Meteor Seamount between 300 and 335 m depth.
Remarks. D. ramifera was described by Topsent (1892) from material collected in the Azores (318 m depth), and later re-collected in the same archipelago at 98 m depth (Topsent, 1904). So far, these were the only records in the North Atlantic. Here we discover for the first time the presence of this species in the Great Meteor seamount (between 300 and 335 m depth). The specimens analysed in this work have a similar external morphology compared to the ones described by Topsent (i.e., small, elongated to branching sponge with finger-like extensions), and similar spicule composition. However, the spicules’ sizes are in general smaller from those presented in the original description (Table 2). Discotriaenes have a smaller cladome, 124–213 µm in the analysed material versus the 300 µm in diameter in the original description; acanthomicroxeas (22.8–32.6 µm vs 40–45 µm long) and acanthorhabds are also smaller (3.9–13.9 µm vs 20–25 µm long), but see Discussion for more details on these differences.
| Habitus | Size | Desmas | Phyllotriaenes | Oxeas | Microxeas | Locality | |
|---|---|---|---|---|---|---|---|
| 1Macandrewia azorica Gray, 1859 (Holotype BMNH 1851.7.28.16) | Cyathiform to flabellate, with a short stem and undulating rounded margins; outer surface smooth, with small, irregular but evenly distributed pores, 37–58 µm in diameter | 120 × 120 mm with a short stem, 30 mm long; walls 6–9 mm thick | Smooth, complex, strongly branched at the end with a loose terminal articulation, 255–438–724 × 8.5–19.0–30.8 µm in size (n = 22)* | Cladome: with strongly incised clades, 297–363–456 µm (n = 11) in diameter; rhabdome: conic and short, 157–163–167 × 17.5–19.9–22.2 µm in size (n = 3)* |
Small, fusiform and thick, 532–652–780 × 10.5–15.1–19.4 µm (n = 8)* | Very common, fusiform, 38–55–96 × 2.5–3.9–7.9 µm* | S. Miguel island, Azores (depth unknown) |
| M. cf. azorica (MNHN-IP-2008-220) | Flabellate to undulate masses with thin lamellas; smooth surfaces; colour beige to light brown | 67 × 50 mm in size; walls are rounded and undulate, 3–5 mm thick | 212–281–343 × 16–34–51 µm (n = 24) | Cladome: very indented, 194–267–333 µm (n = 20); rhabdome: 62–99–129 × 11.6–14.4–17.8 µm (n = 12) | 215–246–301 × 6.8–7.8–9.1 µm (n = 4) | 33.3–55.0–83.6 × 2.5–3.9–5.1 µm | Atlantis Seamount (420 m depth) |
| 2M. clavatella (Schmidt, 1870) (unknown type) | Obconic, seated on a short pedicel, summit flattened or depressed, or convexly rounded, bearing several oscules 0.25–1.0 mm in diameter; pores 0.035–0.04 mm in diameter, dispersed over the sides of the sponge; colour greyish-white | – | Usually smooth, 50–100 × 14–19 µm in size; tubercles are short and well rounded | 130 µm in length. | Fusiform, slender, 390 × 13 µm in size | Fusiform, sometimes with an ellipsoidal centrotylus, usually curved, 55 × 4 µm, in size | Florida, U.S.A. (278–494 m depth) |
| 3M. robusta Topsent, 1904 (unknow type) | Very hard sponges, simple in shape, with thick and short pedicel; top of the sponge can be curved or slightly depressed; water canals visible | – | Monocrepid, smooth, with short and thick tubercles, forming a very strong zygosis; 40 µm diameter | Cladome: scarcely ramified with very indented edges, 165–230 µm; rhabdome: conic, thick, 100–140 × 28–33 µm | Fusiform, slightly curved, 330–400 × 8–12 µm | Smooth, curve, thickened in the center, 20–60 × 4–7 µm | Azores (1,165 m depth) |
| M. robusta Topsent, 1904 (MNHN-IP-2008-216) | Ficiform to globular in shape, with a short and thick pedicel; surface smooth with openings and water canals visible to the naked eye; colour beige to light brown | 18–20 mm high, 14–22 mm in diameter | 248–362 (n = 2) × 17–22–31 µm (n = 22) | Cladome: variable in shape, indented on the edges, 154–228–309 µm (n = 20); rhabdome: 46–91–141 × 13–19–25 µm (n = 10) | 203–329 × 7.2–8.2 µm (n = 3) | 34.6–57.4–79.2 × 3.1–4.7–6.9 µm | Hyéres Seamount (705 m depth) |
| 3M. ramosa Topsent, 1904 (unknow type) | Encrusting with an extensive base where it stands two or three trunks that are slender, subcylindrical, with the top divided into short and obtuse branches | – | Zygosis interlocks with rounded tubercles | Cladome: large, foliated, thin, fully divided, 80–120 µm; rhabdome: conic, 75 × 13 µm | Fusiform, 200–300 × 5–6 µm | Smooth, slightly curved, thickened in the center, 50–65 × 4–5 µm | Azores (1,360 m depth) |
| M. schusterae sp. nov. (Holotype MNHN-IP-2018-87) | Foliate macandrewia with thick and contorted lamellas, usually attached to the substrate by a large pedicel; surface are smooth; colour light brown to white | 94 mm height, 142 mm wide at the top and 45 mm wide at the base; lamellas are 7–10 mm thick | Compact and irregular skeleton, with smooth, short and blunt clones, 301–386–463 × 10.2–19.9–39.2 µm (n = 27) | Cladome: incised especially in the edges, 177–304–420 µm; rhabdome: 67–119–178 × 13–21–26 µm (n = 13) | Smooth, round tips, 263–437–620 × 8.1–12.4–16.0 µm (n = 20) | Smooth, round tips, 43.8–67.9–95.2 × 2.5–4.3–7.7 µm | Gorringe Seamount (605–675 m depth) |
| M. schusterae sp. nov. (Paratype MNHN-IP-2018-88) | Foliate macandrewia with thick lamellas | 107 mm height, 22 mm wide at the base and 145 mm at the top; lamellas are 7–9 mm thick | 326–449–612 × 13.6–27.7–49.6 µm (n = 24) | Cladome: 187–325–457 µm; rhabdome: 94–138–207 × 12–22–31 µm (n = 21) | 302–466–563 × 5.3–10.0–13.3 µm (n = 21) | 53.6–74.0–109.8 × 3.8–6.0–8.3 µm | Gorringe Seamount (605–675 m depth) |
| M. minima sp. nov. (Holotype MNHN-IP-2008-222) | Round shape with a very small pedicel, smooth surface, pores are visible and scattered on the top; colour is white | 15–20 mm height, 17–20 mm wide, 16–17 mm in diameter; base 6 mm wide | Compact and irregular skeleton; clones are robust, usually smooth in the center, 268–318–348 (n = 10) × 7–29–50 µm | Cladome: incised ornamented by tubercles, 136–222–284 µm; rhabdome: conic and short, 58–99–136 × 14–19–25 µm (n = 13) | Smooth, 197–251–316 × 7.5–11.9–16.2 µm (n = 4) | Often curved, tips are blunt, 25.9–48.3–74.2 × 3.1–4.4–7.0 µm | Great Meteor Seamount (615 m depth) |
Notes:
‘–’ no information/not mentioned.
Discodermia cf. ramifera Topsent, 1892
Material. MNHN-IP-2008-210 (1993-02-02, Atlantis Seamount, epibenthic Warén dredge, DW258, 34°00′N, 30°12′W, 420 m, Seamount 2 campaign).
Comparative material examined. D. ramifera (MNHN-IP-2008-204, Great Meteor Seamount; MNHN-IP-2008-207, Great Meteor Seamount; MNHN-IP-2008-213, Great Meteor Seamount; MNHN-IP-2008-214, Great Meteor Seamount), Discodermia verrucosa Topsent, 1928 (MNHN-IP-2008-205, Atlantis Seamount; MNHN-IP-2008-206, Plato Seamount; HBOM 003:00869, Madeira; HBOM 003:00870, Madeira; HBOM 003:00868, Selvagens; HBOM 003:00640, Canary Islands; RMNH6237, Selvagens), D. kellyae sp. nov. (holotype MNHN-IP-2008-208, Plato Seamount), D. arbor sp. nov. (holotype MNHN-IP-2008-211, Great Meteor Seamount).
Description (MNHN-IP-2008-210). Small fragment, 20 × 10 mm in size, of elongated shape, with a smooth surface; subdermal water canals are visible, giving a striated appearance to the sponge; colour is beige in ethanol.
Skeleton. Ectosomal skeleton is formed by a layer of overlapped discotriaenes, and several microscleres spread through the surface; choanosome is formed by irregular tetraclone desmas, oxeas crossing the interior of the sponge and numerous microscleres spread through the interior of the sponge.
Spicules (MNHN-IP-2008-210).
Tetraclone desmas, irregular, with smooth clones and very tuberculated on the extremities, 400–455–534 × 30–51–82 µm in size; tubercles are smooth;
Discotriaenes, cladome varies from oval to indented in shape, usually flat, 195–328–560 µm in diameter; rhabdome is short, conical, with a blunt tip, 20–42–68 × 9.5–20.3–37.9 µm in size;
Oxeas, are present, but all of them were broken;
Acanthomicroxeas, very abundant, spinous, with sharp tips, 24.6–39.0–59.8 × 1.8–3.3–5.4 µm in size;
-
Acanthorhabds, small, abundant, spinous, with rounded extremities, 15.2–20.2–24.2 × 2.1–2.9–4.4 µm.
Distribution. This specimen was collected in the Atlantis Seamount at 420 m depth.
Remarks. Although the external morphology, type of spicules and desma ornamentation are in agreement with the description of D. ramifera, the spicules sizes of this specimen are significantly larger when compared to the ones found in the Great Meteor (Table 2). For this reason, we consider this species as D. cf. ramifera.
Discodermia verrucosa Topsent, 1928
Figures 2F, 12–13 and Table 2
Material examined. MNHN-IP-2008-205 (1993-02-02, Atlantis Seamount, beam trawl, CP257, 34°04′N, 30°15′W, 338 m), MNHN-IP-2008-206 (1993-02-01, Plato Seamount, epibenthic Warén dredge, DW246, 33°14′N, 29°36′W, 520 m). All from Seamount 2 campaign.
Comparative material examined. D. ramifera (MNHN-IP-2008-204, Great Meteor Seamount; MNHN-IP-2008-207, Great Meteor Seamount; MNHN-IP-2008-213, Great Meteor Seamount; MNHN-IP-2008-214, Great Meteor Seamount), D. kellyae sp. nov. (holotype MNHN-IP-2008-208, Plato Seamount), D. arbor sp. nov. (holotype MNHN-IP-2008-211, Great Meteor Seamount).
Diagnosis. Cup-shaped to spherical sponges with numerous warts/protuberances, and extremely tuberculated tetraclone desmas (emended after Topsent, 1928).
Description (MNHN-IP-2008-205). Spherical polymorphic sponge with several round protuberances, 15–20 mm high and 12–13 mm wide, with a rough surface (Fig. 2F); pores cannot be seen with naked eye; colour varies from whitish to light brown in ethanol.
Skeleton. Ectosome composed of a compact layer of discotriaenes, usually overlapping each other, numerous microscleres (acanthomicroxeas and acanthorhabds) spread through the surface, and oxeas perforating the sponges’ surface; occasionally, bundles of oxeas can be observed; choanosome with strongly tuberculated and compact tetraclone desmas (Fig. 12), forming an irregular net with dispersed microscleres in the interior of the sponge.

Figure 12: Skeleton of Discodermia verrucosa Topsent, 1928, specimen MNHN-IP-2008-205.
(A) Overview of tetraclone desmas, (B) and (C) irregular and compact net of tetraclone desmas, (D) detail of the strongly tuberculated zygosis.{kind=link}
Spicules (MNHN-IP-2008-205).
Tetraclone desmas, large, robust, mostly with tubercles spread through the entire clone, although some parts can be smooth, 106–170–278 × 19–34–46 µm in size (Figs. 12A–12C); zygoses very robust and extremely tuberculate (Fig. 12D);
Discotriaenes, irregular in shape, from round to oval, often indented (Figs. 13A–13D); cladome smooth, slightly concave, 102–153–222 µm in diameter; rhabdome is short with a conical shape, 15–25–47 × 5–8–13 µm (Fig. 13D);
Oxeas, long, smooth with rounded ends; length not presented here because they were all broken due to their large size.
Acanthomicroxeas, spinous, slightly curved with pointed ends, 22.8–35.2–53.5 × 1.3–2.2–3.9 µm (Fig. 13E).
Acanthorhabds, cylindrical, spinous, with blunt tips, 7.5–12.9–19.0 × 1.2–1.6–3.0 µm in size (Fig. 13F).
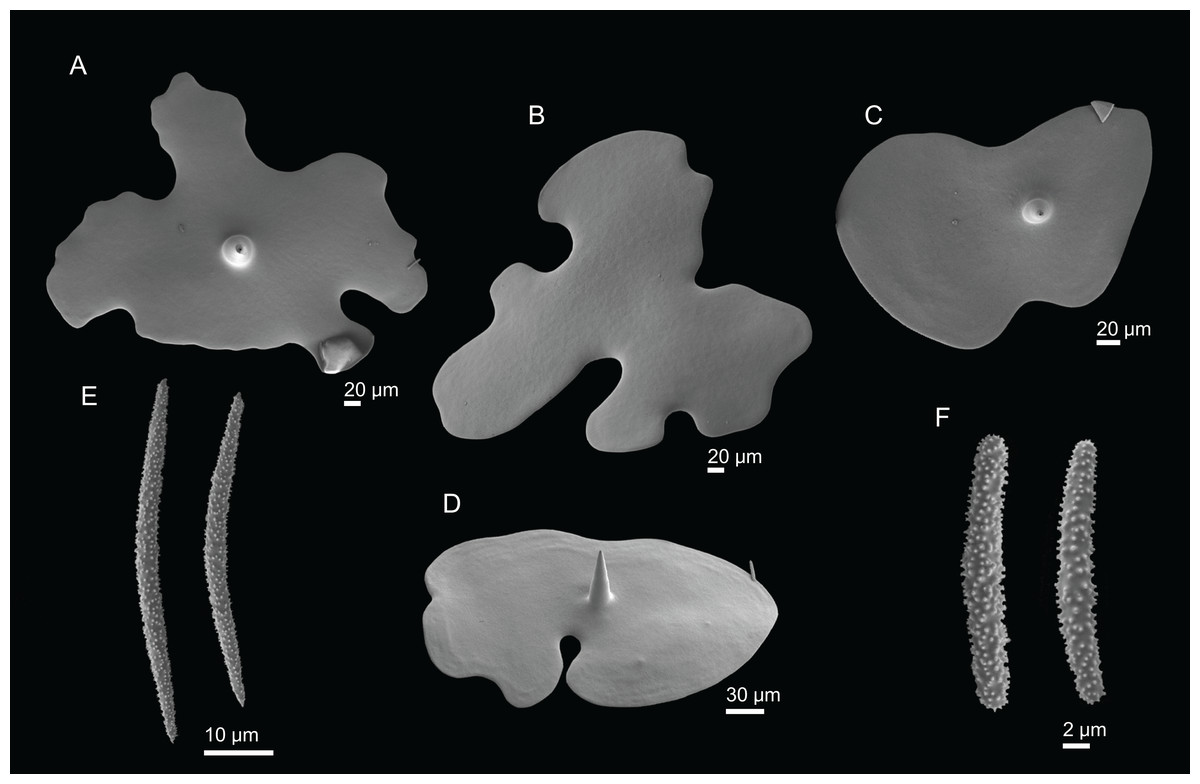
Figure 13: Spicules of Discodermia verrucosa Topsent, 1928, specimen MNHN-IP-2008-205.
(A)–(D) Upper and lower view of discotriaenes, (E) acanthomicroxeas, (F) acanthorhabds.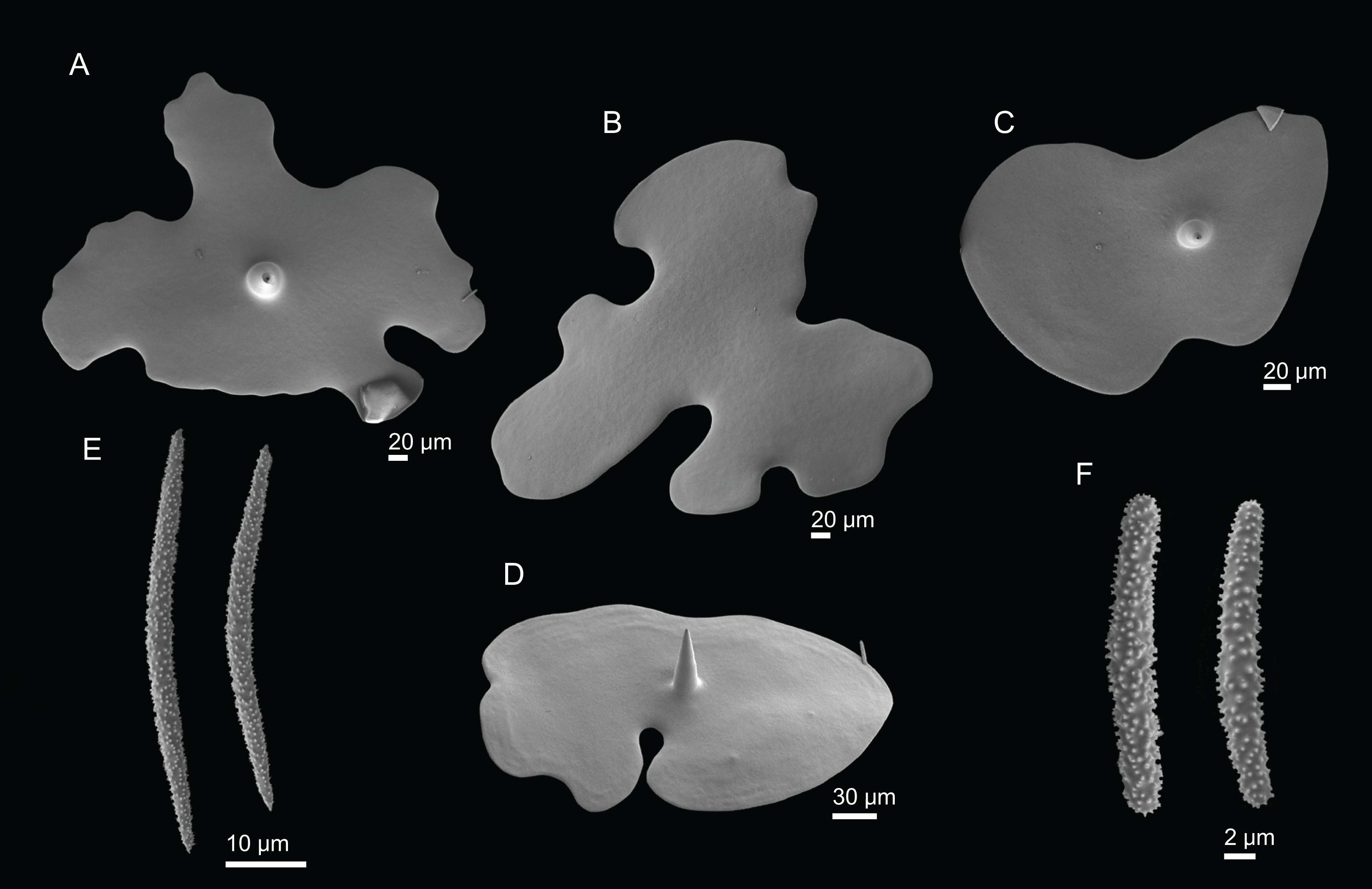{kind=link}
Distribution. Specimens of D. verrucosa were found in Atlantis and Plato Seamounts between 338 and 580 m depth.
Remarks. Discodermia verrucosa was first found in the Canary Islands and described by Topsent (1928). The species differs from the D. ramifera on the habitus and sculpture of desmas. D. verrucosa has a cup to spherical shape with several rounded protuberances/warts and strongly tuberculated tetraclones. On the other hand, D. ramifera has an elongated to branching shape and smooth tetraclone desmas only tuberculated in the extremities. The specimens analysed in this study overall match the description of D. verrucosa, apart from two differences: (1) the discotriaenes are much smaller and (2) the microscleres present a wider size range when compared to the original description (see Table 2).
Discodermia arbor sp. nov.
Figures 2G, 14–15 and Table 2
Urn:lsid:zoobank.org:act:7A732A92-8D8B-4D73-97B1-CD53E9494121
Holotype. MHNH-IP-2008-211 (1993-01-11, Great Meteor Seamount, beam trawl, DW159, 29°44′N, 28°20′W, 330 m, Seamount 2 campaign).
Comparative material examined. D. ramifera (MNHN-IP-2008-204, Great Meteor Seamount; MNHN-IP-2008-207, Great Meteor Seamount; MNHN-IP-2008-213, Great Meteor Seamount; MNHN-IP-2008-214, Great Meteor Seamount), D. verrucosa (MNHN-IP-2008-205, Atlantis Seamount; MNHN-IP-2008-206, Plato Seamount; HBOM 003:00869, Madeira; HBOM 003:00870, Madeira; HBOM 003:00868, Selvagens; HBOM 003:00640, Canary Islands; RMNH6237, Selvagens), D. kellyae sp. nov. (holotype MNHN-IP-2008-208, Plato Seamount).
Diagnosis. Discodermia of tree-like appearance; discotriaenes vary from square to circular shape and can also be indented.
Description (holotype MHNH-IP-2008-211). Discodermia of tree-like appearance (Fig. 2G), with a relatively long stem, 15 mm, where it extends on top into three branches; the stem is wider at the base, 12 mm, and thinner on top, 7.5 mm; branches are irregular and 13–28 mm long; surface is smooth but some rugosities/protuberances are visible; full sponge length is 58 mm; the sponge was attached to the substrate by the stem; colour is beige in ethanol.
Skeleton. Ectosome has a layer of overlapped discotriaenes of variables sizes (Figs. 14A and 14B) with numerous microscleres beneath them; choanosome is composed of an irregular net of tetraclone desmas (Figs. 14C and 14D) and spread microscleres; near the surface, tetraclones are more intricate, rugose, with very complex and strong zygoses near the water canals (Fig. 14C); in the interior part of the sponge, the tetraclones still form an intricate and irregular net, but there is more space between the desmas.
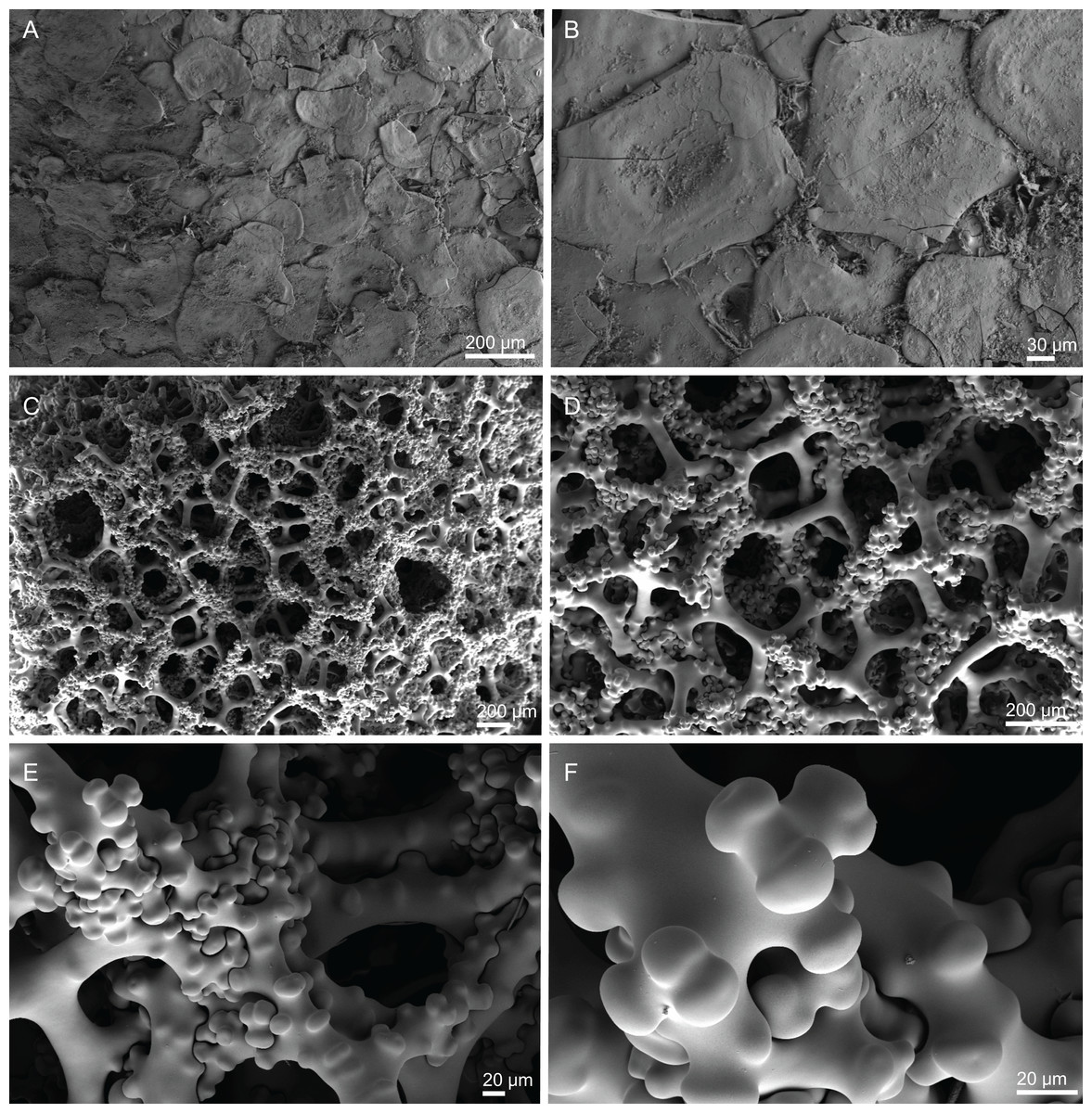
Figure 14: Surface and skeleton of Discodermia arbor sp. nov., holotype MNHN-IP-2008-211.
(A) Overview of the surface, (B) detail of the surface showing the overlapped discotriaenes, (C) overview of choanosomal tetraclone desmas, (D) detail of tetraclone desmas, (E) complex zygoses between several desmas, (F) detail of the desmas ornamentation, showing smooth tubercles.{kind=link}
Spicules (holotype MHNH-IP-2008-211).
Tetraclone desmas, thick, irregular, ornamentation varies according with the location of the desmas, i.e, near the surface the clones have usually tubercles spread through the entire ray (Figs. 14C and 14D) while in the interior they are smoother; tubercles on the zygome are smooth and sometimes subdivided (Fig. 14F); zygoses are very complex and robust (Figs. 14E and 14F), giving a hard consistency to this sponge; tetraclones are 181–392–567 × 15–36–56 µm in size;
Discotriaenes, very variable in shape, from “square” to “circular” shape, or with indented cladomes (Figs. 15A–15E); cladome is smooth with some protuberances, 148–256–396 µm in diameter; rhabdome is relatively short with blunt tips, 34–53–71 × 15–21–24 µm in size;
Acanthomicroxeas, slightly curved, covered by numerous spines with sharp tips, 24.1–35.1–50.1 × 1.4–2.3–3.5 µm in size (Figs. 15H and 15I);
Acanthorhabds, small, with several spines, usually with blunt tips, but they can also be unequal and have a sharp tip in one of the extremities, 6.7–16.1–25.9 × 1.1–2.2–4.3 µm in size (Figs. 15F and 15G);
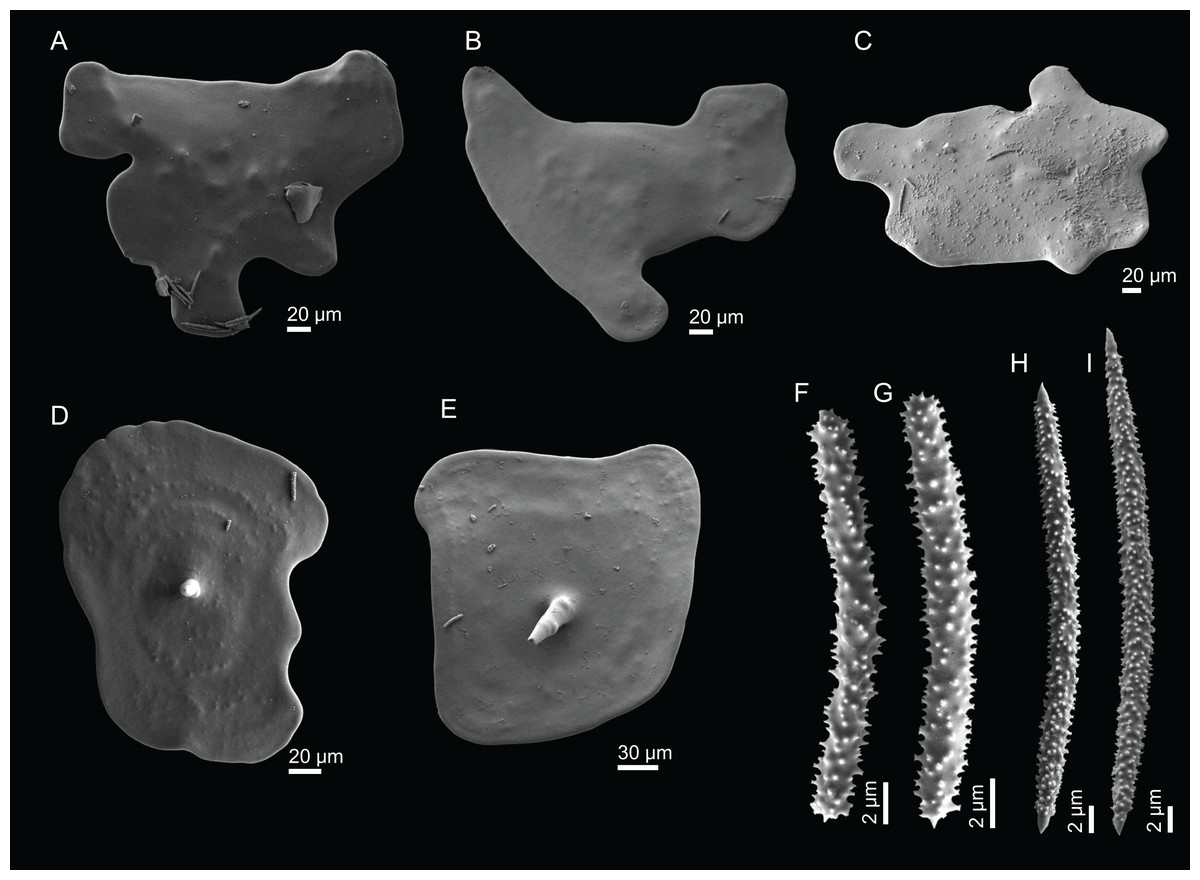
Figure 15: Spicules of Discodermia arbor sp. nov., holotype MNHN-IP-2008-211.
