
Genome-wide characterization of the auxin response factor (ARF) gene family of litchi (Litchi chinensis Sonn.): phylogenetic analysis, miRNA regulation and expression changes during fruit abscission
- Published
- Accepted
- Received
- Academic Editor
- Genlou Sun
- Subject Areas
- Agricultural Science, Molecular Biology, Plant Science
- Keywords
- Litchi, Auxin, Auxin response factors, miRNA, Abscission
- Copyright
- © 2019 Zhang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Genome-wide characterization of the auxin response factor (ARF) gene family of litchi (Litchi chinensis Sonn.): phylogenetic analysis, miRNA regulation and expression changes during fruit abscission. PeerJ 7:e6677 https://doi.org/10.7717/peerj.6677
Abstract
Auxin response factors (ARFs) play fundamental roles in modulating various biological processes including fruit development and abscission via regulating the expression of auxin response genes. Currently, little is known about roles of ARFs in litchi (Litchi chinensis Sonn.), an economically important subtropical fruit tree whose production is suffering from fruit abscission. In this study, a genome-wide analysis of ARFs was conducted for litchi, 39 ARF genes (LcARFs) were identified. Conserved domain analysis showed that all the LcARFs identified have the signature B3 DNA-binding (B3) and ARF (Aux_rep) domains, with only 23 members having the dimerization domain (Aux_IAA). The number of exons in LcARF genes ranges from 2 to 16, suggesting a large variation for the gene structure of LcARFs. Phylogenetic analysis showed that the 39 LcARFs could be divided into three main groups: class I, II, and III. In total, 23 LcARFs were found to be potential targets of small RNAs, with three conserved and one novel miRNA-ARF (miRN43-ARF9) regulatory pathways discovered in litchi. Expression patterns were used to evaluate candidate LcARFs involved in various developmental processes, especially in flower formation and organ abscission. The results revealed that most ARF genes likely acted as repressors in litchi fruit abscission, that is, ARF2D/2E, 7A/7B, 9A/9B, 16A/16B, while a few LcARFs, such as LcARF5A/B, might be positively involved in this process. These findings provide useful information and resources for further studies on the roles of ARF genes in litchi growth and development, especially in the process of fruit abscission.
Introduction
Auxin plays a central role in numerous aspects of plant developmental and physiological processes, including embryogenesis, apical dominance, vascular elongation, flowering, fruit development, and lateral root initiation (Woodward & Bartel, 2005; Fleming, 2006). Auxin response factors (ARFs) are a group of important transcription factors in the auxin signaling pathway, which can activate or repress the expression of early/primary auxin response genes by binding to the auxin response element (AuxRE) site in their promoter regions (Liscum & Reed, 2002; Guilfoyle & Hagen, 2007). A typical ARF is characterized by a highly conserved N-terminal B3-type DNA binding domain (DBD) that recognized the AuxRE motif, an activation domain or repression domain, and a carboxy-terminal dimerization domain (domain III/IV), which is involved in protein–protein interactions by dimerizing with auxin/indole-3-acetic acid (Aux/IAA) family genes (Kim, Harter & Theologis, 1997; Guilfoyle & Hagen, 2007; Piya et al., 2014).
Auxin response factors exert pivotal function in the regulation of plant growth and development through the auxin signaling pathway (Kepinski & Leyser, 2005; Li et al., 2016). Due to the significance of ARFs, genome-wide characterization of ARFs have been completed in many species such as model plants Arabidopsis (Hagen & Guilfoyle, 2002) and Solanum lycopersicum (Zouine et al., 2014), important crops such as soybean (Ha et al., 2013; Le et al., 2016) maize (Xing et al., 2011; Wang et al., 2012), fruit trees like citrus (Citrus sinensis) (Xie et al., 2015) apple (Malus domestica) (Luo et al., 2014), and so on. From embryogenesis to flowering, mutants in members of ARFs exhibit diverse phenotypes, which show their unique and redundant functions for plant development. For instance, in Arabidopsis thaliana, arf1 and arf2 mutations affect leaf senescence and floral organ abscission (Ellis et al., 2005) and the loss of AtARF3 causes defects in gynoecium patterning (Nemhauser, Feldman & Zambryski, 2000; Liu et al., 2014a). AtARF5 influences embryo, root, and shoot development (Krogan et al., 2012; Crawford et al., 2015). AtARF9 acts in suspensor cells to mediate hypophysis specification (Rademacher et al., 2012), and AtARF10/16/17 play vital roles in negatively regulating seed germination and post-germination activities (Liu et al., 2007).
Recently, small RNAs, especially miRNAs, have been emerging as critical regulators in almost all aspects of plant growth and development. Many members of the ARF family have been reported to be targets of miRNAs. In Arabidopsis, AtARF6 and AtARF8 are targets of miR167 (Wu, Tian & Reed, 2006), AtARF10/16/17 are targets of miR160 (Liu et al., 2007, 2010), and ARF2/3/4 are targets of trans-acting siRNAs (tasiRNAs) generated from miR390-targeted TAS3 gene (trans-acting siRNA gene 3) (Allen et al., 2005; Axtell et al., 2006). These targeting relationships of miRNA on ARF genes are widely conserved in land plants (Xia, Xu & Meyers, 2017) and also in horticultural plants (Chen et al., 2018b). It has been reported that down-regulation of ARF6 and ARF8 by miR167 leads to floral development defects and female sterility in tomatoes (Liu et al., 2014b). Down-regulation of sly-miR160, increasing the expression of its targets SlARF10/16/17, regulates auxin-mediated ovary patterning as well as floral organ abscission and lateral organ lamina outgrowth (Damodharan, Zhao & Arazi, 2016). In Arabidopsis and tomato, the tasiRNA-mediated regulation of ARF2 is involved in controlling the onset of leaf senescence and floral organ abscission (Ellis et al., 2005; Lim et al., 2010; Guan et al., 2014; Ren et al., 2017).
Litchi (Litchi chinensis sonn.), an important economic fruit tree in southern China, usually undergo serious fruit abscission before harvest, leading to a low yield. Many studies demonstrate ARFs play critical roles in regulating plant organ abscission (Ellis et al., 2005; Kuang et al., 2012; Guan et al., 2014; Xie et al., 2015; Xu et al., 2015), while which ARFs are involved or more important than other ARFs in the fruit abscission in litchi remains elusive. Here, we identified 39 ARF genes in litchi. Gene structure, phylogeny, and targeting relationship with miRNAs were characterized. The expression of LcARFs was examined in diverse tissues and in the process of fruit abscission which was induced by three different treatments. Among them, ARF2D/2E, 7A/7B, 9A/9B, 16A/16B, and LcARF5A/B, were found to be associated with litchi fruit abscission. Our results offer new knowledge and resources to study the function of plant ARF genes and their roles in the fruit abscission in litchi.
