
‘Rolling’ stoneflies (Insecta: Plecoptera) from mid-Cretaceous Burmese amber
- Published
- Accepted
- Received
- Academic Editor
- Joseph Gillespie
- Subject Areas
- Entomology, Paleontology, Taxonomy, Freshwater Biology
- Keywords
- Paleontology, Cretaceous, Amber, Plecoptera, Stoneflies, Burmese, New family, New genus, New species, Rolling Stones
- Copyright
- © 2018 Sroka et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. ‘Rolling’ stoneflies (Insecta: Plecoptera) from mid-Cretaceous Burmese amber. PeerJ 6:e5354 https://doi.org/10.7717/peerj.5354
Abstract
This contribution describes seven new species of fossil stoneflies from Cretaceous Burmese amber, all of which are dedicated to present and past members of the Rolling Stones. Two species—Petroperla mickjaggeri gen. nov. sp. nov. and Lapisperla keithrichardsi gen. nov. sp. nov.—are placed in a new family Petroperlidae within the stemline of Systellognatha. The first Cretaceous larval specimen of Acroneuriinae, Electroneuria ronwoodi gen. nov. sp. nov., is also described along with another four new species that are placed within the Acroneuriinae genus Largusoperla Chen et al., 2018: Largusoperla charliewattsi sp. nov., Largusoperla brianjonesi sp. nov., Largusoperla micktaylori sp. nov., and Largusoperla billwymani sp. nov. Additional specimens of Acroneuriinae are described without formal assignment to new species due to insufficient preservation. Implications for stonefly phylogeny and palaeobiogeography are discussed.
Introduction
Stoneflies (Plecoptera) is a small insect order with approximately 3,700 extant and 250 fossil species currently described (DeWalt et al., 2018). Extant stonefly diversity is highest in the temperate regions of the Northern Hemisphere (Fochetti & De Figueroa, 2008), a pattern probably consistently followed throughout their history (Sinitshenkova, 1997). Stoneflies in general have aquatic larvae and usually prefer cold streams with relatively high amount of dissolved oxygen (Zwick, 1973; Hynes, 1976), although there are both extant and extinct Jurassic taxa known from hypotrophic lakes (Brittain, 1990; Sinitshenkova & Zherikin, 1996).
Plecoptera is considered either as sister group to all other Neoptera or as one of the basal clades within Polyneoptera (Zwick, 2009; Misof et al., 2014; Wang et al., 2016). Phylogeny of individual lineages within the order was analysed by Zwick (1973, 1980). Sinitshenkova (1987) presented a phylogeny that also included fossil taxa. However, partly due to the limited number of characters visible in some fossil taxa, her topology is not supported by reliable apomorphies of individual clades. The most recent and widely accepted phylogeny of Plecoptera was published by Zwick (2000) based on morphological characters of extant species only.
The first molecular phylogeny was published by Thomas et al. (2000) using 18S gene only. A more complex study using six molecular markers (12S, 16S, 18S, 28S, COII, H3) and morphological data was performed by Terry (2003). Results of this study were largely congruent with Zwick (2000), recovering all major groups, i.e. Antarctoperlaria, Arctoperlaria, Systellognatha, Euholognatha (without Megaleuctra), and Perloidea as monophyletic. Recently, Chen et al. (2018) proposed a molecular phylogeny of Systellognatha inferred from mitochondrial genome sequences.
Stoneflies are relatively rare in the fossil record, which might be related to their delicate bodies and their preferred lotic habitat preventing fossilization. Stoneflies date back to the Carboniferous, with the alleged stem-Plecoptera Gulou carpenteri Béthoux et al., 2011 from the Pennsylvanian of China (Béthoux et al., 2011), although there is only indirect evidence for its placement within Plecoptera. Numerous stonefly fossils attributed to the crown group are known from the Permian, with the diversity culminating in the Jurassic and declining in the Early Cretaceous (Sinitshenkova, 1997; Liu & Ren, 2006).
The best-preserved fossil specimens are found in amber, allowing the observation of even minute morphological details. However, amber inclusions of stoneflies are in general very rare. Only one species attributed to the Perlidae subfamily Acroneuriinae (Stark & Lenz, 1992) was described from Miocene Dominican amber. Three species of Capniidae and Leuctridae were recognized in Rovno amber (Sinitshenkova, 2009), and 19 species out of five families (Taeniopterygidae, Leuctridae, Nemouridae, Perlidae, Perlodidae) are described from Eocene Baltic amber, all but two of which are attributed to extant genera (Caruso & Wichard, 2010, 2011; Chen, 2018a). The occurrence of Plecoptera in Upper Cretaceous Siberian amber is also known (Wichard & Weitschat, 1996). Only recently, Chen, Wang & Du (2018) and Chen (2018b, 2018c) reported the first Plecoptera from mid-Cretaceous Burmese amber with the descriptions of the new genera Largusoperla Chen et al., 2018 (Perlidae: Acroneuriinae) including four species, and Pinguisoperla Chen, 2018, including a single species Pinguisoperla yangzhouensis.
In this study, we add four new species of Largusoperla and describe the first larval specimen of stoneflies in Burmese amber. More importantly, we report two new genera within a new family of fossil stoneflies, which probably represent stem group representatives of Systellognatha. We discuss their systematic affinities and evaluate possible implications for Plecoptera phylogeny. We also provide novel insights into several aspects of the palaeocology and palaeobiogeography of stoneflies.
Material and Methods
Specimens
All pieces of fossil-bearing Burmese amber originate from the type locality Hukawng Valley, Kachin State, Myanmar. The exact outcrop among the various amber mines in this valley is unknown, because the specimens were acquired from traders. A review of these amber deposits and relative geological history is available in Zherikhin & Ross (2000), Grimaldi, Engel & Nascimbene (2002), and Ross et al. (2010). UePb zircon dating (Shi et al., 2012) constrained this amber to a maximum age of 98.79 ± 0.62 Ma, which is equivalent to the earliest Cenomanian (Gradstein, Ogg & Smith, 2004).
Imaging
The material was studied under a Leica M205 C (Leica Corporation, Wetzlar, Germany) and Olympus SZX7 (Olympus Corporation, Tokyo, Japan) stereomicroscope. Leica Z16 APO Macroscope with Leica Application Suite Version 3.1.8 and Helicon Focus Pro was used to obtain stacked photographs with extended depth of field. Photographs were sharpened and adjusted in contrast and tonality in Adobe Photoshop™ version CS6 (Adobe Systems Incorporated, San Jose, CA, USA).
High-resolution μ-CT scanning with a Bruker Skyscan 1272 tomograph was performed on specimen SMNS BU-79, but yielded no sufficient contrast to produce any useful tomographic image.
Terminology
Abbreviations for wing veins used throughout the text follow Béthoux (2005): C, costa; ScP, subcosta posterior; R, radius; RA, radius anterior; RP, radius posterior; M, media; MA, media anterior; MP, media posterior; Cu, cubitus; CuA, cubitus anterior; CuP, cubitus posterior; AA, analis anterior; arc, arculus (secondarily strenghtened cross vein between M and Cu).
All type specimens examined are housed in the State Museum of Natural History, Stuttgart, Germany (SMNS) and catalogue numbers are specified for individual species below.
When discussing the affinities of individual taxa, we refer to the phylogenetic system of Plecoptera proposed by Zwick (2000).
Nomenclature
The electronic version of this article in portable document format will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub:486E9A01-EF59-41D7-B001-4AD6D7FBB11F. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central, and CLOCKSS.
Etymology of ‘Rolling’ stoneflies
Burmese amber is one of the oldest resins with insect inclusions, and stoneflies are one of the oldest pterygote lineages. What lies closer at hand than to link fossil stoneflies in ancient stones with the Rolling Stones and to name the new species after the members of the oldest and greatest Rock ‘n’ Roll Band in the world. The discerning reader will notice that the new family and genera are named after ‘the Stones,’ and all present and former members of the Rolling Stones are honoured with their own species.
Systematic Palaeontology
Class Insecta Linnaeus, 1758
Subclass Pterygota Lang, 1888
Order Plecoptera Burmeister, 1839
Suborder Arctoperlaria Zwick, 1973
Infraorder Systellognatha Zwick, 2000
Petroperlidae, fam. nov.
urn:lsid:zoobank.org:act:3F9EB209-A5DD-49C9-A5D6-7952F05F94A4
Type genus. Petroperla gen. nov.
Diagnosis. Glossae and paraglossae of approximately same size (plesiomorphy of Systellognatha), rather stout labial palps (plesiomorphy of Systellognatha), short first tarsomere (apical tarsomere 3.5× longer than first tarsomere) (apomorphy of Systellognatha); euplantulae present on tarsi (plesiomorphy of Systellognatha); setose arolium (apomorphy of Systellognatha); forewing with numerous crossveins in costal field (plesiomorphy of Systellognatha); vein RA almost reaching wing apex (apomorphy of Petroperlidae), proximal origin of vein RP (just distal to 1/4 of wing length) (plesiomorphy of Systellognatha).
Petroperla gen. nov.
urn:lsid:zoobank.org:act:68557BD3-F4E1-43BB-A10B-68D796971769
Type species. Petroperla mickjaggeri, gen. et sp. nov.
Diagnosis. By monotypy, as for the type species.
Etymology. The first part of the compound noun refers to the Rolling Stones and is derived from Latin ‘petra,’ meaning ‘stone,’ the second part ‘perla,’ refers to the stonefly genus Perla.
urn:lsid:zoobank.org:act:838EDFF8-BD85-4F83-88F6-87E38701A941

Figure 1: Petroperla mickjaggeri gen. nov. sp. nov., holotype SMNS BU-79, photographs.
(A) Dorsal view. (B) Ventral view. (C) Head in frontoventral view. Abbreviations: gl, glossa; pgl, paraglossa; pmx, maxillary palp; plb, labial palp. (D) Ventral tip of abdomen with subgenital plate: sgp.{kind=link}
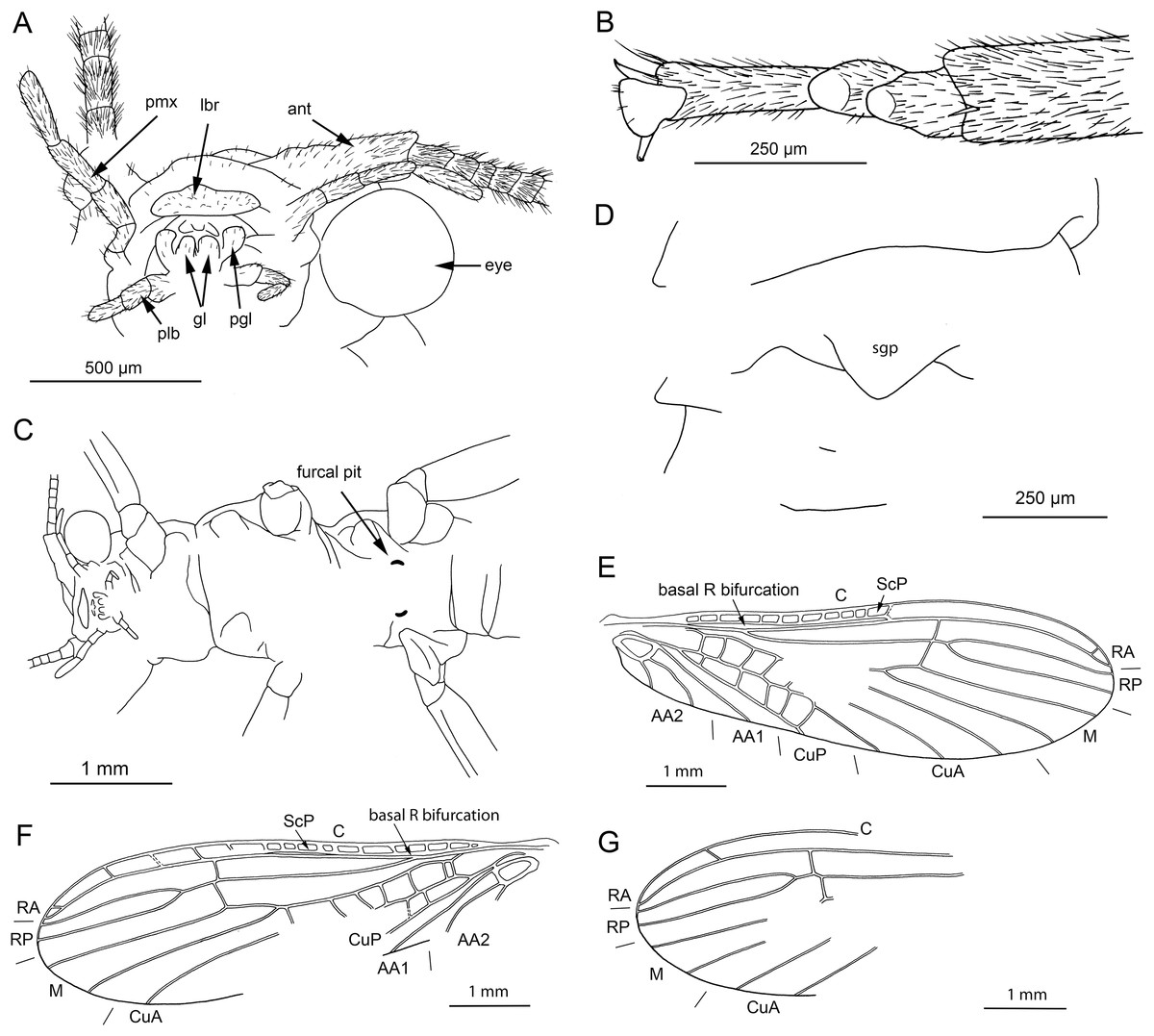
Figure 2: Petroperla mickjaggeri gen. nov. sp. nov., holotype SMNS BU-79, line drawings.