(A)–(C) Top view of discotriaenes, (D) and (E) bottom view of discotriaenes showing the rhabdome, (F) and (G) acanthorhabds, (H) and (I) acanthomicroxeas.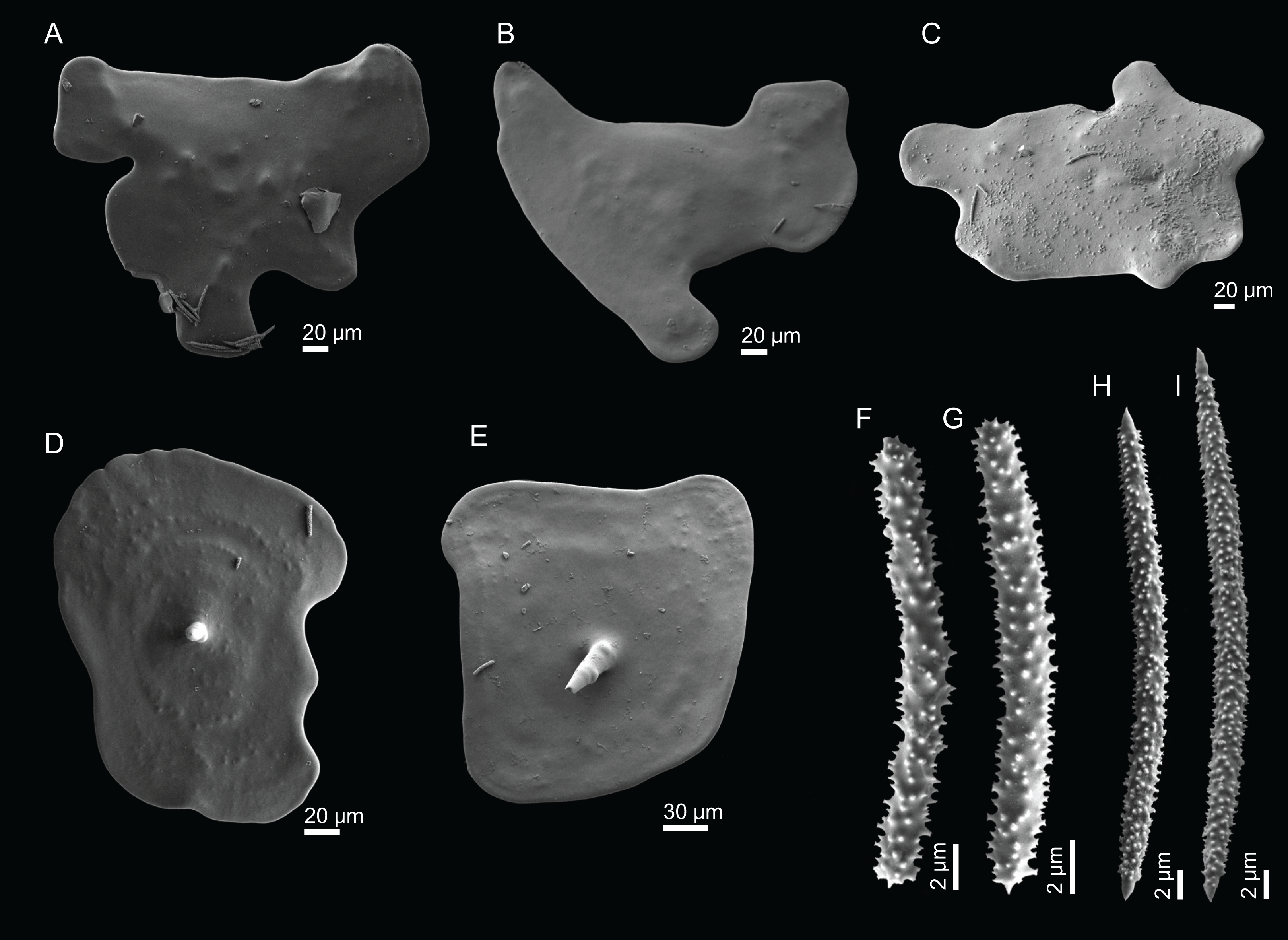{kind=link}
Etymology. From the latin arbor = tree; this Discodermia looks like a small tree.
Distribution. D. arbor sp. nov. is only know from the Great Meteor Seamount, where it was found at 330 m depth.
Remarks. Discodermia arbor sp. nov. is here described as a new species constituting the eighth Discodermia species reported to the North Atlantic and Mediterranean Sea. Its tree-like shape is very distinct from the other Discodermia spp. recorded for this area. Besides that, this species does not have oxeas, a spicule type that was reported in all Discodermia species in the North Atlantic except for D. polymorpha from the Mediterranean Sea. Although D. arbor sp. nov. shares the absence of oxeas with D. polymorpha, they have very different habitus, desmas ornamentation and size of microscleres (but see Remarks under D. kellyae sp. nov. for a more detailed comparison of all Discodermia species in the North Atlantic and Mediterranean Sea).
Discodermia kellyae sp. nov.
Figures 2H, 16–17 and Table 2
urn:lsid:zoobank.org:act:E7A06142-4AF7-404E-B369-B30240ADE5F4
Holotype. MNHN-IP-2008-208 (1993-02-03, Plato Seamount, beam trawl, DW247, 33°14’N, 29°35’W, 580 m, Seamount 2 campaign).
Comparative material examined. D. ramifera (MNHN-IP-2008-204, Great Meteor Seamount; MNHN-IP-2008-207, Great Meteor Seamount; MNHN-IP-2008-213, Great Meteor Seamount; MNHN-IP-2008-214, Great Meteor Seamount), D. verrucosa (MNHN-IP-2008-205, Atlantis Seamount; MNHN-IP-2008-206, Plato Seamount; HBOM 003:00869, Madeira; HBOM 003:00870, Madeira; HBOM 003:00868, Selvagens; HBOM 003:00640, Canary Islands; RMNH6237, Selvagens), D. arbor sp. nov. (holotype MNHN-IP-2008-211, Great Meteor Seamount).
Diagnosis. Massive, spherical, irregular, Discodermia of bulb appearance, with smooth tetraclone desmas.
Description (holotype MNHN-IP-2008-208). Massive sponge, irregular appearance, with large protuberances of round shape, 53 mm high and 31 mm wide; surface is irregular with a rugose appearance; the basal part of the sponge is not evident, since there is no obvious mark in the sponge that shows where it was attached to the substrate; colour is beige to light brown in alcohol (Fig. 2H).
Skeleton. Ectosome is composed of a layer of overlapped discotriaenes (Figs. 16A and 16B) of different sizes with several microscleres spread through the surface; openings are surrounded by these microscleres; choanosome is composed by an irregular net of tetraclone desmas (Figs. 16C and 16D), forming large areas between them, usually near the ectosome; the rays of the tetraclones articulate into a complex zygosis; several microscleres and some strongyles are spread loosely in the choanosome.
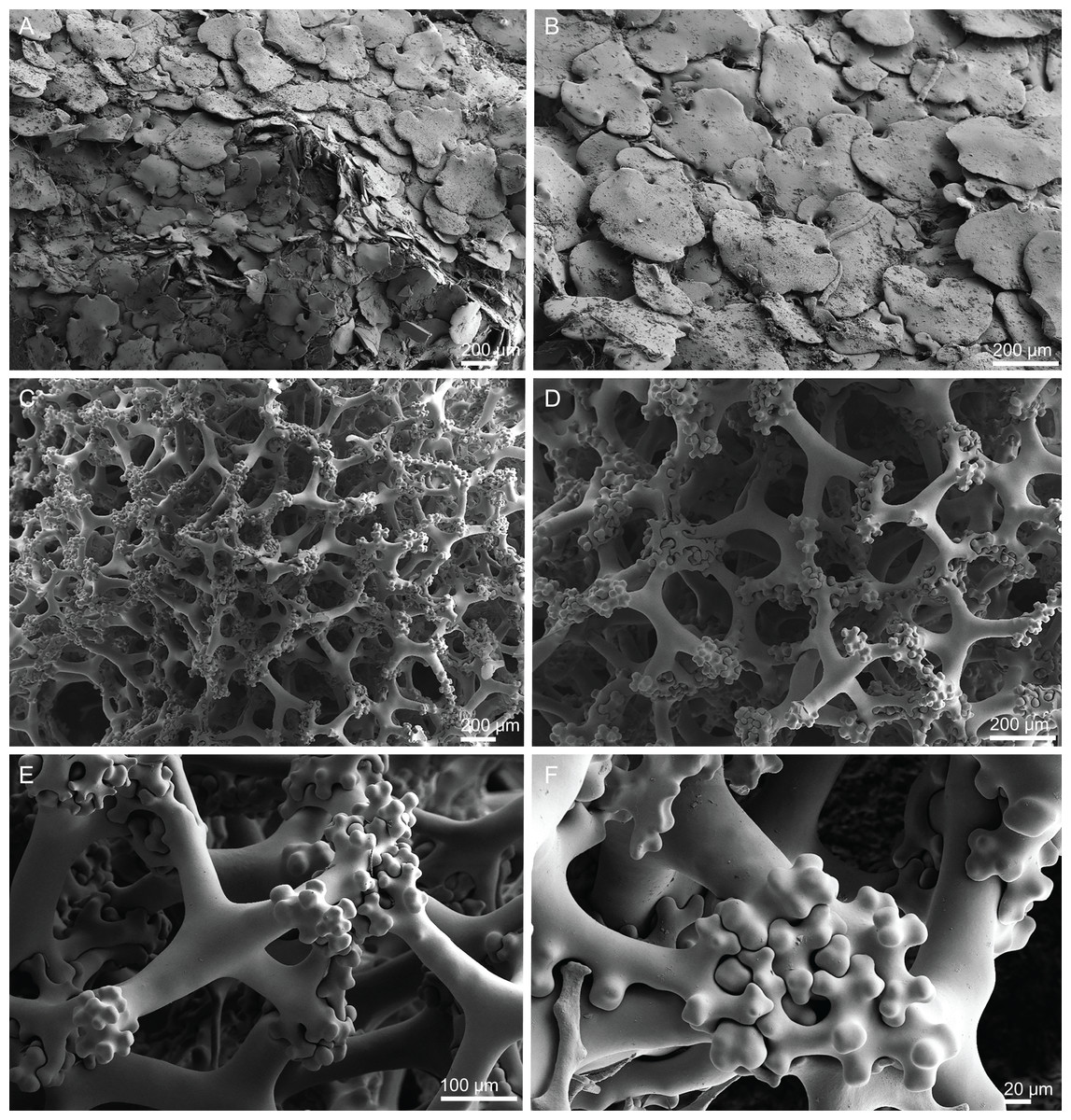
Figure 16: Surface and skeleton of Discodermia kellyae sp. nov., holotype MNHN-IP-2008-208.
(A) Overview of the surface, (B) overlapped discotriaenes on the surface, (C) overview of choanosomal tetraclone desmas, (D) tetraclone demas, (E) detail of a tetraclone desma showing their sculpture and ornamentation, (F) detail of the zygosis.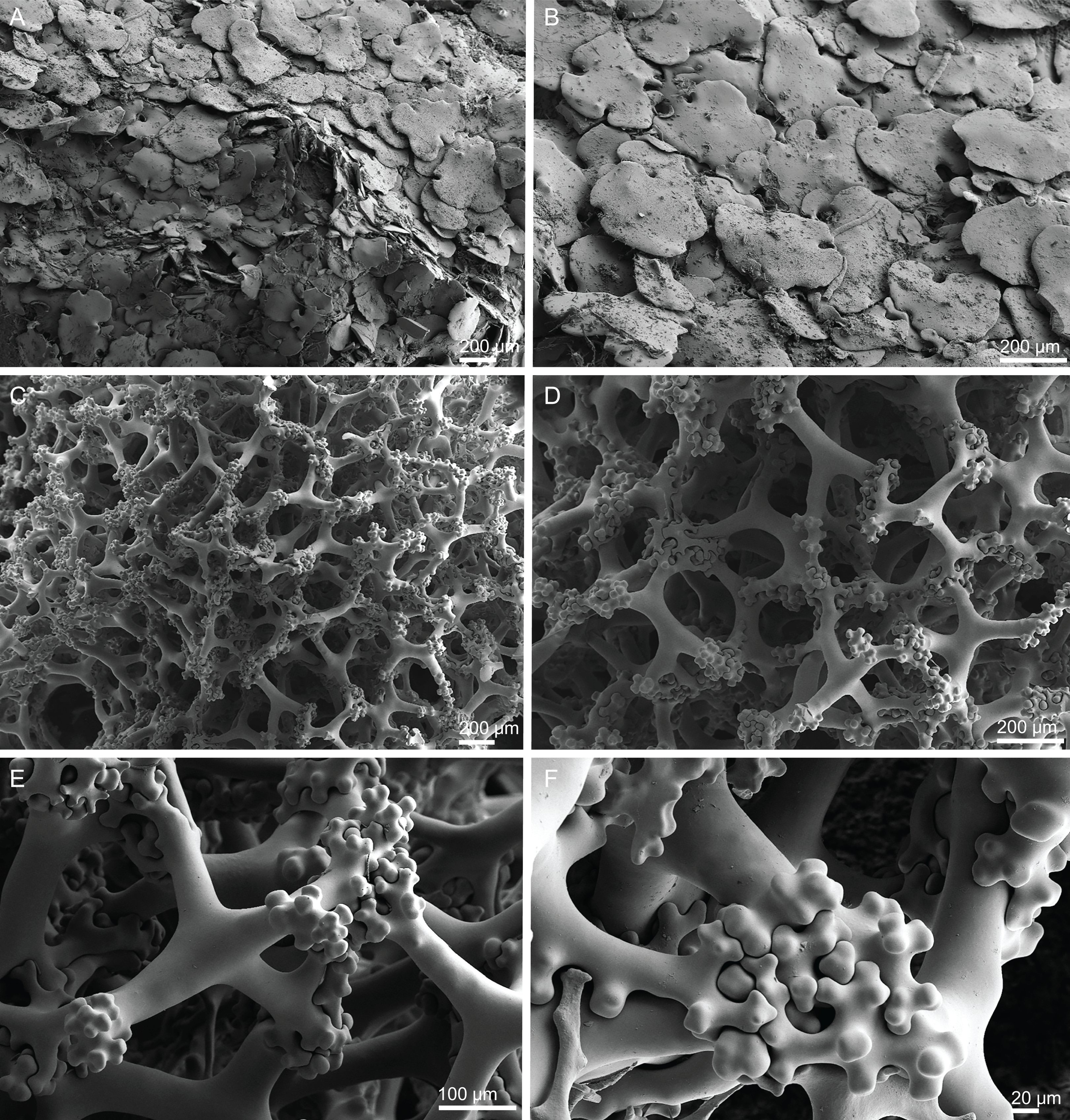{kind=link}
Spicules (holotype MNHN-IP-2008-208).
Tetraclone desmas, compact, irregular, with smooth and thick clones, 112–338–589 × 20–42–76 µm in size (Figs. 16C and 16D); the termination of the clones has several tubercles, resulting in very complex and large zygoses (Figs. 16D–16F); tubercles of the clones are smooth (Fig. 16F).
Discotriaenes, irregular, with diverse shapes and sizes; cladomes vary from oval to indented discs, and they are either flat or concave, 121–289–425 µm in diameter (Figs. 16A, 16B and 17A–17G); rhabdome is also very variable in size, 36–78–119 × 13–30–44 µm, with a blunt or sharp tip.
Strongyles, with one of the tips rounded and the other one sharp, sometimes resembling a crochet needle, 418–444 × 6.0–7.9 µm in size (Figs. 17H and 17I);
Acanthomicroxeas, very abundant, long, straight to curved, covered by numerous spines, with sharp tips, 16.7–43.2–66.5 × 1.5–2.5–3.7 µm in size (Figs. 17J and 17K);
Acanthorhabds, very abundant, with blunt tips, covered by numerous spines, very variable in size, 5.3–13.3–24.9 × 1.2–2.1–3.7 µm (Figs. 17L and 17M).
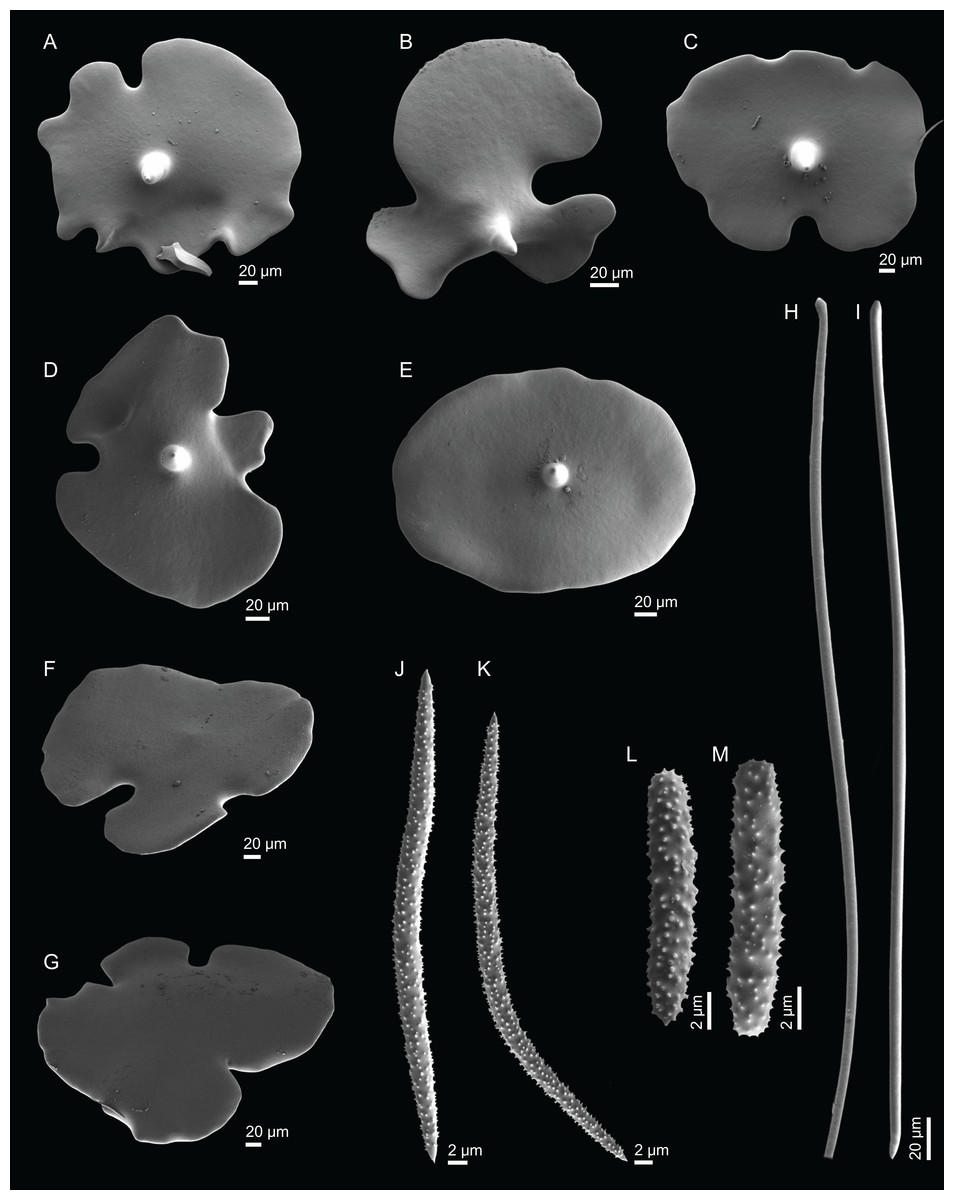
Figure 17: Spicules of Discodermia kellyae sp. nov., holotype MNHN-IP-2008-208.
(A)–(E) Bottom view of discotriaenes, (F and G) top view of discotriaenes, (H and I) strongyles, (J and K) acanthomicroxeas, (L and M) acanthorhabds.{kind=link}
Etymology. Named after Dr. Michelle Kelly from the National Institute of Water and Atmospheric Research (NIWA) in recognition of her work on taxonomy and systematics of Porifera, particularly on lithistid demosponges of New Zealand.
Distribution. D. kellyae sp. nov. is only known from its type locality, the Plato Seamount at 580 m depth.
Remarks. The identification of species belonging to the genus Discodermia is particularly challenging due to the few and very variable morphological characters used for the distinction of species (Pisera & Vacelet, 2011). Moreover, for some species we are limited to the original descriptions where detailed information of skeletal composition and spicule sizes, or images are lacking.
In the North Atlantic and Mediterranean Sea, a total of nine species have been described, including the two described species in this study (Table 2). Despite the high plasticity of morphological characters, the main differences between species are (1) habitus, (2) the sculpture and size of the desmas, (3) size and shape of the discotriaenes, and (4) size and shape of the microscleres. We propose D. kellyae sp. nov. as a new species based on (1) the habitus of this sponge: the polymorphic sponge of bulb appearance contrasts with the massively encrusting shape of D. adhaerens, the spherical to irregular masses in D. polymorpha, the cup-shaped with numerous warts/protuberances in D. verrucosa, the elongated with several finger-like extensions in D. ramifera, the tree-like shape of D. arbor, the cluster of knobby fingers in D. dissoluta and the irregular mushroom shape of D. polydiscus; (2) tetraclones of D. kellyae sp. nov. have similar ornamentation to the ones found in D. ramifera (tetraclones with smooth clones that are tuberculated in the zygomes), however, they are more compact and thicker (24–32–48 µm vs 20–42–76 µm) resembling the ones present in D. verrucosa; the other species have slender and smooth desmas without strong/complex zygoses; (3) the intraspecific size range of discotriaenes is usually wide, and similar between the different species, but in D. kellyae sp. nov. the size range of the cladomes is very large, 121–425 µm, and this can only be observed in D. verrucosa (200–560 µm) and D. arbor sp. nov. (148–396 µm); besides that, the shape of the rhabdome is also variable in D. kellyae sp. nov., where the tips of the rhabdomes can be blunt or sharp; (4) the size of the acanthomicroxeas in D. kellyae sp. nov. is larger (16.7–43.2–66.5 µm) compared to the other species, except when compared to D. dissoluta (41.6–68.0 µm; however, these values were taken from Pisera & Pomponi, 2015 where the authors presented a detailed description of the species, since in the original description, the species was poorly described and no measurements were given); (5) D. kellyae sp. nov., along with D. arbor sp. nov., are the only species with a wide acanthorhabds size range (5.3–13.3–24.9 µm and 6.7–16.1–25.9 µm, respectively) while the other species have a considerably narrower range (Table 2).
The species D. inscripta (Schmidt, 1879) was not included here for comparison because the type material was deciduous and the species is therefore considered incertae sedis (Pisera & Lévi, 2002d).
Family Macandrewiidae Schrammen, 1924
Genus Macandrewia Gray, 1859
Diagnosis. Macandrewiidae with phyllotriaenes/discotriaenes as ectosomal megascleres; choanosmal megascleres are oxeas and desmas with a triaenose crepsis; microscleres are microxeas (emended after Pisera & Lévi, 2002e).
Definition. Polymorphic Macandrewiidae; ectosomal spicules are dentate phyllotriaenes and/or discotriaenes; desmas are smooth with a triaenose (rarely monaxial) crepsis, and a terminal zygosis; oxeas are smooth; microscleres are microxeas (emended after Pisera & Lévi, 2002b).
Type species. Macandrewia azorica Gray, 1859 (type by monotypy).
Macandrewia cf. azorica Gray, 1859
Figures 2I, 18–19 and Table 3
Material. MNHN-IP-2008-217 (1993-02-03, Atlantis Seamount, beam trawl, DW263, 34°26′N, 30°32′W, 610 m), MNHN-IP-2008-220 (1993-02-03, Atlantis Seamount, epibenthic Warén dredge, DW258, 34°00′N, 30°12′W, 1,000 m), MNHN-IP-2008-225 (1993-02-06, Tyro Seamount, epibenthic Warén dredge, DW277, 34°00′N, 28°21′W, 1,000 m), MNHN-IP-2008-226 (1993-01, no data about station, 500 m), MNHN-IP-2008-229 (1993-01-06, Gran Canaria, epibenthic Warén dredge, DW129, 28°08′N, 15°52′W, 480 m), MNHN-IP-2008-249a (1993-01-06, Hyères Seamount, epibenthic Warén dredge, DW202, 31°16′N, 28°43′W, 640 m). All from Seamount 2 campaign.
Comparative material examined. M. azorica (holotype BMNH 1851.7.28.16, S. Miguel island, Azores; HBOM 003:00784, Selvagens), M. robusta (MNHN-IP-2008-216, Hyères Seamount; MNHN-IP-2008-224, Hyères Seamount), M. schusterae sp. nov. (holotype MNHN-IP-2018-87 and paratype MNHN-IP-2018-90, Gorringe Seamount), M. minima sp. nov. (MNHN-IP-2008-222, Great Meteor Seamount).
Description (MNHN-IP-2008-220). Polymorphic sponges attached to the substrate by a thick pedicel/stem, 67 × 50 mm in size; lamellas are thin, rounded and undulate, 3–5 mm thick (Fig. 2I); inner surface (top) has openings visible to the naked eye, around 224 µm in size (Fig. 18A); outer surface is smooth with several little openings spread randomly through the entire sponge, 40–83 µm in size (Fig. 18B); colour is beige to light brown in ethanol.
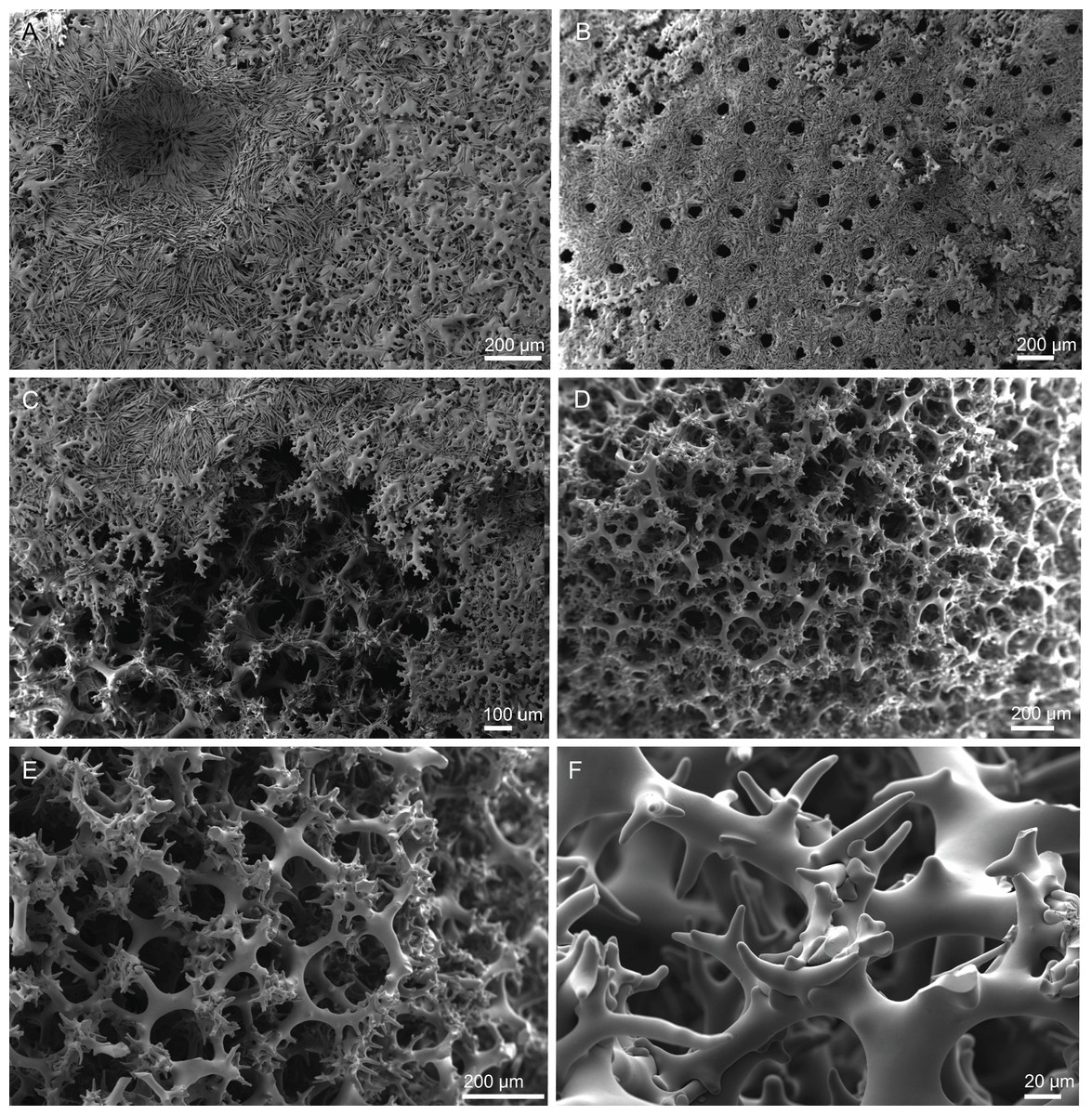
Figure 18: Surface and skeleton of Macandrewia cf. azorica Gray, 1859, specimen MNHN-IP-2008-220.
(A) Upper/inner surface with large openings, (B) lower/outer surface with several small openings, (C) division between ectosome and choanosome: top of the image showing the ectosome formed by phyllotriaenes and microxeas, and the bottom showing the desmas, (D) choanosomal desmas, (E) choanosomal desmas resembling tetraclones, (F) detail of the sculpture of desmas and zygoses.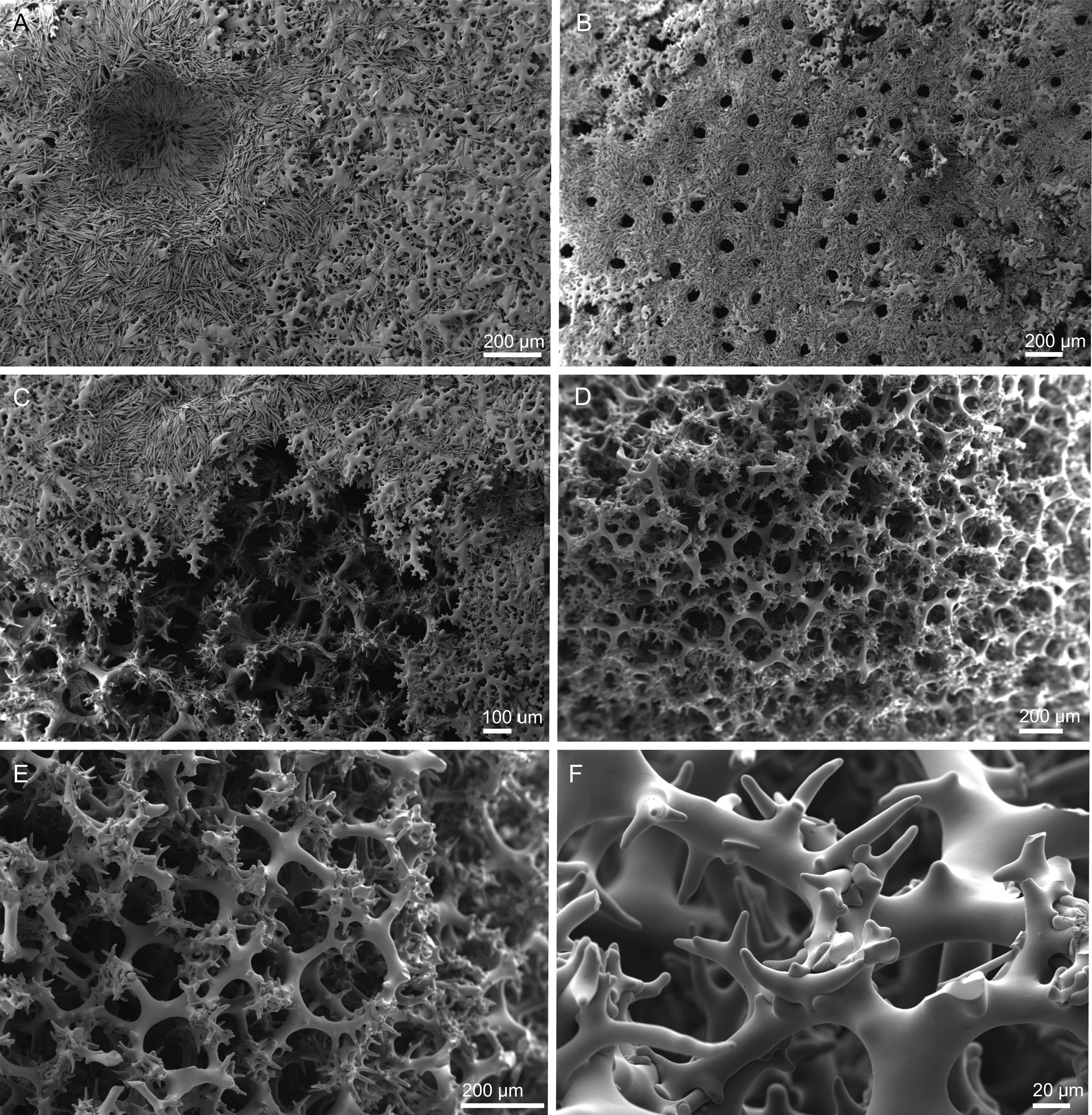{kind=link}
Skeleton. Ectosome formed by a layer of overlapped phyllotriaenes covered by numerous microxeas (Figs. 18A–18C); small openings are surrounded by microxeas (Fig. 18A) whereas larger openings are delimited by both phyllotriaenes and microxeas (Fig. 18B); choanosomal skeleton formed by a regular and solid network of desmas with a triaenose crepsis, resembling tetraclone desmas (Figs. 18D and 18E), some oxeas and microxeas are spread in the interior of the sponge.
Spicules (MNHN-IP-2008-220).