Materials and Methods
Plant materials and treatments
The young fruitlet used for RNA-seq in our study were collected from 9-year-old litchi trees (L. chinensis Sonn. cv. “Feizixiao”) in an orchard located at South China Agricultural University (Guangzhou, China). Three treatments have been performed 25 day after anthesis. The three treatments included ethephon (ETH), girdling plus defoliation (GPD), and dipping in 20 mg/L 2, 4-D for 1 min after girdling plus defoliation (GPDD). Details can refer to Li et al. (2015) and Peng et al. (2013). Samples of “Feizixiao” (FZX) for qRT-PCR were obtained from the orchard of Guangzhou Fruit Research Institute (Guangdong, China). Tissues including fruit-bearing shoots (FBS), young leaves (YL), mature leaves (ML), female flower (FF), male flower (MF), undeterminated-sex flower (USF), and young fruit (YF) of 25 days after fertilization were collected from different directions of each tree. After separation, all tissues were quickly frozen in liquid nitrogen and stored at −80 °C before usage.
Identification and phylogenetic analyses of LcARFs
Amino acid sequences of 25 and 23 ARFs from rice (Wang et al., 2007) and Arabidopsis (Okushima et al., 2005; Wang et al., 2007) were downloaded from UniProt (http://www.uniprot.org/) and 22 ARFs from tomato (Zouine et al., 2014) were downloaded from Sol Genomics Network (https://solgenomics.net/). These ARFs were used as bait to identify potential ARFs in litchi genome and obtained by BLAST analysis in TBtools (Chen et al., 2018a), with the cutoff E-value set at 1e−10. All sequences were then further validated by conserved domain search against the CDD (http://www.ncbi.nlm.nih.gov/cdd/) and PFAM (http://pfam.xfam.org/) databases. Based on the optimized alignment of amino acid sequences of LcARFs proteins, achieved by TrimAL 1.3 (http://phylemon2.bioinfo.cipf.es/index.html) after the initial sequence alignment in ClustalX 2.1, a maximum likelihood (ML) tree was constructed in MEGA 7.0 software with a bootstrap of 1,000 replicates. Intron and exon distribution patterns and genome structure were analyzed and visualized in TBtools (Chen et al., 2018a). The details of predicted protein sequences of LcARFs were shown in Datas S1 and S2.
LcARF targets of miRNAs and RNA-seq analysis
Most of the ARF-targeting miRNA/tasiRNAs were identified in our previous study (Ma et al., 2018). To further validate the targeting relationship, target sites were verified by psRNATarget (http://plantgrn.noble.org/psRNATarget) (Dai & Zhao, 2011) and miRBase (http://www.mirbase.org/).
RNA-seq analysis was carried on as described previously (Li et al., 2013). These RNA-seq data were normalized using the TPM method (Wagner, Kin & Lynch, 2012). Based on the sequences of the identified LcARFs, a BLASTN search was conducted to find their gene counterparts in previous data (Li et al., 2013). Finally, global gene expression profiles were visualized by heatmap via TBtools software (Chen et al., 2018a).
Quantitative RT-PCR
Total RNA of the above seven tissue samples was extracted by column plant RNAout kit (Tiandz, Beijing, China) according to the manufacturer’s instructions. About two µg total RNA was applied to synthesized first strand cDNA using reverse transcriptase AT311-03 (TransGen Biotech, Beijing, China). PCR primers (Table S1) of LcARFs and reference genes GAPDH and EF (Zhong et al., 2011) were designed by Primer Premier 5.0. qRT-PCR was performed according to the manufacturer’s specifications of THUNDERBIRD qPCR MIX QPS-201 (Toyobo, Shanghai, China) on a LightCycler 480 (Roche, Rotkreuz, Switzerland). Each expression profile was independently verified in three biological replicates. Relative expression level of each gene was calculated by the 2−ΔΔCt method (Livak & Schmittgen, 2001). Significance analysis was conducted in SPSS version 22.0 and visualized by SigmaPlot 12.5.
Results
Identification and phylogenetic analysis of LcARF genes
To identify all ARF members in litchi, protein sequences of ARFs in rice, Arabidopsis and tomato were used as queries in BLASTP to search against a litchi annotated gene database. A total of 68 potential ARF genes were identified. After redundant result elimination and further conserved domain validation, 39 LcARFs were ultimately obtained. Among them, 23 protein sequences contained B3, ARF, and the Aux/IAA domains. Length of these litchi ARF proteins ranged from 260 (LcARF1B) to 1,200 (LcARF4A) and the relative molecular mass of them varied from 29,522.5 (LcARF1B) to 144,936.9 Da (LcARF16C), with PIs in the range of 5.28 (LcARF16C) to 8.64 (LcARF19A) (Table S2). According to the subcellular localization predictor (CELLO v.2.5; http://cello.life.nctu.edu.tw), most LcARFs were predicted to be located in the nucleus.
A phylogenetic tree was generated using the ML method based on alignment of litchi ARFs with their orthologs from rice, Arabidopsis and tomato (Fig. 1). All 109 ARFs fell into three broad groups: class I, II, and III, which contained 53, 34, and 22 members, respectively. Obviously, most ARFs of the four species were clustered together in the first two classes. Litchi ARFs were named according to their positions with orthologs from the other three species in the tree. Thus, the 39 LcARFs could also be assigned to three separate clusters as well. Class I contained LcARF1/2/3/4/9/18 (1A/B/C/D, 2A/B/C/D/E/F, 3A/B, 4A/B/C, 9A/B, 18A/B); Class II included LcARF5/6/7/8/19 (5A/B, 6A/B/C/D, 7A/B, 8A/B, 19A/B); Class III included LcARF10/16/17 (10A/B, 16A/B/C/D, 17A/B).