(A) Head in frontoventral view. Abbreviations: ant, antenna; gl, glossa; lbr, labrum; pgl, paraglossa; pmx, maxillary palp; plb, labial palp. (B) Tarsus in ventral view. (C) Head and thorax in ventral view. (D) Subgenital plate; sgp. (E) Right forewing. (F) Left forewing. (G) Left hind wing.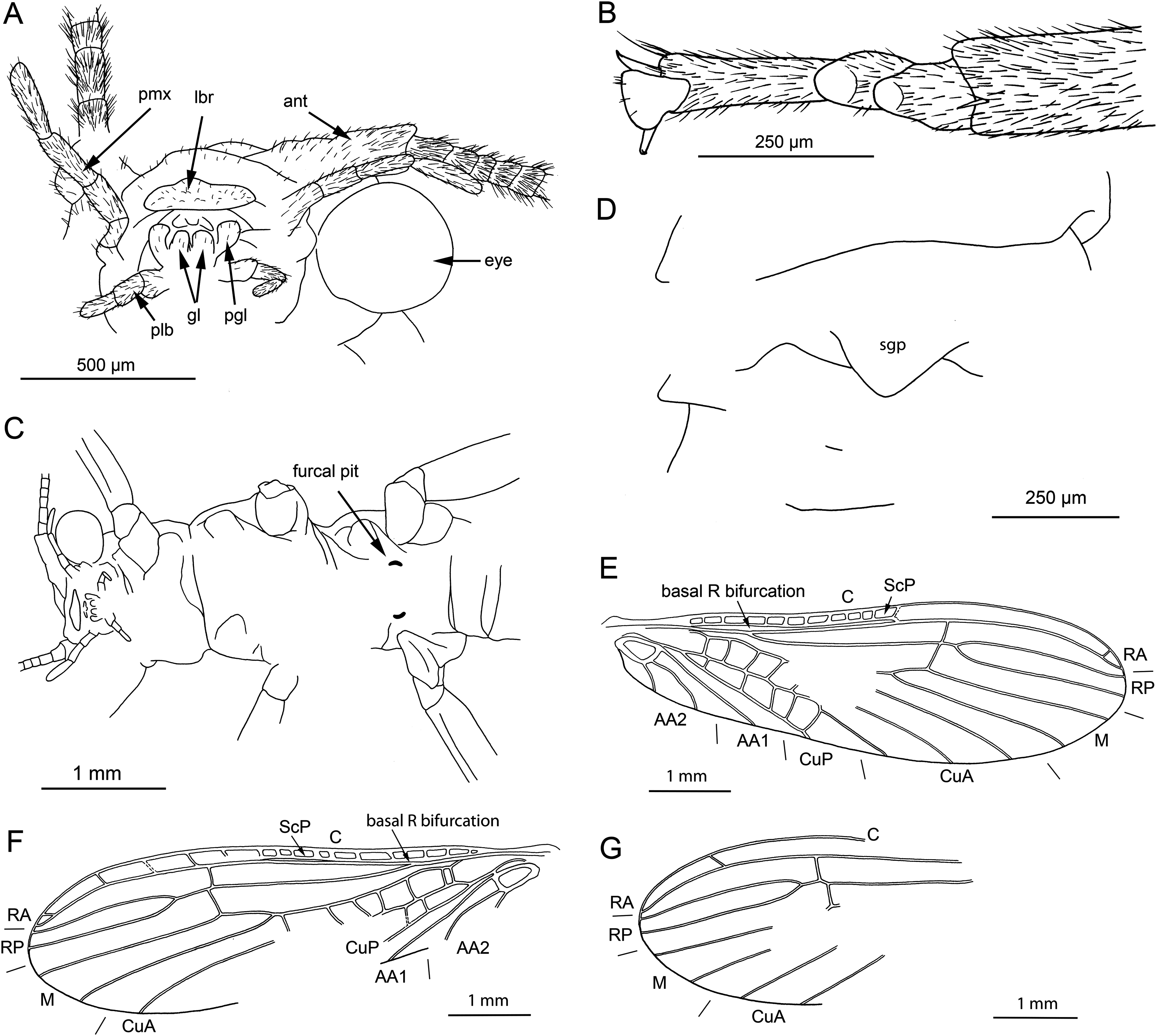{kind=link}
Diagnosis. Thoracic gill remnants absent, abdominal segments not extended posterolaterally.
Etymology. The name refers to Sir Mick Jagger, founding member, harmonica player, and lead singer of the Rolling Stones.
Material. Holotype specimen: SMNS BU-79, female.
Description.
Body length 6.2 mm.
Head. Colour greyish, original cuticular colouration not preserved. All three ocelli fully developed. Median ocellus slightly subequal in size to lateral ocelli. Antennae 3.6 mm long (approximately 0.6× body length). Individual segments covered with short hair-like setae. Antennal segments in middle third of antenna approximately 2.2× longer than wide. Maxillary palps five-segmented, two basal palpomeres short, three distal palpomeres longer, of approximately same length. Labium with glossae and paraglossae of approximately same size, labial palps rather stout, three-segmented (Figs. 1C and 2A). Two basal palpomeres short, apical palpomere longer. Other mouthparts not observable in detail.
Thorax. Prothorax approximately quadrangular. Colour greyish, original cuticular colouration not preserved (Figs. 1A and 1B).
Forewings (Figs. 2E and 2F): length 6.5 mm, width 2 mm; costal field with numerous crossveins (12 crossveins visible on right forewing and 17 on left forewing, including area distal to ScP, badly visible due to preservation); ScP reaching RA just distally to 1/2 of wing length; RA simple, almost reaching wing apex; single crossvein between RA and RP; RP originating from R just distally to 1/4 of wing length; RP with two distal branches, originating at 2/3 of wing length; single crossvein between RP and M; M approximated to R basally, diverging close to origin of RP; M with two branches, originating just distally to 1/2 of wing length; occurrence of arculus and up to six additional crossveins between M and CuA; CuA branched distally, with three branches; up to five crossveins between CuA and CuP; CuP simple and straight; AA1 simple; AA2 with two branches.
Hind wings (Fig. 2G, venation visible only in apical portion): length 5.6 mm; RA simple, almost reaching wing apex; single crossvein between RA and RP; RP with two distal branches, originating at 2/3 of wing length; single crossvein between RP and M; M with two branches; branching of Cu and AA not recognizable.
Thoracic sterna (Fig. 2C) with one pair of dark patches centrally on mesosternum and two pairs on metasternum. Metasternum (Fig. 2C) with apparent arched furcal pits (not apparent on pro- and mesosternum due to state of preservation). Thoracic gill remnants absent.
Legs slender, covered with very short spine-like setae (thicker setae near posterior margin of femora and on dorsal surface of tibiae. Single tibial spur present apically (Fig. 2B). Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.5× longer than first two combined (Fig. 2B). Arolium present, approximately as wide as width of apical tarsomere, with setae on arolium. Euplantulae present on tarsomeres 1 and 2.
Abdomen. Colour greyish, original colouration not preserved. Individual segments not distinctly enlarged posteromedially. No abdominal gill remnants recognizable. Subgenital plate poorly visible, probably triangular, without notch medially (Figs. 1D and 2D). Cerci short, 1 mm in length, covered with very short setae (Fig. 1A).
Lapisperla gen. nov.
urn:lsid:zoobank.org:act:05FAF13D-5548-4EFB-AFA1-CA30EA1A0B7B
Type species. Lapisperla keithrichardsi, gen. et sp. nov.
Diagnosis. By monotypy, as for the type species.
Etymology. The first part of the name refers to the Rolling Stones and is derived from Latin ‘lapis,’ meaning ‘stone,’ the suffix ‘perla’ refers to the stonefly genus Perla.
urn:lsid:zoobank.org:act:586DF169-34E5-4E6F-89EC-58E7D67BF6CB

Figure 3: Lapisperla keithrichardsi gen. nov. sp. nov., holotype SMNS BU-313, photographs.
(A) Dorsal view. (B) Ventral view. (C) Abdomen in ventral view. (D) Tarsus of right middle leg in lateral view. Abbreviations: ar, arolium; eu, euplantula.{kind=link}
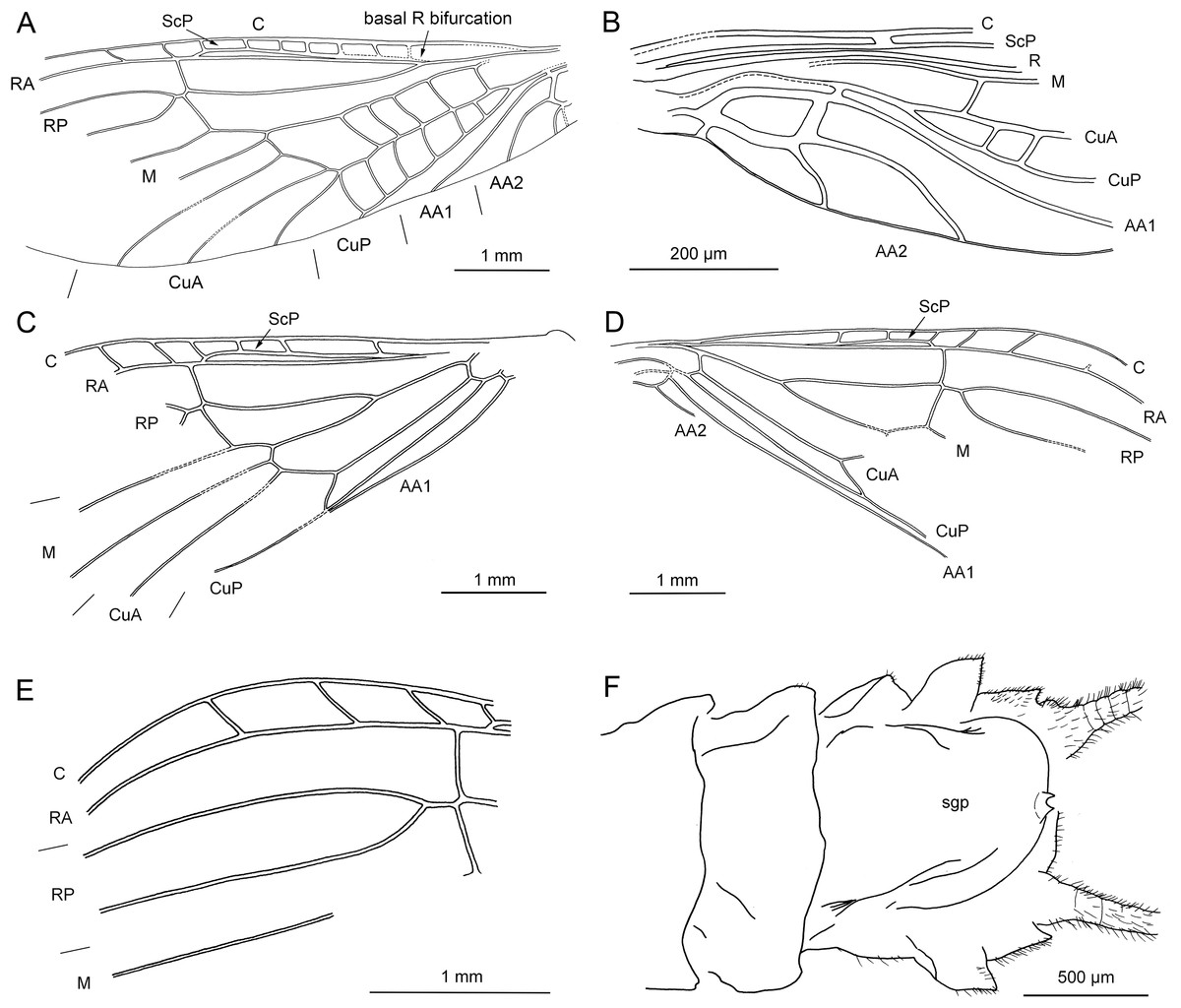
Figure 4: Lapisperla keithrichardsi gen. nov. sp. nov., holotype SMNS BU-313, line drawings.