Desmas, with a triaenose crepsis, compact, forming a regular mesh, resembling tetraclones; rays are smooth with branches, especially on the termination of the clone, measuring 212–281–343 × 16–34–51 µm in size; branches have blunt ends and their size is very variable, 34–54–74 × 5.9–8.3–11.5 µm in size (Figs. 18D–18F);
Phyllotriaenes, with particularly incised cladome with 194–267–333 µm in diameter, and a short conical-shaped rhabdome, 62–99–129 × 11.6–14.4–17.8 µm in size; cladomes are very variable, from a simple (Fig. 19A) to a very complex and incised shape (Figs. 19B and 19C);
Oxeas, smooth, slightly curved with pointed ends, 215–246–301 × 6.8–7.8–9.1 µm in size (Fig. 19D);
Microxeas, smooth, fusiform with blunt tips, slightly curved, very abundant, 33.3–55.0–83.6 × 2.5–3.9–5.1 µm in size (Fig. 19E).
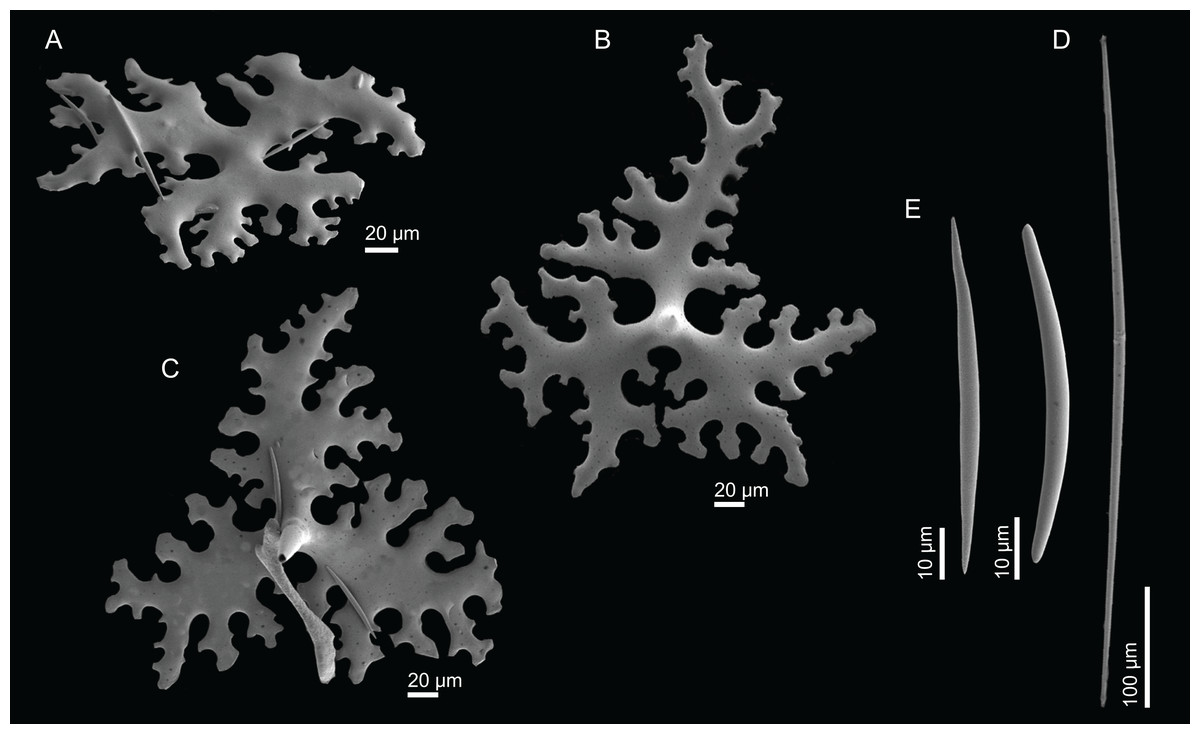
Figure 19: Spicules of Macandrewia cf. azorica Gray, 1859, specimen MNHN-IP-2008-220.
(A)–(C) Phyllotriaenes with a very incised cladome, (D) oxeas, (E) microxeas.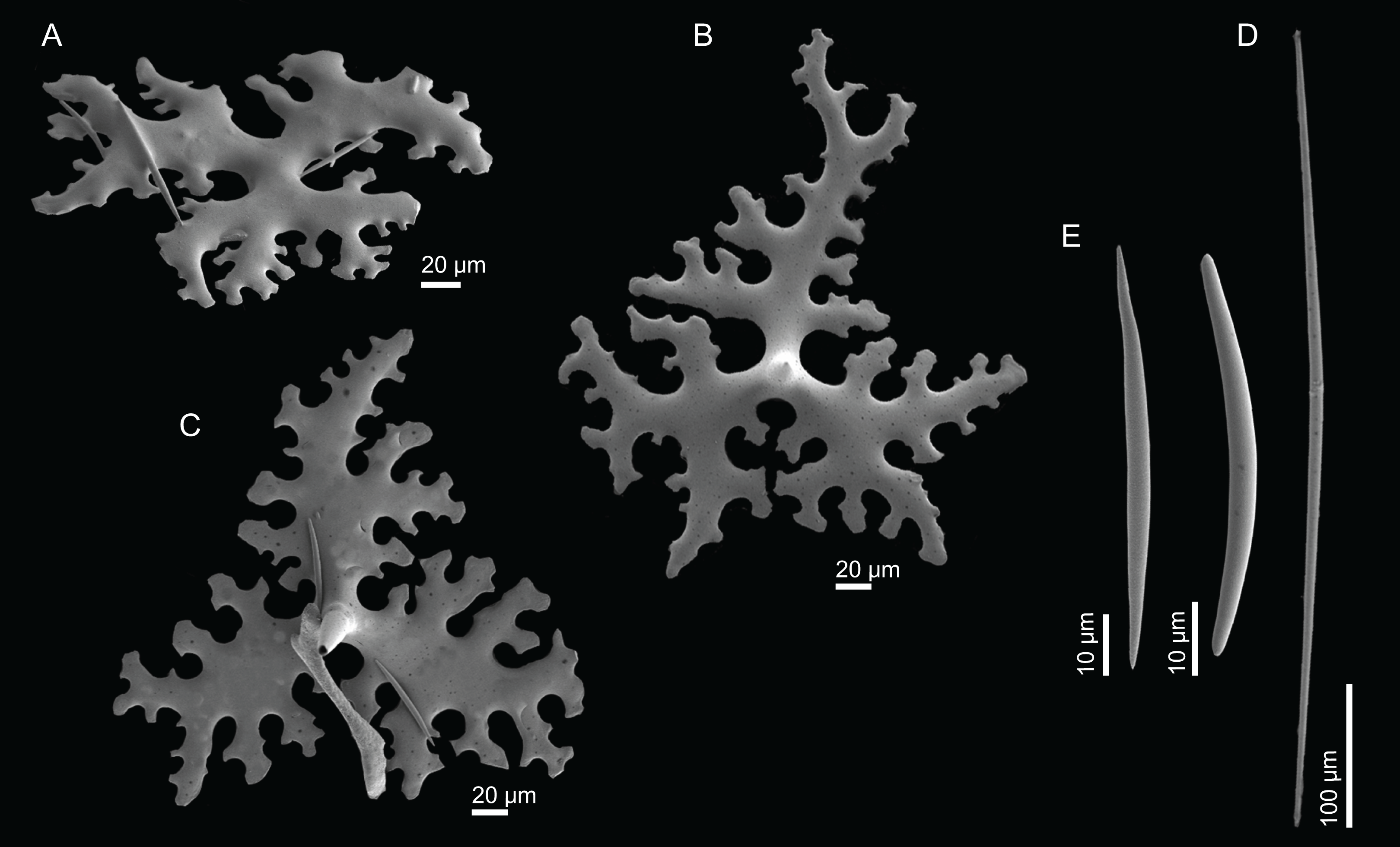{kind=link}
Distribution. The specimens were found on the Atlantis Seamount between 420 and 610 m depth, and one specimen was collected in Gran Canaria at 480 m depth.
Remarks. Pisera & Lévi (2002d) re-described and illustrated the holotype of M. azorica, a specimen collected in the Azores archipelago. Since we also had access to the holotype of M. azorica we have made new measurements of the spicules, in order to fill the gaps of some spicule’s measurements missing in the redescription. The comparison of the holotype of M. azorica with the specimens collected during the campaigns Seamount 1 and 2, lead us to consider these specimens as M. cf. azorica. Although very similar in the habitus they differ from the holotype in two features: (1) desmas are considerably more robust and thicker, resembling tetraclones (MNHN-IP-2008-220: 16–34–51 µm width vs holotype BMNH 1851.7.28.16: 8.5–19.0–30.8 µm width), forming compact network, while in the redescription of the holotype, the desmas have a “variable morphology” resembling tetraclones or rhizoclones, with strongly branched clones at the tip, forming a complex and loose articulation (Pisera & Lévi, 2002e); (2) the size of the cladome of the phyllotriaenes (MNHN-IP-2008-220: 194–267–333 µm in diameter vs holotype BMNH 1851.7.28.16: 297–363–456 µm in diameter) and oxeas (MNHN-IP-2008-220: 215–246–301 µm length vs holotype BMNH 1851.7.28.16: 532–652–780 µm length) is considerably smaller (Table 3).
Nineteen large specimens were found in the same station in the Hyères seamount (station DW202), suggesting that the species may be forming a sponge ground in this area of the seamount.
Macandrewia robusta Topsent, 1904
Figures 2J, 20–21 and Table 3
Material. MNHN-IP-2008-216, two specimens (1993-01-16, Hyères Seamount, epibenthic Warén dredge, DW184, 31°24′N, 28°52′W, 705 m), MNHN-IP-2008-224 two specimens (1993-01-16, Hyères Seamount, epibenthic Warén dredge, DW184, 31°24′N, 28°52′W, 705 m). All from Seamount 2 campaign.
Comparative material examined. M. azorica (holotype BMNH 1851.7.28.16, S. Miguel island, Azores; HBOM 003:00784, Selvagens), M. cf. azorica (MNHN-IP-2008-217, Atlantis Seamount; MNHN-IP-2008-220, Atlantis Seamount; MNHN-IP-2008-225, Tyro Seamount; MNHN-IP-2008-226, no data; MNHN-IP-2008-229, Gran Canaria; MNHN-IP-2008-249a, Hyères Seamount), M. schusterae sp. nov. (holotype MNHN-IP-2018-87 and paratype MNHN-IP-2018-90, Gorringe Seamount), M. minima sp. nov. (MNHN-IP-2008-222, Great Meteor Seamount).
Diagnosis. Small ficiform to globular shape Macandrewia with a flattened top and a short and thick pedicel.
Description (MNHN-IP-2008-216). Small sponges with a ficiform to globular shape, 18–20 × 14–22 mm in size, attached to the substrate by a short and thick pedicel (8 mm in height and 16 mm width) (Fig. 2J); top of the sponge is flattened, smooth, where openings can be observed in small clusters leading to water canals giving a striated appearance to the sponge; openings and the subdermal water canals visible to the naked eye; lateral walls of the sponge are smooth with small openings spread evenly through this surface; in some individuals, the top or upper surface has a slight depression; colour varies from beige to light brown in alcohol.
Skeleton. Ectosome is composed of a layer of overlapped phyllotriaenes and numerous microxeas; these microxeas surround the openings radially; choanosomal skeleton formed by desmas, oxeas and dispersed microxeas; desmas form an irregular and very dense mesh (Fig. 20).
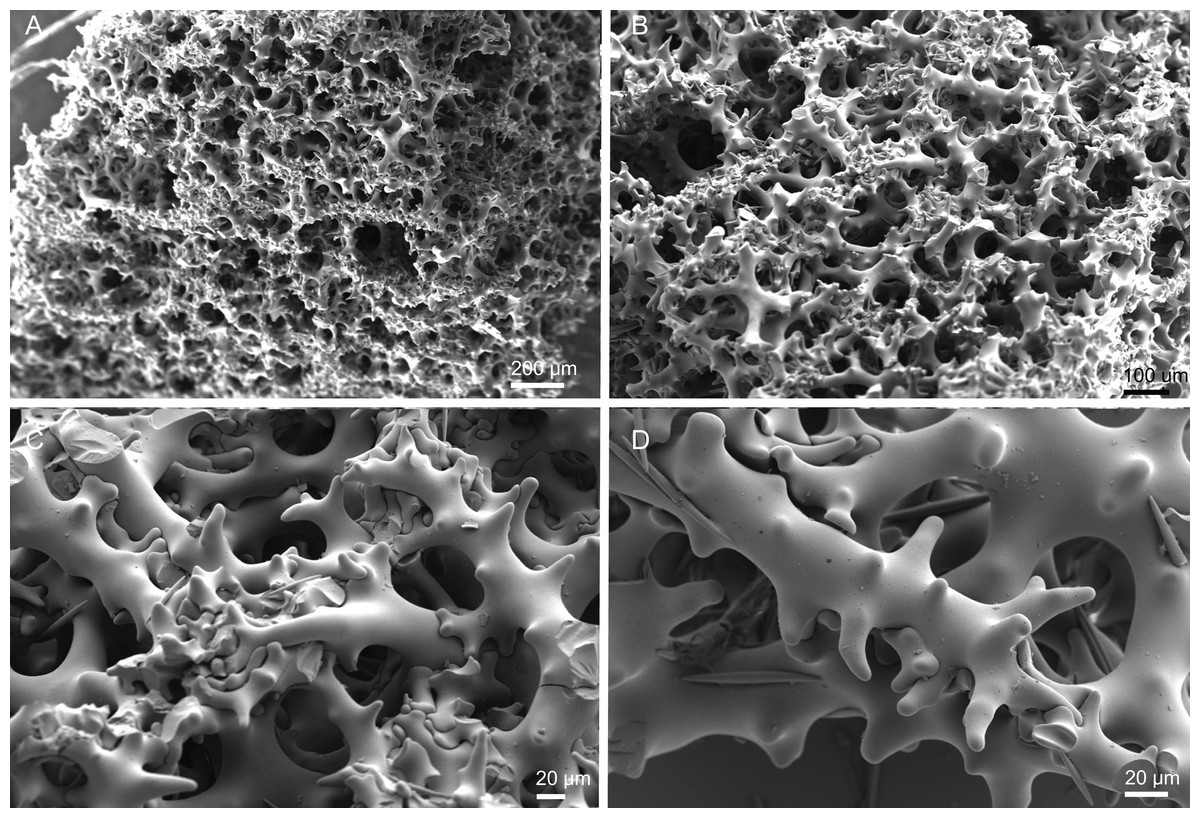
Figure 20: Skeleton of Macandrewia robusta Topsent, 1904, specimen MNHN-IP-2008-216.
(A) Overview of choanosomal desmas, (B) desmas, (C) zygoses, (D) sculpture of the desmas.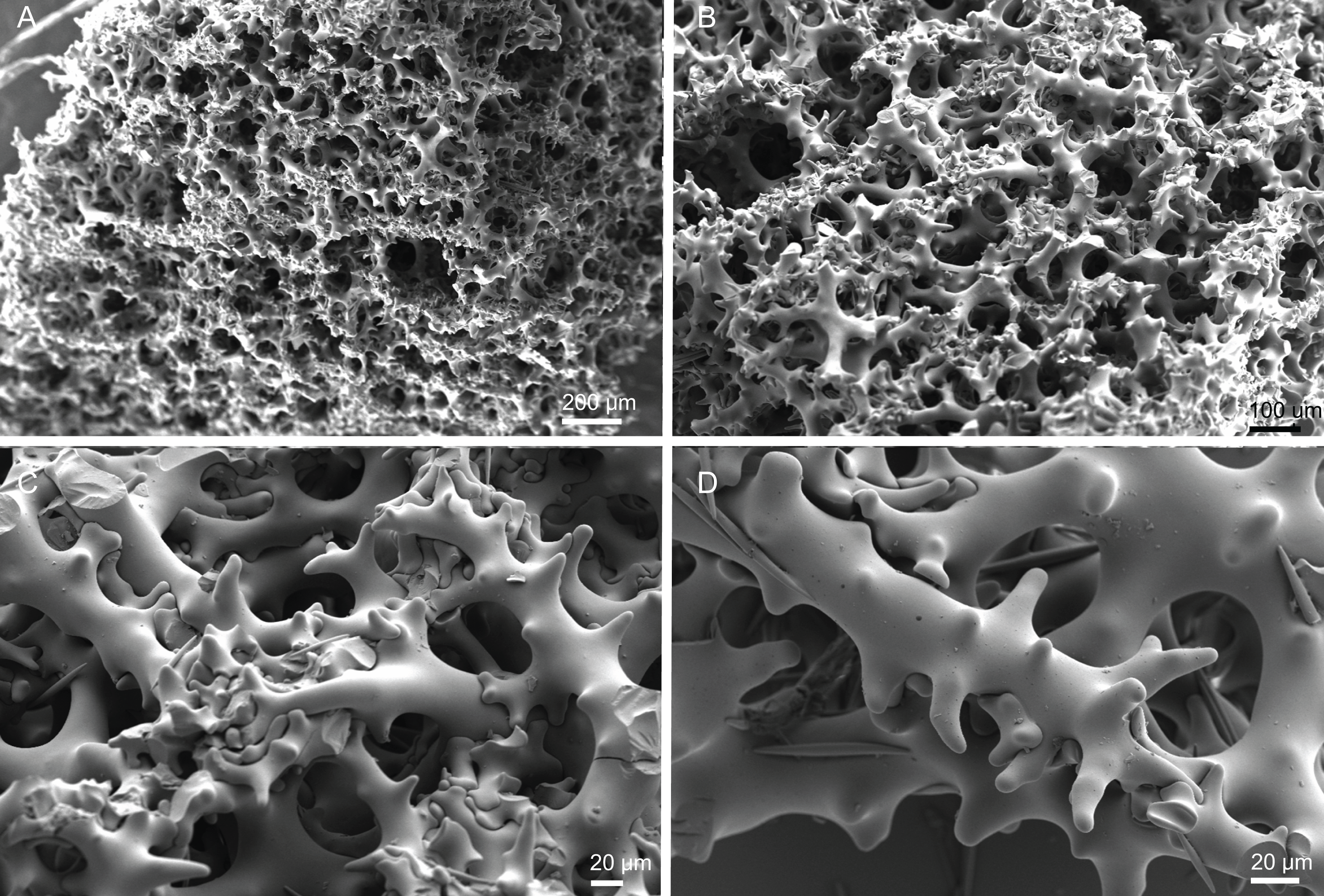{kind=link}
Spicules (MNHN-IP-2008-216).
Desmas, with a triaenose crepsis, compact, robust, with smooth clones that are very branched, 248–362 µm in size and 17–22–31 µm thick (Figs. 20A and 20B); clones have several short (18–41–75 µm), thick (7–10–12 µm) and blunt branches (Figs. 20D and 20E); the zygosis, that can be formed by numerous clones, is strong and complex (Fig. 20D).
Phyllotriaenes, very variable in shape, with a cladome particularly indented on the edges, 15–228–309 µm in diameter, with a conical rhabdome 46–91–141 × 13–19–25 µm in size (Figs. 21A–21D);
Oxeas, smooth with rounded tips, 203–329 × 7.2–8.2 µm thick (Fig. 21F).
Microxeas, smooth, with rounded extremities, slightly curved, 34.6–57.4–79.2 × 3.1–4.7–6.9 µm wide (Fig. 21E).
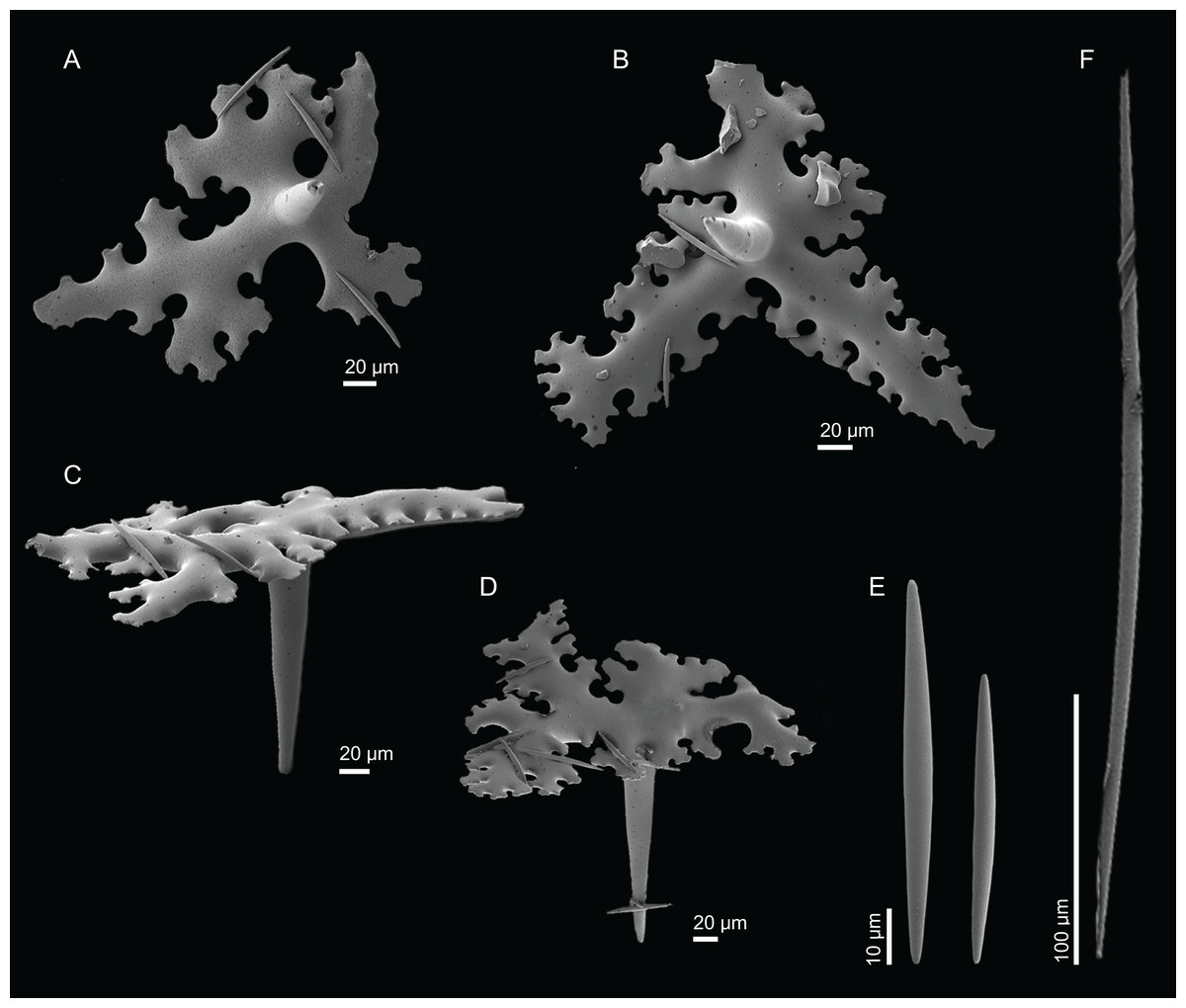
Figure 21: Spicules of Macandrewia robusta Topsent, 1904, specimen MNHN-IP-2008-216.
(A)–(D) Phyllotriaenes, (E) microxeas, (F) oxea.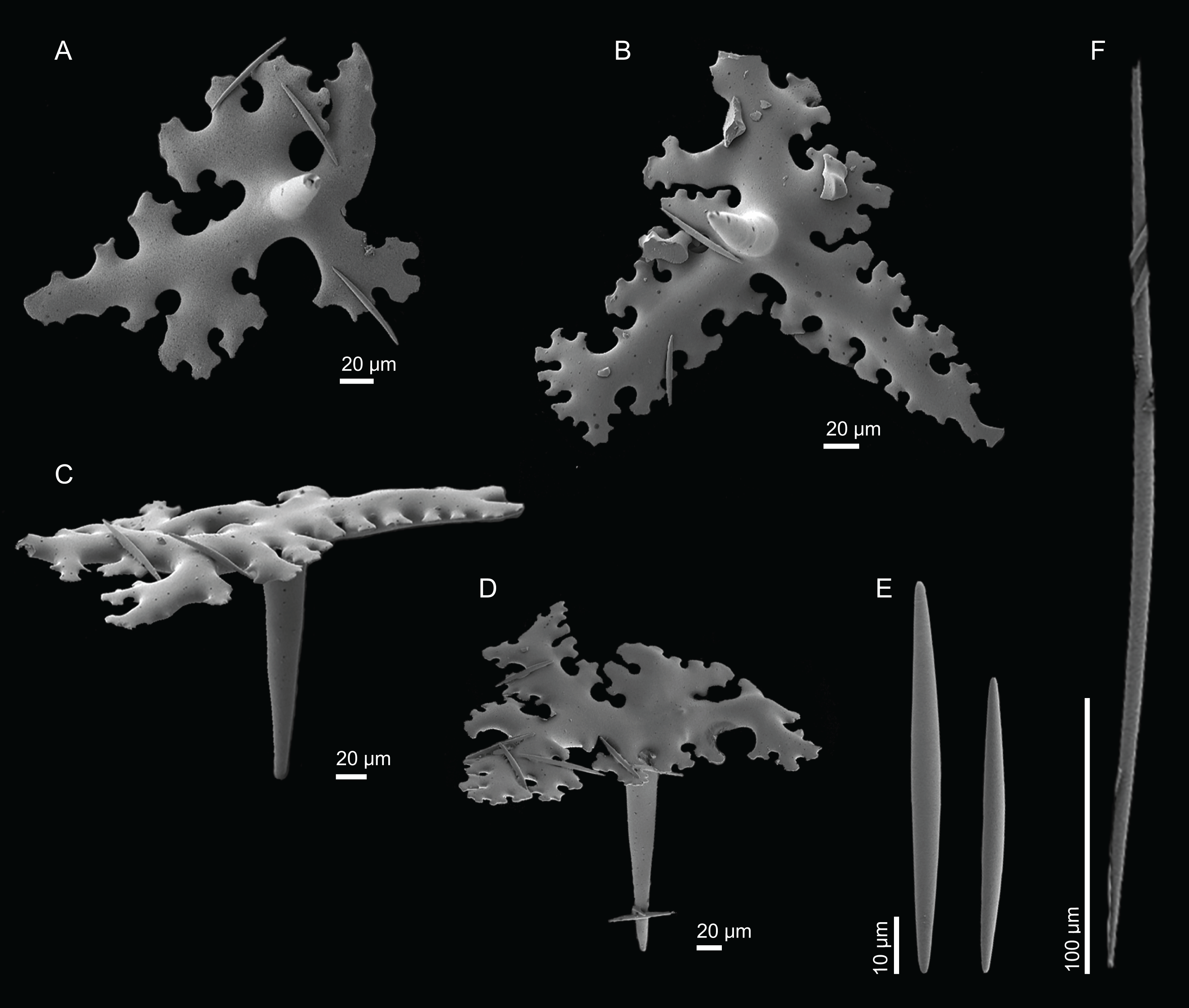{kind=link}
Distribution. These specimens were found on Hyères seamount at 705 m depth.
Remarks. In the specimens here examined, phyllotriaenes (165–230 µm vs 154–309; Table 3) and oxeas (330–400 vs 203–309; Table 3) are smaller when compared to previous records for the species (Topsent, 1904). However, M. robusta has a very distinct habitus in relation to the other Macandrewia described for the North Atlantic Ocean (Table 3). Its ficiform to globular shape, with a short and thick pedicel, contrasts with the cyathiform to flabellate shape with undulating rounded margins in M. azorica, the encrusting with standing trunks of M. ramosa, the foliate with thick lamellas in M. schusterae sp. nov., or the globular shape with a small pedicel as in M. minima sp. nov. (descriptions of the latter two below). Differences in spicule sizes were observed in another species analysed in this work as well as in other studies (see ‘Spicules dimensions’ section in the Discussion for further information regarding this topic).
Two specimens from the Seamount 2 collection could not be confidently identified down to species level (MNHN-IP-2008-228 and MNHN-IP-2018-94). They are very small fragments, seemingly encrusting, and most likely it is a Macandrewia at an early stage of development. The spicules were measured and they fall within the size range found in M. robusta.
Macandrewia schusterae sp. nov.
Figures 2K, 22–23 and Table 3
urn:lsid:zoobank.org:act:2BA2C1EF-8FAB-4C91-89CB-DCB59DDA61EB
Holotype. MNHN-IP-2018-87 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°28′N, 11°29’W, 605–675 m, Seamount 1 campaign).
Paratype. MNHN-IP-2018-88 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°28’N, 11°29′W, 605–675 m, Seamount 1 campaign).
Other material. MNHN-IP-2018-90, six specimens (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°28′N, 11°29′W, 605–675 m, Seamount 1 campaign), MNHN-IP-2018-89 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°28′N, 11°29′W, 605–675 m, Seamount 1 campaign), MNHN-IP-2018-91 (1988-09-26, Gorringe Seamount, beam trawl, CP28, 36°28′N, 11°29′W, 605–675 m, Seamount 1 campaign), MNHN-IP-2008-219 (1993-02-06, Tyro Seamount, epibenthic Warén dredge, DW279, 33°56′N, 28°24′W, 805 m, Seamount 2 campaign), MNHN-IP-2008-230 (1993-02-01, Plato Seamount, epibenthic Warén dredge, DW246, 33°14′N, 29°36′W, 520 m, Seamount 2 campaign).
Comparative material examined. M. azorica (holotype BMNH 1851.7.28.16, S. Miguel island, Azores; HBOM 003:00784, Selvagens), M. cf. azorica (MNHN-IP-2008-217, Atlantis Seamount; MNHN-IP-2008-220, Atlantis Seamount; MNHN-IP-2008-225, Tyro Seamount; MNHN-IP-2008-226, no data; MNHN-IP-2008-229, Gran Canaria; MNHN-IP-2008-249a, Hyères Seamount), M. robusta (MNHN-IP-2008-216, Hyéres Seamount; MNHN-IP-2008-224, Hyères Seamount), M. minima sp. nov. MNHN-IP-2008-222 (Great Meteor Seamount).
Diagnosis. Foliate to vase shaped Macandrewia with thick, irregular and undulated lamellas, with a small pedicel.
Description (holotype MNHN-IP-2018-87). Massive, foliate to vase shape with undulate lamellas, 94 mm high and 142 mm wide at the top and 45 mm wide at the base, usually attached to the substrate by a large pedicel; lamellas are generally irregular and contorted; walls are thick, 7–10 mm (Fig. 2K); interior surface with openings slightly elevated and evenly distributed, 278–378 µm in diameter (Fig. 22A); subdermal water canals are visible on the inner surface; external surface is smooth and covered by small openings, 29–98 µm in size (Fig. 22B); colour light brown to white in alcohol.
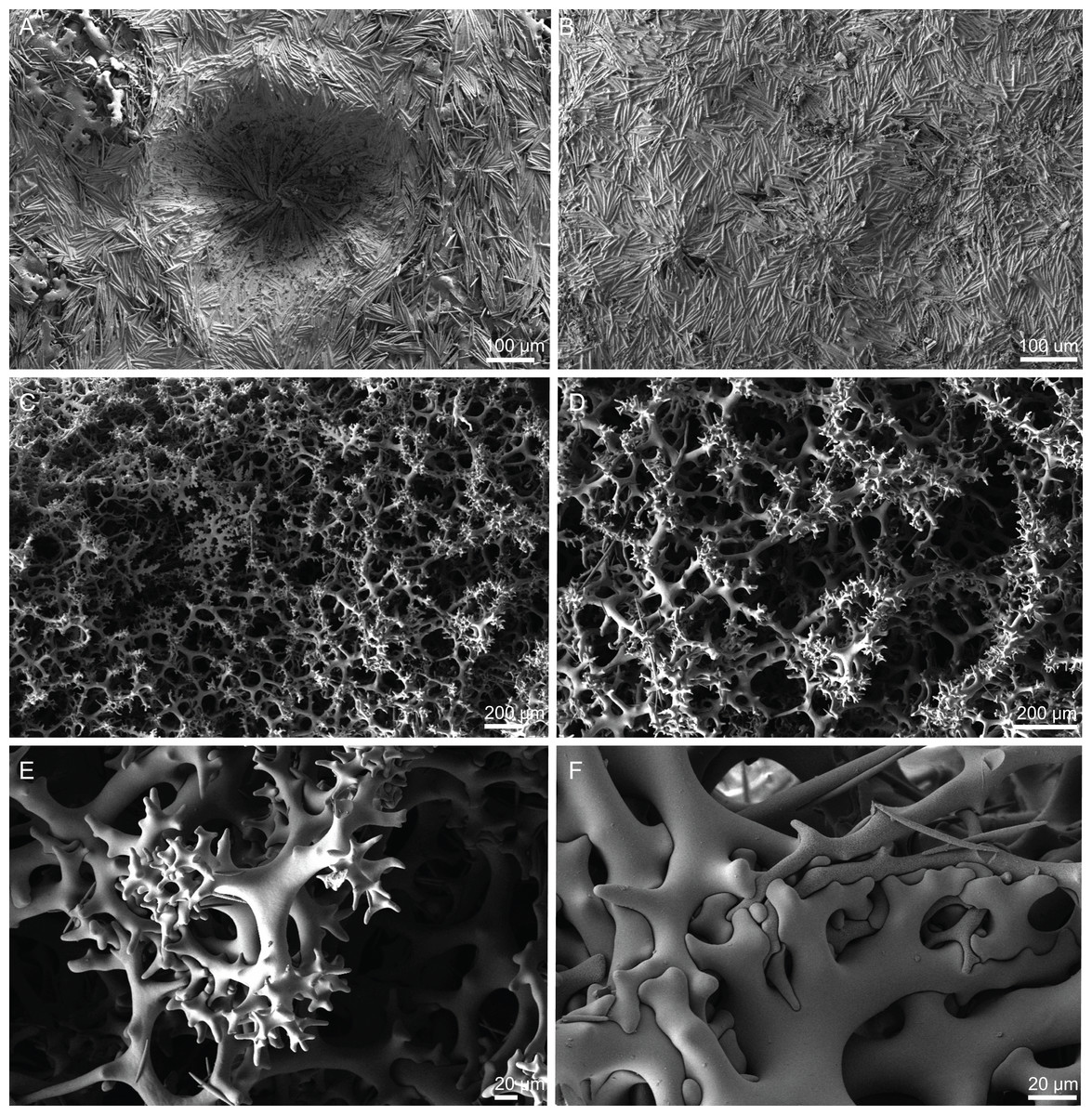
Figure 22: Surface and skeleton of Macandrewia schusterae sp. nov., holotype MNHN-IP-2018-87.