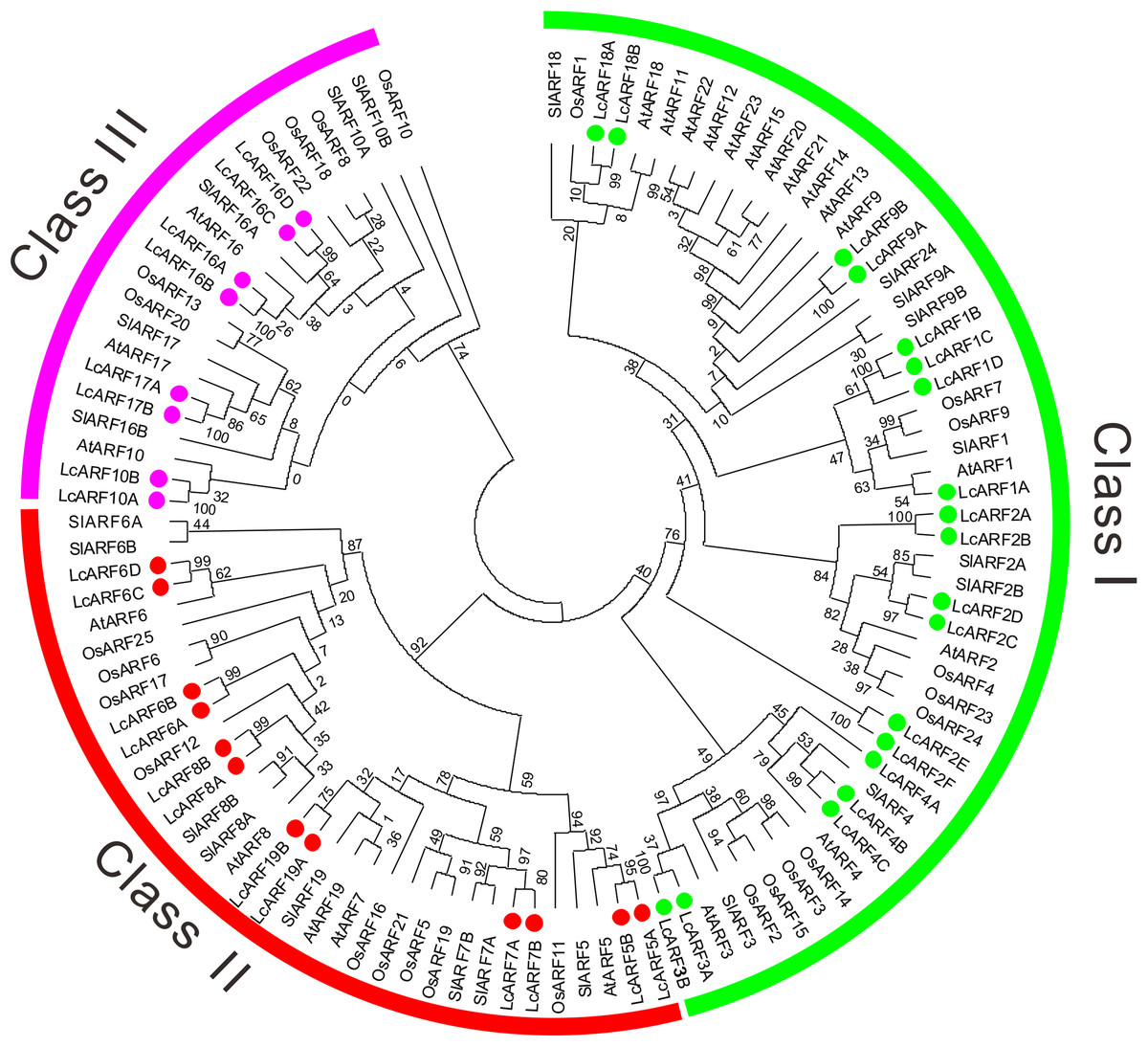
Figure 1: Phylogenetic analysis of ARFs from litchi, rice, Arabidopsis, and tomato.
The phylogenetic tree was generated using the maximum likelihood method with the JTT matrix-based model (Kumar, Stecher & Tamura, 2016) and the bootstrap test was carried out with 1,000 bootstrap replicates. Numbers on the nodes indicate the credibility values of each clay. Three subgroups were shown as Class I, II, and III.{kind=link}
Gene structure and conserved domains of litchi ARFs
To better understand the structure evolution of LcARF genes, their gene structure (intron/exon number and positions) and functional domains were analyzed. As shown in Fig. 2, 23 LcARFs harbored the typical ARF protein structure which composed of a highly conserved DBD in the N-terminal region with a plant specific B3-type subdomain, an Auxin-resp subdomain, and an AUX_IAA dimerization subdomains. The remaining 16 LcARFs contained only B3 and Auxin_resp subdomains. Gene structure analysis revealed that the exon number of class III (2–4 exons) was significantly less than the other two groups (8–17 exons for class II and 10–16 exons for class I). These results provided additional evidence confirming the phylogenic relationships among LcARFs.
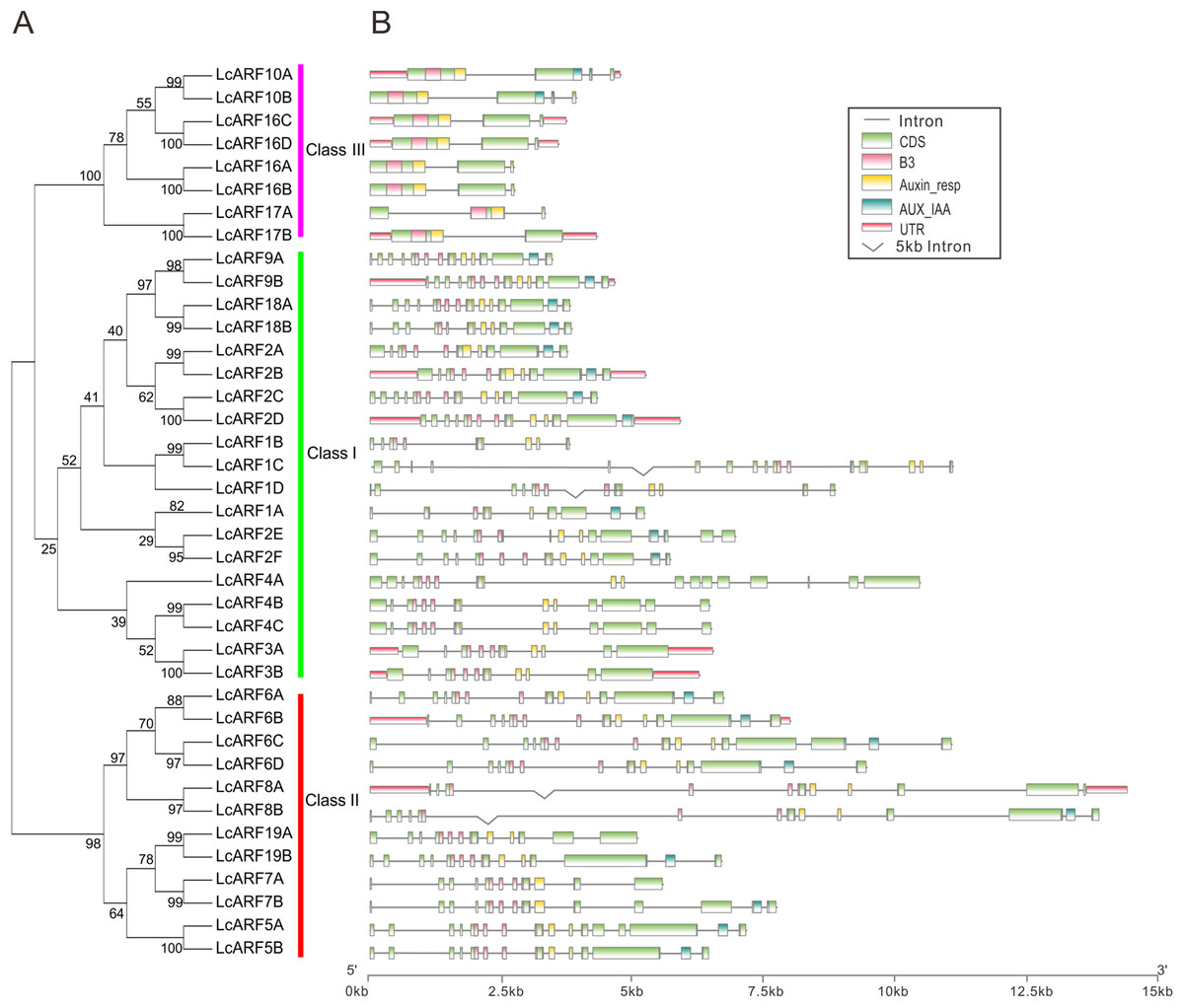
Figure 2: Phylogenetic relationship, exon–intron structure, conserved domains analyses of LcARFs.