(A) Left forewing. (B) Right forewing base. (C) Left hind wing. (D) Right hind wing. (E) Right hind wing from ventral side. (F) Tip of ventral abdomen with subgenital plate, sgp.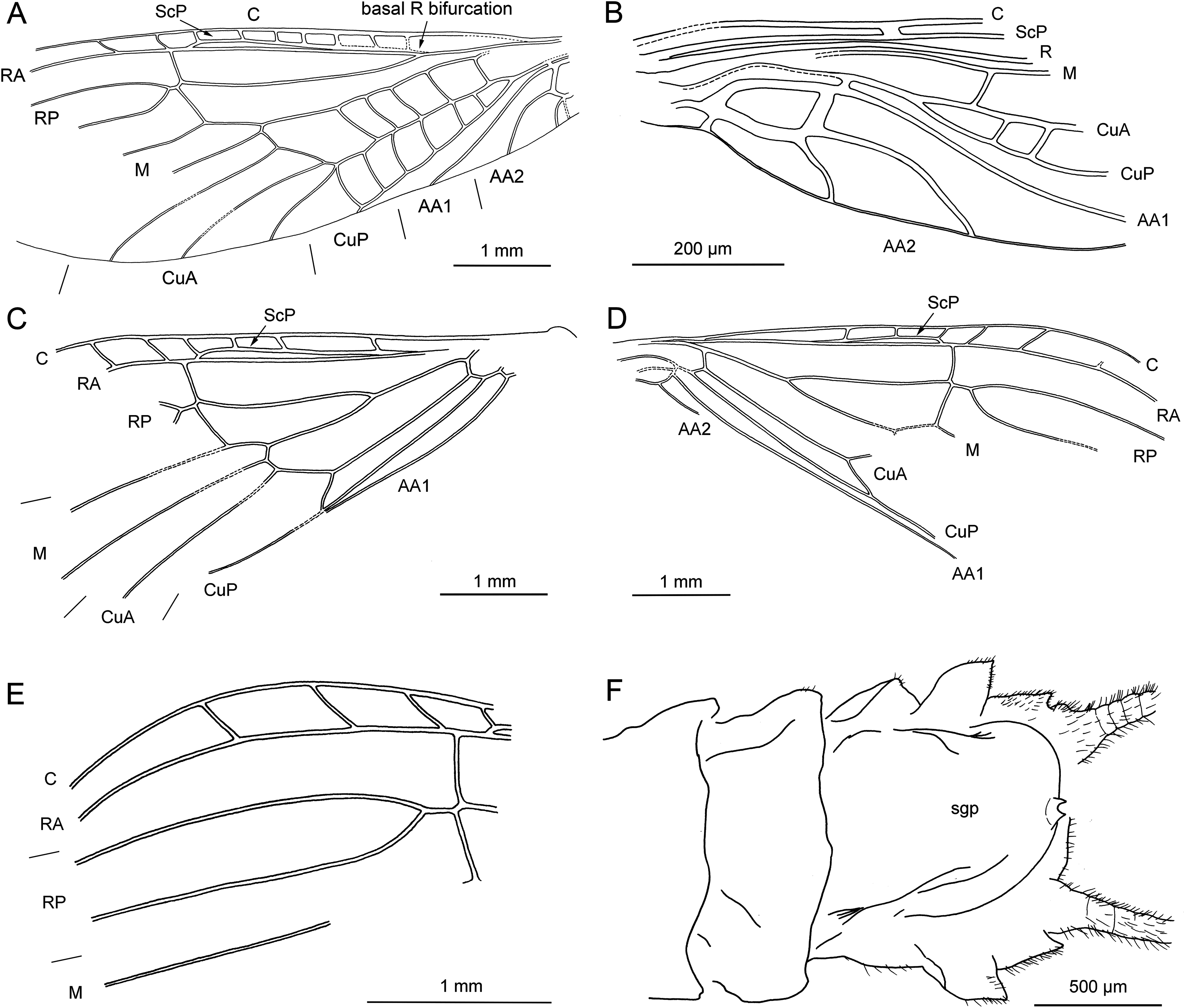{kind=link}
Diagnosis. Prominent subgenital plate, abdominal segments extended posterolaterally, thoracic gill remnants present.
Etymology. The name refers to Keith Richards, founding member and guitar player of the Rolling Stones, master of the ancient art of weaving.
Material. Holotype specimen: SMNS BU-313, female.
Description.
Body length 6.1 mm (without head).
Head. Not preserved.
Thorax. Prothorax approximately quadrangular (Figs. 3A and 3B). Original cuticular colouration not preserved.
Forewings (Figs. 4A and 4B): right forewing present only as short basal fragment, 2.3 mm long. Left forewing almost complete, except for apical portion, length of preserved part approximately 6 mm, width 2.4 mm; costal field with nine visible crossveins, including area distal to ScP with two visible crossveins; ScP reaching RA just proximally to RA–RP crossvein; RA simple; single crossvein between RA and RP; RP originating from R relatively basally, approximately 1.8 mm from wing base; RP with two visible distal branches, originating just distally to RA–RP crossvein; single crossvein between RP and M; M approximated to R basally; M with two visible branches, originating approximately 3.6 mm from wing base; occurrence of arculus and five additional crossveins between M and CuA; CuA branched distally, with three branches; six crossveins between CuA and CuP; CuP simple and straight; AA1 simple; AA2 with three branches.
Hind wings (Figs. 4C–4E): apical portions missing, length of preserved parts approximately 5.7 mm (right hind wing) and 4.9 (left hind wing); costal field with six visible crossveins, including area distal to ScP with three crossveins; ScP reaching RA just proximally to RA–RP crossvein; RA simple, close to apex not approximated to C; single crossvein between RA and RP; RP with two branches, originating just distally to RA–RP crossvein; single crossvein between RP and M; M with two branches; CuA, CuP, and AA1 simple. Venation pattern of AA2 not recognizable.
Visible gill remnants ventrolateral between meso- and metathorax. Legs slender, covered with very short spine-like setae. Single apical tibial spur present (Fig. 3D). Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.6× longer than first two combined (Fig. 3D). Long hair-like setae near base of claws. Arolium present, approximately as wide as width of apical tarsomere. Arolium equipped with setae. Euplantulae present apically on tarsomeres 1 and 2 (Fig. 3D).
Abdomen. Individual segments with pronounced posterolateral extensions (Figs. 3C and 4F). No abdominal gill remnants recognizable. Prominent subgenital plate, rounded posteriorly, with slightly elevated notch posteromedially (Figs. 3C and 4F). Cerci broken off, only short basal parts preserved, densely covered with short setae.
Remarks on generic composition of Petroperlidae. Lapisperla keithrichardsi sp. nov. differs from Petroperla mickjaggeri sp. nov. in having a more prominent subgenital plate (badly visible, but much smaller in Petroperla mickjaggeri sp. nov.). Abdominal segments of Lapisperla keithrichardsi sp. nov. are equipped with posterolateral projections (no projections in Petroperla mickjaggeri sp. nov.), and meso- and metathorax of Lapisperla keithrichardsi sp. nov. ventrolaterally with gill remnants (apparently absent in Petroperla mickjaggeri sp. nov.). We consider these differences pronounced enough to justify a placement of these species in two separate genera within Petroperlidae.
Order Plecoptera Burmeister, 1839
Suborder Arctoperlaria Zwick, 1973
Infraorder Systellognatha Zwick, 2000
Superfamily Perloidea Latreille, 1802
Family Perlidae Latreille, 1802
Subfamily Acroneuriinae Klapálek, 1914
Electroneuria, gen. nov.
urn:lsid:zoobank.org:act:3A4CA23C-2D23-4F8E-9492-93C490AE736C
Type species. Electroneuria ronwoodi, gen. et sp. nov.
Diagnosis. By monotypy, as for the type species.
Etymology. Latin ‘electrum’ refers to both amber and electric guitars, the suffix ‘–neuria’ to the stonefly subfamily Acroneuriinae.
urn:lsid:zoobank.org:act:A52885BB-E2CF-49E8-871B-C4CE8577002C

Figure 5: Electroneuria ronwoodi gen. nov. sp. nov., holotype SMNS BU-306, photographs.
(A) Dorsal view. (B) Ventral view. (C) Head with exposed maxilla, mx. (D) Head, occipital row of spines, marked by arrows. (E) Tarsus of left foreleg. (F) Setae on abdominal terga.{kind=link}
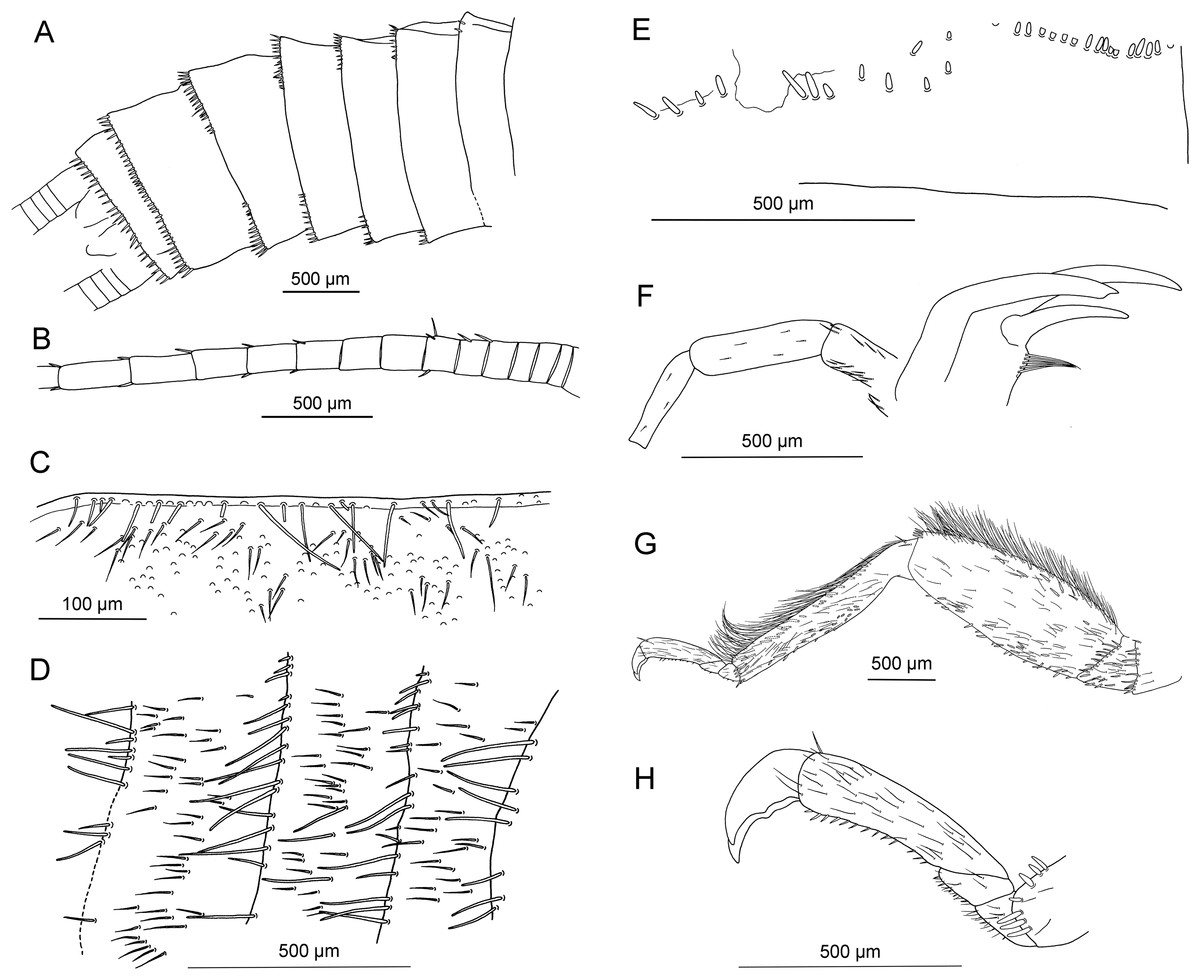
Figure 6: Electroneuria ronwoodi gen. nov. sp. nov., holotype SMNS BU-306, line drawings.