(A) Internal surface with large openings, (B) exterior surface with small openings, (C) overview of choanosomal skeleton, (D) choanosomal desmas, (E) detail of the sculpture of desmas, (F) zygoses.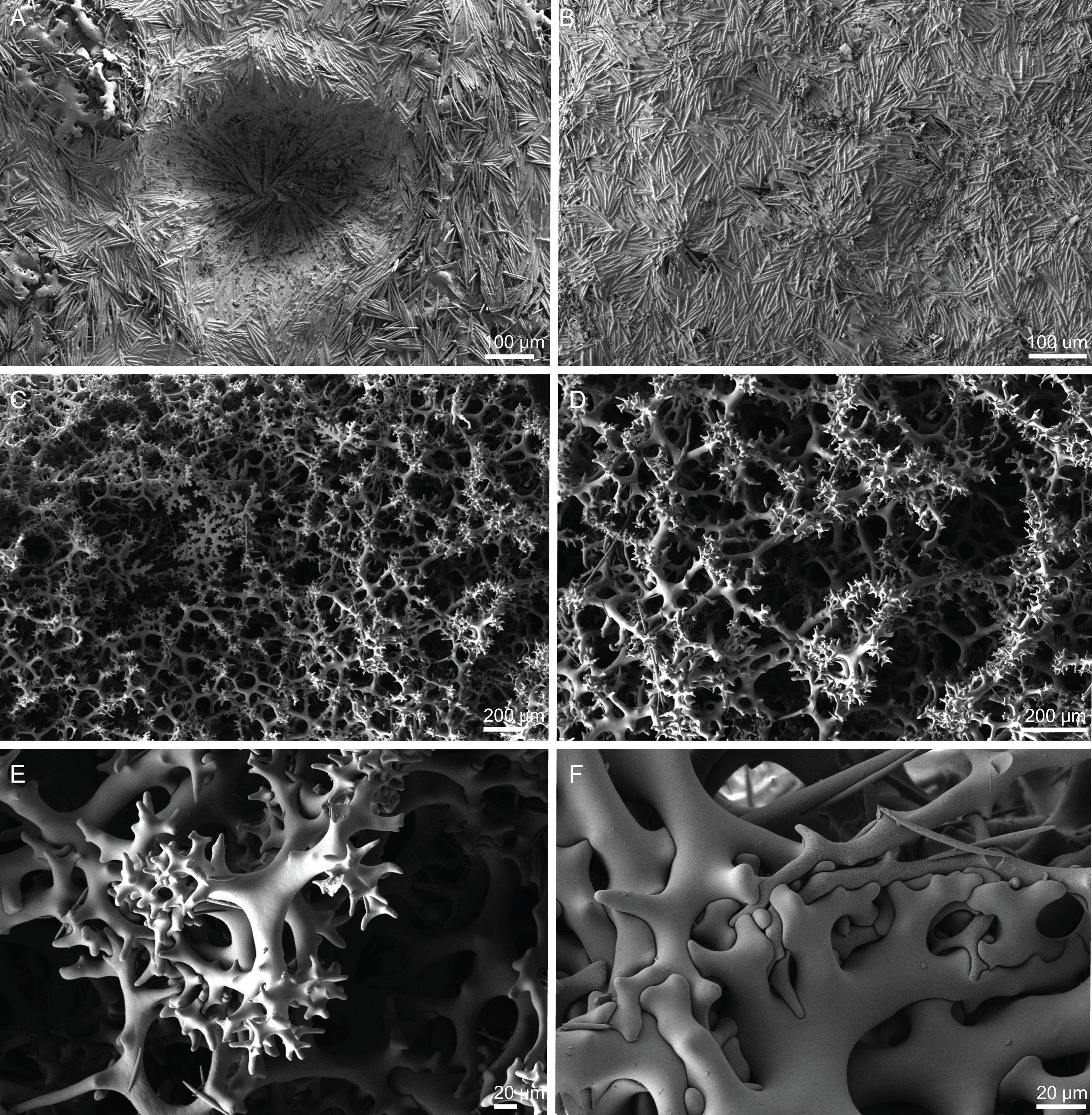{kind=link}
Skeleton. Ectosome has phyllotriaenes that are covered by numerous microxeas, surrounding the openings radially (Figs. 22A and 22B); choanosome has desmas, oxeas and dispersed microxeas; desmas are compact, irregular and create a dense network (Figs. 22C and 22D).
Spicules (holotype MNHN-IP-2018-87).
Desmas, with a triaenose crepsis, smooth, irregular, forming an intricate and complex net, 301–386–463 × 10.2–19.9–39.2 µm in size (Figs. 22C–22E); clones have the terminations splitting in several branches that are usually short, and blunt, 17–37–78 × 5–9–15 µm in size; zygoses is complex and solid (Fig. 22F).
Phyllotriaenes, cladome it is particularly incised on the edges, 177–304–420 µm in diameter; short rhabdome 67–119–178 × 13–21–26 µm in size (Figs. 23A–23E).
Oxeas, smooth with rounded tips, 263–437–620 × 8.1–12.4–16.0 µm in size (Fig. 23F).
Microxeas, smooth, with round edges, 43.8–67.9–95.2 × 2.5–4.3–7.7 µm size (Fig. 23G).
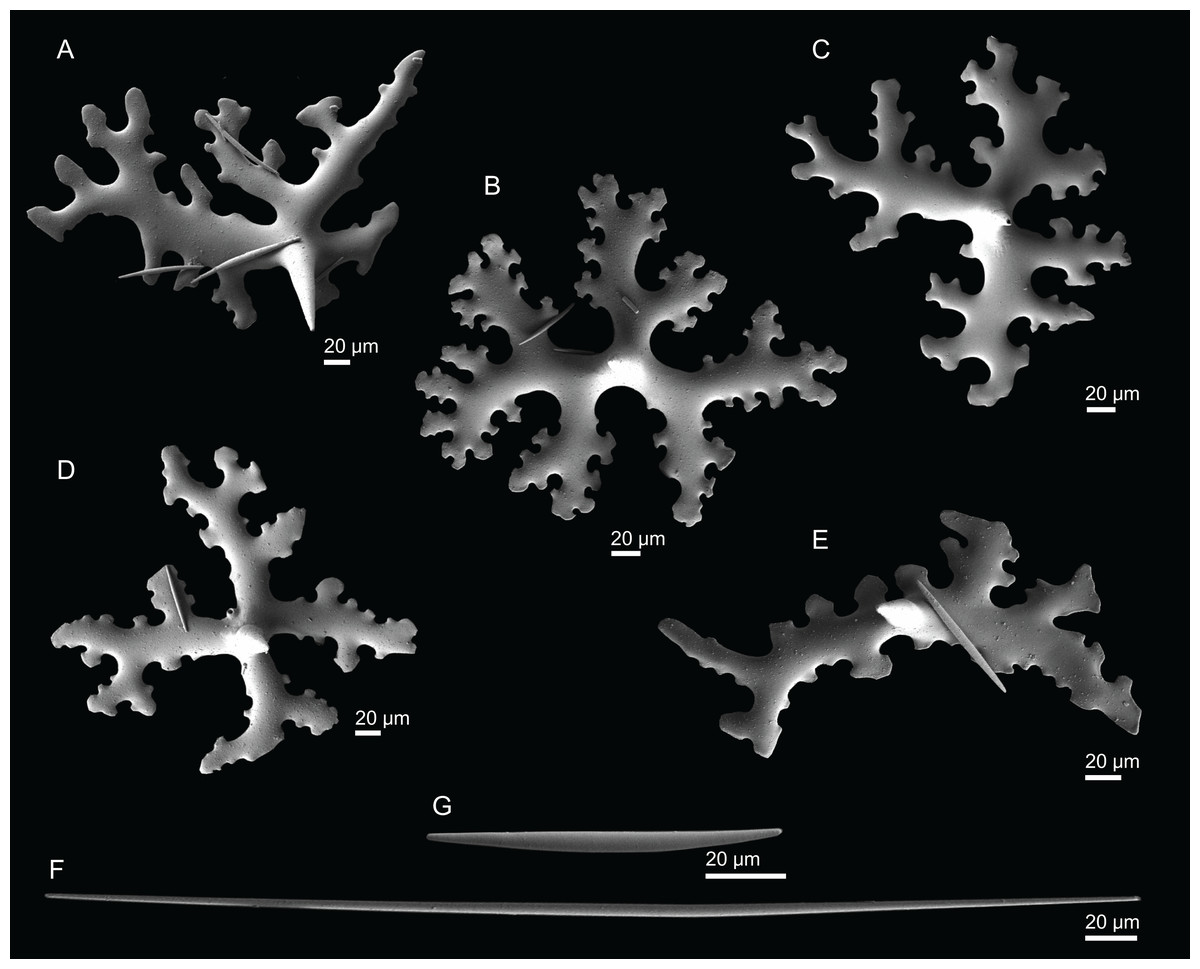
Figure 23: Spicules of Macandrewia schusterae sp. nov., holotype MNHN-IP-2018-87.
(A)–(E) Phyllotriaenes, (F) oxea, (G) microxea.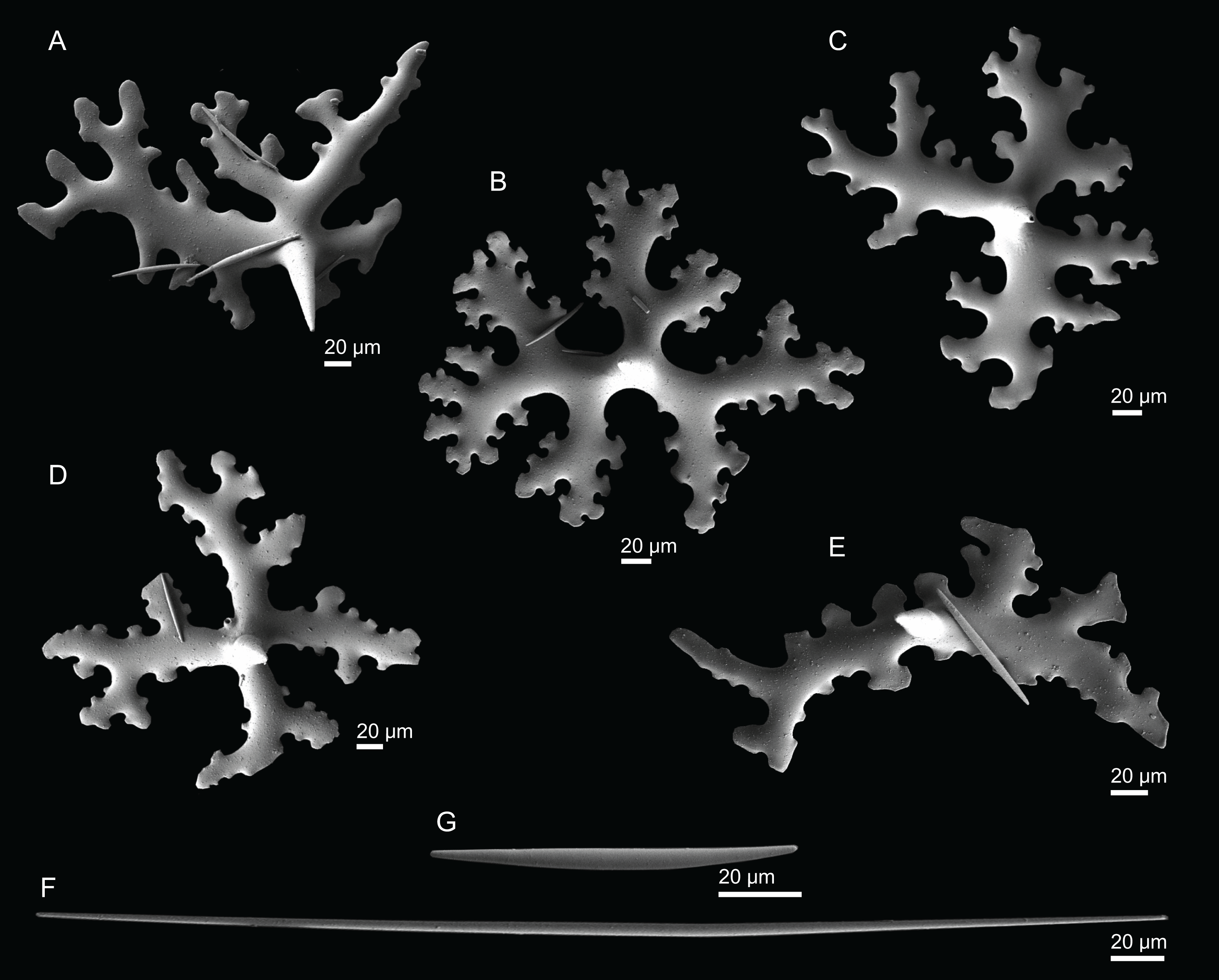{kind=link}
Etymology. Named after Dr. Astrid Schuster for her contributions in the field of molecular paleobiology of lithistid demosponges.
Distribution. This specimen was found on Gorringe, Tyro and Plato Seamounts between 520 and 805 m depth.
Remarks. M. schusterae sp. nov. is here proposed as a new species due to its particular habit, the sculpture of the desmas and size of the spicules. M. schusterae sp. nov. has a foliate shape with contorted lamellas, sometimes resembling a Leiodermatium sp., that contrasts with the ficiform to globular shape with a flattened top in M. robusta, the flabellate to undulate masses with thin lamellas in M. azorica, the ramose shape in M. ramosa Topsent, 1904 and the small ball shape in M. minima sp. nov. The desmas have a different sculpture compared to the other Macandrewia species, as the zygomes have extremely ramified long and thin branches, forming a very strong zygosis (Figs. 22C–22E). This new species also presents a relatively wide range of spicule sizes, mainly on phyllotrianes (cladome: 177–304–420 µm; rhabdome: 67–119–178 µm), oxeas (263–437–620 µm) and microxeas (43.8–67.9–95.2 µm), a feature that is not so common on the other species (Table 3).
Macandrewia minima sp. nov.
Figures 2L, 24–25 and Table 3
urn:lsid:zoobank.org:act:E405AE49-5636-4778-9B07-ED39E9EBB7BE
Holotype. MNHN-IP-2008-222 (1993-01-11, Great Meteor Seamount, epibenthic Warén dredge, DW148, 30°12′N, 28°25′W, 615 m, Seamount 2 campaign).
Comparative material. M. azorica (holotype BMNH 1851.7.28.16, S. Miguel island, Azores; HBOM 003:00784, Selvagens), M. cf. azorica (MNHN-IP-2008-217, Atlantis Seamount; MNHN-IP-2008-220, Atlantis Seamount; MNHN-IP-2008-225, Tyro Seamount; MNHN-IP-2008-226, no data; MNHN-IP-2008-229, Gran Canaria; MNHN-IP-2008-249a, Hyères Seamount), M. robusta (MNHN-IP-2008-216, Hyéres Seamount; MNHN-IP-2008-224, Hyères Seamount), M. schusterae sp. nov. (holotype MNHN-IP-2018-87 and paratype MNHN-IP-2018-90, Gorringe Seamount).
Diagnosis. Small ball shaped Macandrewia with tuberculated phyllotriaenes.
Description (holotype MNHN-IP-2008-222). Small sponge of round-globular shape, 15–20 × 16–17 mm in size, with a very shorth and slender pedicel; surface is smooth with visible openings scattered on the top, 34–69 µm in diameter; colour beige to white in alcohol (Fig. 2L).
Skeleton. Ectosome has a layer of phyllotriaenes covered by large amounts of microxeas; microxeas surround the openings radially (Figs. 24A and 24B); choanosome has desmas, with a triaenose crepsis, forming a compact and irregular network (Fig. 24C); oxeas and microxeas are spread through the choanosome but in small amounts compared to the ectosome.
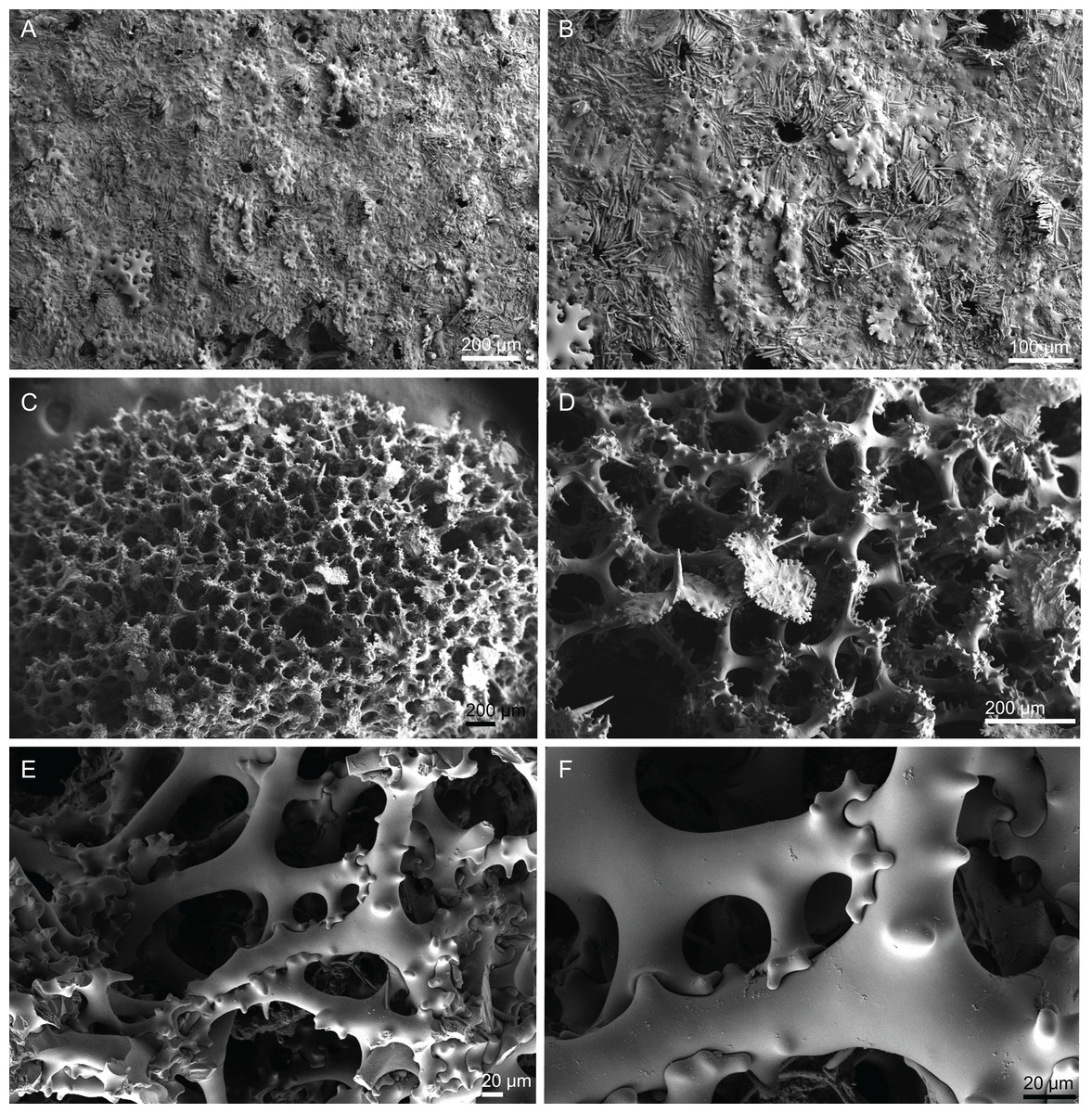
Figure 24: Surface and skeleton of Macandrewia minima sp. nov., holotype MNHN-IP-2008-222.
(A) Surface, (B) close-up of the surface showing the microxeas surrounding the pores radially, (C) overview of desmas, (D) choanosomal desmas, (E) sculpture of desmas, (F) zygosis.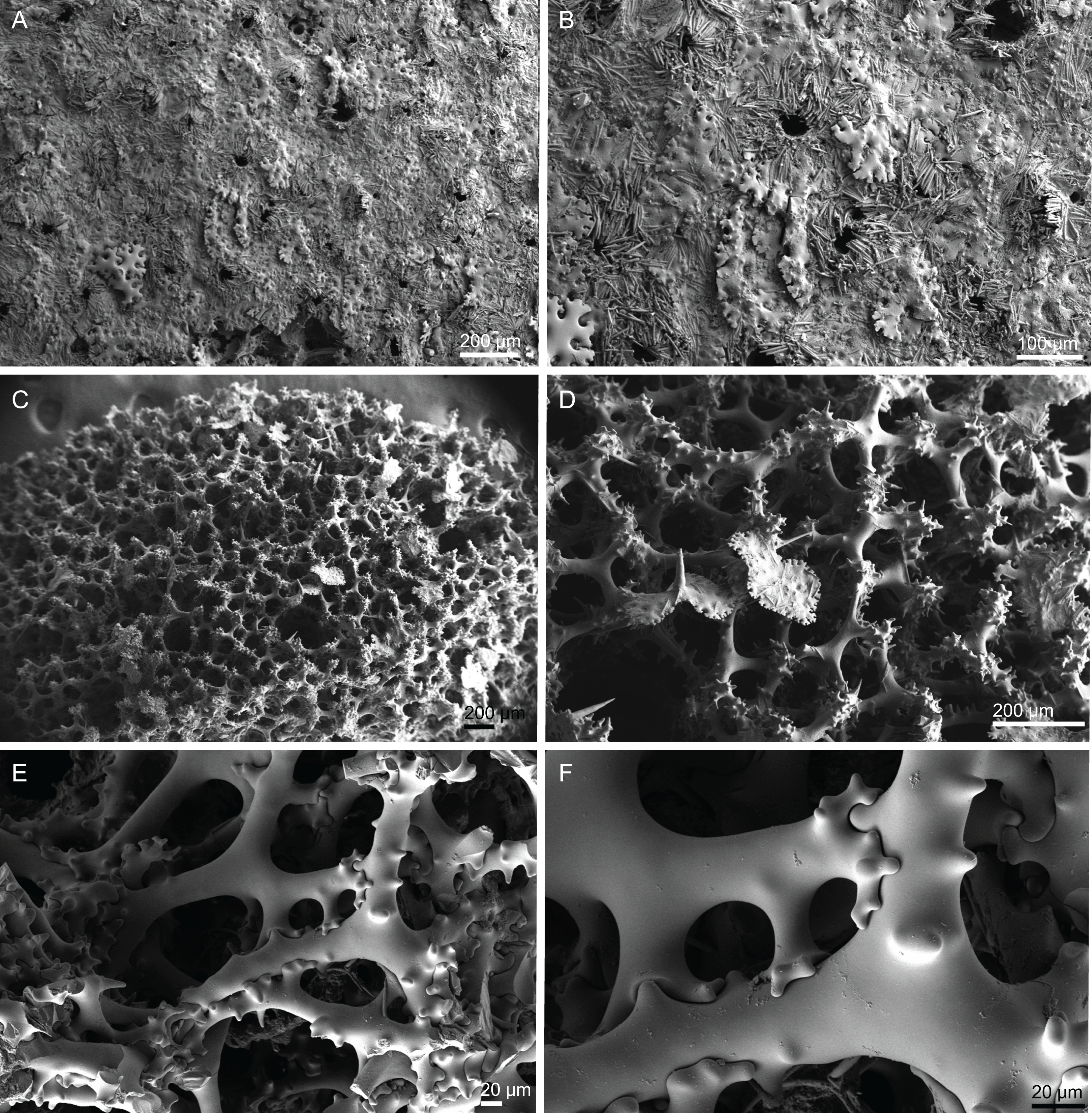{kind=link}
Spicules (holotype MNHN-IP-2008-222).
Desmas, with a triaenose crepsis, robust, usually smooth in the centre, but some branches/rugosities can also be observed, 268–318–348 µm in length and 7–29–50 µm thick (Figs. 24C–24E); clones extremities split in several small branches, 17–37–78 × 5–9–15 in size; zygosis is complex and strong giving a bulb appearance to this part of the desma (Figs. 24D–24F).
Phyllotriaenes, cladome generally more compact, with incised clades that are ornamented by tubercles, 136–222–284 µm in diameter; short rhabdome with a conical shape, 58–99–136 × 14–19–25 µm in size (Figs. 25A–25D).
Oxeas, slightly curved, 197–251–316 × 7.5–11.9–16.2 µm in size (Fig. 25E).
Microxeas, often curved, tips are blunt, 25.9–48.3–74.2 × 3.1–4.4–7.0 µm in size (Figs. 25F and 25G).
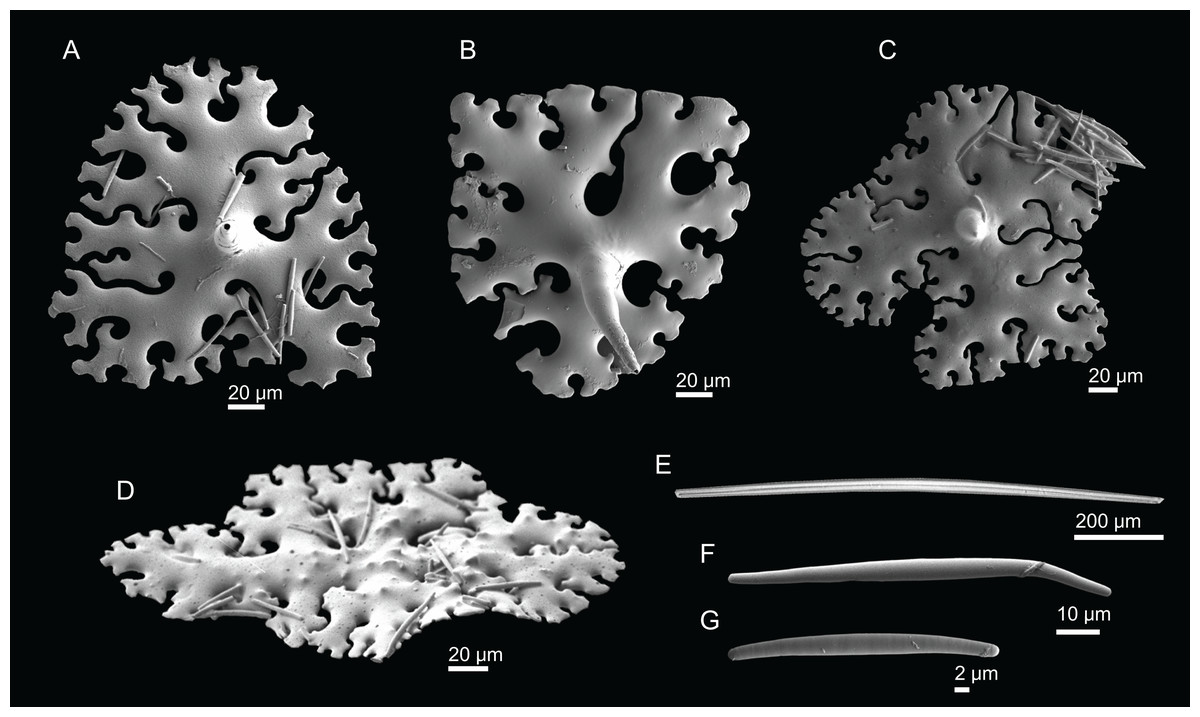
Figure 25: Spicules of Macandrewia minima sp. nov., holotype MNHN-IP-2008-222.
(A)–(C) Bottom view of the cladomes of the phyllotriaenes, (D) top view of cladome showing the small protuberances, (E) oxea-tips are broken, (F) and (G) microxeas.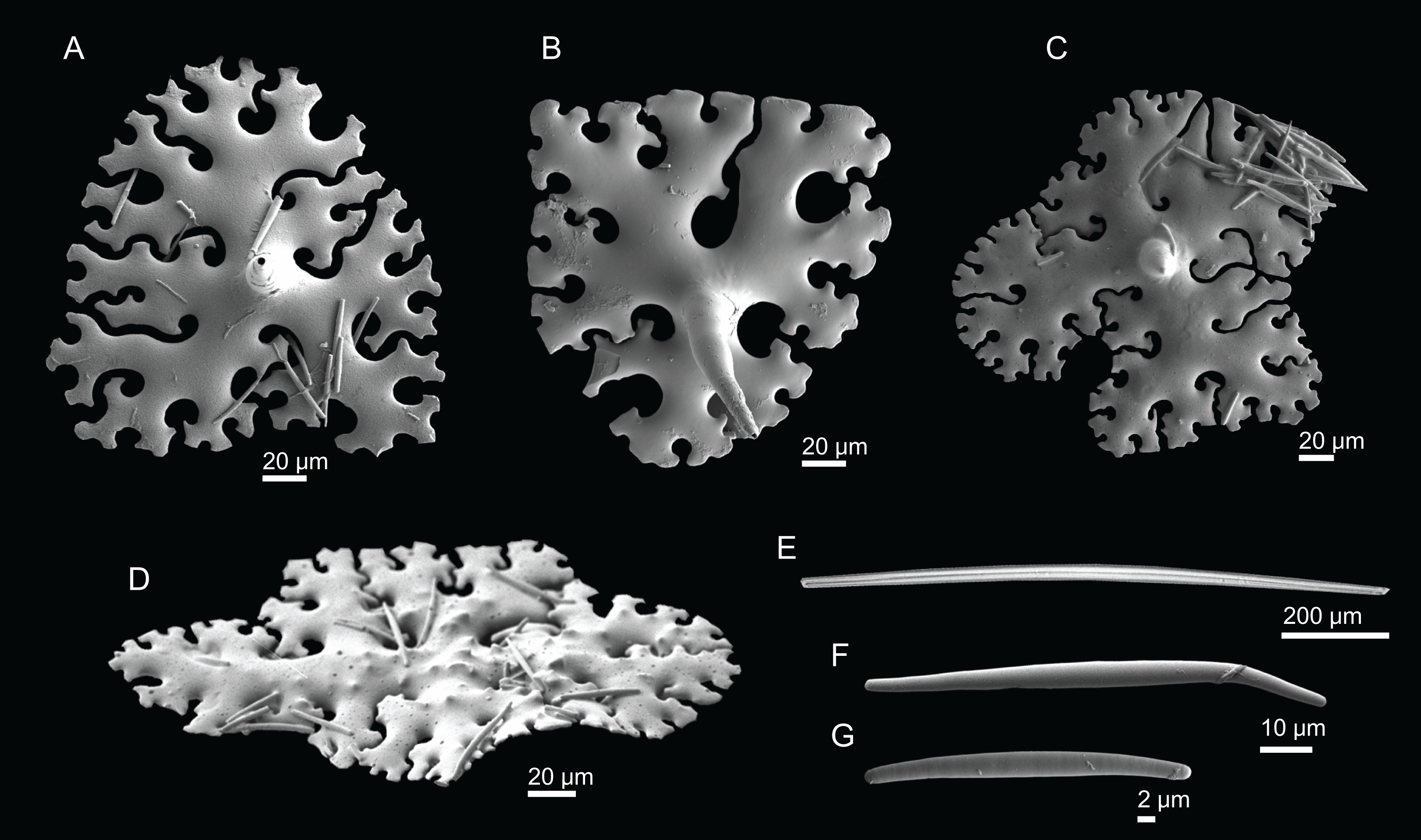{kind=link}
Etymology. From the Latin minima = small.