(A) Phylogenetic relationship among the litchi ARF proteins. The unrooted tree was generated using the maximum likelihood method by JTT matrix-based model. The reliability was assessed using 1,000 bootstrap replicates. Three clusters are labeled as Class I, Class II, and Class III. (B) Exon–intron structure and conserved domains of LcARFs. Information of exon, intron, and functional domain was obtained from model gene annotation and results of NCBI CDD search and visualized by TBtools. B3: B3 DNA-binding domain; Auxin-resp: ARF domain; AUX_IAA: C-terminal dimerization domain. Lengths of exons and introns and domains of each LcARF protein were exhibited proportionally.{kind=link}
Analyses of miRNA targeting LcARFs
A total of 22 out of the 39 LcARF genes were found to be the targets of miRNA (Fig. 3). All group members of class III (LcARF10A/B, 16A/B/C/D, and 17A/B) were found to be targeted by Lc-miR160. LcARF6A/B/C/D and 8A/B were members of class II and all of them were collectively targeted by Lc-miR167. Two kinds of miRNA targeting patterns were observed in the class I. One group comprising LcARF2E/2F/4B/4C/3A/3B were targeted by tasiARFs. In litchi, LcTAS3 are divided into two subgroups, long TAS3 genes(LcTAS3_1 and LcTAS3_2 ) and short TAS3 genes (LcTAS3_3 and LcTAS3_4 ), which trigger to produce one or two tasiARFs when cleaved by miR390 (Ma et al., 2018). Additionally, LcARF2E/2F incorporated one target site of tasiARF, while LcARF3A/3B/4B/4C contained two, which is in accordance with our previous study (Xia, Xu & Meyers, 2017). Interestingly, in the other group, a novel miRNA-ARF pathway was discovered, in which LcARF9A/B was targeted by the Lc-miRN43.
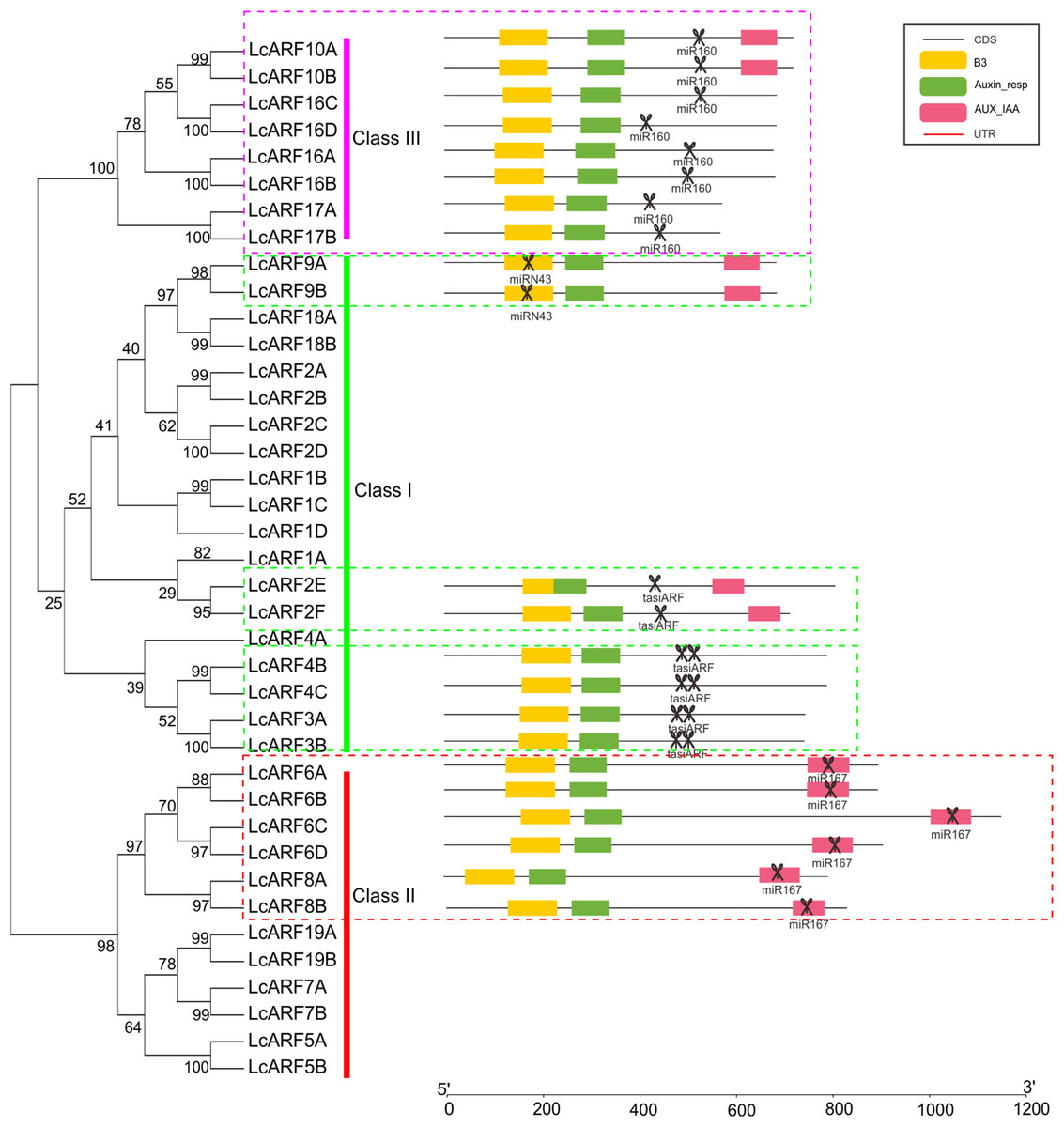
Figure 3: LcARFs targeted by some LcmiRNAs.