(A) Abdominal sterna. (B) Detail of right cercus. (C) Anterior margin of pronotum. (D) Setation of abdominal terga. (E) Head, occipital row of spines. (F) Maxilla. (G) Right middle leg. (H) Tarsus of right middle leg.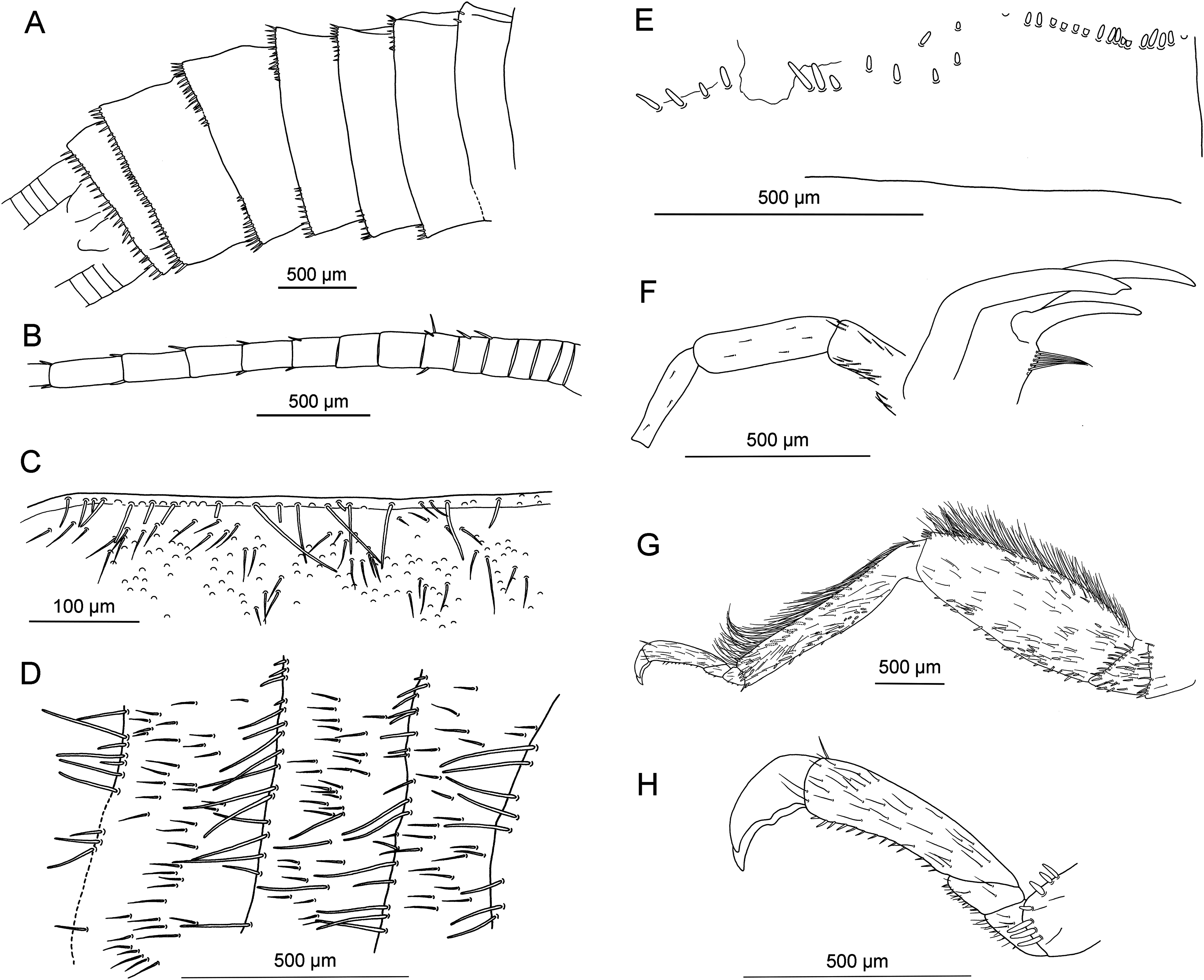{kind=link}
Diagnosis. Larva with occipital spinule row complete medially; fringe of long thin setae laterally on pronotum; long hair-like setae on surface of wingpads and abdominal terga; posterior margin of abdominal terga with numerous very long thin setae; cerci long, with only short setae.
Etymology. The name of this immature specimen refers to Ronnie Wood, guitar player of the Rolling Stones since 1975, and youngest member of the Rolling Stones.
Material. Holotype specimen SMNS BU-306, larva.
Description. Body length 8.1 mm (Figs. 5A and 5B).
Head. Antennae 5.5 mm long (approximately 0.7× body length). Length of antennal segments in the middle third of antenna approximately equal to segment width. Mouthparts of predaceous type. Mandibles with only apical part of right mandible recognizable, with four rounded teeth. Left maxilla well visible (Figs. 5C and 6F), with thin galea slightly shorter than lacinia. Lacinia with two long, prominent pointed teeth, apical tooth longer. Row of six long setae on inner margin of lacinia, situated basally from subapical tooth, on slightly elevated hump. Left maxillary palp with three recognizable elongated palpomeres, basal part not well visible. Distal palpomere probably missing. Sparse short setae present on all visible palpomeres, more dense on proximal palpomere. Other mouthparts not recognizable.
Occipital row of short spinules medially regular, in lateral parts scattered goups of longer spinules present (Figs. 5D and 6E).
Thorax. Pronotum covered with minute, hair-like setae on surface. Longer setae along margins (Fig. 6C). Pronounced wingpads (same size on meso- and metathorax), posterior notal contour not apparent. Thoracic gills badly preserved, indistinctly visible on left side between meso- and metathorax. Legs covered with numerous setae of various size and shape (Fig. 6G). Regular row of long, hair-like setae along outer margin of femora (length approximately 0.5× femur width) and tibiae (length approximately 1.5× tibia width). Tarsi with first two tarsomeres very short, third one approximately 2× longer than first two combined. Two claws with indistinctly pronounced denticles (Figs. 5E and 6H).
Abdomen. Posterior margin of terga with long setae (posterior tergal spinule fringe sensu Stark & Gaufin, 1976). Numerous long, intercalary setae on surface of terga (Figs. 5F and 6D). Sterna with short spine-like setae along posterior margin; row of these setae complete in two last segments (Fig. 6A). No abdominal gill remnants recognizable. Paraprocts bluntly pointed apically. Cerci 6 mm long (approximately 0.7× body length). Individual segments with short, spine-like setae (Fig. 6B).
Order Plecoptera Burmeister, 1839
Suborder Arctoperlaria Zwick, 1973
Infraorder Systellognatha Zwick, 2000
Superfamily Perloidea Latreille, 1802
Family Perlidae Latreille, 1802
Subfamily Acroneuriinae Klapálek, 1914
Genus Largusoperla Chen et al., 2018
urn:lsid:zoobank.org:act:43BA0BA8-7818-47FE-BA37-E38F140A457C

Figure 7: Largusoperla charliewattsi sp. nov., holotype SMNS BU-10, photographs.
(A) Dorsal view, arrow marks rounded pronotum. (B) Head in ventral view. (C) Meso- and metathorax in ventral view with furcal pits, fp. (D) Ventral tip of abdomen with hammer, paraprocts, pp, and cercus, ce. (E) Tarsus of right hind leg. (F) Tarsus of left hind leg with euplantulae, eu.{kind=link}
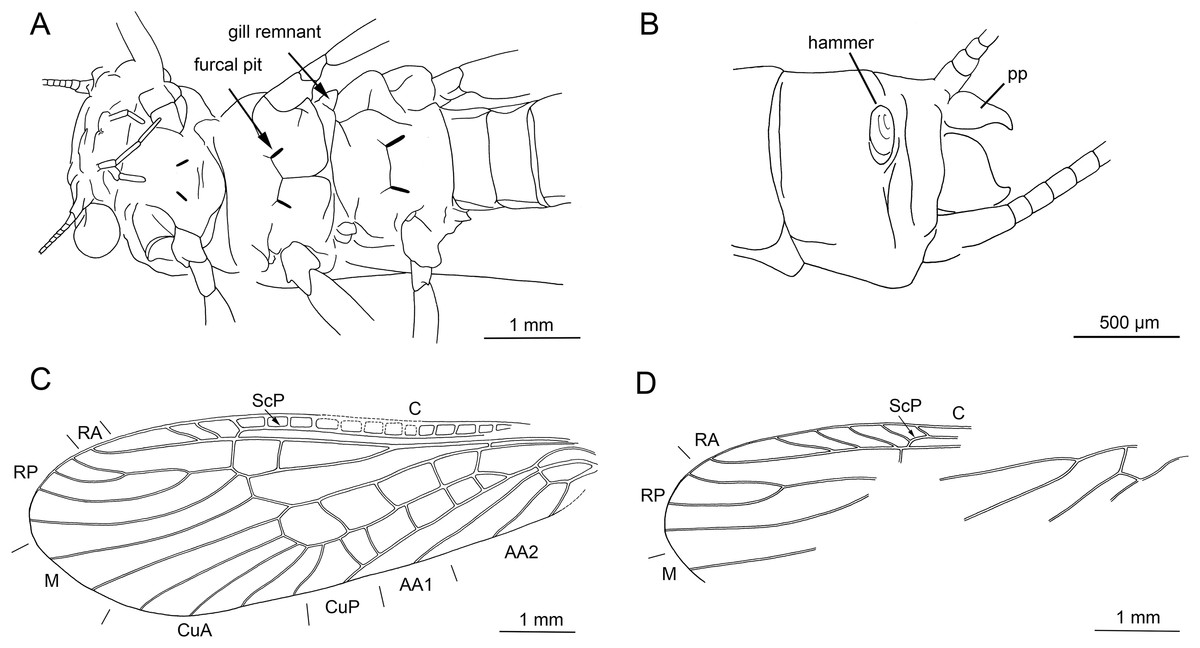
Figure 8: Largusoperla charliewattsi sp. nov., holotype SMNS BU-10, line drawings.
(A) Head and thorax in ventral view with furcal pits and gill remnants. (B) Ventral tip of abdomen with hammer and paraprocts, pp. (C) Left forewing. (D) Left hind wing.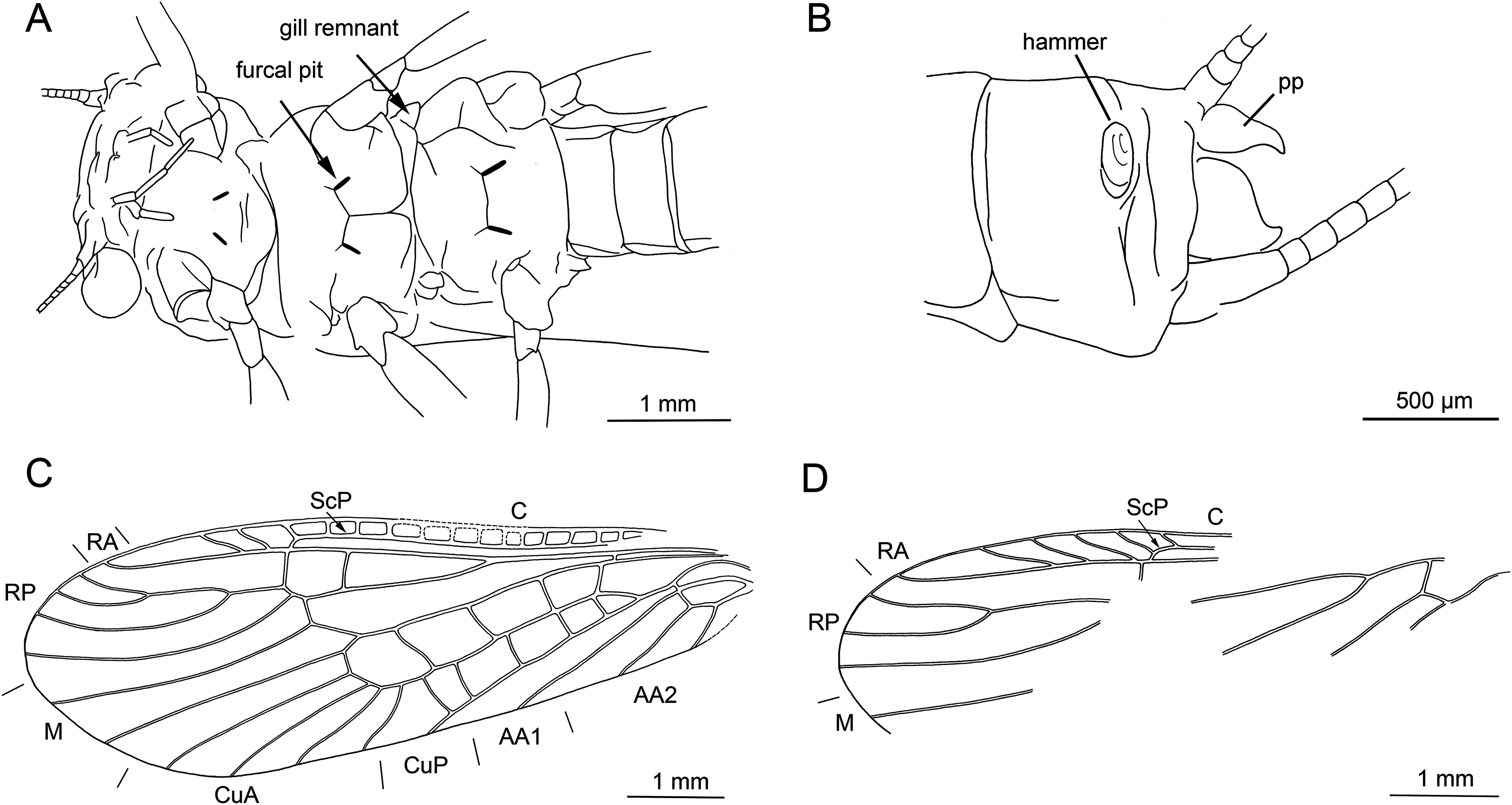{kind=link}
Diagnosis. Large paraprocts; hammer knob-shaped, elongated transversally; pronotum not distinctly widened anteriorly.