Distribution. Only known from its type locality, the Great Meteor Seamount at 615 m depth.
Remarks. M. minima sp. nov. differs from the other Macandrewia in the considerably smaller size of its spicules (see Table 3), its globular shape and in the characteristic tubercles of the phyllotriaenes (only observed in this species).
Family Phymaraphiniidae Schrammen, 1924
Genus Exsuperantia Özdikmen, 2009
Synonymy. Rimella Schmidt, 1879: 21 (preoccupied); Racodiscula sensu Topsent, 1928, 1904, 1892 (wrong genus identification).
Diagnosis. Clavate to columnar Phymaraphiniidae with phyllotriaenes as ectosomal spicules (Pisera & Lévi, 2002f).
Definiton. Clavate to columnar, globular knob-like small sponges. Desmas are triders with smooth and/or tuberculated tubercles. Other spicules are smooth phyllo- to discotriaenes and subtylostyles to tylotes as megascleres, and acanthomicroxeas, acanthorhabds and streptasters as microscleres (emended after Carvalho & Pisera, 2019; Pisera & Lévi, 2002e).
Type species. Exsuperantia clava (Schmidt, 1879) (type by monotypy).
Exsuperantia archipelagus Carvalho & Pisera, 2019
Figures 3A, 26–27 and Table 4
| Species | Habitus | Size | Triders | Phylo- to discotriaenes | Subtylostyles to tylotes | Acanthomicroxeas | Acanthorhabds | Amphiasters | Locality |
|---|---|---|---|---|---|---|---|---|---|
|
1E. clava (Schmidt, 1879) (Syntype MZUS PO146) |
Cylindrical to clavate | Up to 30 mm long, 10 mm thick | 230–320 µm in size | – | Present | Fusiform, occasionally centrotylotes | Present | Slender rays | Cuba (depth unknown) |
| 2E. archipelagus Carvalho & Pisera, 2019 (Holotype MNHN DT-782/1) | Columnar to ficiform, with or without lateral protuberances or branches; surface is smooth, with marked water canals; colour beige to whitish | 20–30 mm × 10–20 mm | 409–693 × 52–98 µm in size | Phyllotriaenes. Cladome: 640–890 µm in diameter; rhabdome: 229–320 × 71 µm | Subtylostyles to tylostyles: 260–1114 × 3–38 µm in size | 31–47 × 2.9–4.2 µm | 18–24 × 2.3–4.1 µm | 15–19 × 1.2–1.7 µm | Azores (168–594 m depth) |
|
E. archipelagus (MNHN-IP-2008-196). |
Columnar to ficiform in habitus, with or without lateral protuberance; water canals visible on the surface; colour beige | 22–23 × 8–18 mm | 260–362–464 (n = 7) × 15–29–44 µm thick (n = 15) | Phyllotriaenes. Cladome: 199–358–470 µm in diameter (=11); rhabdome: 140 × 34.4 µm (n = 1) | 296–515–618 × 6.1–9.7–13.4 µm (n = 11) | 18.6–25.0–44.0 × 1.4–2.2–3.7 µm | 7.2–12.0–15.9 × 1.2–1.8–3.2 µm | 5.3–8.6–15.0 µm (n = 15) | Hyères Seamount (310 m depth) |
|
E. levii sp. nov. (Holotype MNHN-IP-2008-201) |
Clusters of globular to ficiform knob-like short fingers | 293–346–503 µm in size, 28–45–67 µm thick (n = 12) | Phyllo- to discotriaenes. Cladome: 143–299–486 diameter (n = 20); rhabdome: 25–73–130 × 10 –28–44 µm (n = 10) |
234–307–436 µm × 8.6–9.8–11.3 µm (n = 6) | 21.6–28.2–35.6 × 1.7–2.6–3.8 µm | 10.3–14.1–19.3 × 1.9–2.7–3.5 µm (n = 25) | 7.4–10.0–14.8 µm (n = 20). | Hyères Seamount (480 m depth) |
Synonymy. Racodiscula sensu Topsent, 1892 (Topsent, 1904, 1928) (wrong generic assignment); Exsuperantia sp. Carvalho, Pomponi & Xavier (2015).
Material. MNHN-IP-2008-191 (1993-02-06, Tyro Seamount, epibenthic Warén dredge, st. DW277, 34°00′N, 28°21′W, 1,000 m), MNHN-IP-2008-192 (1993-02-03, Atlantis Seamount, epibenthic Warén dredge, st. DW265, 34°29′N, 30°36′W, 545 m), MNHN-IP-2008-195 (1993-02-02, Atlantis Seamount, beam trawl, st. CP257, 34°04′N, 30°15′W, 338 m), MNHN-IP-2008-196 (1993-01-17, Hyères Seamount, epibenthic Warén dredge, st. DW188, 31°30′N, 29°00′W, 310 m), MNHN-IP-2008-199 (1993-02-02, Atlantis Seamount, epibenthic Warén dredge, st. DW258, 34°00′N, 30°12′W, 420 m), MNHN-IP-2008-200 (1993-01-31, Plato Seamount, epibenthic Warén dredge, st. DW242, 33°12′N, 28°57′W, 710 m), MNHN-IP-2008-202 (1993-02-02, Atlantis Seamount, epibenthic Warén dredge, st. DW254, 34°05′N, 30°13′W, 480 m), MNHN-IP-2008-240 (1993-02-01, Plato Seamount, epibenthic Warén dredge, st. DW246, 33°14′N, 29°36′W, 520 m), MNHN-IP-2008-243 (1993-02-02, Atlantis Seamount, epibenthic Warén dredge, st. DW258, 34°00′N, 30°12′W, 420 m). All from the Seamount 2 campaign.
Comparative material examined. E. archipelagus (holotype MNHN DT 782/1 Azores; paratype MNHN DT 782/2, Azores; paratype DOP 1976, Azores); E. levii sp. nov. (holotype MNHN-IP-2008-201, Hyères Seamount).
Diagnosis. Columnar to ficiform Exsuperantia with trider-type desmas that have smooth tubercles (few presenting rugosities).
Description (MNHN-IP-2008-196). Small phymarapiniid 22–23 × 8–18 mm in size, columnar to ficiform in habitus, with or without lateral protuberances (Fig. 3A); some specimens have a “V” shape morphology; surface is smooth with conspicuous subdermal water canals giving a striped appearance to the sponge; oscula or pores are not visible; colour beige in ethanol.
Skeleton. Ectosome is formed by a layer of phyllotriaenes covered by large amounts of microscleres: openings are surrounded by these microscleres; choanosomal skeleton is mainly built of trider-type desmas, that form a regular network with large spaces in between (Fig. 26); some subtylostyles (Fig. 26B) and microscleres are also present and spread through the skeleton.
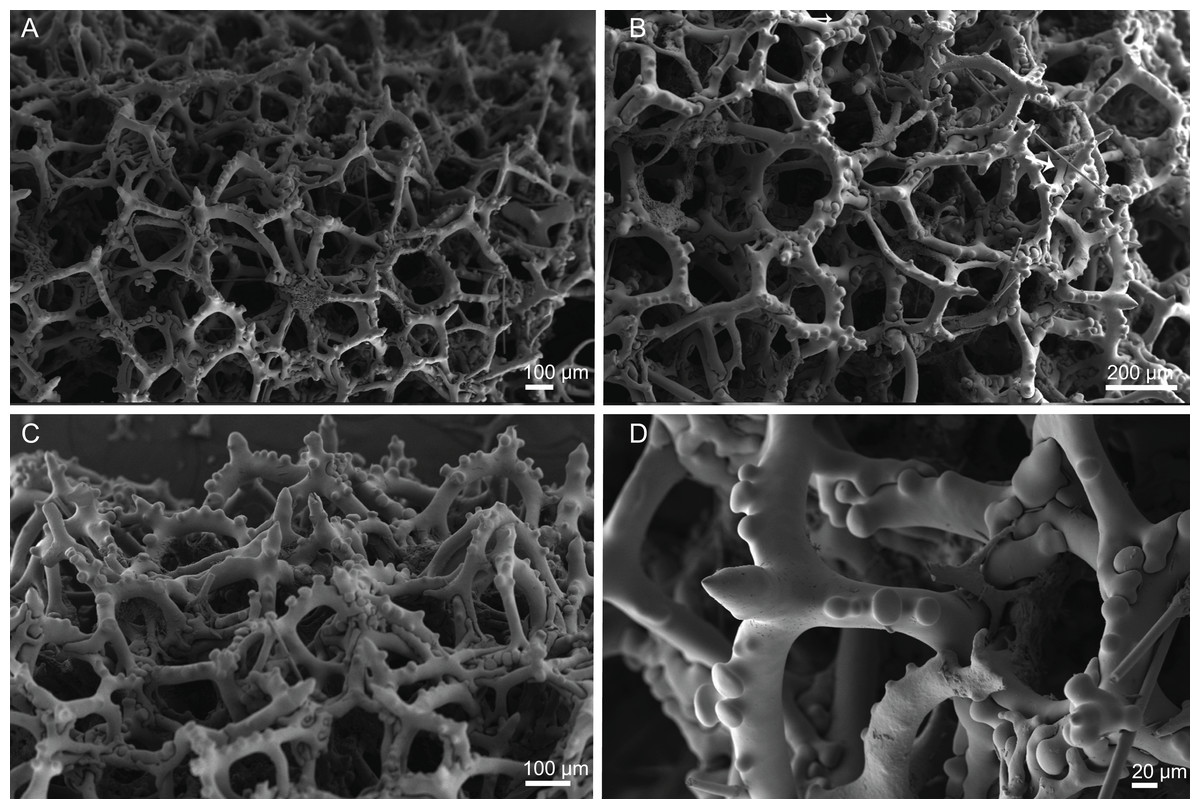
Figure 26: Skeleton of Exsuperantia archipelagus Carvalho & Pisera, 2019, specimen MNHN-IP-2008-196.
(A) Overview of choanosomal triders, (B) subtylostyles crossing the skeleton, (C) detail of trider-type desmas, (D) zygosis and close up of a trider showing the desma ornamentation.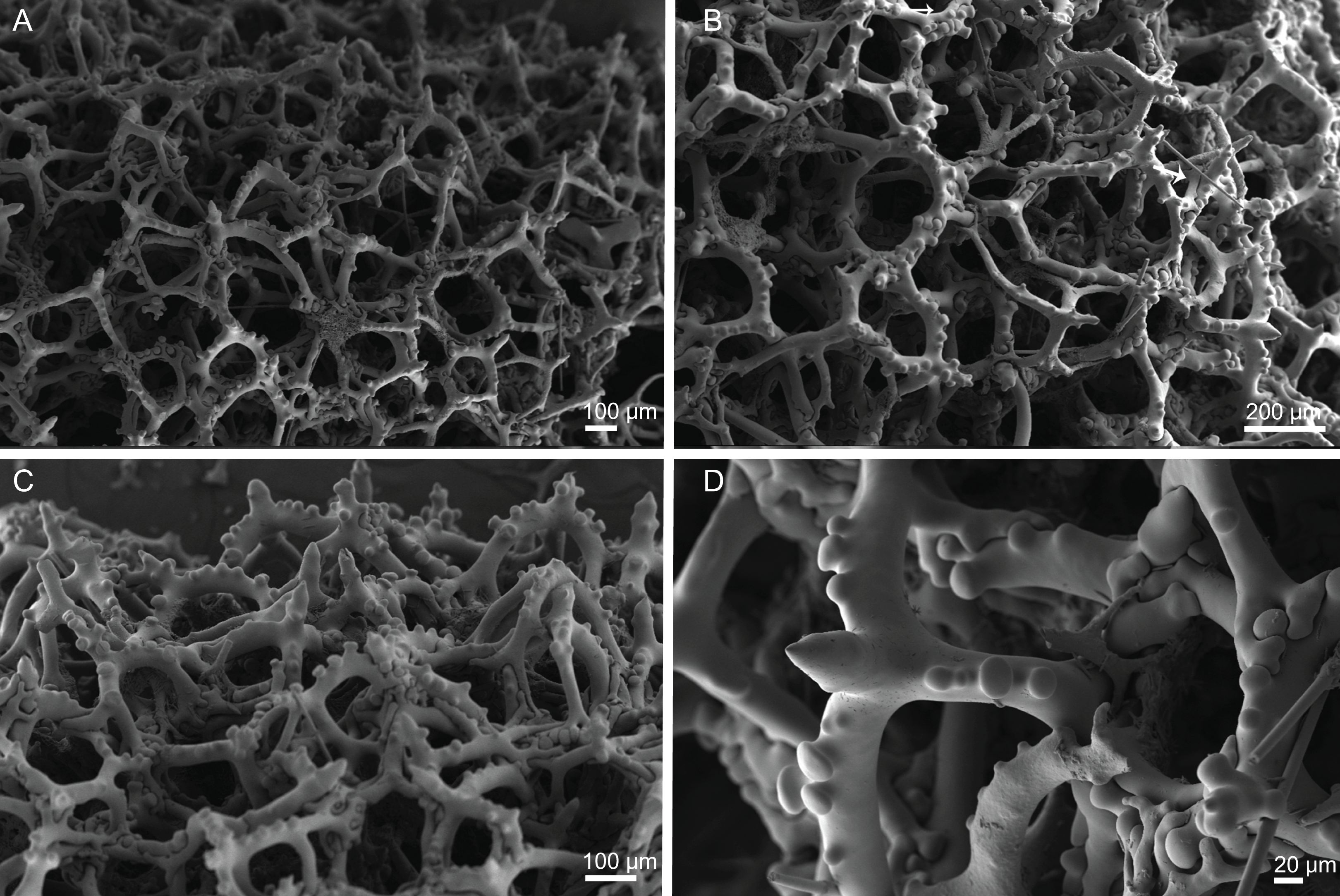{kind=link}
Spicules (MNHN-IP-2008-196).
Trider-type desmas, smooth, very tuberculated, 261–342–419 × 23–30–44 µm in size (Figs. 26A–26C); tubercles are smooth, sometimes with rugosities, 7–10–11 µm diameter; tip of the trider is smooth and has a conical shape (Fig. 26D).
Phyllotriaenes, irregular, smooth cladome 412–450–493 µm in diameter (Figs. 27A–27D), long rhabdome, 43–75–126 µm in size, with pointed tip (Fig. 27A).
Subtylostyles to tylotes, smooth, 401–542–629 × 6.0–9.9–12.8 µm in size (Fig. 26B).
Acanthomicroxeas, slender, with sharp tips, 16.8–22.1–28.1 × 1.6–2.2–3.1 µm (Fig. 27E).
Acanthorhabds, thick with blunt ends, 9.8–12.7–17.5 × 1.2–2.0–2.8 µm (Fig. 27F).
Amphiasters, with several arms covered by spines, 5.0–6.6–8.6 µm long (Figs. 27G and 27H).

Figure 27: Spicules of Exsuperantia archipelagus Carvalho & Pisera, 2019, specimen MNHN-IP-2008-196.
(A)–(D) Phyllotriaenes, (E) acanthomicroxeas, (F) acanthorhabds, (G) and (H) streptasters/amphiasters.{kind=link}
Distribution. E. archipelagus was found in Tyro, Hyères, Atlantis, and Plato Seamounts between 280 and 1,000 m depth and also in Gran Canaria island at 660 m depth.
Remarks. The size of the spicules measured in these specimens are considerable smaller when compared to the type material (Carvalho & Pisera, 2019) (Table 4), but see Discussion for more information regarding this topic.
Exsuperantia levii sp. nov.
Figures 3B, 28–29 and Table 4
urn:lsid:zoobank.org:act:24B5934A-4767-4429-B172-A649C4CE0D83
Holotype. MNHN-IP-2008-201 (1993-01-16, Hyères Seamount, epibenthic Warén dredge, st. DW182, 31°23′N, 28°54′W, 480 m, Seamount 2 campaign).
Comparative material examined. E. archipelagus (holotype MNHN DT 782/1 Azores; paratype MNHN DT 782/2, Azores; paratype DOP 1976, Azores).
Diagnosis. Clusters of globular to ficiform knob-like short fingers with apical osculum; phyllo- to discotriaenes as ectosomal megascleres.
Description (holotype MNHN-IP-2008-201). Clusters of globular to ficiform knob-like short fingers, 30 mm in length and 29 mm wide; oscula, approximately 2 mm in diameter, are located on the top of the knobs (Fig. 3B); surface is rugose with a striated appearance due to the visible subdermal water canals; colour is brown in ethanol.
Skeleton. Ectosome is composed by phyllo- to discotriaenes that are very variable in shape, and several microscleres; choanosomal skeleton has regular and articulated triders, forming an irregular and relatively loose network (Fig. 28); subtylostyles are present crossing the skeleton (Figs. 28A and 28B); microscleres are present and very abundant, except for streptasters that are less numerous.

Figure 28: Skeleton of Exsuperantia levii sp. nov., specimen MNHN-IP-2008-201.
(A) Outline of trider-type desmas, (B) triders, (C) zygosis, (D) detail of a trider desma.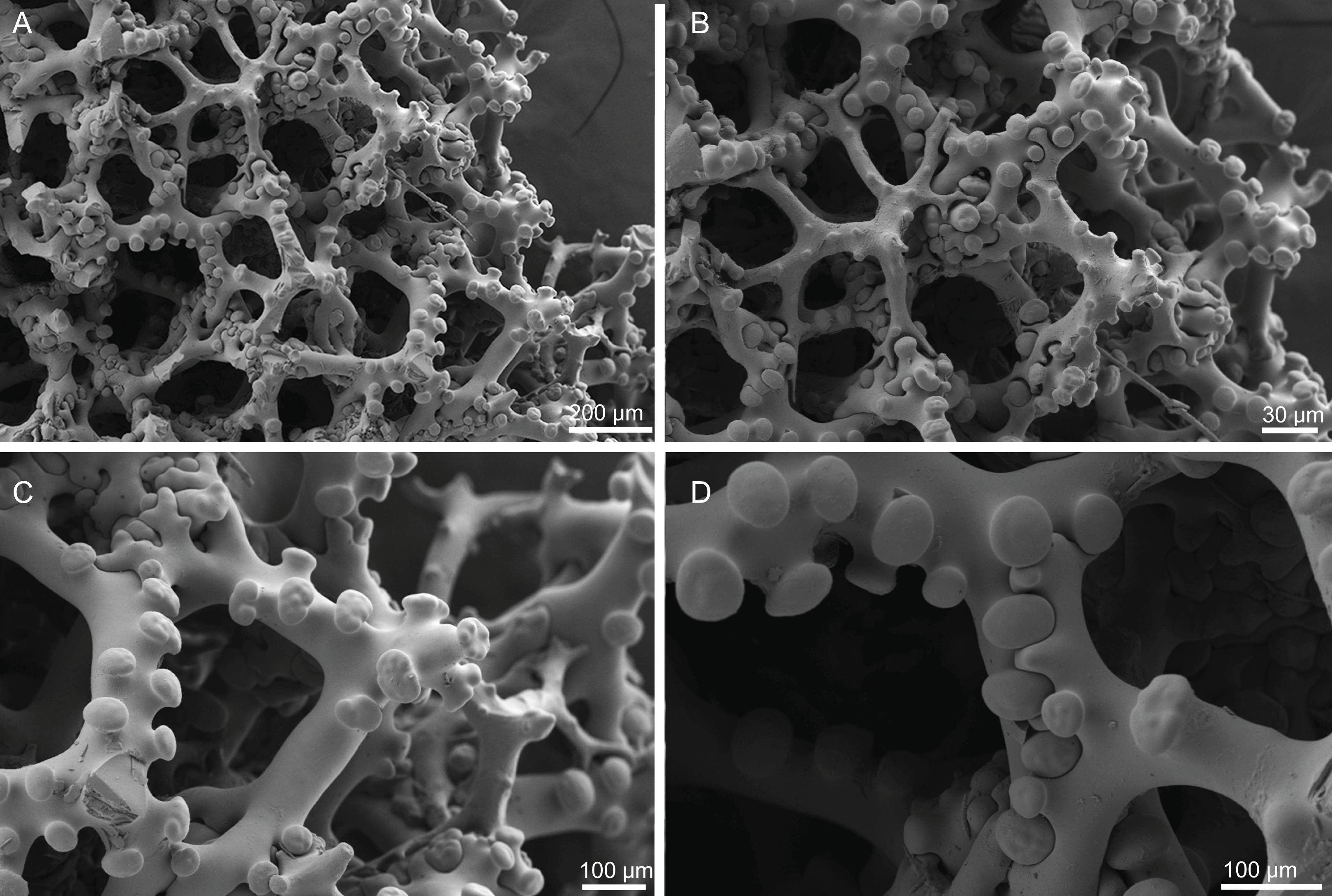{kind=link}
Spicules (holotype MNHN-IP-2008-201).
Trider-type desmas, regular, smooth, 293–346–503 × 28–45–67 µm in size, with large and flattened tubercles that can be smooth or very tuberculated, 15.4–21.2–29.9 µm in diameter (Figs. 28A–28D).
Phyllo- to discotriaenes, smooth, cladome very variable in shape, 143–299–486 µm in diameter; rhabdome has a conical shape and a sharp tip, 25–73–130 µm × 10–28–44 µm in size (Figs. 29A–29F).
Subtylostyles, smooth, large, slightly curved, 234–307–436 × 8.6–9.8–11.3 µm in size (Figs. 28A and 28B).
Acanthomicroxeas, thin, slightly curved, with sharp tips, 21.5–26.2–31.6 × 1.8–2.9–4.1 µm (Figs. 29G and 29H); occasionally, these spicules are irregular, and exhibit one sharp and one blunt tip, resembling an intermediate stage between an acanthomicroxea and an acanthorhabd (Fig. 29I);
Acanthorhabds, small, robust, 9.3–15.1–22.5 × 1.6–2.8–3.8 µm in size (Figs. 29J and 29K).
Amphiasters, thin with spiny arms, 5.9–8.2–11.5 µm long (Figs. 29L and 29M).
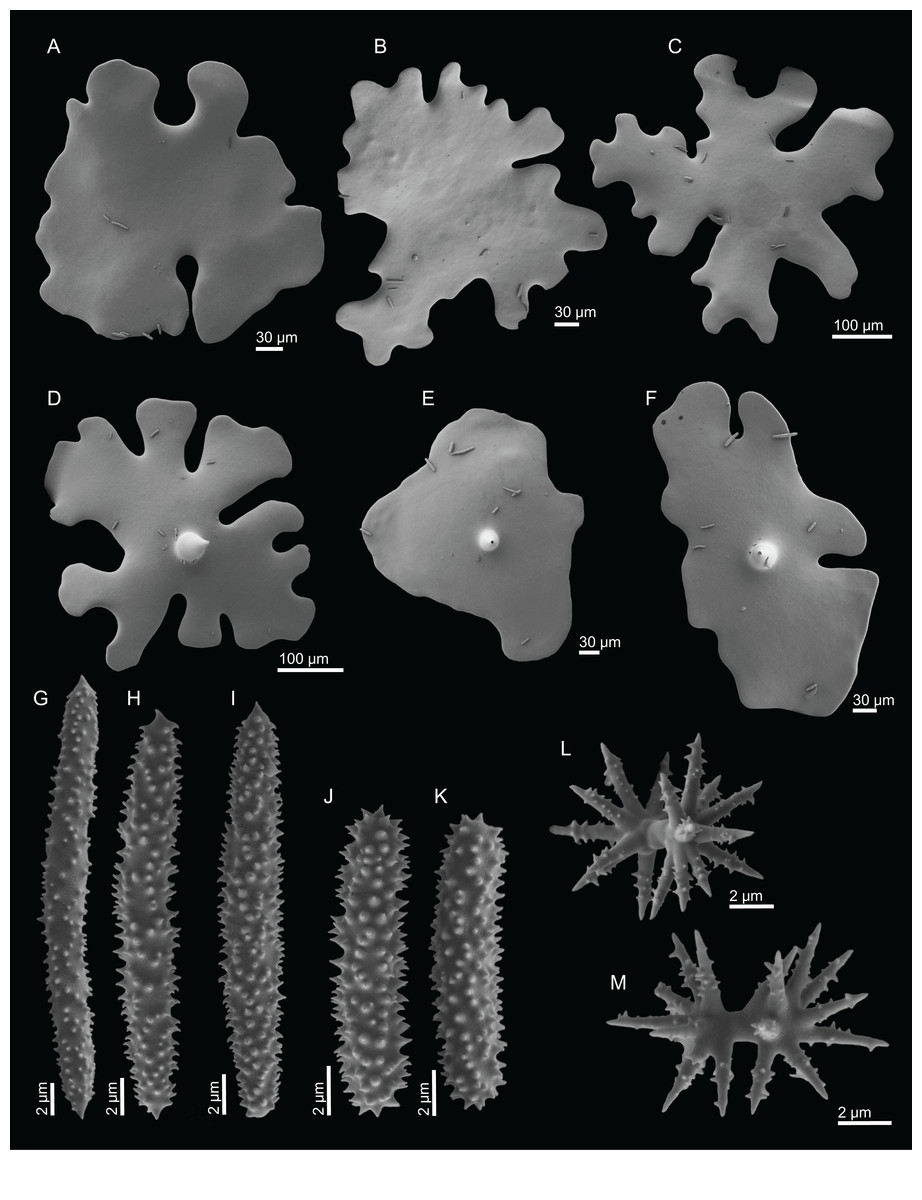
Figure 29: Spicules of Exsuperantia levii sp. nov., specimen IP-2008-201.
(A)–(C) Top view of cladomes of phyllo- to discotriaenes, (D) and (E) bottom view of cladomes, (G)–(I) acanthomicroxeas, (J) and (K) acanthorhabds, (L) and (M) streptasters/amphiasters.{kind=link}
Distribution. Exsuperantia levii sp. nov. is known from its type locality, the Hyères Seamount at 480 m depth.
Etymology. Named after Professor Claude Lévi from the Muséum National d’Histoire Naturelle Paris (MNHN) for his lifelong contribution to the taxonomy and systematics of Porifera, including lithistid sponges.
Remarks. Recently, a revision of the genus Exsuperantia allowed to clarify some taxonomic problems by establishing two species, E. clava (NWA) and E. archipelagus (NEA), that were previously considered a single species (Carvalho & Pisera, 2019). According to the authors, the main differences between these two species are the desmas morphology and ornamentation.
Here we propose E. levii sp. nov. as a new species, third of the genus, based not only on desmas morphology and ornamentation, but also on the habitus of this new species. The trider-type desmas on E. levii sp. nov. resemble the ones found in E. clava, i.e., the tubercles are ornamented and the tip of the trider has a tubercle, while in E. archipelagus it usually has a conical shape. In general, the size of the spicules of E. levii sp. nov. is smaller when compared to the holotype E. archipelagus (unfortunately the size of spicules of the E. clava is not known, with exception of the desmas, since the type material was deciduous and microscleres were not present (Pisera & Lévi, 2002f)), however, the most distinct feature is the habitus of E. levii sp. nov.: a cluster of globular knob-like fingers with large apical oscula on top, contrasting with the columnar to ficiform morphology of the other two species.
Suborder Spirophorina Bergquist & Hogg, 1969
Family Azoricidae Sollas, 1888
Genus Leiodermatium Schmidt, 1870
Diagnosis. Azoricidae with spiny rhizoclones and diactines as megascleres; ectosomal spicules and microscleres are absent (Pisera & Lévi, 2002i).
Definition. Lamellate, plate-like, foliose, vase- or ear-shaped Azoricidae; oscules are visible; choanosomal desmas are spiny rhizoclones; megascleres are diactines; microscleres are absent (Kelly, 2007; modified from Pisera & Lévi, 2002h).
Type species. Leiodermatium lynceus Schmidt, 1870.
Leiodermatium lynceus Schmidt, 1870
Figures 3C, 30–31 and Table 5
| Habitus | Size | Rhizoclones | Oxeas | Locality | |
|---|---|---|---|---|---|
| 1Leiodermatium lynceus Schmidt, 1870 (Holotype MZUS PO145) | Foliate or vase to ear-shape; outer surface with large and elevated oscules (500–750 µm) and inner surface with small pores (156–188 µm) | 60 × 30 mm in size; walls are 3–4 mm thick | – | Not found in the holotype | Portugal (depth unknown) |
| L. lynceus Schmidt, 1870 (MNHN-IP-2008-93) | Foliate to undulate polymorphic masses, with large oscules in the outer surface (243–269 µm) of the sponge and small pores in the external one (68–145 µm); colour beige to brown | 90–93 mm wide; walls 5–12 mm thick | 156–179–223 µm (n = 6) long and 8.4–19.4–49.9 µm thick (n = 30) | up to 1mm long and 8.5–9.6–10.7 µm thick (n = 5) | Gorringe seamount (305–320 m depth) |
| 2Leiodermatium pfeifferae (Carter, 1873) (unknown type) | Flattish, cabbage-like, infoliated, with branched sinuous laminae, vertically, widely separated, and proliferous; ostia (vents) are little raised on papillary eminences and scattered over the inner of the laminae; pores are on the outer laminae | 360 mm in diameter and 280 mm vertical diameter; walls 6–17 mm thick | – | Fusiform, growing on the edge/margins of the specimen (measurements were not given) | Madeira (684 m depth) |
| L. tuba sp. nov. (Holotype MNHN-IP-2018-72) | Large sponges, lamellate vase to contorted walls, sometimes forming a tube, with smooth and similar surfaces; colour is beige to brown | 138 mm long and 93 mm wide; walls 4–5 mm thick | 141–173–211 (n = 4) × 12.1–18.7–31.0 µm (n = 19) | up to 1 mm long and 5.9–8.1–9.8 µm thick (n = 3) | Gorringe seamount (805–830 m depth) |
| L. tuba sp. nov. (Paratype MNHN-IP-2018-73) | Small fragment, of tubular shape, with thin walls; outer surface is smooth but with a stripe appearance due the water canals underneath the surface; inner surface has a white appearance given the numerous oxeas piercing the surface | 65 mm long and 25 mm wide; walls are 5–6 mm thick | Gorringe seamount (805–830 m depth) | ||
|
3L. deciduum (Schmidt, 1879) (Holotype MZUS PO167). Incertae sedis |
Ear shaped, or irregular vase shaped sponge; upper side of chaonosomal skeleton, with numerous oscula, 500 µm in diameter; lower side of choanosome with numerous pores, 200–250 µm in diameter | 35 mm high, 32 mm wide; walls 10 mm thick | – | – | Gulf of Mexico (183–1,472 m depth) |
Notes:
‘–’ indicates the information was not given in the description.
Synonym. Azorica pfeifferae var. tenuilaminaris Sollas, 1888 (genus transfer and junior synonym).
Material examined. MNHN-IP-2018-93 (1988-09-24, Gorringe Seamount, beam trawl, st. CP20, 36°33.7′N, 11°30.1′W, 305–320 m, Seamount 1 campaign), MNHN-IP-2008-239 (1993-01-16, Hyères Seamount, epibenthic Warén dredge, st. DW182, 31°23′N, 28°54′W, 480 m, Seamount 2 campaign).
Comparative material examined. L. tuba sp. nov. (holotype MNHN-IP-2018-72, Gorringe Seamount; paratype MNHN-IP-2018-73, Gorringe Seamount).
Diagnosis. Foliate to undulate polymorphic masses, with large openings in the outer surface of the sponge and small openings in the inner surface.
Description (MNHN-IP-2018-93). Large foliate to undulate irregular masses, with thick lamellas, 5–12 mm, that in some cases can form cups/funnels (Fig. 3C); inner and outer surfaces are different from each other, and it is possible to distinguished at naked eye; outer surface has larger openings slightly elevated from the surface, 243–269 µm in diameter, (Figs. 3C, 30A and 30C) while the inner surface is smooth with small openings, 68–145 µm in diameter, evenly distributed (Figs. 3C, 30B and 30D); both surfaces are heavily protruded by long oxeas; colour varies from beige to brown in ethanol.

Figure 30: Surface and skeleton of Leiodermatium lynceus Schmidt, 1870, specimen MNHN-IP-2018-93.