miR160 targets LcARF10/16/17, miR167 targets LcARF6/8, and miRN43 targets LcARF9. Additionally, the LcTAS3 was targeted by miR390 and then triggered the production of tasiARF which target LcARF2E/2F/4B/4C/3A/3B.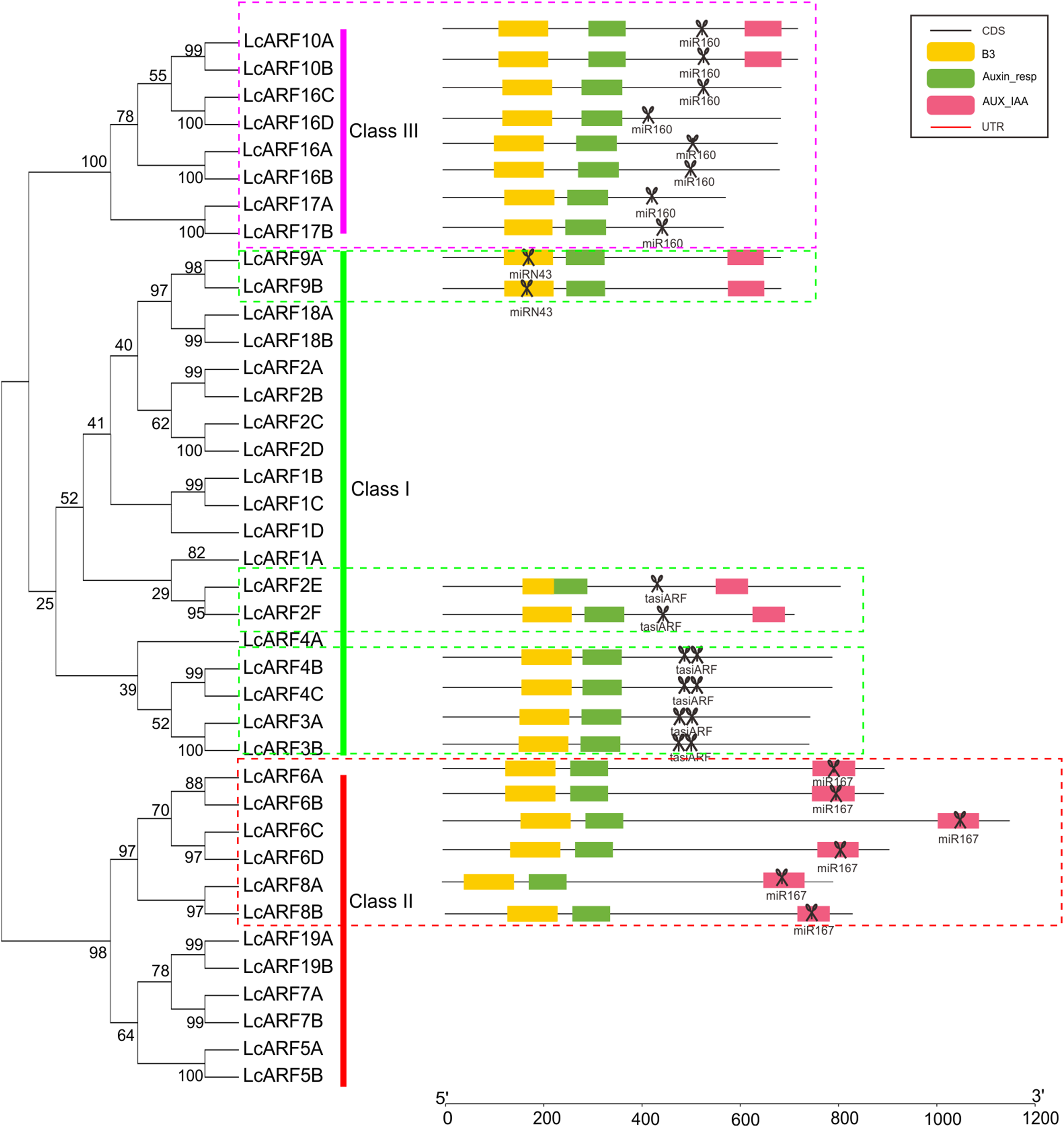{kind=link}
Expression of LcARF genes in different organs and tissues
A large body of evidence support the importance of ARF genes in plant growth and development (Ellis et al., 2005; Guilfoyle & Hagen, 2007; Lim et al., 2010; Li et al., 2016). To explore how LcARF genes function in the development of litchi, we examined their expression levels in various litchi organs/tissues by qRT-PCR. Seven organs/tissues samples, composing FBS, MF, FF, USF, ML, YL, and YFs in “FZX” were collected and tested. As shown in Fig. 4A, expression of all ARFs was detected at least in several tissues. Generally, most LcARFs show low expression in YL and YF but high in USF. LcARFs from different clades showed various expression patterns. LcARFs from class I and III were detected to be higher expressed in ML while those from class II were lowly accumulated, suggesting that these LcARFs in different classes may perform different functions in ML development (Fig. 4A). Additionally, as is shown in Fig. 4B, 10 LcARFs (LcARF2B, 3A, 4A/B, 5A/B, 6D, 9A/B, 18B) were significantly higher expressed in FF than MF, implying their potential function in ovule and ovary might be derived from FFs. Remarkably, three LcARFs (LcARF8A, LcARF10A/B) were of significantly higher expression in MF than FF, indicated that they might play roles in MF formation. Additionally, a table for LcARFs and their potential function was concluded in Table S3 according to an early study which may be useful for further functional validation.