Etymology. The name refers to Charlie Watts, drummer of the Rolling Stones, which is most adequate in regard of the pronounced drumming apparatus of the new species.
Material. Holotype specimen: SMNS BU-10, male.
Description. Body length 8.1 mm (Fig. 7A).
Head. Colour brown, marginal areas paler. Three fully developed ocelli. Median ocellus slightly subequal in size to lateral ocelli. Antennae 5.4 mm long (approximately 0.7× body length). Individual segments covered with short hair-like setae. Antennal segments in the middle third of antenna length approximately 2.5× longer than wide.
Maxillary palps slender, five elongated palpomeres covered with short hair-like setae (Fig. 7B). Labial palps rather elongated, three palpomeres present. First palpomere short (Fig. 7B). Other mouthparts not recognizable.
Thorax. Prothorax approximately quadrangular. Colour brown with pale median longitudinal line.
Forewings (Fig. 8C): length 8 mm, width 2.6 mm; costal field with 15 visible crossveins (including two crossveins distal to ScP); ScP reaching RA just proximally to 2/3 of wing length; RA simple; two crossveins between RA and RP; RP originating from R just distally to 1/3 of wing length; RP with four distal branches; single crossvein between RP and M; M slightly approximated to R basally; M with two branches, originating just distally to 1/2 of wing length; occurrence of arculus and four additional crossveins between M and CuA; CuA branched distally, with four branches; five crossveins between CuA and CuP; CuP simple and straight; single cubito-anal crossvein reaching anal cell; A1 simple; AA2 with two branches, originating directly from anal cell.
Hind wings (Fig. 8D, venation details observable only in apical portion): length 6.9 mm; costal field with five visible crossveins (including three crossveins distal to ScP); ScP reaching RA distal to 1/2 of wing length; RA simple; single crossvein between RA and RP; RP originating from common stem with M at 1/4 of wing length; RP with three distal branches; branching of M, Cu and AA not recognizable.
Thoracic sterna (Figs. 7C and 8A) with apparent oblique furcal pits, converging anteriorly. Transverse meso- and metasternal ridge connecting anterior corners of respective furcal pits. Longitudinal median ridge apparent on mesofurcasternum. Y-ridge connecting posterior corners of furcal pits absent. Gill remnants recognizable on all thoracic segments (Fig. 8A).
Legs slender, covered with very short hair-like setae. Two thickened tibial spurs present apically on tibia, along with several additional long spine-like setae (Fig. 7E). Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.5× longer than first two combined. Arolium present, slightly wider than width of apical tarsomere. Euplantulae present on tarsomeres 2 and 3 (Figs. 7E and 7F).
Abdomen. Colour brown, ventral side paler. Individual segments not distinctly enlarged posteromedially. Elevated, knob-shaped and transversally elongated hammer near posterior margin of sternum IX (Figs. 7D and 8B). Enlarged dark brown hook-like paraprocts between cerci (Figs. 7D and 8B). Cerci short, 2.1 mm in length (approximately 0.3× body length). Slightly moniliform in shape; segments covered with short hair-like setae (Fig. 7D).
Affinities. Largusoperla charliewattsi sp. nov. exhibits a combination of morphological characters allowing attribution to the family Perlidae (presence of long and slender palps, vestiges of thoracic gills, euplantulae, short first tarsomere, and forewings with numerous crossveins in the basal half of the costal field, see Zwick, 1980, 2000). The presence of sclerotized and strongly recurved paraprocts and well-developed hammer allow a placement into the subfamily Acroneuriinae (Stark & Gaufin, 1976; Zwick, 1980, 2000).
Within Acroneuriinae, Largusoperla charliewattsi sp. nov. can be attributed to the recently described genus Largusoperla, as defined by Chen, Wang & Du (2018), based on the following diagnostic characters: triocellate; pronotum with pale median area; sternum IX with a small elevated lobe; large paraprocts sclerotized and strongly upcurved; short cerci, no longer than 1/2 of the abdomen length. Some characters stated by Chen, Wang & Du (2018) as diagnostic for the genus are not visible in Largusoperla charliewattsi sp. nov., possibly due to the state of preservation (head with dark stigma covering ocelli; abdominal terga with darker lateral markings). Another diagnostic character (abdominal segments posterolaterally extended) is not very pronounced in Largusoperla charliewattsi sp. nov. and is probably variable within the genus.
In Pinguisoperla Chen, 2018, the second Perlidae genus described from Burmese amber, the subfamilial attribution is uncertain (Chen, 2018b), and it might also belong to Acroneuriinae. Nevertheless, any affinity of Largusoperla charliewattsi sp. nov. to Pinguisoperla can be excluded based on the absence of enlarged, plump basal segment of the cercus, which constitutes the main diagnostic character of Pinguisoperla.
Up to now, four species of Largusoperla have been described, all based on male specimens from the Burmese amber, namely Largusoperla acus Chen et al., 2018, Largusoperla flata Chen et al., 2018, Largusoperla arcus Chen et al., 2018, and Largusoperla difformitatem Chen, 2018.
Largusoperla charliewattsi sp. nov. differs from Largusoperla acus in having less prominent posterolateral extensions on abdominal segments, pronotum not distinctly widened anteriorly, and paraprocts without needle-like apices. Largusoperla flata can be discriminated from Largusoperla charliewattsi sp. nov. by having entirely different shape of paraprocts (see Chen, Wang & Du, 2018, figs. 12 and 13; Figs. 7D and 8B). Largusoperla arcus possesses a more circular hammer on sternum IX, compared to a transversally rather elongated hammer in Largusoperla charliewattsi sp. nov. and paraprocts with apices distinctly diverging, contrary to more parallel oriented paraprocts in Largusoperla charliewattsi sp. nov. Largusoperla difformitatem differs by longer, highly divergent paraprocts and smaller hammer.
urn:lsid:zoobank.org:act:4AB77845-FAB5-4440-8496-935CBD1249B0

Figure 9: Largusoperla billwymani sp. nov., holotype SMNS BU-229, photographs.
(A) Dorsal view, arrow marks trapezoid pronotum. (B) Head in ventral view. (C) Ventral tip of abdomen with hammer, paraprocts, pp, and cercus, ce. (D) Tarsus of left hind leg. (E) Tarsus of right hind leg.{kind=link}

Figure 10: Largusoperla billwymani sp. nov., holotype SMNS BU-229, line drawings.
(A) Left forewing. (B) Right forewing. (C) Head and thorax in ventral view. (D) Right hind wing. (E) Head in ventral view. (F) Ventral tip of abdomen with hammer and paraprocts, pp.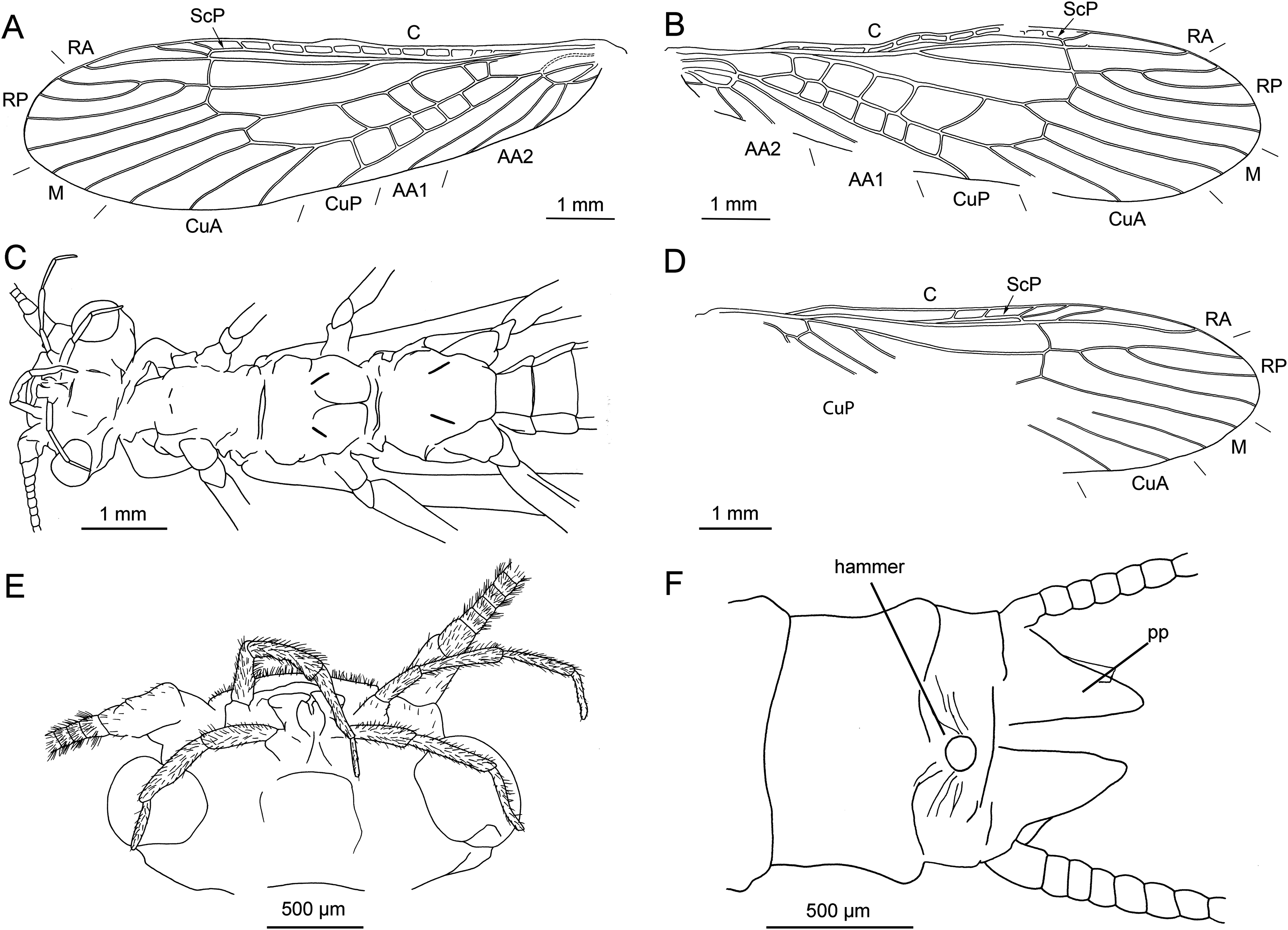{kind=link}
Diagnosis. Very large paraprocts, knob-shaped, circular hammer, slender body, trapezoidal pronotum.
Etymology. The name refers to Bill Wyman, former bass player of the Rolling Stones until 1991.
Material. Holotype specimen: SMNS BU-229, male.
Description. Body length 10.2 mm (Fig. 9A).
Head. Colour brown without distinct markings. Surface of head capsule with numerous short hair-like setae. Three fully developed ocelli. Median ocellus subequal in size to lateral ocelli. Antennae 7.6 mm long (approximately 0.7× body length). Individual segments covered with short hair-like setae. Antennal segments in the middle third of antenna length approximately 2.5× longer than wide.