(A) Overview of the outer surface with larger and elevated oscula (surface not digested in nitric acid), (B) overview of the inner surface with smaller and depressed pores (surface not digested), (C) overview of the outer surface with larger and elevated pores (digested surface in nitric acid), (D) overview of the inner surface with smaller pores (digested surface), (E) detail of the oscula, (F) detail of the pores.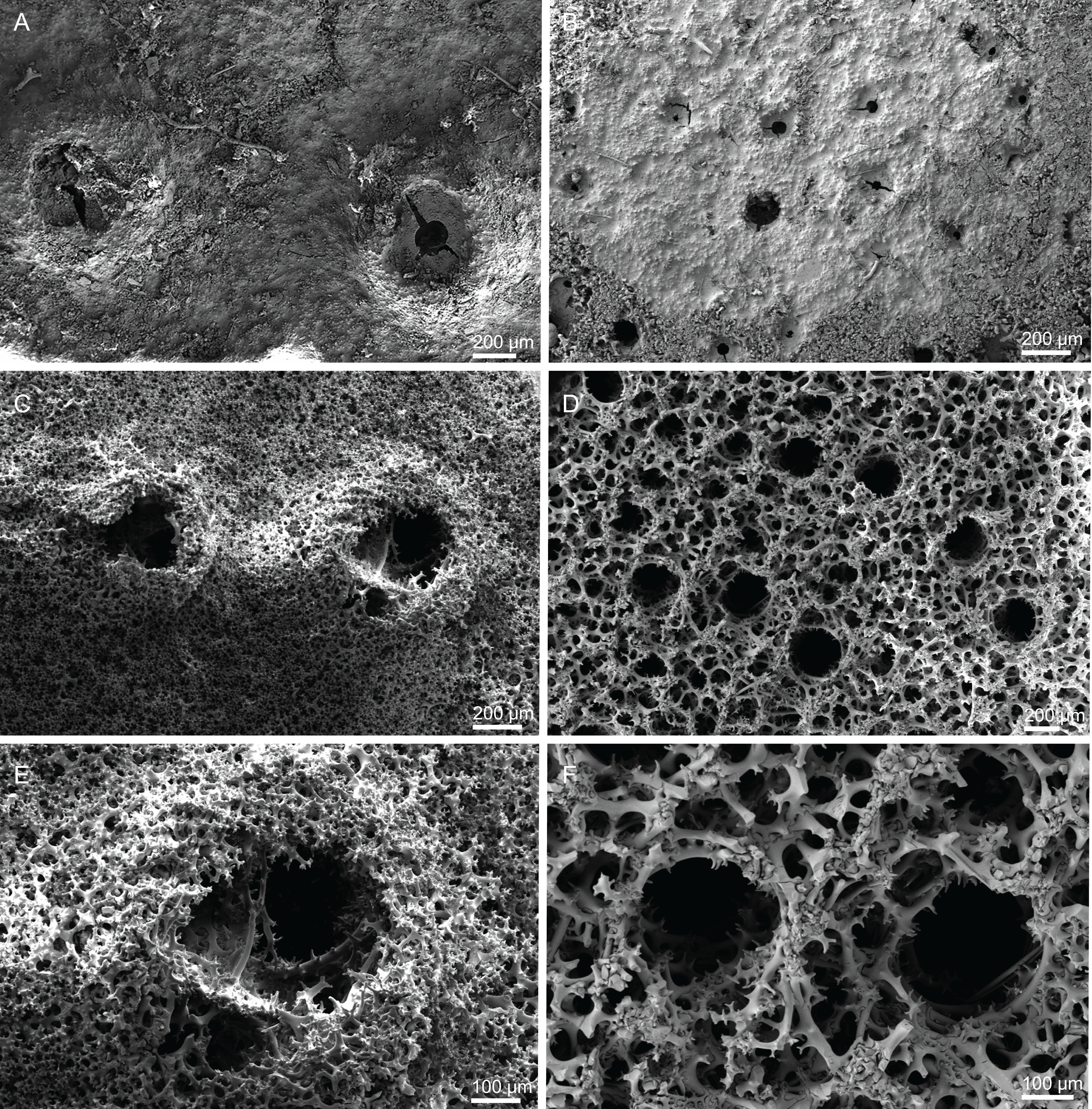{kind=link}
Skeleton. A very intricate, irregular and dense mesh of rhizoclones desmas extremely branched and spiny (Figs. 31A–31C); the body of this sponge is mainly built of desmas, giving them a stony consistency; near the openings and water canals, the arms of the desmas are more elongated; large oxeas cross the skeleton and perforate the surface; no microscleres.

Figure 31: Skeleton of Leiodermatium lynceus Schmidt, 1870, specimen MNHN-IP-2018-93.
(A) Overview of choanosomal desmas, (B) rhizoclone desmas forming a very compact mesh, (C) detail of rhizoclones, (D) ornamentation of rhizoclone desmas, (E) zygosis between several rays, (F) detail of the sculpture of the desma.{kind=link}
Spicules (MNHN-IP-2018-93).
Rhizoclones, very spiny arms with multifurcating spines tips, 156–179–223 µm long and 8.4–19.4–49.9 µm thick (Figs. 31A–31F); zygosis is complex and robust (Fig. 31E).
Oxeas, smooth, straight or curved, up to 1 mm long and 8.5–9.6–10.7 µm thick.
Distribution. These specimens were found on the Gorringe and Hyères Seamounts, between 305 and 480 m depth.
Remarks. Within Tetractinellida, the genus Leiodermatium is particularly difficult from a taxonomic standpoint, given the few characters available to distinguish and describe the different species. In the North Atlantic, only two species have been described to date—L. lynceus Schmidt, 1870 and L. pfeifferae (Carter, 1873); the former from specimens collected off the coast of Portugal, and the later from Madeira island i.e. both from the NEA but unknown depths. Later, Carter (1876) formally explained the differences between these two species: (1) L. lynceus has large oscula located on outer surface while in L. pfeifferae they are on the inner surface; (2) L. pfeifferae has numerous fusiform oxeas on the edge of the laminae, while L. lynceus has “isolated acerates” (Schmidt, 1870) (however they were not found in the redescription of the holotype L. lynceus (Pisera & Lévi, 2002i)). Another important detail, is the difference between the thickness of the laminae on both species, L. lynceus has thinner (3–4 mm) laminae compared to L. pfeifferae (6–17 mm; see Table 5).
In addition to these two currently recognized species, Poritella deciduum (Schmidt, 1879), was also assigned to this genus (Lendenfeld, 1903) but this allocation is considered questionable (Pisera & Lévi, 2002i). Also, Sollas (1888) reported a number of varieties of L. pfeifferae from the material collected in the course of the Challenger expedition in the Atlantic, viz. A. pfeifferae tenuilaminaris (Bahia, Brazil, unknown depth) and A. pfeifferae tenuilaminaris osculis disjunctis (Bermuda, 795–1965 m depth). However, the material was deciduous and therefore the descriptions are incomplete (see also review in Kelly, 2007). Records of L. lynceus and L. pfeifferae for the western Atlantic (e.g. Van Soest & Stentoft, 1988) need to be carefully re-assessed, as they may represent different and likely undescribed species given that several putatively new Leiodermatium species have been reported for the tropical western Atlantic (Schuster et al., 2019) but still lack formal description. Topsent (1892) reports one specimen of Azorica pfeifferae for the Azores (st. 234, 454 m depth) with a strong blue coloration. However, from the illustration provided, it appears that the specimen has elevated openings on the external surface, thereby conforming to L. lynceus.
The specimens analysed in this study are very similar to the holotype of L. lynceus regarding the morphology, surfaces and the ornamentation of the desmas. The only difference lays on the size of the openings: the holotype has large oscula on the outer surface, 500–750 µm in diameter, while in our specimen oscula are 243–269 µm in diameter; the same happens in relation to the pores of the inner surface of the holotype, which are 156–188 µm in diameter, against 68–145 µm in our specimen (Table 5).
Leiodermatium tuba sp. nov.
Figures 3D, 32–33 and Table 5
urn:lsid:zoobank.org:act:041DAB82-B538-4EB9-A43A-1E3E79B67CF8
Holotype. MNHN-IP-2018-72 (1988-09-23, Gorringe Seamount, beam trawl, st. CP11, 36°26.4′N, 11°40.2′W, 805–830 m, Seamount 1 campaign).
Paratype. MNHN-IP-2018-73 (1988-09-23, Gorringe Seamount, beam trawl, st. CP11, 36°26.4′N, 11°40.2′W, 805–830 m, Seamount 1 campaign).
Other material. MNHN-IP-2018-74 (1988-09-25, Gorringe Seamount, epibenthic Warén dredge, st. DW25, 36°49.7′N, 11°03.3′W, 970–1,035 m, Seamount 1 campaign); MNHN-IP-2018-75 (1988-09-23, Gorringe Seamount, beam trawl, st. CP11, 36°26.4′N, 11°40.2′W, 805–830 m, Seamount 1 campaign); MNHN-IP-2018-76 (1988-09-23, Gorringe Seamount, beam trawl, st. CP11, 36°26.4′N, 11°40.2′W, 805–830 m, Seamount 1 campaign); MNHN-IP-2008-235 (1993-01-31, Plato Seamount, epibenthic Warén dredge, st. DW242, 33°12′N, 28°57′W, 710 m, Seamount 2 campaign); MNHN-IP-2008-237 (1993-02-03, Atlantis Seamount, epibenthic Warén dredge, st. DW265, 34°29′N, 30°36′W, 545 m, Seamount 2 campaign); MNHN-IP-2008-249b (1993-01-19, Hyères Seamount, epibenthic Warén dredge, st. DW202, 31°16′N, 28°43′W, 640 m, Seamount 2 campaign); MNHN-IP-2008-253 (1993-01-11, Great Meteor Seamount, epibenthic Warén dredge, st. DW159, 29°44′N, 28°20′W, 330 m, Seamount 2 campaign); MNHN-IP-2008-255 (1993-01-06, Gran Canaria, epibenthic Warén dredge, st. DW130, 28°09′N, 15°53′W, 660 m, Seamount 2 campaign).
Comparative material examined. L. lynceus (MNHN-IP-2018-93, Gorringe Seamount; MNHN-IP-2008-239, Hyères Seamount).
Diagnosis. Massive lamellate vase to contorted walls, sometimes forming a cone, with smooth and similar surfaces.
Description (holotype MNHN-IP-2018-72). Lamellate vase with contorted thin walls, 4–5 mm, occasionally forming a cone (Fig. 3D); this specimen consists of three fragments, the largest one is 138 mm long and 93 mm wide; surfaces are identical when observed with the naked-eye, given they are both smooth, but some differences can be noticed when observed under the stereomicroscope: outer surface has slightly larger depressed openings, 266–322 µm in diameter, (Figs. 32A and 32C) and a striated appearance due to the water canals underneath the surface; inner surface (Figs. 32B and 32D) has a whitish appearance caused by the presence of numerous oxeas covering the smaller depressed openings; openings are 186–261 µm in diameter; specimen coloration varies from light beige to brown in ethanol.

Figure 32: Surface of Leiodermatium tuba sp. nov., holotype MNHN-IP-2018-72.
(A) Outer surface with depressed and slightly larger pores, (B) inner surface, with small and depressed pores, (C) detail of a pore from the outer surface, (D) detail of a pore from the inner surface.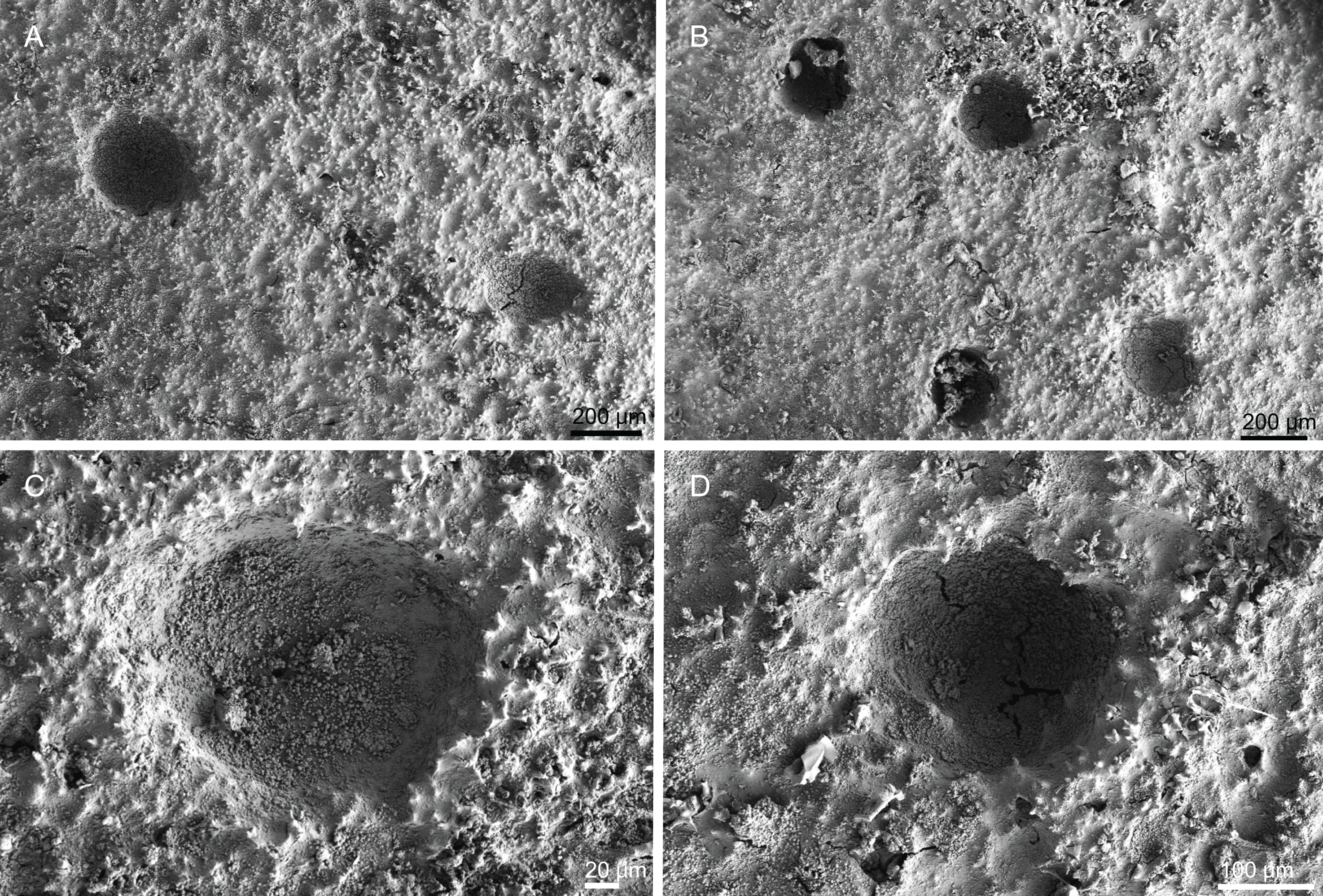{kind=link}
Skeleton. There is no clear distinction between the ectosome and choanosome since there is no special arrangement of spicules or different spicules in the ectosome; choanosomal skeleton is composed by very spiny rhizoclones desmas, forming a complex, branching and compact network (Figs. 33A and 33B); other megascleres are oxeas across the skeleton; microscleres are not present.

Figure 33: Skeleton and spicules of Leiodermatium tuba sp. nov., holotype MNHN-IP-2018-72.
(A) Overview of choanosomal desmas showing the water cannals, (B) rhizoclones desmas, (C) ornamentation of the rays of the desmas, (D) close up on the ornamentation of the desmas, (E) zygosis, (F) zygosis and sculpture of rhizoclones, (G) and (H) oxeas.{kind=link}
Spicules (holotype MNHN-IP-2018-72).
Rhizoclones, spiny, 141–173–211 × 12.1–18.7–31.0 µm in size, with single to multifurcate spiny tips (Figs. 33A–33F); zygoses are strong where several clones can articulate with each other, making the skeleton very dense and robust (Figs. 33E and 33F).
Oxeas, thin, curved, up to 1 mm long and 5.9–8.1–9.8 µm thick (Figs. 33G and 33H).
Etymology. From the Latin tubae= trumpet; since some lamellas in this species have a conical shape resembling a trumpet.
Distribution. The type locality is the Gorringe Seamount at 805–830 m depth. Other specimens were found in Plato, Hyères, Atlantis and Gorringe Seamounts between 545 and 1,035 m, and in Gran Canaria at 660 m.
Remarks. L. tuba sp. nov. exhibits a distinct external morphology and surface ornamentation compared to the other two Leiodermatium species recorded for the North Atlantic, i.e. L. lynceus and L. pfeifferae. Firstly, in L. tuba sp. nov. both surfaces look similar at the naked eye (smooth and with slightly depressed openings; Fig. 32) whereas in L. lynceus and L. pfeifferae, the openings are elevated (depending on the surface) and this is a very distinctive feature (see above remarks under L. lynceus). Additionally, the inner surface of L. tuba sp. nov. is pierced by numerous oxeas providing a whitish colour to this surface. Pisera & Lévi (2002h) discussed the possibility of a new species of Leiodermatium being reported as L. lynceus due to the absence of larger oscules on the outer side. However, it is not clear from their account to which specimens they were referring to nor their characteristics. Perhaps they conform to L. tuba sp. nov. here described.
Another important observation is the bathymetric range where the Leiodermatium spp. were collected in this study. L. tuba sp. nov. was usually found deeper (330–830 m depth) than L. lynceus (305–320 m depth) (see “Diversity” section and Supplemental Material S1).
Family SIPHONIDIIDAE Lendenfeld, 1903
Genus Siphonidium Schmidt, 1879
Synonymy. Siphonidiella Burton, 1928 (junior synonym), Tremaulidium Schmidt, 1879 (junior synonym).
Diagnosis. Siphonidiidae with fistules; choanosmal megascleres are rhizoclones desmas, exotylostyles and/or styles (emended after Pisera & Lévi (2002f)).
Definition. Polymorphic Siphonidiidae, encrusting, massive irregular, hemispherical or irregularly cylindrical to club-shape with fistules; without special ectosomal spicules; rhizoclone desmas, exotylostyles and styles as choanosomal spicules (emended after Pisera & Lévi (2002f)).
Type species. Leiodermatium ramosum Schmidt, 1870 (type by original designation).
Siphonidium elongatus sp. nov.
Figures 3E, 34–35 and Table 6
| Habitus | Size | Rhizoclones | Exotylostyles | Strongyles | Locality | |
|---|---|---|---|---|---|---|
| 1Siphonidium ramosum (Schmidt, 1879) (Holotype MCZ 6321, 6322) | Small, irregular massive to cylindrical, with numerous small fistules | 20–55 mm high, 10 mm wide; fistules are 1–2 mm in diameter | Massive rhizoclones, strongly tuberculated, 180–220 µm in size | With spinose heads: 160–220 µm × 2–3 µm | Not present | Florida (depth unkonw) |
| 2Siphonidium ramosum (Schmidt, 1879) | – | – | – | 800–1,000 µm long, 4–6 µm thick | – | Azores (349–793 m) |
| 3Siphonidium dubium Lévi, 1959 (Holotype) | Massive and hard sponge with a large base, that its subdivided into three lobes 2 cm long barely separated, that ended on a flat surface; surface reticulated and covered by numerous pores; ostia, 1–1.5 cm in diameter; colour light beige when alive and dark brown in ethanol | – | Compact | Not present | Abundant, grouped perpendicular to the surface, tip slightly rugose and rounded, 600–800 µm | Principe, Gulf of Guinea (50 m depth) |
| Siphonidium geminum (Schmidt, 1879) (Holotype MNHN DT 2194) | Flat and irregular incrusting basis bearing simple or bifurcate cone shaped prolongations with round ends; surface is covered by a finely corrugated cuticula | – | Irregular, like “three-roots” and later they are bumpy and hard | – | Not present | Gulf of Mexico (240 m depth) |
|
Siphonidium elongatus sp. nov. (Holotype MNHN-IP-2008-236) |
Cylindrical to arborescent, sometimes bulb shape; surface is smooth and exhibits fistules that are often closed but may be open; colour is beige to brown | 33–49 × 2–9 mm in size | 123–197–267 × 10.4–23.5–40.3 µm; desmas branches, 13.8–30.9–88.2 µm long | 173–363–504 × 2.9–5.1–6.6 µm (n = 4) | Not present | Atlantis seamount (545 m depth) |
|
Siphonidium elongatus sp. nov. (Paratype MNHN-IP-2018-79) |
Cylindrical, elongated with several fistules; colour is brown | 54 × 4–5 mm in size | 129–210–326 × 12.1–22.1–34.0 µm; desmas branches, 16.8–45.4–83.7 µm long | 248–393 × 6.5–12.4 µm (n = 3) | Not present | Gorringe seamount (605–675 m depth) |
urn:lsid:zoobank.org:act:26B193F9-2588-4479-ACB2-27AD1945DEE4
Holotype. MNHN-IP-2008-236 (1993-02-03, Atlantis Seamount, epibenthic Warén dredge, st. DW265, 34°29′N, 30°36′W, 545 m, Seamount 2 campaign).
Paratype. MNHN-IP-2018-79 (1988-09-26, Gorringe Seamount, beam trawl, st. CP28, 36°38′N, 11°29.8′W, 605–675 m, Seamount 1 campaign).
Other material. MNHN-IP-2008-232 (1993-01-06, Gran Canaria, epibenthic Warén dredge, st. DW128, 28°08′N, 15°52′W, 470 m, Seamount 2 campaign), MNHN-IP-2008-245 (1993-01-16, Hyères seamount, epibenthic Warén dredge, st. DW182, 31°23′N, 28°54′W, 480 m, Seamount 2 campaign), MNHN-IP-2008-256 (no data), MNHN-IP-2018-80 (no data), MNHN-IP-2018-81 (1988-09-26, Gorringe Seamount, beam trawl, st. CP28, 36°34.9′N, 11°28.4′W, 605–675 m, Seamount 1 campaign), MNHN-IP-2018-78 (1988-09-24, Gorringe seamount, epibenthic Warén dredge, st. DW21, 36°34.9′N, 11°28.4′W, 460–480 m, Seamount 1 campaign), MNHN-IP-2018-82 (1988-10-08, Lion seamount, epibenthic Warén dredge, st. DW63, 35°1.4′N, 15°34.4′W, 630 m, Seamount 1 campaign), MNHN-IP-2018-83 (1988-09-26, Gorringe seamount, epibenthic Warén dredge, st. CP28, 36°38.0′N, 11°29.8′W, 605–675 m, Seamount 1 campaign).
Diagnosis. Polymorphic sponge, cylindrical to arborescent, with several fistules; rhizoclones with slim arms ornamented with microspines along the edges; exotylostyles to styles as other choanosomal megascleres.
Description (holotype MNHN-IP-2008-236). Polymorphic sponge, cylindrical to arborescent Siphonidiidae, sometimes of bulb shape, attached by the base to the substrate; small, 33–49 mm high, thin, 2–9 mm wide (but can be 14 mm wide); surface is smooth and exhibits fistules spread through the sponge pointed in several directions, 1–8 mm long and 1–4 thick (Fig. 3E); fistules are usually close-ended, but when open, it is possible to see the subdermal water canals emerging from the interior of the sponge; extremely hard sponge (stony consistency); colour varies from beige to brown in ethanol.
Skeleton. No clear distinction of the spicules between the ectosome and choanosome, with exception of the desmas of the surface that are different from the interior of the skeleton: a layer of flattened, fused and modified desmas, resembling a puzzle, constitutes the surface of the sponge (Figs. 34B and 35B); these modified desmas, resembling a shield, contribute to the hardness of this species; some wrinkles can also be observed on the surface of the sponge (Fig. 35A); choanosome is formed by an extremely dense, compact and irregular net of rhizoclone desmas, exotylostyles and rarely styles, crossing through the skeleton; several water canals can be observed in a cross section of the sponge, as large holes (Figs. 34A and 34C) surrounded by the desmas that here are slightly more elongated (Fig. 34C); desmas from the fistules are different from the ones in the ‘body’ of the sponge, i.e., usually the desmas of the fistules are longer and looser (Fig. 35C) while in the ‘body’ they are very dense and compact (Fig. 34C).
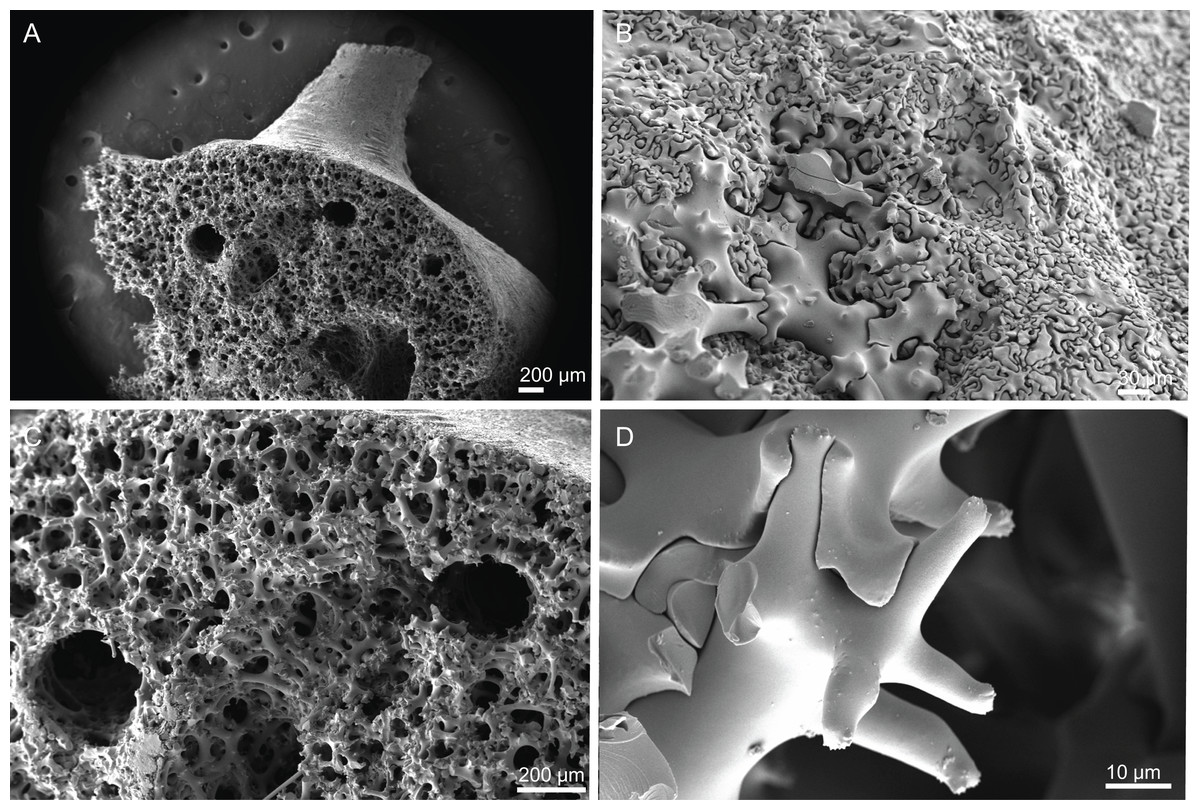
Figure 34: Surface and skeleton of Siphonidium elongatus sp. nov., holotype MNHN-IP-2008-236.
(A) Overview of a fragment of the specimen showing a fistule, (B) surface of the sponge composed of modified desmas, (C) overview of the chonosomal desmas showing the water canals in the rhizoclones, (D) zygosis showing the ornamentation of desmas tips.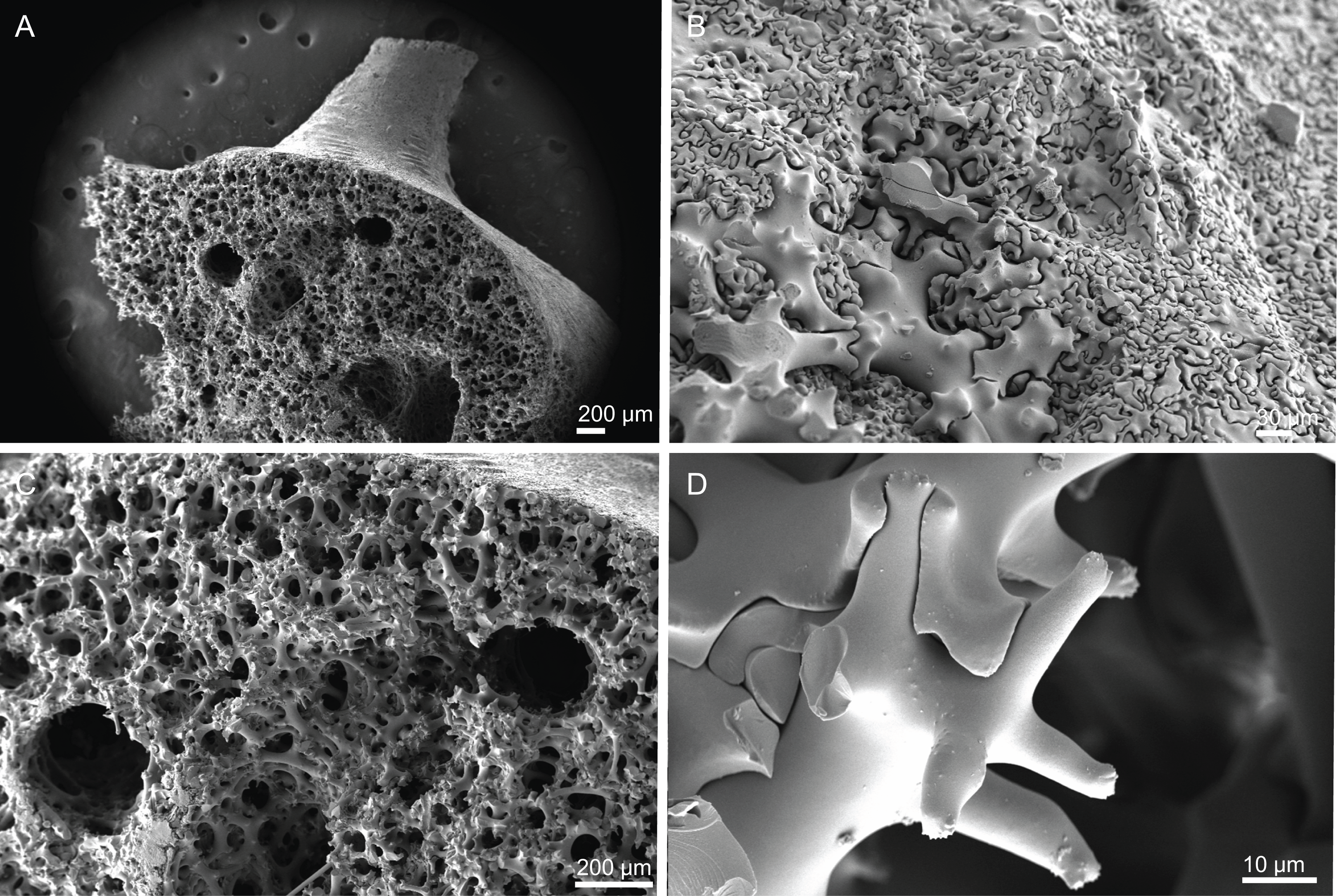{kind=link}
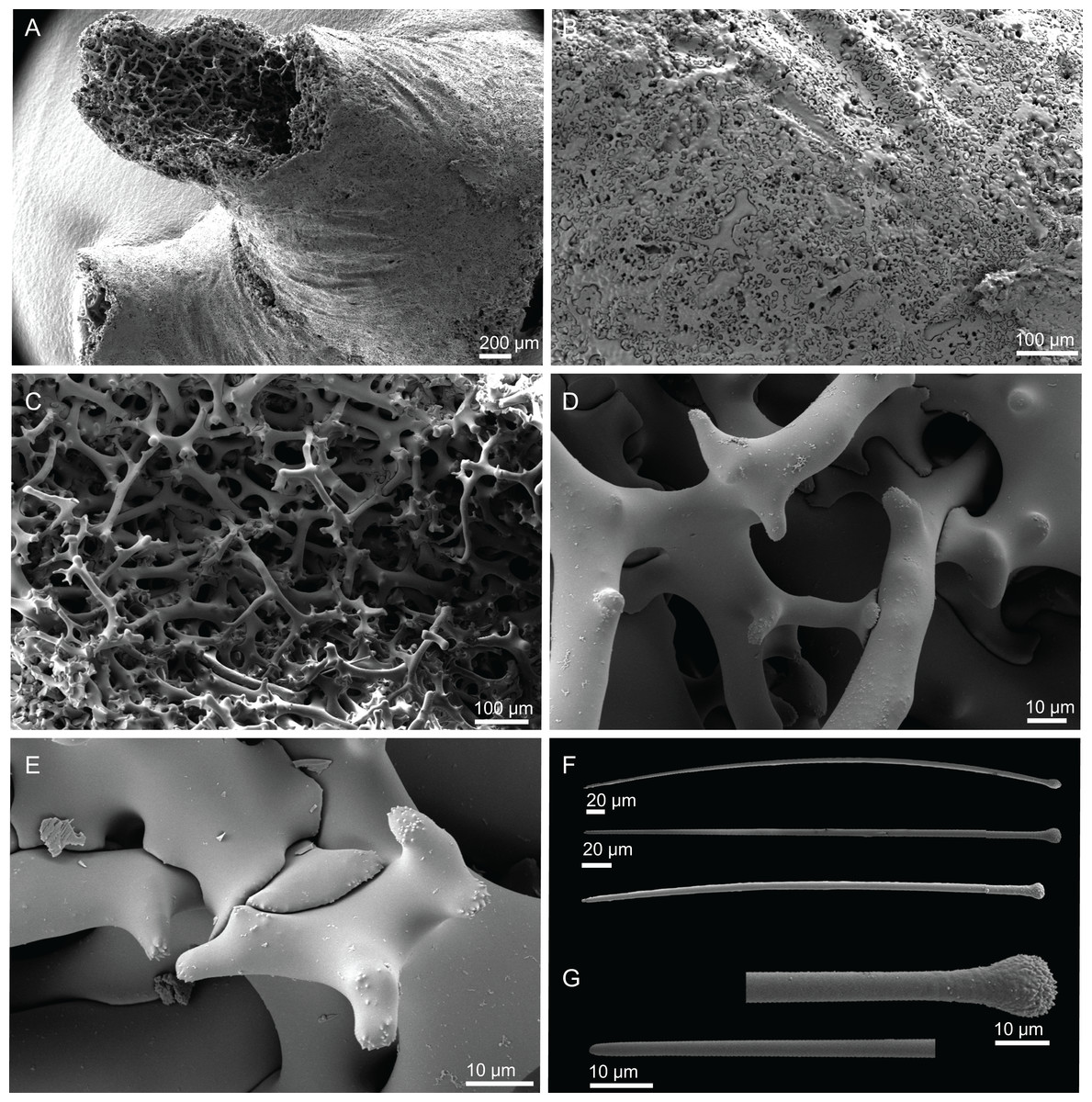
Figure 35: Surface, skeleton and spicules of Siphonidium elongatus sp. nov., holotype MNHN-IP-2008-236.