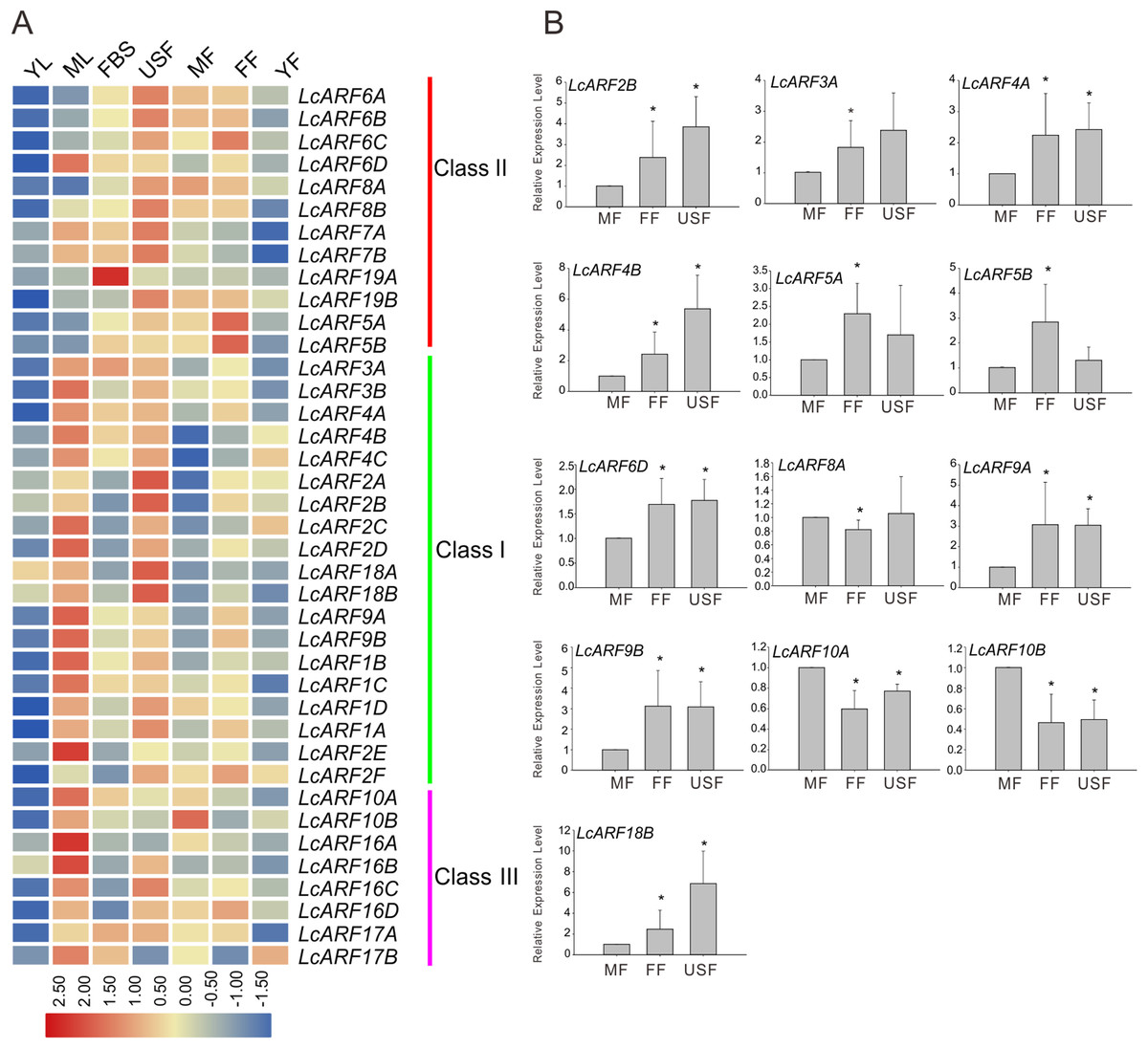
Figure 4: Expression profiles of LcARF genes in various tissues of “FZX” by qRT-PCR.
(A) Expression of all LcARFs in different tissues. The heatmap was generated based on the relative expression values of 39 LcARFs obtained by qRT-PCR in seven different tissues and organs. Red and blue were represented relatively high and lower expression (log2 ratio), respectively. Every sample has three biological replicates. YF, young fruit (25 days after fertilization); FF, female flower; MF, male flower; USF, undeterminated-sex flowers; ML, mature leaves; YL, young leaves; FBS, fruit-bearing shoots. (B) Relative abundance of LcARFs significantly expressed in FF. Data from three independent biological replicates are shown with standard deviation (SD). Asterisks on the top of bars indicate significant differences as determined by Student’s t-test (*P, 0.05).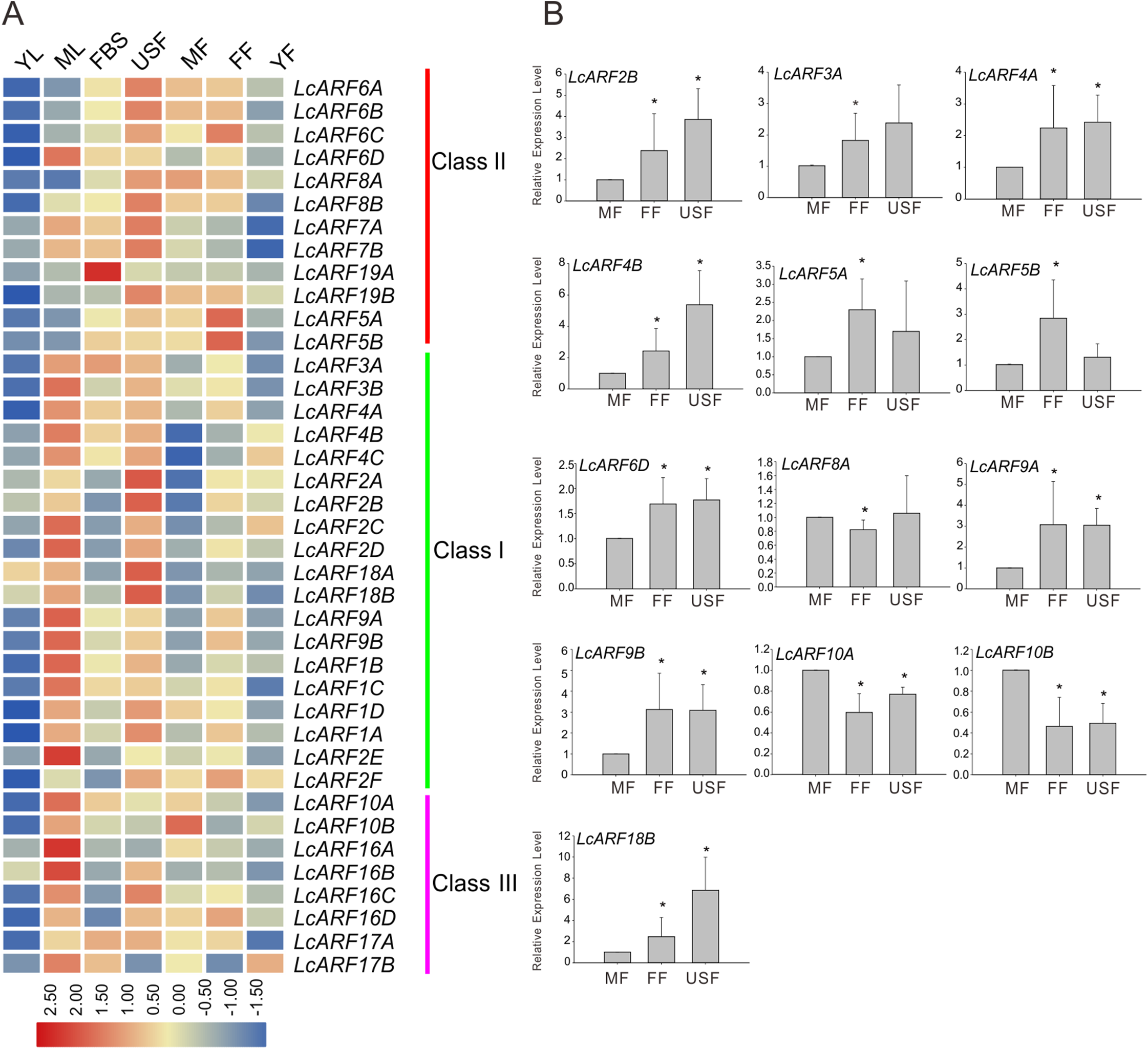{kind=link}
Expression profiles of LcARF genes in response to ETH, GPD, and GPDD treatment
Auxin is a critical signal in the abscission of fruits in plants (Blanusa et al., 2005; Meir et al., 2010; Xie et al., 2013). To explore the role of LcARFs in fruitlet abscission in litchi, RNA-seq was carried on and transcription levels of LcARF genes in the abscission zone were investigated