Maxillary palps slender, with five elongated palpomeres covered with short hair-like setae (Figs. 9B and 10E). Palpomeres 1 and 5 short, palpomeres 2–4 approximately equal in length. Labium with glossae much smaller than paraglossae. Labial palps elongated, palpomere 2 longest, palpomeres 1 and 3 slightly shorter (Figs. 9B, 10C and 10E). All palpomeres covered with short hair-like setae. Other mouthparts not recognizable.
Thorax. Prothorax trapezoidal, distinctly wider anteriorly. Surface of pronotum with numerous short hair-like setae. Colour brown with pale longitudinal median band, poorly visible due to state of preservation.
Forewings (Figs. 10A and 10B): length 9.3 mm, width 2.5 mm; costal field with up to 13 crossveins (single crossvein distal to ScP faintly visible on right forewing); ScP reaching RA approximately at 2/3 of wing length; RA simple; single crossvein between RA and RP; RP originating from R distal to 1/3 of wing length; RP with four distal branches; single crossvein between RP and M; M approximated to R basally; M with two branches, originating proximally to 2/3 of wing length; occurrence of arculus and five additional crossveins between M and CuA; CuA branched distally, with three branches; five to six crossveins between CuA and CuP; CuP simple and straight; single cubito-anal crossvein reaching anal cell; AA1 simple; AA2 with two branches originating directly from anal cell.
Hind wings (Fig. 10D): length 7.8 mm; costal field with five visible crossveins (including single crossvein distal to ScP); ScP reaching RA proximal to 2/3 of wing length; RA simple; single crossvein between RA and RP; RP originating from common stem with M at 1/3 of wing length; RP with four distal branches; single crossvein between RP and M; branching of M, Cu and AA not recognizable.
Thoracic sterna (Fig. 10C) with apparent oblique furcal pits, converging anteriorly. Transverse meso- and metasternal ridge connecting anterior corners of respective furcal pits. Longitudinal median ridge apparent on mesofurcasternum, bifurcated and connecting anterior corners of furcal pits. Y-ridge connecting posterior corners of furcal pits absent. Gill remnants recognizable on all thoracic segments (Fig. 10C).
Legs slender, covered with short hair-like setae. Several longer and thicker spine-like setae occasionally on tibia. Two thickened tibial spurs and several setae apically on tibia (Figs. 9D and 9E). Very long hair-like setae subapically on tarsomere 3, approximately as long as claw length. Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.6× longer than first two combined. Arolium present, slightly wider than width of apical tarsomere. Euplantulae present on tarsomeres 2 and 3 (Fig. 9E).
Abdomen. Colour brown, ventral side paler. Individual segments not distinctly enlarged posteromedially. Elevated, knob-shaped and circular hammer near posterior margin of sternum IX (Figs. 9C and 10F).
Enlarged hook-like paraprocts between cerci (Figs. 9C and 10F). Paraprocts with convex lateral margins, not diverging apically. Number of hair-like setae apparent on sterna VIII, IX (Fig. 9C).
Cerci very short, consisting of 13 segments, 1.7 mm in length (approximately 0.2× body length). Slightly moniliform in shape; segments covered with long hair-like setae (Fig. 9C).
Affinities. Largusoperla billwymani sp. nov. share all the diagnostic characters of the Perlidae subfamily Acroneuriinae and the genus Largusoperla, as discussed above.
Largusoperla billwymani sp. nov. clearly differs from Largusoperla charliewattsi sp. nov. based on the general shape of body and wings (more slender with narrower wings in Largusoperla billwymani sp. nov.) and pronotum shape (distinctly trapezoidal in Largusoperla billwymani sp. nov., more oblong-shaped in Largusoperla charliewattsi sp. nov.). Largusoperla billwymani sp. nov. also exhibits larger paraprocts in comparison with L. charliewattsi sp. nov. and a different shape of hammer (larger, circular in Largusoperla billwymani sp. nov. contrary to transversally elongated in Largusoperla charliewattsi sp. nov.).
The paraprocts of Largusoperla billwymani sp. nov. are similar to Largusoperla flata, but slightly differ in shape, lateral contour being concave in Largusoperla flata (Chen, Wang & Du, 2018, figs. 12 and 13) and convex in Largusoperla billwymani sp. nov. (Figs. 9C and 10F). Largusoperla billwymani sp. nov. also exhibits narrower pronotum and more circular hammer compared to Largusoperla flata. Another difference can be found in the forewing venation, vein RP in Largusoperla flata is three-branched with bifurcation on the posterior branch (Chen, Wang & Du, 2018, fig. 11), whereas Largusoperla billwymani sp. nov. has a four-branched RP with bifurcations on anterior branches (Figs. 10A and 10B). Nevertheless, wing venation characters must be taken cautiously, since they are subject to large intraspecific variability (Béthoux et al., 2011).
Another two species Largusoperla acus and Largusoperla arcus have differently shaped paraprocts compared to Largusoperla billwymani sp. nov. (Largusoperla acus with strongly constricted, needle-shaped apices and Largusoperla arcus with apices distinctly curved outward). Largusoperla difformitatem can be distinguished from Largusoperla billwymani sp. nov. by having a smaller, transversally elongated hammer and also highly divergent paraprocts.
urn:lsid:zoobank.org:act:17E6ED82-03C8-4824-9176-51C34EFDF66F

Figure 11: Largusoperla micktaylori sp. nov., holotype SMNS BU-227, photographs.
(A) Dorsal view. (B) Ventral view. (C) Head in ventral view. (D) Three-lobed (arrows) subgenital plate.{kind=link}
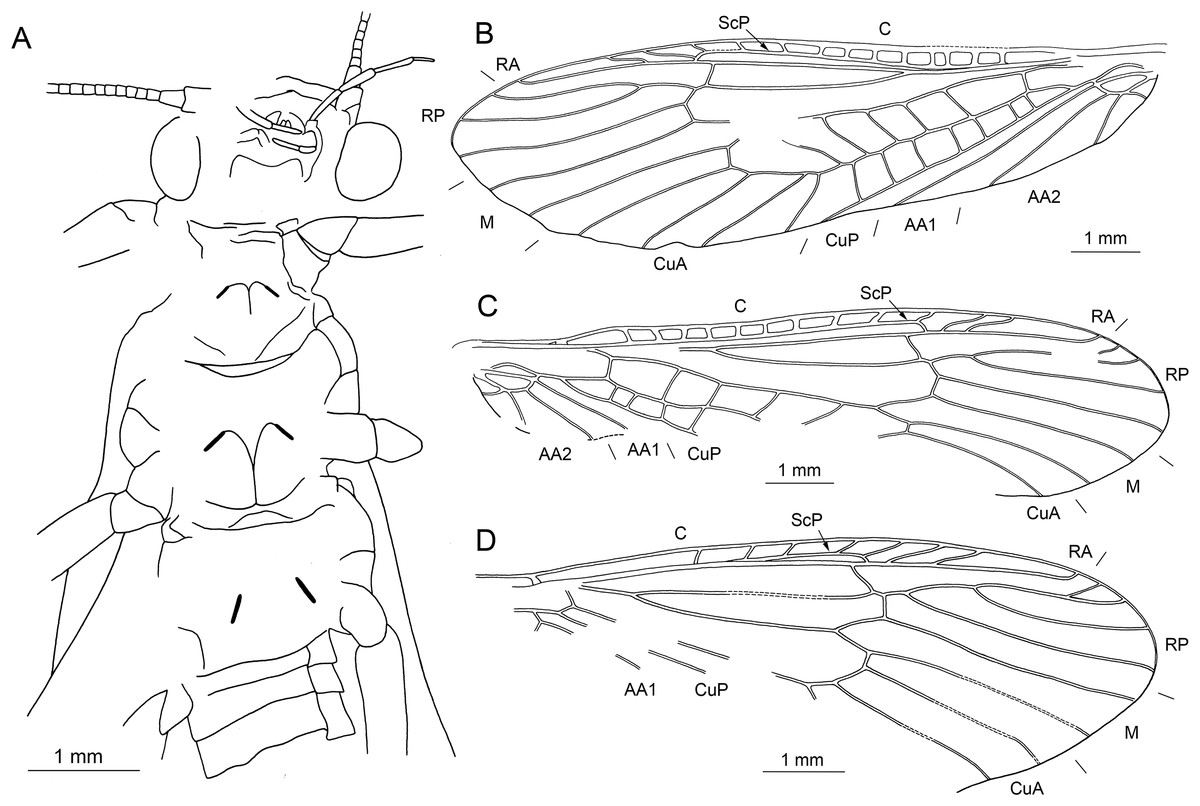
Figure 12: Largusoperla micktaylori sp. nov., holotype SMNS BU-227, line drawings.
(A) Head and thorax in ventral view. (B) Left forewing. (C) Right forewing. (D) Right hind wing.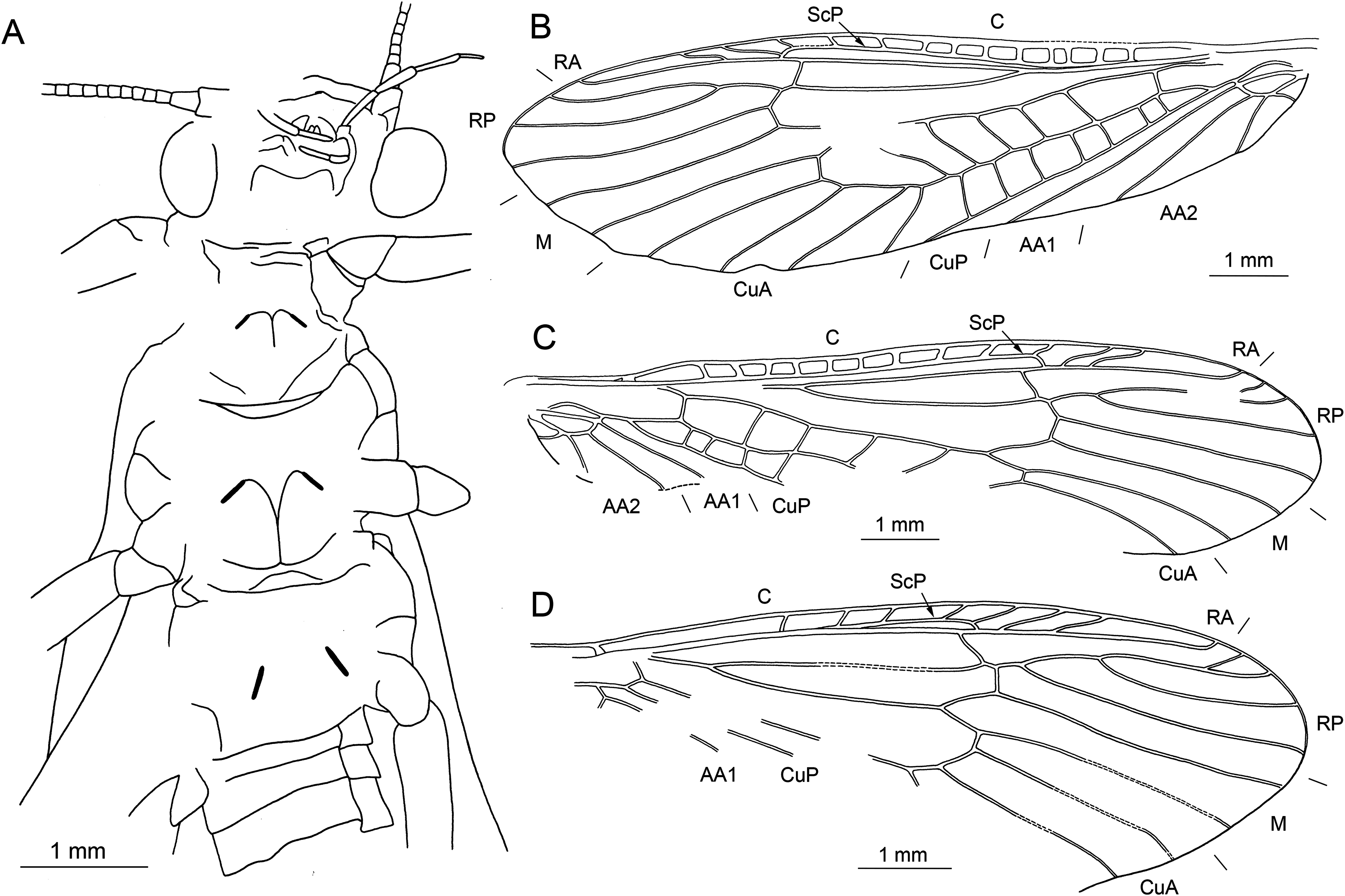{kind=link}

Figure 13: Largusoperla micktaylori sp. nov., paratype SMNS BU-312, photographs.