(A) Overview of a fistule, (B) close up of the modified desmas from the surface of the fistule, (C) loose rhizoclones of the fistule, (D) detail of the desmas, (E) close up of the ornamentation of the desmas of the fistules, (F) exotylostyles, (G) detail of the spiky pin-shaped head and tip of the exotylostyles.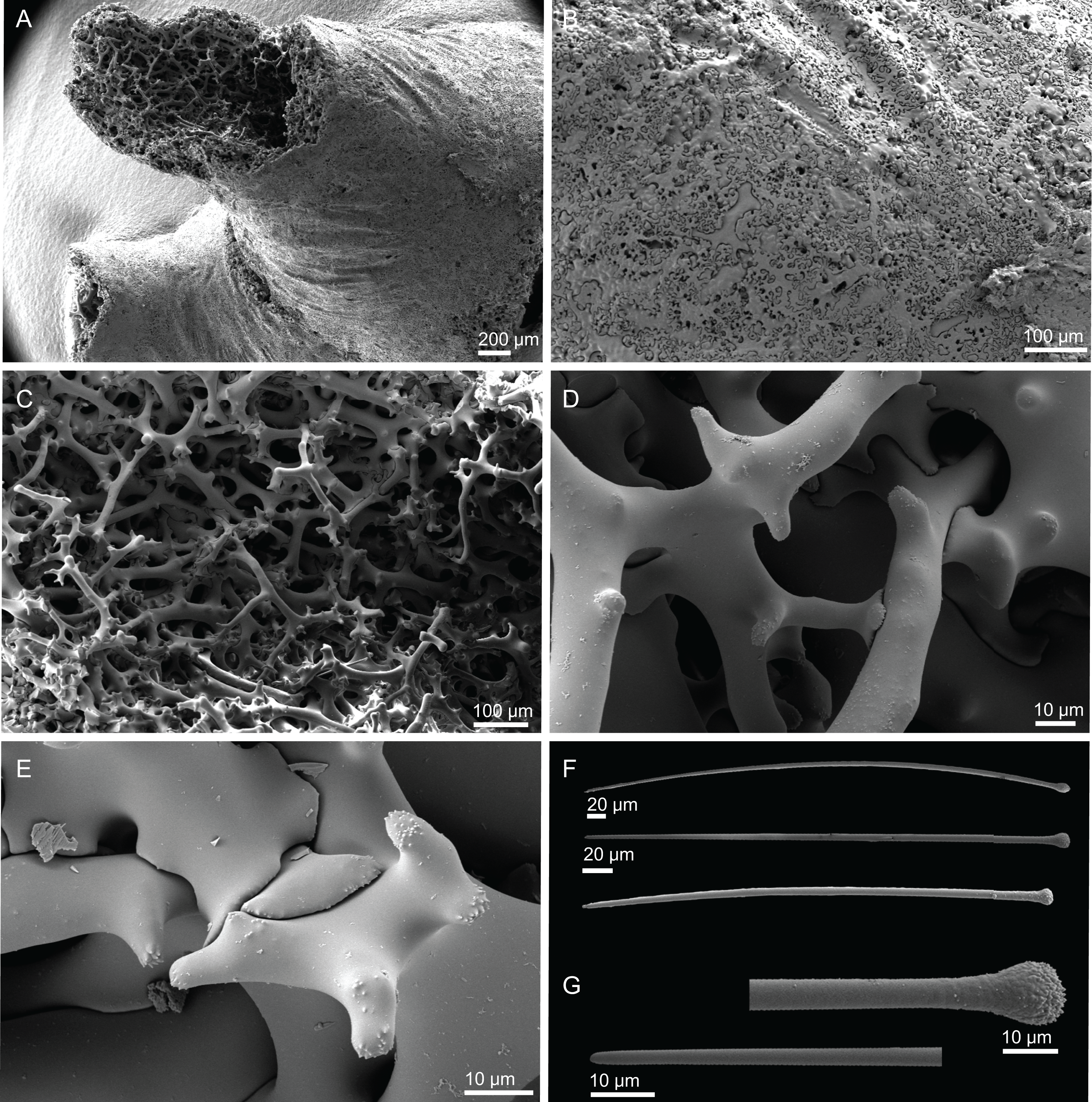{kind=link}
Spicules (holotype MNHN-IP-2008-236).
Rhizoclone desmas, extremely dense especially near the surface, 123–197–267 μm long and 10.4–23.5–40.3 μm wide (Figs. 34A–34C); clones are smooth with several finger-like branches, that can be smooth or ornamented with microspines on the tips, 13.8–30.9–88.2 μm long (Figs. 34D, 35D and 35E); zygoses can be formed by several rays or just some, but it is always solid and complex (Figs. 34F and 35E);
Exotylostyles, pin-shaped, with spiny heads and pointed tips, straight or slightly curved, not very abundant, 173–363–504 μm in length and 2.9–5.1–6.6 μm in width (Figs. 35F and 35G); some exotylostyles look underdeveloped and resemble styles.
Distribution. Siphonidium elongatus sp. nov. was found in the Atlantis, Hyéres, Lion, and Gorringe seamounts, and in Gran Canaria, between 470 and 675 m depth
Etymology. From the latin elongatus = elongated, due to an elongated shape of the desmas, especially those composing the fistules.
Remarks. Three species of Siphonidium have been described in the Atlantic Ocean, and only one, S. ramosum, has been reported for both sides of the North Atlantic (Schmidt, 1879; Topsent, 1928, 1904, 1892; Van Soest, 2017; Van Soest & Stentoft, 1988) and Mediterranean Sea (Longo, Mastrototaro & Corriero, 2005; Vacelet, 1969; Zibrowius & Taviani, 2005). With the redescription of S. ramosum in (Pisera & Lévi, 2002g), a detailed account of the external morphology and spicules was given, allowing a better definition of the species. Despite the relatively similar habitus of S. ramosum and S. elongatus sp. nov., the main difference between these two species relies on the desmas morphology and ornamentation: S. elongatus sp. nov. has very spiny rhizoclones with slim arms ornamented with microspines on the edges, contrasting with the tuberculated rhizoclones of S. ramosum. Another distinct feature, is the presence of styles in S. elongatus sp. nov. (even though they are rare) that were never mentioned in the redescription of S. ramosum. Furthermore, when S. elongatus sp. nov. is compared with the other North Atlantic species, its external morphology and spicules differ: S. dubium Lévi, 1959 is a massive sponge with a large base, subdivided into three lobes and the only one within the genus with strongyles; S. geminum (Schmidt, 1879) has a flat and irregular incrusting base with simple or bifurcated cone shape.
Topsent (1904) presented a small description of S. ramosum from several specimens found in the Azores. In his account, the shape and the ornamentation of the desmas are not explicitly described or illustrated, but the spicules sizes are given and are much larger than the ones described by Schmidt (1879) from material collected in the Gulf of Mexico (Table 6). The spicules sizes in S. elongatus sp. nov. are more similar to the ones in S. ramosum described by Schmidt than to the one described by Topsent. It was previously stated by Van Soest (2017), that the S. ramosum reported from the Azores, is most likely a different species due to the difference in the spicules sizes when compared to the type material. A revision of Topsent’s material would be required to clarify this question.
Order BUBARIDA Morrow & Cárdenas, 2015
Family DESMANTHIDAE Topsent, 1893
Genus Petromica Topsent, 1898
Synonymy. Monanthus Kirkpatrick, 1903 (junior synonym).
Diagnosis. Massive, encrusting or globular shape Desmanthidae with desmas branching in various planes forming a loosely articulated or non-articulated choanosomal skeletal structure. (List-Armitage & Hooper, 2002; Pisera & Lévi, 2002h).
Definition. Massive, encrusting or globular in shape, with or without fistule-like papillae. Surface smooth, hispid, conules can be present. Compressible to rigid, or soft to fragile sponges. Acrepid or monocrepid smooth desmas, branched in several planes. Desmas can be isolated, non-articulated, fused, or dispersed in the ectosome and choanosome; zygomes vary from simple to complex; zygosis when present, is rarely fully articulated in the skeleton turning into a loose skeleton. Other megascleres are oxeas, where the tips can vary from sharp to blunt. Microscleres not present (List-Armitage & Hooper, 2002; Muricy et al., 2001; Pisera & Lévi, 2002h)
Type species. Petromica (Petromica) grimaldii Topsent, 1898 (type by monotype).
Subgenus Petromica Topsent, 1898
Diagnosis. Firm and rigid sponge, with or without papillae, with acrepid or monocrepid desmas that can form a loose or well-formed skeleton. Oxeas present and variable in size (List-Armitage & Hooper, 2002).
Petromica (Petromica) grimaldii Topsent, 1898
Figures 3F, 36–37 and Table 7
| Habitus | Size | Monocrepid desmas | Anisoxeas | Strongyloxeas | Locality | |
|---|---|---|---|---|---|---|
| 1Petromica (Chaladesma) ciocalyptoides (Van Soest & Zea, 1986) (Holotype RMNH 1309) | Basal mass buried in the sand, with proeminent tapering fistules; colour pale yellow-orange alive and white in alcohol | 1 cm thick; fistules are large, 100 mm long, 3–9 mm thick | Smooth, long clads, 600–700 µm, clads up to 300 µm, epirhabd 180–300 × 10–28 µm; no zygosis | – | Sometimes modified to styles, 378–592 × 5–22 µm | Saba Bank, Colombia (34 m depth) |
| 2Petromica (Chaladesma) citrina Muricy, Hajdu, Minervino, Madeira & Peixinho, 2001 (Holotype MNRJ 580) | Thickly encrusting to massive, irregular, with small cone-shaped or digitiorm surface projections and large papillae (1–9); colour alive bright orange-yellow, pale yellow in ethanol | Base: 30–90 × 16–60 mm wide, 4–20 mm thick | 180–337–620 µm, epirhab 40–87.4–190 × 9.8–32.6 µm; cladii 50–126–300 µm. long | – | Usually both extremities are acerate, but occasionally stylote or strongylote can be present, 320–527–780 × 3.2–26 µm | São Sebastião island, Brazil (25 m depth) |
|
3Petromica (Petromica) grimaldii Topsent, 1898 (Holotype MNHN DT 850) |
Massive sponge, large in the base and slimer on top, covered with conules | Very variable, large specimens up to 40 mm high and 30 mm diameter | Up to 570 µm in size | 956–1250 × 23–30 µm | 700–1280 × 23 µm | Azores (200–599 m depth) |
| Petromica (Petromica) grimaldii (MNHN-IP-2008-92) | Massive sponge with a soft and rugose surface; papillae absent | Small, 21 mm height and 14 mm width | 347–499–652 µm × 8–22–80 µm | 890–1,213–1,376 × 14.5–22.5–28.9 µm | 541–1,122–1,561 × 13.1–21.1–33.2 µm (n = 29) | Gorringe Seamount (255–265 m depth) |
Synonym. Petromica grimaldii Topsent, 1898 accepted, alternate representation (subgenus assignment).
Material. MNHN-IP-2018-92 (1998-09-24, Gorringe Seamount, epibenthic Warén dredge, st. DW16, 36°31.1′N, 11°32.5′W, 255–265 m, Seamount 1 campaign).
Diagnosis. Small sponge with a conulose surface and no papillae; desmas are monocrepid.
Description (MNHN-IP-2018-92). Fragile, soft, massive sponge with a soft and conulose surface; small, 21 mm height and 14 mm width; colour white in ethanol (Fig. 3F); specimen in poor condition.
Skeleton. No clear distinction between ectosome and choanosome; skeleton is composed of smooth monocrepid desmas (Figs. 36A–36D), poorly articulated (Fig. 36E), forming a loose, confuse and irregular skeleton; other megascleres are anisoxeas and strongyloxeas, rarely tylostyles, usually arranged in bundles (Fig. 36B); it is also possible to observe a zygosis between the desmas and the oxeas (Fig. 36F); microscleres are absent.
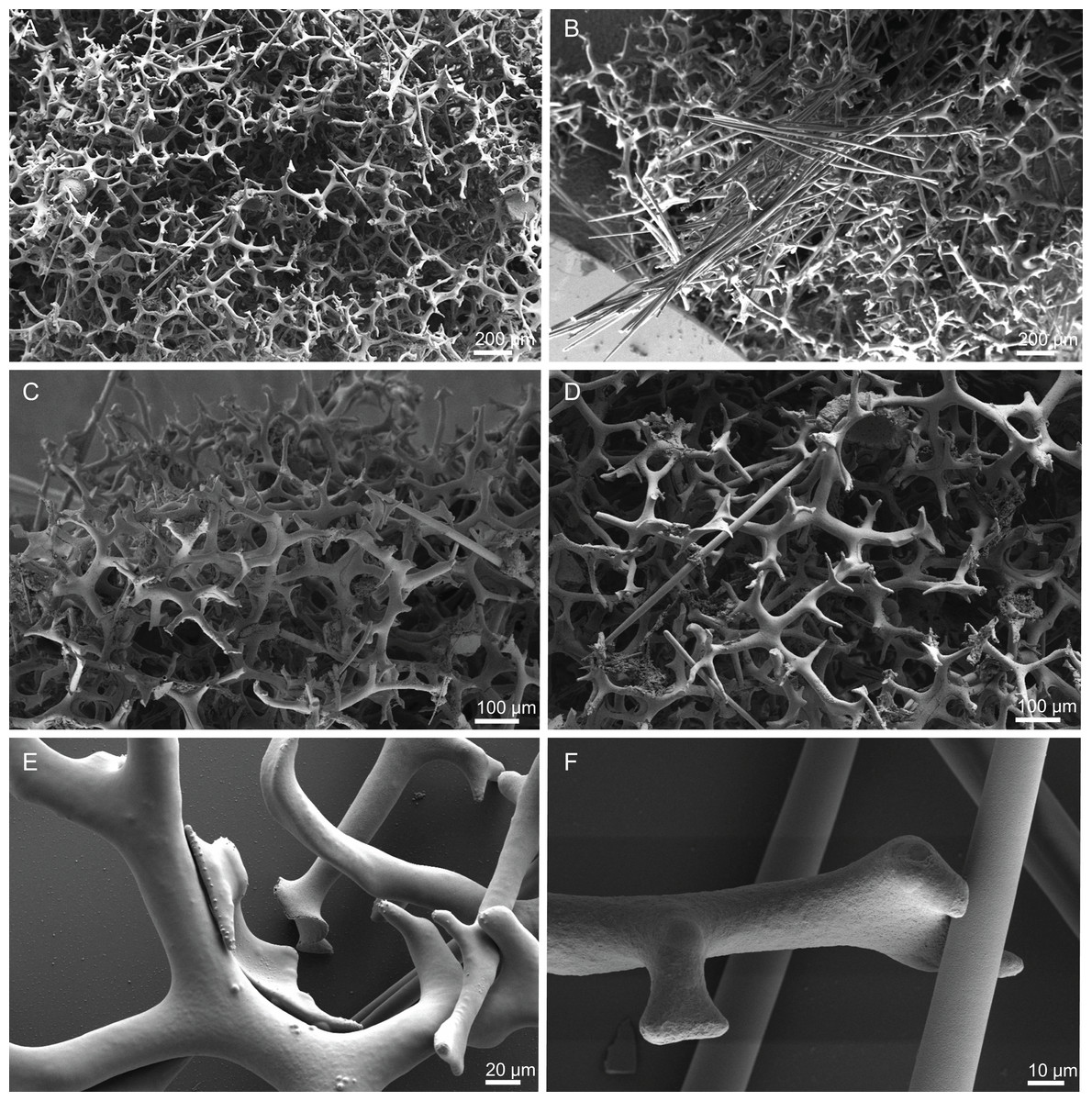
Figure 36: Skeleton of Petromica (Petromica) grimaldii Topsent, 1898, specimen MNHN-IP-2008-92.
(A) Overview of monocrepid desmas, (B) bundles of anisoxeas and strongyloxeas, (C) monocrepid desmas, (D) detail of anisoxeas and strongyloxeas crossing the desmas, (E) zygosis, (F) zygosis between a desma and an oxea.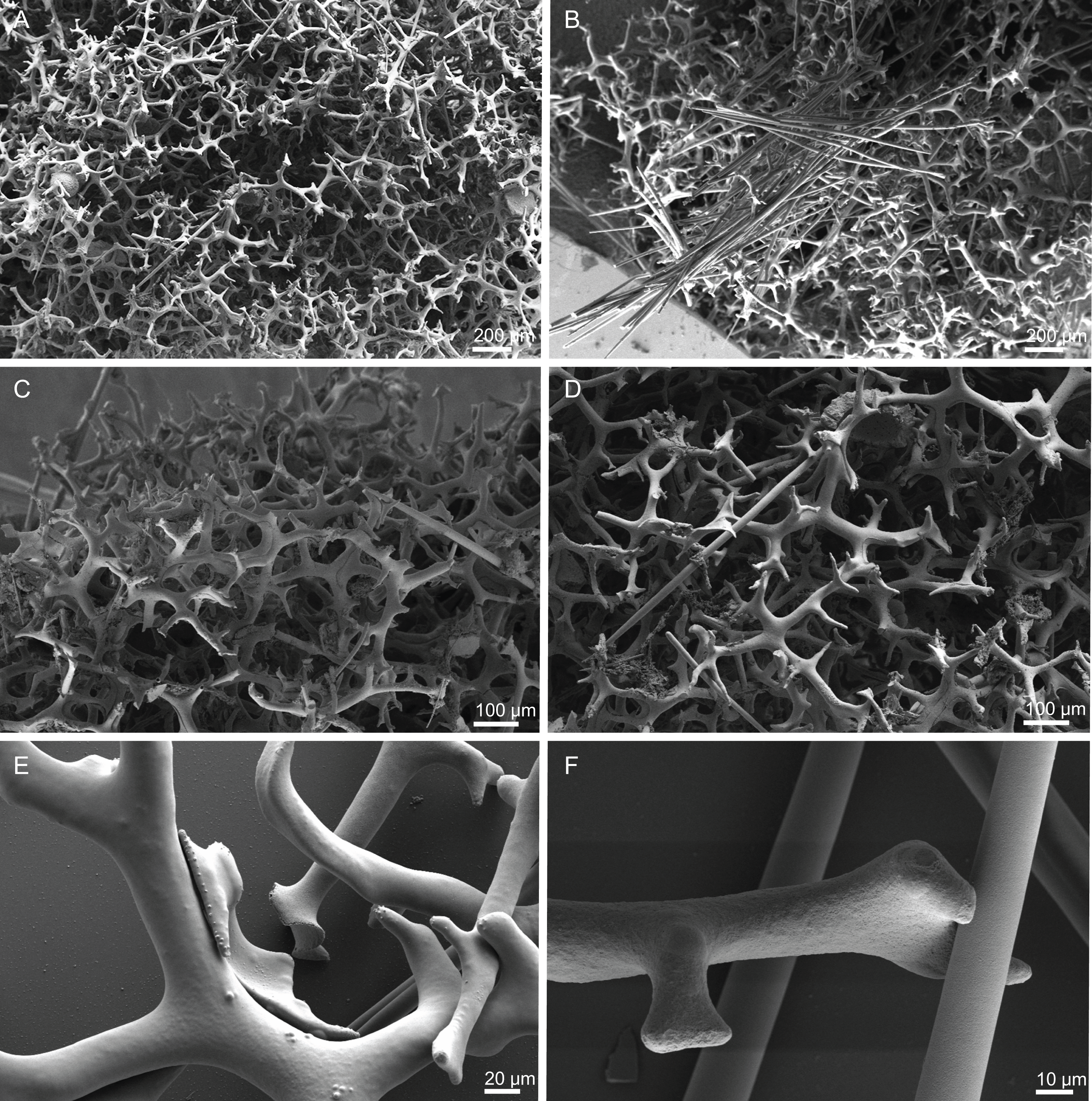{kind=link}
Spicules (MNHN-IP-2018-92).
Monocrepid desmas, smooth, with branches in several planes, except for the tips which can have some ornamentation with a spiny appearance (Figs. 36 and 37A), 347–499–652 × 8–22–80 µm in size; tips are 45–82–147 × 10.6–15.2–30.8 µm in size; zygomes are spiny, mainly in the inner part, about 57–118–207 µm in size (Fig. 37B);
Anisoxeas, very abundant, fusiform, smooth, with acerate tips, 890–1,213–1,376 µm long and 14.5–22.5–28.9 µm thick (Fig. 37C);
Strongyloxeas, smooth, can be straight or curved, with one acerate tip and one blunt tip, 541–1,122–1561 µm long and 13.1–21.1–33.2 µm wide (Figs. 37D and 37E).
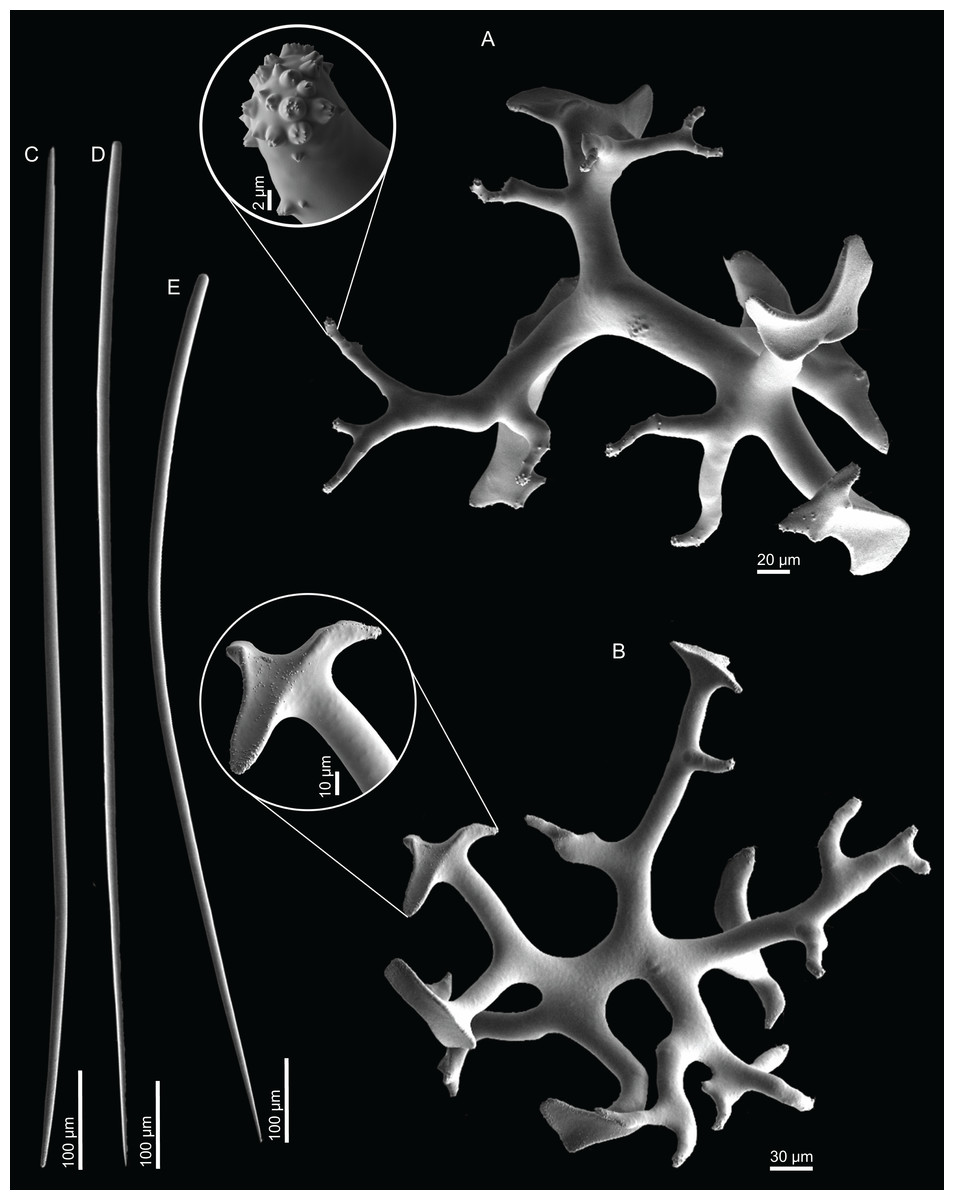
Figure 37: Spicules of Petromica (Petromica) grimaldii Topsent, 1898, specimen MNHN-IP-2008-92.
(A) Monocrepid desma pointing a detail on the ornamentation of the tips of the desma, (B) monocrepid desma showing a close up of the zygome, (C) Anisoxea, (D) and (E) strongyloxeas.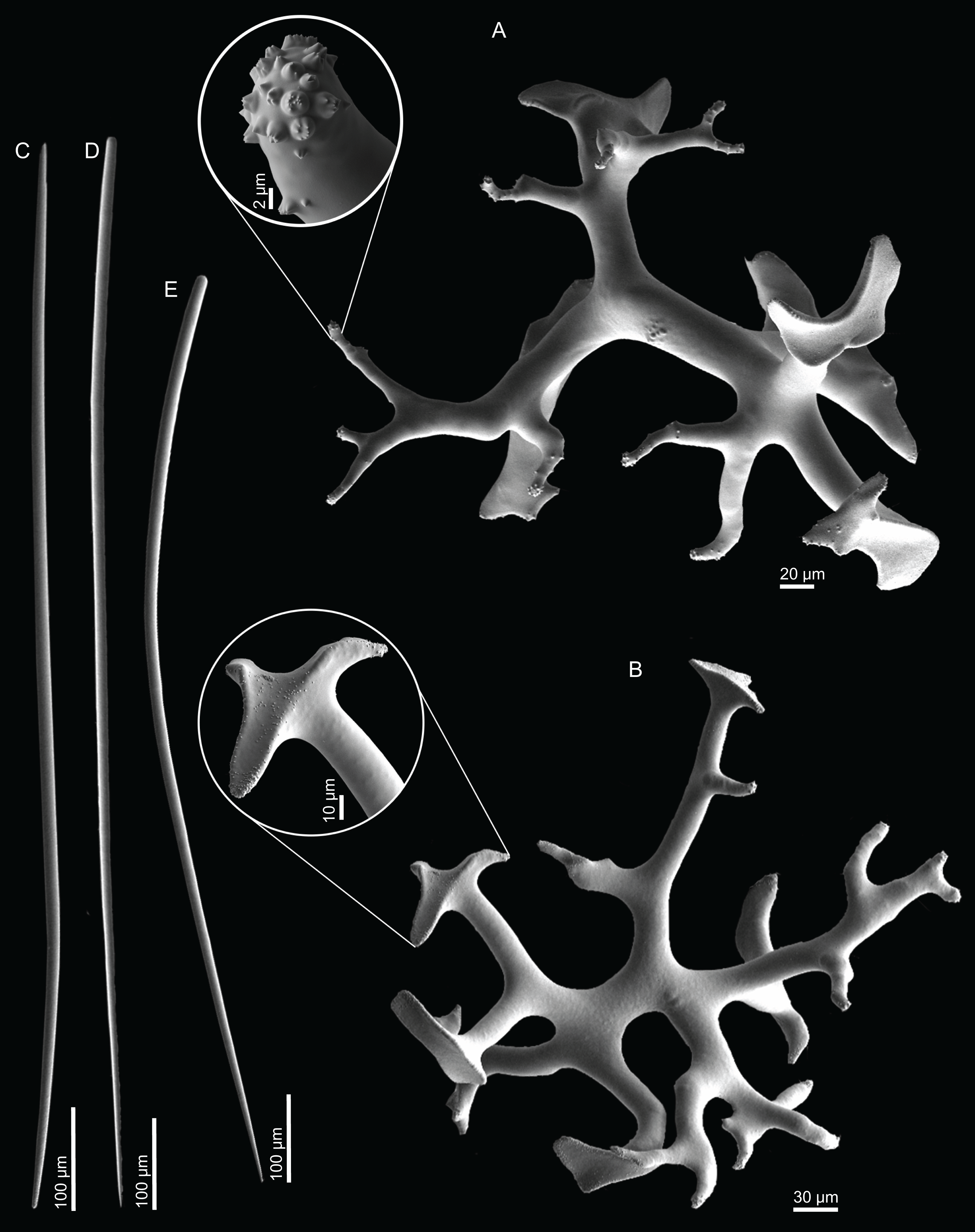{kind=link}
Distribution. This specimen was found on the Gorringe seamount between 255 and 265 m depth.
Remarks. Petromica is a widely distributed genus, and so far, eight species have been described. In the North Atlantic, three species have been reported, P. (Chaladesma) ciocalyptoides and P. (Chaladesma) citrina to the NWA and P. (Petromica) grimaldii from the NEA and MED (Table 7). P. (P.) grimaldii was first described from the Azores archipelago by Topsent (1898) where it was found to be a very common sponge, collected throughout the archipelago between 200 and 914 m depth (Topsent, 1928, 1904, 1898). This species has been also reported from the MED (Boury-Esnault, Pansini & Uriz, 1994; Pulitzer-Finali, 1972) and since microspine desmas’ terminations were absent, P. (P.) massalis Dendy, 1905 (a species from the Indian Ocean) and P. (P.) grimaldii were synonymized (Pulitzer-Finali, 1972). According to Muricy et al. (2001), these microspines are not present in all desmas in the same specimen and they can be rare. Therefore, the absence of microspines in the desmas is not enough to distinguish one species from another. A more detailed examination of the specimens from the MED would be necessary to allow to clarify this uncertainty (Muricy et al., 2001) and make sure the Petromica found in MED are in fact P. (P.) grimaldii. In the specimen examined in this study spicules sizes are very similar to those of the holotype (from the Azores) and the microspines in the termination of the desmas are present and very evident (Fig. 37).
Diversity
The specimens described in the present work constitute the first records of lithistid demosponges for these two groups of NEA seamounts, except for Exsuperantia archipelagus. The Meteor seamount group harbours a more diverse lithistid fauna, 15 species, compared to the Lusitanian seamount group, where six species are recorded (Table 8). At a smaller scale, the Hyères seamount is the most diverse where eight species, namely N. pomponiae sp. nov., M. cf. azorica, M. robusta., E. archipelagus, E. levii sp. nov., L. lynceus, L. tuba sp. nov. and S. elongatus sp. nov. were found, followed by the Gorringe and Atlantis (six species), Plato and Great Meteor (five species), Tyro (three species) and Lion seamount (one species). Two specimens were found on the Antialtair and Ampère seamount (one on each) but it was not possible to identify them down to species level because they were small and incrusting specimens, possibly young individuals of M. robusta. The majority of the species have a restricted distribution and are found only in one or two seamounts, except E. archipelagus, M. cf. azorica, M. schusterae sp. nov., L. tuba sp. nov. and S. elongatus sp. nov., that are distributed between three to five different seamounts. Four species were also sampled in Gran Canaria, M. cf. azorica (480 m depth), E. archipelagus (660 m depth), L. tuba sp. nov. (660 m depth) and S. elongatus sp. nov. (470 m depth), the two latter representing the first records for the Canary Islands. New bathymetric records were also reported for three species, viz. M. robusta (705 m), D. ramifera (300–420 m), D. verrucosa (338–520 m) (Fig. 38).