under three treatments (ETH, GPD, GPDD). Both the ETH and GPD treatments promote fruitlet abscission while GPDD delays the process (Peng et al., 2013; Li et al., 2015). As shown in Fig. 5, the transcript expression of most LcARFs were decreased after treated by ETH and GPD, but an upsurge was present after GPDD treatment, which was corresponding to the process of promoting or inhibiting of fruitlet shedding, respectively. It seemed that there was a negative correlation between LcARFs and the fruit abscission and among these LcARFs, those from two groups (group 2 and group 3) were particularly representative. Moreover, LcARFs from group 3 were more sensitive to GPDD treatment along with stronger expression than those from group 2. Thus, we could deduce that LcARFs from group 3, including ARF2D/2E, 7A/7B, 9A/9B, 16A/16B, played major roles on the litchi fruitlet abscission. In contrast, a few LcARFs, such as group 1 including LcARF5A/5B, showed an opposite expression pattern, suggesting that they might function to accelerate the process of abscission. There were as well some LcARFs with no significant expression change and seemed to be unrelated to fruitlet abscission.
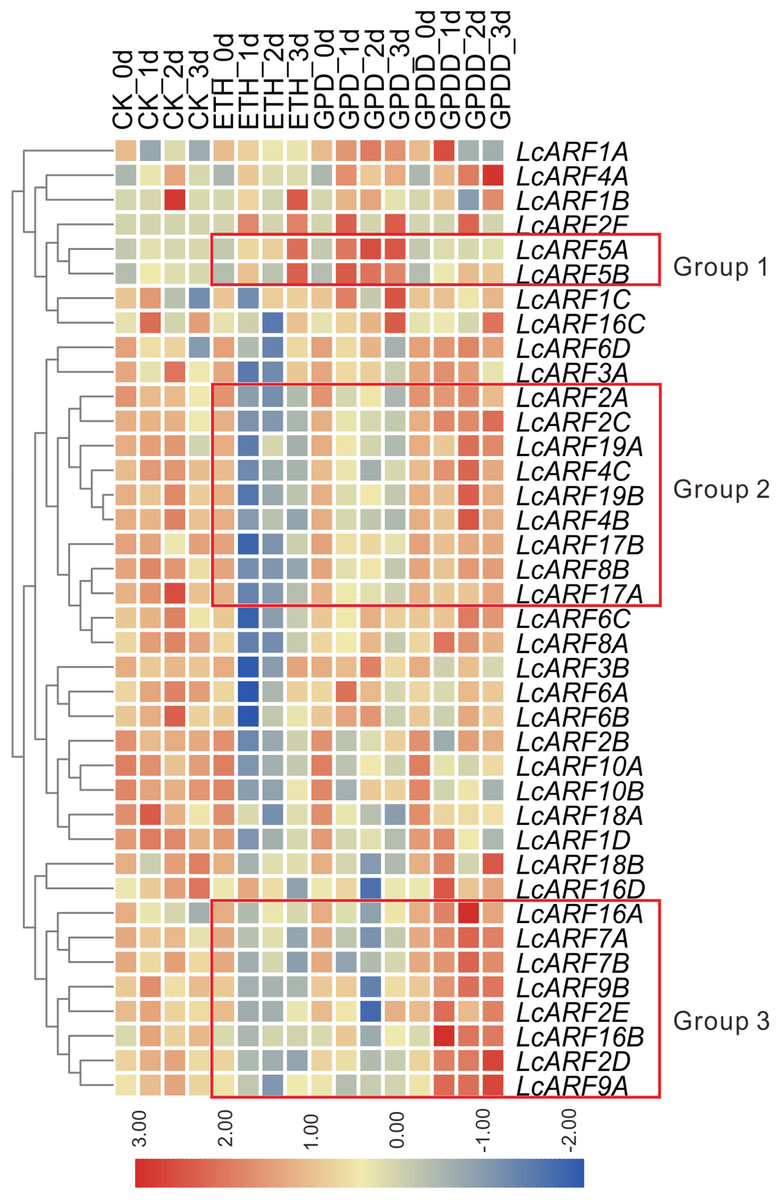
Figure 5: Expression profiles of LcARF genes in response to ETH, GPD, and GPDD treatments.