(A) Dorsal view. (B) Ventral view. (C) Head in ventral view. (D) Tarsus of right hind leg. (E) Ventral tip of abdomen with subgenital plate, paraprocts, and cerci. (F) Three-lobed (arrows) subgenital plate.{kind=link}
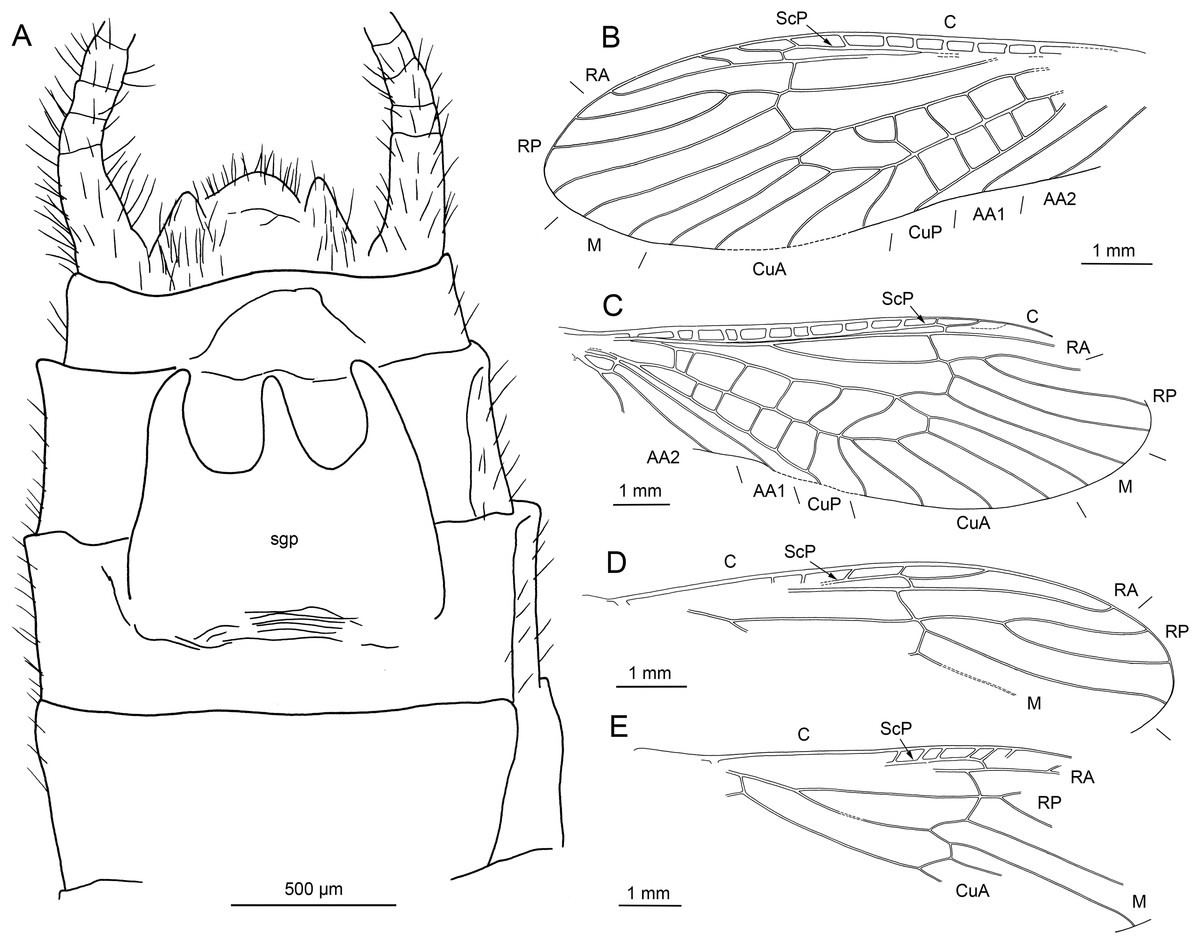
Figure 14: Largusoperla micktaylori sp. nov., paratype SMNS BU-312, line drawings.
(A) Ventral tip of abdomen with subgenital plate, paraprocts, and cerci. (B) Left forewing. (C) Right forewing. (D) Left hind wing (ventral view). (E) Right hind wing.{kind=link}
Diagnosis. Posterior margin of subgenital plate with three long, narrow, apically pointed processes.
Etymology. The name refers to Mick Taylor, guitar player of the Rolling Stones between 1969 and 1975 with unmatched virtuosity and dexterity, which is reflected by the finger-like, three-lobed subgenital plate of the new species.
Material. Holotype specimen SMNS BU-227, female; paratype specimen SMNS BU-312, female.
Description. Body length 8.6–10.5 mm (Figs. 11A, 11B, 13A and 13B).

Figure 15: Largusoperla brianjonesi sp. nov., holotype SMNS BU-311, photographs.
(A) Dorsal view. (B) Ventral view. (C) Head in dorsal view. (D) Bi-lobed (arrows) subgenital plate.{kind=link}
Head. Colour brown without distinct markings. Three fully developed ocelli. Median ocellus slightly subequal in size to lateral ocelli. Antennae 7.2–7.4 mm long (approximately 0.8× body length). Individual segments covered with short hair-like setae. Antennal segments in the middle third of antenna length approximately 2.2× longer than wide.
Maxillary palps slender, with five elongated palpomeres covered with short hair-like setae (Figs. 11C and 13C). Palpomeres 1 and 5 short, palpomeres 2–4 approximately equal in length. Labium with glossae much smaller than paraglossae. Labial palps elongated, three-segmented. All palpomeres of approximately same length (Figs. 11C and 13C). All palpomeres covered with short hair-like setae. Other mouthparts not recognizable.
Thorax. Pronotum trapezoidal, anteriorly slightly wider. Colour brown with central longitudinal pale band.
Forewings (Figs. 12B, 12C, 14B and 14C): length 10–10.7 mm, width 3.1–3.5 mm; costal field with 9–14 crossveins, including one to two crossveins distal to ScP; ScP reaching RA just proximally to 2/3 of wing length; RA simple; single crossvein between RA and RP; RP originating from R just distally to 1/3 of wing length; RP with three to four distal branches; single crossvein between RP and M; M slightly approximated to R basally; M with two branches, originating proximal to 2/3 of wing length; occurrence of arculus and three to five crossveins between M and CuA; CuA branched distally, with four branches; three to seven crossveins between CuA and CuP; CuP simple and straight; single cubito-anal crossvein reaching anal cell; AA1 simple; AA2 with three branches originating directly from anal cell.
Hind wings (Figs. 12D, 14D and 14E): length 8.7–8.8 mm; costal field with seven to eight crossveins, including one to two crossveins distal to ScP; ScP reaching RA distal to 1/2 of wing length; RA simple; single crossvein between RA and RP; RP originating from common stem with M at approximately 1/4 of wing length; RP with three to four distal branches; single crossvein between RP and M; M with two branches originating distal to 1/2 of wing length; CuA with two branches; CuP and A recognizable only partially.
Thoracic sterna (Fig. 12A) with apparent oblique furcal pits, converging anteriorly. Longitudinal median ridge apparent on mesofurcasternum, bifurcated in the middle of its length and connecting anterior corners of furcal pits. Y-ridge connecting posterior corners of furcal pits absent. Transverse meso- and metasternal ridge connecting anterior corners of respective furcal pits indistinct. Remnants of thoracic gills recognizable.
Legs slender, covered with short hair-like setae. Two thickened tibial spurs apically on tibia. Very long hair-like setae subapically on tarsomere 3, approximately as long as claw length. Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.5× longer than first two combined. Arolium present, slightly wider than width of apical tarsomere. Euplantulae prominent, present on tarsomeres 2 and 3.
Abdomen. Individual segments not distinctly enlarged posterolaterally. Cerci short, 2.2–2.3 mm in length (approximately 0.2× body length), covered with short setae (Figs. 13E and 14A). Subgenital plate with three long, narrow, apically pointed processes on posterior margin (Figs. 11D, 13E, 13F and 14A). All three processes of same length, one situated medially and two laterally.
Affinities. Largusoperla micktaylori sp. nov. shares with male representatives of the genus Largusoperla traits typical for the Perlidae (presence of long and slender palps, vestiges of thoracic gills, euplantulae, short first tarsomere, and forewings with numerous crossveins in the basal half of the costal field). Relative size of glossae and paraglossae is apparent, with paraglossae distinctly larger (Figs. 11C and 13C) and thus congruent with the assignment to Perlidae. The placement to the subfamily or tribe within Perlidae can be done only based on characters of male genitalia, which is inapplicable for Largusoperla micktaylori sp. nov. However, based on the presence of key generic diagnostic characters of Largusoperla (triocellate; pronotum with pale median area; short cerci, no longer than 1/2 of the abdomen length) together with overall high similarity in the arrangement of tarsi, mouthparts, wing venation and simultaneous occurrence with Largusoperla males, we include Largusoperla micktaylori sp. nov. into Largusoperla. From Pinguisoperla Chen, 2018, another Perlidae genus described from the Burmese amber, Largusoperla micktaylori sp. nov. clearly differs by the absence of an enlarged, plump basal segment of the cercus, which constitutes the main diagnostic character of Pinguisoperla.
The subgenital plate in Largusoperla micktaylori sp. nov. with three pronounced extensions on posterior margin is rather unusual and unique among known stoneflies and represents a crucial diagnostic character of Largusoperla micktaylori sp. nov. Since the shape of the subgenital plate is exactly the same in both specimens and since they also share other morphological characteristics (body size, arrangement of mouthparts, general wing venation pattern), we consider these specimens to represent a single new species.
Individual specimens of Largusoperla micktaylori sp. nov. differ from each other in the wing venation details such as number of crossveins and RP branches (Figs. 12B, 12C, 14B and 14C). However, these differences correspond with common intraspecific variability, as documented for recent Perlidae by Béthoux et al. (2011). In the holotype, the number of RP branches even differs between right and left forewing, being four and three, respectively, a phenomenon not uncommon in extant taxa.
urn:lsid:zoobank.org:act:3FF737E9-D935-48D7-901A-A1F5FBF61CE9
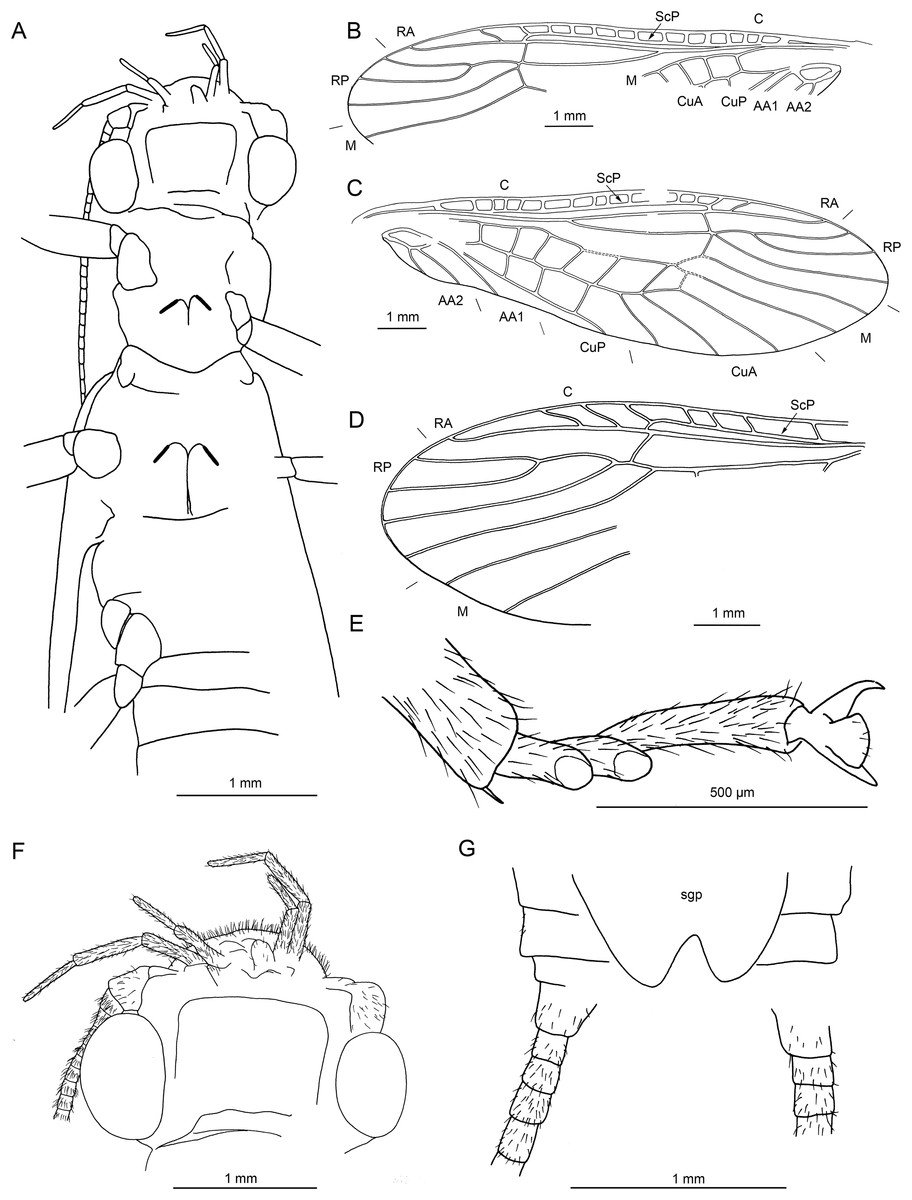
Figure 16: Largusoperla brianjonesi sp. nov., holotype SMNS BU-311, line drawings.