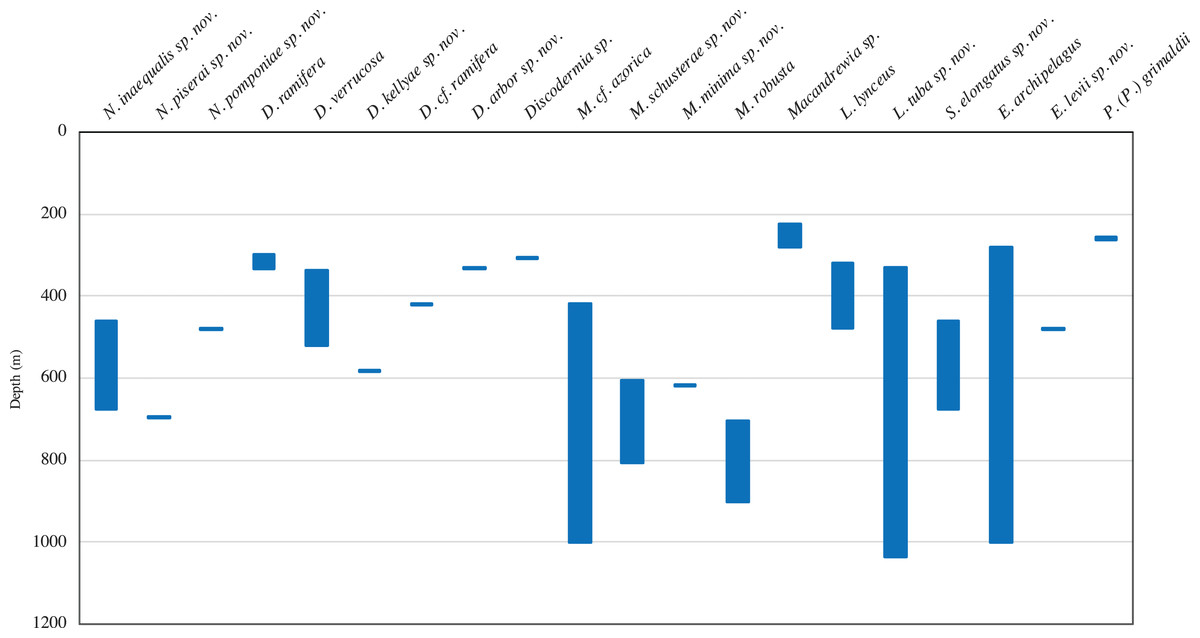
Figure 38: Bathymetric distribution of the lithistid demosponges collected during the Seamount 1 and Seamount 2 expeditions on the Northeast Atlantic seamounts.
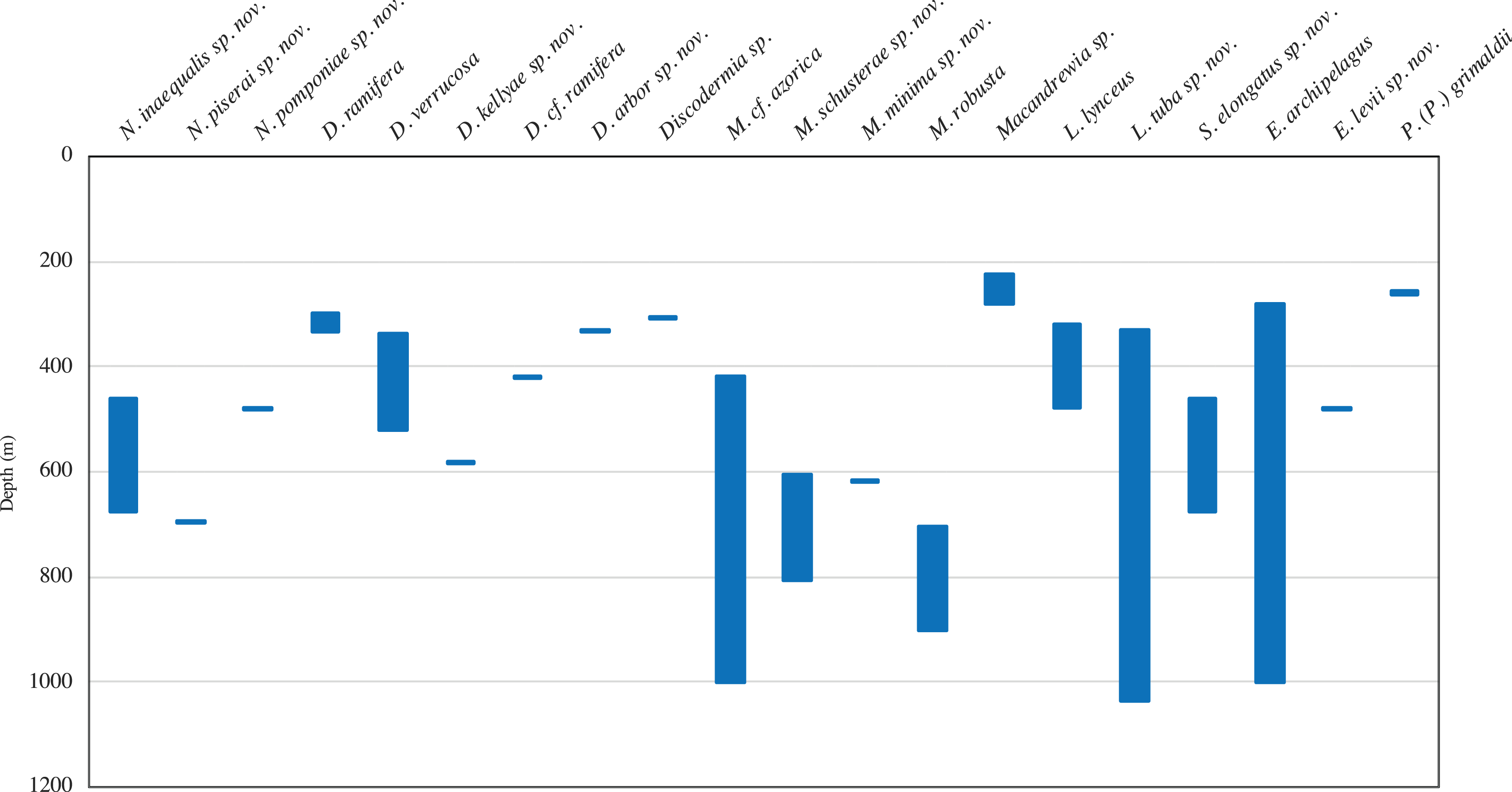{kind=link}
| Seamounts | Oceanic archipelagos | Continental shelf/slope | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | ATR | ATL | TYR | PLT | HYR | MET | LIO | GOR | AMP | AZO | MAD | SEL | CAN | PT | MED | MOR |
| CORALLISTIDAE Sollas, 1888 | ||||||||||||||||
| Corallistes elegantior Schmidt, 1870 | ○ | |||||||||||||||
| Corallistes masoni (Bowerbank, 1869) | ○ | ○ | ||||||||||||||
| Isabella harborbranchi Carvalho, Pomponi & Xavier, 2015 | ○ | |||||||||||||||
| Neophrissospongia endoumensis Pisera & Vacelet, 2011 | ○ | |||||||||||||||
| Neophrissospongia nana Manconi & Serusi, 2008 | ○ | |||||||||||||||
| Neophrissospongia nolitangere (Schmidt, 1870) | ○ | ○ | ○ | ○ | ○ | |||||||||||
| Neophrissospongia radjae Pisera & Vacelet, 2011 | ○ | |||||||||||||||
| Neoschrammeniella bowerbankii (Johnson, 1863) | ○ | ○ | ||||||||||||||
| Neoschrammeniella inaequalis sp. nov. | • | |||||||||||||||
| Neoschrammeniella piserai sp. nov. | • | |||||||||||||||
| Neoschrammeniella pomponiae sp. nov. | • | |||||||||||||||
| Neoschrammeniella sp. | • | |||||||||||||||
| THEONELLIDAE Lendenfeld, 1903 | ||||||||||||||||
| Discodermia arbor sp. nov. | • | |||||||||||||||
| Discodermia kellyae sp. nov. | • | |||||||||||||||
| Discodermia polydiscus (Bowerbank, 1869) | ○ | ○ | ||||||||||||||
| Discodermia polymorpha Pisera & Vacelet, 2011 | ○ | |||||||||||||||
| Discodermia ramifera Topsent, 1892 | • | ○ | ||||||||||||||
| Discodermia cf. ramifera | • | |||||||||||||||
| Discodermia sp. | • | |||||||||||||||
| Discodermia verrucosa Topsent, 1928 | • | • | ○ | ○ | ○ | |||||||||||
| Theonella annulata Lendenfeld, 1907 | ○ | |||||||||||||||
| MACANDREWIIDAE Schrammen, 1924 | ||||||||||||||||
| Macandrewia azorica Gray, 1859 | ○ | ○ | ○ | |||||||||||||
| Macandrewia cf. azorica | • | • | • | • | ||||||||||||
| Macandrewia schusterae sp. nov. | • | • | • | |||||||||||||
| Macandrewia minima sp. nov. | • | |||||||||||||||
| Macandrewia ramosa Topsent, 1904 | ○ | |||||||||||||||
| Macandrewia robusta Topsent, 1904 | • | ○ | ||||||||||||||
| Macandrewia sp. | • | • | ||||||||||||||
| PHYMARAPHINIIDAE Schrammen, 1924 | ||||||||||||||||
| Exsuperantia archipelagus Carvalho and Pisera, 2018 | • | • | • | • | ○ | ○ | • | |||||||||
| Exsuperantia levii sp. nov. | • | |||||||||||||||
| AZORICIDAE Sollas, 1888 | ||||||||||||||||
| Leiodermatium lynceus Schmidt, 1870 | • | • | ○ | ○ | ○ | ○* | ||||||||||
| Leiodermatium pfeifferae (Carter, 1876) | ○ | ○ | ○ | |||||||||||||
| Leiodermatium tuba sp. nov. | • | • | • | • | • | • | ||||||||||
| SCLERITODERMIDAE Sollas, 1888 | ||||||||||||||||
| Aciculites mediterranea Manconi, Serusi & Pisera, 2006 | ○ | |||||||||||||||
| Microscleroderma lamina Perez et al., 2004 | ○ | |||||||||||||||
| SIPHONIDIIDAE Lendenfeld, 1903 | ||||||||||||||||
| Gastrophanella phoeniciensis Perez et al., 2004 | ○ | |||||||||||||||
| Siphonidium elongatus sp. nov. | • | • | • | • | • | |||||||||||
| Siphonidium ramosum (Schmidt, 1870) | ? | ○ | ||||||||||||||
| DESMANTHIDAE Topsent, 1893 | ||||||||||||||||
| Desmanthus incrustans (Topsent, 1889) | ○ | |||||||||||||||
| Petromica (Petromica) grimaldii Topsent, 1898 | • | ○ | ○ | |||||||||||||
| Sulcastrella tenens (Vacelet, 1969) | ○ | |||||||||||||||
| Total of species/taxa 36 | 1 | 6 | 3 | 5 | 8 | 5 | 1 | 6 | 1 | 11 | 7 | 2 | 10 | 2 | 15 | 1 |
Notes:
AMP, Ampere seamount; ATL, Atlantis seamount; ATR, Antialtair seamount; AZO, Azores; CAN, Canaries; GOR, Gorringe seamount; HYR, Hyères seamount; LIO, Lion; MAD, Madeira; MED, Mediterranean Sea; MET, Great Meteor seamount; MOR, Morocco; PLT, Plato seamount; PT, Portugal; SEL, Selvagens; TYR, Tyro seamount.
Sources of the literature records: Bowerbank (1869); Carter (1876); Carvalho, Pomponi & Xavier (2015); Carvalho & Pisera (2019); Cruz (2002); du Bocage (1869); Gray (1859); Johnson (1863); Lendenfeld (1907); Longo, Mastrototaro & Corriero (2005); Magnino et al. (1999); Maldonado et al. (2015); Manconi, Serusi & Pisera (2006); Manconi & Serusi (2008); Perez et al. (2004); Pisera & Vacelet (2011); Pulitzer-Finali (1972); Schmidt (1870); Topsent (1889, 1892, 1898, 1904, 1928); Vacelet (1969).
?The assignment of the specimens examined by Topsent in Azores need to be revised in order to clarify if it is in fact S. ramosum.
Some of the examined material was of very small size and/or in poor condition, which hampered its identification to lower taxonomic levels. These specimens were therefore not identified and are not included in this manuscript (see Supplemental Material).
Discussion
Diversity and biogeographic patterns
With the present work, we describe for the first time the lithistid fauna of two seamount groups of the NEA, the Great Meteor and the Lusitanian seamounts. All of the 17 species here reported constitute new records for these seamounts and ten are new to science. The only exception is E. archipelagus previously reported for the Great Meteor Seamount as Exsuperantia sp. (Cárdenas et al., 2011). These 10 newly described species add to the 17 species previously reported for the NEA, representing an increase of approximately 60% of the lithistid diversity of this area. These findings show how understudied the fauna of these ecosystems is and suggests that additional species are likely to be found as survey efforts increase. It also concurs with previous studies made for other invertebrate groups based on material collected from the same seamounts where several new species were described (Berning, Harmelin & Bader, 2017; Cárdenas et al., 2018; George & Schminke, 2002; Gofas, 2007; Souto, Berning & Ostrovsky, 2016). The Great Meteor group, appears to harbour a more diverse lithistid fauna, with a total of 15 species (nine new to science), whereas in the Lusitanian group, six species were recorded (four new to science). Interestingly, only a relatively small proportion of the lithistid species known from the NEA (7 out of 17) were found during the present study. Finally, the finding of 19 large specimens of M. cf. azorica in the same station in the Hyères Seamount (st. DW202), suggests that this species may occur in relatively larger densities, possibly forming a sponge ground in this area. However, this would require verification with other sampling and observation tools such as remotely operated or autonomous underwater vehicles (ROV/AUV). Such finding would add on to the aggregations dominated by Leiodermatium pfeifferae, recently reported on three seamounts in the Western Mediterranean Sea (Maldonado et al., 2015), which suggests that some extant lithistids may still form highly structured habitats comparable to the Mesozoic reefs (Maldonado et al., 2015; Reid, 1967),
Several paradigms in seamount ecology, including the seamount endemism hypothesis, have been heavily debated in recent years, with some authors considering seamounts as places of high endemism (de Forges, Koslow & Poore, 2000), while others attributed the observed patterns to sample bias (Samadi et al., 2006; see also McClain, 2007; Rowden et al., 2010). In our study, the majority of the species (Neoschrammeniella inaequalis sp. nov., N. piserai sp. nov., N. pomponiae sp. nov., Discodermia. arbor sp. nov., D. kellyae sp. nov., D. ramifera, Macandrewia minima sp. nov., M. robusta, Leiodermatium lynceus and Exsuperantia levii sp. nov.) were only found on one of the seamounts. These findings concur with a study on lithistids of the Norfolk Ridge (New Caledonia) where the authors reported 16 species (seven new to science, including a new genus) with the half of the species (eight) restricted to one seamount (Schlacher-Hoenlinger, Pisera & Hooper, 2005). On the other hand, five species, M. cf. azorica, M. schusterae sp. nov., S. elongatus sp. nov., L. tuba sp. nov. and E. archipelagus, have a wider distribution (found in three to five seamounts), and the latter three are shared between the two seamount groups. The differences in diversity and distribution found in our study may be a result of uneven sampling effort between the different seamounts (between 2 and 35 stations) and the two seamount groups (92 stations in Seamount 1 vs 131 stations in Seamount 2).
When examined at a larger scale, seamounts share most species with the Azores and Canary archipelagos, with seven (D. ramifera, D. verrucosa, M. azorica, M. robusta, E. archipelagus, L. lynceus, P. (P.) grimaldii) and six species (D. verrucosa, M. azorica, E. archipelagus, L. lynceus, L. tuba sp. nov. and S. elongatus sp. nov.) shared, respectively. Given the relative proximity between localities and the oceanographic setting, it would be expected that the Azores would share more species with the Great Meteor group, instead of the Canaries, Madeira, Selvagens and the continental shelf of the Lusitanian group (Fig. 1). However, this is not observed in our study as only two species (D. ramifera and M. robusta) are exclusively shared between the Azores and the Meteor Seamount group. One species (L. lynceus) is common to Azores, Madeira, Canaries and the two groups of seamounts, and two species (M. azorica and D. verrucosa) are shared between the Meteor group and the oceanic islands. All the species found in the Lusitanian group are shared with the archipelagos and/or the Meteor Seamount, with only one exception, N. inaequalis sp. nov. that is exclusively known from the Gorringe Seamount. However, none of the species reported from the Portuguese (Corallistes elegantior Schmidt, 1870) and Moroccan continental shelves (Theonella annulata Lendenfeld, 1907) were found to occur in the Lusitanian seamounts group. It should be noted that the description of C. elegantior is vague and does not provide a detailed characterization of all spicules. Moreover, this species was never observed since its description by Schmidt (1870) in Portugal or in the surrounded areas, thus it should be considered a taxon inquirendum. Neophrissospongia nolitangere Pisera & Vacelet, 2011 a species reported from all oceanic islands (Carvalho, Pomponi & Xavier, 2015; Cruz, 2002; Topsent, 1904) and the Mediterranean sea (Manconi, 2011; Pisera & Vacelet, 2011) and Corallistes masoni Bowerbank, 1869 reported from Madeira (Bowerbank, 1869; Carvalho, Pomponi & Xavier, 2015) and Canary Islands, were also not found in this study. If we compare the diversity between NEA and the Mediterranean Sea, only five species, viz. N. nolitangere, Neoschrammeniella bowerbankii (Johnson, 1863), L. lynceus, L. pfeifferae (Carter, 1876) and Siphonidium ramosum (Schmidt, 1870) out of 36, are shared between these two areas. Finally, whether some of the species here described for the first time are shared with the Northwest Atlantic and/or the Caribbean Sea also remains to be assessed, since the lithistid fauna of these areas is known to be far more diverse than currently reported but awaits formal description (A. Pisera, 2018, personal communication; Schuster et al., 2019). Therefore, and given the still limited and uneven sampling of the various areas, we refrain from considering the species herein described endemic to these seamounts or seamount groups.
Future studies employing a more comprehensive sampling design and modern technologies would be required to test the extent to which an interplay between intrinsic (dispersal potential) and extrinsic (seamount age, isolation and area) factors underpin and shape the observed diversity and endemism patterns of the fauna of these seamounts.
Spicules dimensions
Several morphological features are used in taxonomy and classification of Porifera and among them, the skeletal elements (spicules, fibres) and their arrangement are the most used. This is mainly due to historical reasons, since specimens would be sent for taxonomic assignment, sometime after collection and preservation, and usually having lost some of its live characteristics such as colour or consistency (Bergquist, 1970). Spicules sizes, which occur over a relatively large range are also important for species determination (Bergquist, 1970), altough some studies have shown that biophysical environmental conditions and life cycle can lead to some intraspecific varibaility (Bavestrello, Bonito & Sarà, 1993; Cárdenas & Rapp, 2013; Mercurio et al., 2000). In the case of lithistids sponges, the identification is mainly based on the shape and development of desmas and other accompanying spicules (Bergquist, 1970; Lévi, 1991).
Whether spicule size is as relevant for lithistids as in other taxonomic groups remains to be assessed. However, in the material examined in our study, we have found some differences in the size of the spicules for some species in comparison with the type material. Examples include D. ramifera, D. verrucosa, M. cf. azorica, M. robusta, E. archipelagus and P. (P). grimaldii. Specimens of D. ramifera and D. verrucosa despite being slightly larger than the holotypes and having been sampled at similar depths, present smaller cladomes of the discotriaenes, as well as their acanthomicroxeas and acanthorhabds (Table 2). In the case of M. cf. azorica and M. robusta the same pattern repeats, with exception of the microxeas on both specimens analysed here which are larger than those in the respective holotypes (Table 3). Finally, in E. archipelagus all the spicules are smaller than those in the holotype, even though the specimen itself has nearly the same size as the type material (Table 4). P. (P.) grimaldii is the only one that has slightly larger spicules compared with the type material (Table 7). These variations were also found in other deep water tetractinellids and were assumed to be related to the depth and/or silica concentration, where deeper specimens have larger spicules due to the availability of silica in the water (Cárdenas & Rapp, 2013). However, one cannot find a correlation with the depth since: (1) D. ramifera and D. verrucosa were sampled at similar depths as the holotypes, (2) the depth at which the type material of M. azorica was sampled is unknow preventing us to make any assumption, (3) M. robusta was found at shallower depths in the Hyères seamount and yet its spicules were in general smaller, (4) P. (P.) grimaldii was found within the same depth range as the holotype and has larger spicules, thus the depth seems to not be related with the size of the spicules. The amount of silica in the water does not seem to be related either since these two groups of seamounts have many lithistids, and they possibly require large amounts of silica to build their skeleton. Another explanation is that lithistids are very efficient at removing the silica from the water thus, not requiring large amounts of this element (Alvarez et al., 2017; Maldonado et al., 2015). Since there is no data regarding the biogeochemical parameters of the water column upon the time of collection of the material, it remains unclear if the cause of this variation are abiotic factors or intraspecific variation due to distinctive geographical area, as it was also observed in other astrophorins (Van Soest, Beglinger & de Voogd, 2010) including lithistids (Pisera & Vacelet, 2011).
Conclusions and Identification Key
The discovery of ten new lithistid species in the NE Atlantic seamounts and the additional record of another seven species, emphasises how diverse these ecosystems are and how our knowledge on the diversity of this group of sponges is still limited. Whether the patterns of distribution here reported are due to sampling bias, or true cases of endemism, requires further investigation.
The factors behind the variability on the spicules sizes, found in some species compared to those of the holotypes, remain unclear and more studies are needed in order to shed light on the factors behind this variability. This is particularly important on the field of sponge taxonomy since spicules are a key element for their identification. Future expeditions to these seamounts, with the use of ROVs, will allow us to have a better picture of this diversity and confirm if there are sponge grounds dominated by lithistids in the area.
An identification key of all lithistid species reported to date for the NE Atlantic and Mediterranean Sea is presented below (Table 9).
| 1. | Desmas are dicranoclones | 2. Corallistidae |
| Desmas are tetraclones, ectosomal spicules are phyllotriaenes or discotriaenes | 8. Theonellidae | |
| Desmas have a triaenose crepsis, rarely monaxial crepis | 11. Macandrewiidae | |
| Demas are triders | 13. Phymaraphiniidae | |
| Desmas are rhizoclones, no ectosomal spicules, microscleres absent, raphides may be present | 15. Azoricidae | |
| Desmas are rhizoclones, ectosomal spicules, if present, are rhabds or oxeas, sigmaspires may be present | 17. Scleritodermidae | |
| Desmas are rhizoclones, no ectosomal spicules, exotylostyles present | 20. Siphonidiidae | |
| Demas are monaxial or probably monaxial but not rhizoclones, styles are present | 23. Desmanthidae | |
| 2. | Dichotriaenes with spines and tubercles on the top of the cladome, microscleres are streptasters/amphiasters | 3. Neophrissospongia |
| Dichotriaenes are smooth | 4. | |
| 3. | Cup-shaped, dichotriaenes with very massive, thick and irregular cladomes; dicranoclones extremely tuberculated with a central core; spinose microstyles | N. endoumensis |
| Encrusting thick plate with rounded margins, two types of dichotriaenes: with few tubercles or smooth; triaenes with few tubercles present, but rare; styles/sub-tylostyles | N. nana | |
| Ear- or cup-shaped when young to large to flabellate masses when old; dicranoclones very tuberculated; spinose microtylostyles | N. nolitangere | |
| Clavate in habitus with a narrow and central spongocoel; dicranoclones have sparsely distributed round tubercles; spinose microstylostyles | N. radjae | |
| 4. | One type of microscleres, spirasters with pointed arms | 5. Corallistes |
| Desmas have a root/vine-like appearance, microscleres are two types of microacanthoxeas, spirasters and streptasters | 6. Isabella | |
| Two types of microsclers (metasters and spirasters), oxeas usually present | 7. Neoschrammeniella | |
| 5. | No proper description has been given to this species in the original description and there are no more records of this species. The type material should be re-examined | C. elegantior |
| Sinuously fan-shaped with rounded and thin walls; microscleres are spirasters with long and thin arms | C. masoni | |
| 6. | Irregular rounded sponge of dark purple-brown colour; ectosomal spicules are irregular dichotriaenes, short- and long-shafted triaenes; two types of long oxeas (type I: long and thick with blunt tips; type II long, thin, curved with acerate tips) | I. harborbranchi |
| 7. | Cup-shaped to contorted lamellate masses with thick walls; smooth surface; several thin oxeas in the inner surface; dicranoclones have irregular and high tubercles, that can be subdivided into several smaller tubercles | N. bowerbankii |
| Cup- to flattened cup-shaped with a concave center and rounded edges; smooth surfaces; dichotriaenes are very variable in shape and size; long-shafted triaenes can be present; oxeas are large and thin; dicranoclones of vine-like appearance, with some tubercles that are smooth or rugose | N. inaequalis | |
| Large cup-rectangular in shape with smooth surfaces; dicranoclones are irregular, compact, usually smooth, with few tubercles that are usually smooth; no oxeas; some microscleres are irregular, resembling irregular rhabds with spiny tips | N. piserai | |
| Cup-rounded in shape with a small pedicel; surfaces are crumble and hispid; oxeas are long with sharp tips; dicranoclones are compact, densely covered by numerous and ornamented tubercles | N. pomponiae | |
| 8. | Ectosomal spicules are discotriaenes, desmas are tetraclones, oxeas usually present, microscleres are acanthoxeas and acanthorhabds | 9. Discodermia |
| Ectosomal spicules are phyllotriaenes to discotriaenes, microscleres are acanthorhabds | 10. Theonella | |
| 9. | Tree-like shaped, with a long stem smooth surface with some rugosities/protuberances; discotriaenes of “square” to “circular” shape or with “idented” cladomes; oxeas not present; tetraclones very tuberculated near the surface and smoother in the inner part of the sponge. | D. arbor |
| Massive, irregular in shape, with large protuberances of round shape; rugose surface; discotriaenes very variable in the shape of the cladomes varying from oval to indented, and size of rhabdomes; strongyles with one tip rounded and the other sharp | D. kellyae | |
| Small irregular mushroom shaped, with a concave upper side, a short stem and smooth surface; discotriaenes with a round to oval cladome; tetraclones with smooth rays and strongly branched and tuberculated zygomes; oxeas | D. polydiscus | |
| Small, polymorphic, varying from spherical to irregular masses with protuberances, attached by a short pedicel; smooth surface; discotriaenes have very variable cladomes, from circular and concave to oval with irregular margins; tetraclones are smooth and irregular; oxeas not present | D. polymorpha | |
| Small, elongated and branched with a smooth surface; discotriaenes have a round/oval to irregular and indented cladome; oxeas; tetraclones have smooth rays and tuberculated zygoses, that are usually smooth | D. ramifera | |
| Cup-shaped to spherical polymorphic, with several round protuberances; discotriaenes are round/oval, smooth, often indented; oxeas; tetraclones are large, robust densely covered by tubercles | D. verrucosa | |
| 10. | Tetraclones are tuberculated but sometimes smooth in the center; phyllotriaenes have a simple or bifurcated cladome with rounded edges, and a short rhabdome* | T. annulata |
| 11. | Dentate ectosomal phyllotriaenes/discotriaenes, smooth oxeas, microscleres are smooth microxeas. | 12. Macandrewia |
| 12. | Cyathiform to flabellate, with undulating rounded margins and a short stem; outer surface is smooth with small pores and inner surface is smooth but the oscules have slightly raised margins; desmas are smooth, either resembling tetraclones or rhizoclones, very branched at the end | M. azorica |
| Small round-globular shaped, with a very short and slender pedicel and smooth surface; phyllotriaenes have incised and tuberculated cladomes; desmas with triaenose crepsis, usually smooth but some rugosities can be present | M. minima | |
| Sponge with a vast base where it stands two or more truncks of cylindrical shape, with the top divided into short and obtuse branches | M. ramosa | |
| Ficiform to globular in shape, with a thick and short pedicel; top of the sponge can be curved or slightly depressed; monocrepid desmas are smooth, with short and thick tubercles | M. robusta | |
| Foliate to vase shape, with thick and contorted lamellas and a small pedicel; phyllotriaenes with incised cladomes on the edges; desmas with a triaenose crepsis, smooth, irregular, with the several short and blunt branches at the end | M. schusterae | |
| 13. | Clavate or globular knob-like shaped, ectosomal spicules are phyllo- to discotriaenes; subtylostyles to tylotes; microscleres are acanthorhabds, acanthomicroxeas and steptasters/amphiasters | 14. Exsuperantia |
| 14. | Columnar to ficiform, sometimes with lateral protuberances/branches; smooth surface ectosomal spicules are phyllotriaenes. | E. archipelagus |
| Clusters of globular to ficiform knob-like short fingers with an apical osculum; surface is rugose; ectosomal spicules are phyllo- to discotriaenes | E. levii | |
| 15. | Foliate or vase shaped in habitus, long oxeas, no microscleres | 16. Leiodermatium |
| 16. | Narrow ear-shaped or cylindrical sponge with deeply incised rounded margin; outer surface with oscules located on top of small elevations; inner surface with densely distributed pores | L. lynceus |
| Flattish, cabbage-like, infoliated, with branched sinuous laminae; outer surface has pores and inner surface has scattered ostia slightly raised on papillary eminences; rhizoclones with branched arms | L. pfeifferae | |
| Massive lamellate vase to contorted thin walls, sometimes forming a cone; surfaces are smooth and similar at naked eye; outer surface has slightly depressed openings while the inner surface has several small openings and numerous oxeas | L. tuba | |
| 17. | Ectosomal spicules are acanthorhabds/strongyles, no microscleres | 18. Aciculites |
| Vase or foliate in shape, ectosomal spicules absent, microscleres present | 19. Microscleroderma | |
| 18. | Massive cerebellum-like, sub-oval with a wide base; surface is smooth with subdermal canals covered by a dermal membrane; inhalant areas are irregularly distributed in depressed concavities, while exhalant areas are elevated; anisostrongyle to tylostrongyles, usually with a spinose/rugose head | A. mediterranea |
| 19. | Irregular mass of contorted, irregularly undulating lamellae; two types of oxeas: thick, straight with acerate tips or thin, hair-like oxeas rarely straight; sigmaspires are C- or S-shaped with short spines | M. lamina |
| 20. | With long fistules, exotylostyles with ornamented heads | 21. Siphonidium |
| No fisutles, deep and narrow atrial cavity, ear- or vase-shaped | 22. Gastrophanella | |
| 21. | Small, irregularly massive to cylindrical in shape with numerous small fistules; desmas are tuberculated | S. ramosum |
| Polymorphic, cilindrical to arborescent, sometimes bulb-shaped with numerous fistules; very spiny rhizoclones with slim arms ornamented with microspines in the edges; styles present (rare) | S. elongatus | |
| 22. | Irregular pear-shaped with lateral depressed shallow concavities and an osculum on the top; monaxons with a slightly protruding head | G. phoeniciensis |
| 23. | Massive in habitus, desmas branching in various planes | 24. Petromica |
| Encrusting in habitus, no microscleres | 25. | |
| 24. | Massive with conical form; irregular surface with conules and dispersed pores; desmas are poorly articulated with the tips divided into massive low spines; anisoxeas to styles (or strongyloxeas) and anisorhabds | P. (Petromica) grimaldii |
| 25. | Desmas of the outer layer are trider-like | 26. Desmanthus |
| Desmas hook-like, with pointed spines | 27. Sulcastrella | |
| 26. | Encrsuting with a hispid outer surface; desmas are of two types: trider-like, tri- to tetrapodial, branched in the upper parts and tuberculated in the surface; styles are long | D. incrustans |
| 27. | Small circular encrusting sponge with a hispid surface; monocrepid desmas are irregular, strongly tuberculated in some places and form a solid basal crust; styles are slightly curved | S. tenens |
Notes:
Sources: Carter (1873), Carvalho & Pisera (2019), Lendenfeld (1907), Manconi, Serusi & Pisera (2006), Manconi & Serusi (2008), Perez et al. (2004), Pisera & Lévi (2002a), Pisera & Lévi (2002d), Pisera & Lévi (2002f), Pisera & Lévi (2002g), Pisera & Lévi (2002h), Pisera & Lévi (2002i), Pisera & Vacelet (2011), Topsent (1904) and Vacelet (1969).