Fruit-bearing shoots of “FZX” litchi were obtained at 25 days after anthesis and then carried on ETH, GPD, and GPDD treatment from 0, 1, 2, and 3 days, respectively. CK, control; ETH, treated by ethylene; GPD, girdling plus defoliation; GPDD, dipping in 2, 4-D after GPD treatment. The heatmap was created based on the TPM values of LcARFs from the transcriptome data. In the heatmap, red and blue were represented higher and lower expression (log2 ratio), respectively. Heatmap and hierarchical clustering were performed by average linage (default) method.{kind=link}
Discussion
Litchi is an important tropic fruit tree and massive fruit abscission before harvest usually leads to low and even no production. Auxin is proposed to be one of the endogenous hormones playing significant roles in the regulation of fruit abscission in litchi (Yuan, 1988; Stern & Gazit, 2000). In this study, 39 LcARF genes were identified, which was found more than in the other model plants, such as Arabidopsis (23) (Okushima et al., 2005; Wang et al., 2007), rice (25) (Wang et al., 2007), and tomato (22) (Zouine et al., 2014), implying extensive duplication and diversification of the ARF gene family in litchi. Analysis of conserved motifs revealed that all LcARFs had a typical DBD domain required for efficient binding to AuxRE and a Auxin_resp (Fig. 2) (Hagen & Guilfoyle, 2002; Ha et al., 2013). However, only 23 of 39 LcARFs contain the AUX_IAA domain, which can mediate the dimerization of ARFs or ARF and Aux/IAA protein (Guilfoyle & Hagen, 2007). Lack of the AUX_IAA domain for dimerization makes it interesting to address questions like how these ARFs function and whether they need dimerization with other proteins. In plants, ARFs can function as transcription activators (ADs) or repressors (RDs), according to the amino acid composition of Auxin_resp domain (Guilfoyle & Hagen, 2007). In A. thaliana, ARF ADs and RDs were proposed to contain biased amino acid sequences, which ARF ADs were enriched in glutamine (Q), while RDs were enriched in serine (S), serine and proline (SP), and serine glycine (SG) (Guilfoyle & Hagen, 2001; Tiwari, Hagen & Guilfoyle, 2003). Intriguingly, no ARFs in litchi were enriched in Q, but with SPL and SP/SG enrichment; therefore none of LcARF proteins seems to be an activator (reviewed in Guilfoyle & Hagen (2007)). Further experiments are needed to verify this observation.
Much evidence demonstrates that miRNAs play essential roles in post-transcriptional gene regulation in plants (Jones-Rhoades, Bartel & Bartel, 2006; Li & Zhang, 2016). It has been found that several ARF genes are regulated by a few miRNAs. In our work, 22 out of 39 LcARF genes were found to be targets of miRNAs (Fig. 3) and LcARFs from different classes were displayed in different miRNA targeting patterns. Members of class III (LcARF10A/B, 16A/B/C/D, and 17A/B) were found to be targeted by Lc-miR160, which might affect flower development of litchi, as down-regulation of ARF10/16/17 by miRNA160 is reported to regulate floral organ abscission in tomato (Damodharan, Zhao & Arazi, 2016). Consistent with Arabidopsis, LcARF6/8 were collectively targeted by Lc-miR167. Interestingly, even though it has been reported that LcARF8B is targeted by Lc-miR167 (Ma et al., 2018), LcARF8A was a novel target by Lc-miR167. Notably, in another group, a novel miRNA-ARF pathway was discovered in litchi, in which LcARF9A/B was targeted by Lc-miRN43. In fact, miRN43 is an innovative miRNA as well, for it is unable to find any ortholog in the database of miRase, which may provide a new interaction for us to study for the specific functions within litchi.
The expression profiles can help us screen for candidate LcARF genes with potentially distinct functions. Most LcARFs were higher accumulated in ML than YL including LcARF2, and its Arabidopsis homolog, AtARF2, which has been reported to regulate leaf senescence with high expression in ML (Ellis et al., 2005). Thus, we can deduce that LcARF2 is likely to be involved in leaf senescence in litchi. It has been reported that Arabidopsis ARF genes regulate flower formation, especially gametophyte development, which are critical for sexual reproduction. AtARF2–AtARF4 and AtARF5 play significant roles in regulating both female and male gametophyte development in Arabidopsis (Liu et al., 2017) and ARF6 and ARF8 regulate both stamen and gynoecium maturation (Nagpal, 2005; Ru et al., 2006; Wu, Tian & Reed, 2006; Liu et al., 2014b). Thus their litchi homologs LcARF2B, 3A, 4A/B, 5A/B, 6D, and 8A may be important for female and male gametophyte development in litchi, along with significantly higher accumulation in female or MF. Moreover, LcARF4, 5 and 9 were prominently higher expressed in FF than MF, implying their potential function in ovule and ovary development.
Auxin response factor genes have been reported to be involved in plant organ abscission. Overexpression of SlARF2 in tomato results in flower organ senescence (Ren et al., 2017) and SlARF1, 2, 7, 11, and 19 showed overlapping functions in tomato abscission (Guan et al., 2014). Similar roles of ARFs in abscission were observed in Arabidopsis (Ellis et al., 2005). Our previous studies show that the treatment of GPD in litchi could reduce the transcript level of auxin response factor (LcARF1) mRNA, along with the increase of fruitlet abscission (Kuang et al., 2012). Here in our RNA-seq survey of ARF gene expression, we found that most of the ARF genes show an opposite correlation with the fruit abscission, that is, ARF genes were down-regulated by abscission induced treatments (ETH and GPD), but up-regulated by GPDD, in which the abscission of GPD was inhibited by the addition of 2, 4-D. This suggested that the majority of LcARF genes were negatively involved in litchi fruit abscission, especially LcARFs from group 3 (ARF2D/2E, 7A/7B, 9A/9B, 16A/16B), with more prominent expression after GPDD treatment. By contrast, a few LcARFs, such LcARF5A/5B, showed positive correlation with fruit abscission, indicating that they might serve as contributing factors to fruit shedding.
Conclusion
In this study, a total of 39 ARF genes were identified from the litchi genome. Comprehensive analyses, including phylogenetic relationship, exon–intron structure, conserved domain, and potential targets for small RNAs, revealed that the ARF gene family was expanded in litchi with species-specific features. A novel miRNA-ARF (miRN43-ARF9) regulatory pathway was discovered, which is likely specific in litchi. Expression profiles in various organs and under different abscission-related treatments (ETH, GPD, and GPDD) uncovered the expression diversity of these ARF genes in litchi. Some ARF genes, including ARF2D/2E, 7A/7B, 9A/9B, 16A/16B, and 5A/5B, likely play predominant roles in the process of litchi fruit abscission. These findings provide new knowledge and resources for further functional characterization of ARF genes in litchi.
Supplemental Information
Specific primers of 39 LcARF genes used for quantitative RT-PCR in this study.
Potential functions of LcARFs.
YF, young fruit (25 days after fertilization); FF, female flower; MF, male flower; USF, undeterminated sex flowers; ML, mature leaves; YL, young leaves; FBS, fruit-bearing shoots. The red font represents the genes in group1, group2, and group3 in Fig. 5.