(A) Head and thorax in ventral view. (B) Left forewing. (C) Right forewing. (D) Right hind wing (ventral view). (E) Tarsus in ventral view. (F) Head in ventral view. (G) Bi-lobed subgenital plate.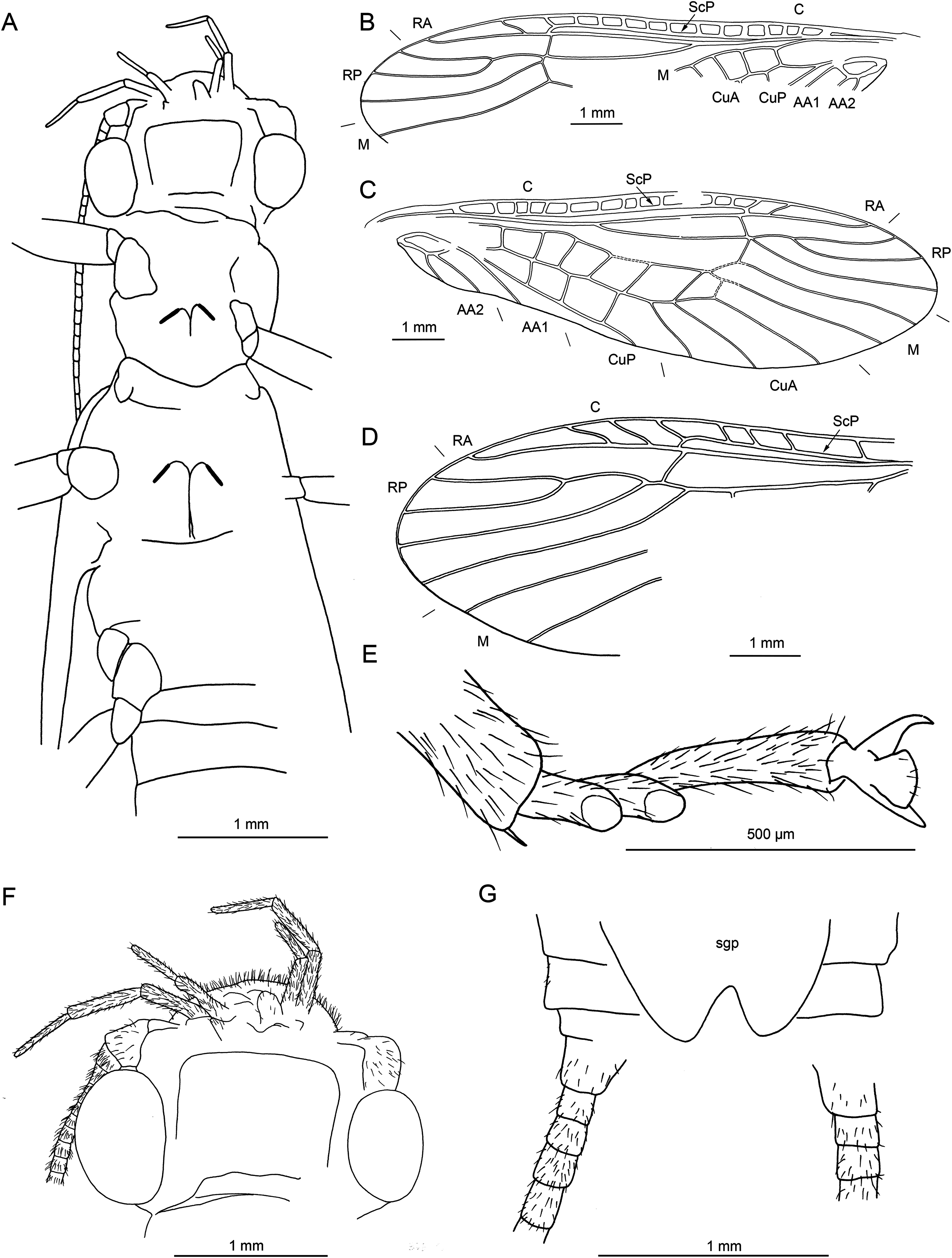{kind=link}
Diagnosis. Posterior margin of subgenital plate with two broadly rounded lobes.
Etymology. The name refers to Brian Jones, founding member and former guitar player of the Rolling Stones until 1969.
Material. Holotype specimen SMNS BU-311, female.
Description. Body length 10.3 mm (Figs. 15A and 15B).
Head. Colour brown, with distinct pale areas (Fig. 15C). Median ocellus slightly subequal in size to lateral ocelli. Antennae 6.4 mm long (approximately 0.6× body length). Individual segments covered with short hair-like setae. Antennal segments in middle third of antenna length approximately 2× longer than wide.
Maxillary palps with three visible elongated palpomeres of approximately same length. Two basal palpomeres not visible, presumably shorter than three distal palpomeres. Glossae and paraglossae not recognizable. Postmentum large. Labial palps rather elongated, three-segmented. First palpomere shortest, second slightly longer, apical palpomere longest (Fig. 16F).
Thorax. Pronotum slightly trapezoidal, wider anteriorly (Fig. 15A). Colour brown, pale longitudinal band medially.
Forewings (Figs. 16B and 16C): length 10.8 mm, width 3.1 mm; costal field with 14 crossveins visible (including single crossvein distal to ScP); ScP reaching RA approximately at 2/3 of wing length; RA simple; single crossvein between RA and RP; RP originating from R distal to 1/3 of wing length; RP with three distal branches; single crossvein between RP and M; M approximated to R basally; M with two branches, originating proximal to 2/3 of wing length; occurrence of arculus and six additional crossveins between M and CuA; CuA branched distally, with three branches; three crossveins between CuA and CuP; CuP simple and straight; AA1 simple; AA2 with two branches originating directly from anal cell.
Hind wings (Fig. 16D, details of venation only visible in apical portion): length 8.9 mm; costal field with seven visible crossveins (including two crossveins distal to ScP); ScP reaching RA distal to 1/2 of wing length; RA simple; single crossvein between RA and RP; RP originating from common stem with M approximately at 1/4 of wing length; RP with four distal branches; single crossvein between RP and M; course and branching of M, CuP and AA unrecognizable.
Thoracic sterna (Fig. 16A) with apparent oblique furcal pits, converging anteriorly. Transverse meso- and metasternal ridge connecting anterior corners of respective furcal pits. Longitudinal median ridge apparent on mesofurcasternum, connecting anterior corners of furcal pits. Y-ridge connecting posterior corners of furcal pits absent. Remnants of thoracic gills recognizable on all thoracic segments.
Legs slender, covered with short hair-like setae. One thickened tibial spur and several setae apically on tibia (Fig. 16E). Tarsi with first two tarsomeres short (approximately equal in size) and apical tarsomere long, approximately 1.5× longer than first two combined. Arolium present, slightly wider than width of apical tarsomere. Euplantulae present on tarsomeres 2 and 3 (Fig. 16E).
Abdomen. Individual segments not distinctly enlarged posterolaterally. Cerci very short, 1.9 mm in length (approximately 0.2× body length). Subgenital plate bilobed with wide rounded notch in middle of posterior margin (Figs. 15D and 16G).
Affinities. Largusoperla brianjonesi sp. nov. is very similar to Largusoperla micktaylori sp. nov., sharing all the diagnostic characters for the family Perlidae and genus Largusoperla as discussed above. At the same time, Largusoperla brianjonesi sp. nov. clearly differs from Largusoperla micktaylori sp. nov. based on the shape of the subgenital plate (with two broad lobes on the posterior margin in Largusoperla brianjonesi sp. nov. in contrast to the presence of three narrow projections in Largusoperla micktaylori sp. nov.).

Figure 17: Largusoperla sp., SMNS BU-228, photographs.
(A) Dorsal view. (B) Head in ventral view. (C) Ventral tip of abdomen with hammer, paraprocts, pp, and cercus, ce. (D) Tarsus of left hind leg.{kind=link}
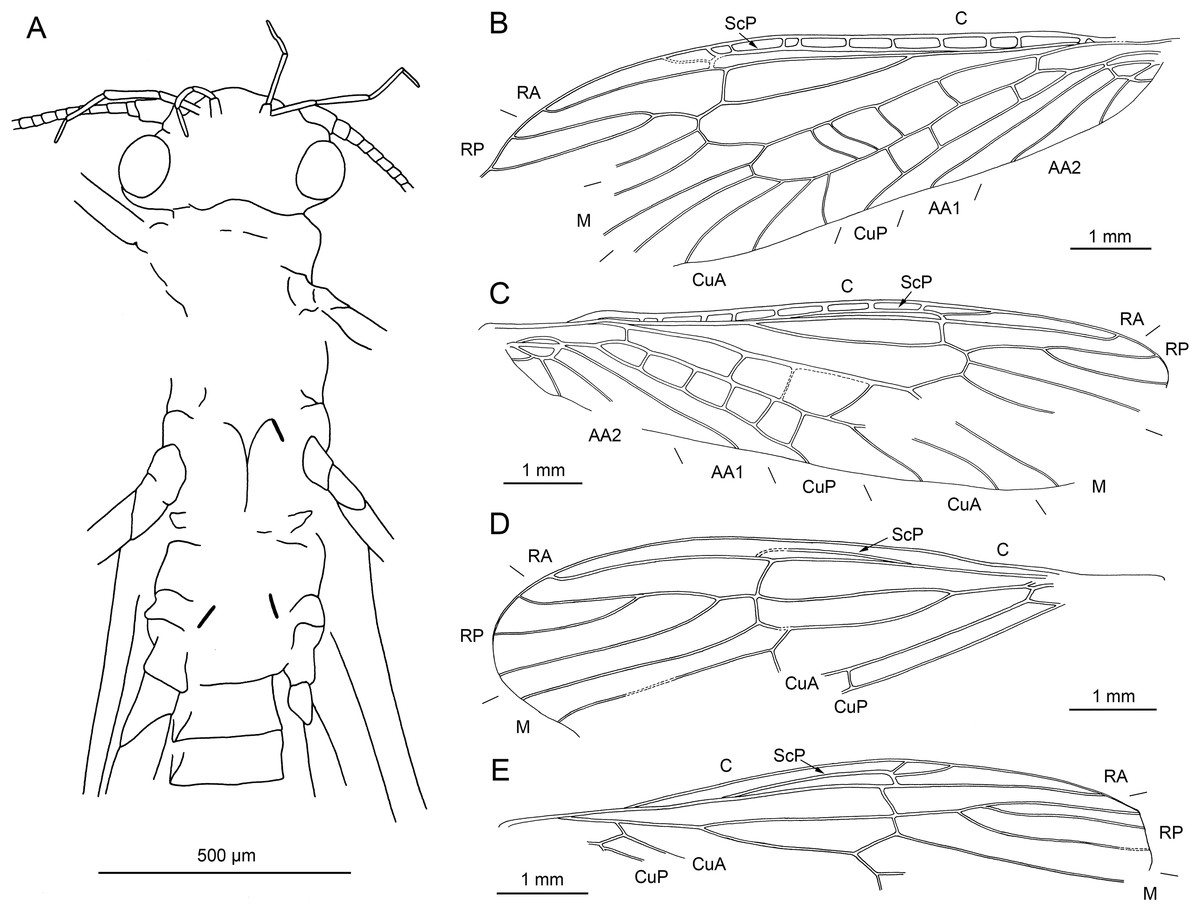
Figure 18: Largusoperla sp., SMNS BU-228, line drawings.
(A) Head and thorax in ventral view. (B) Left forewing. (C) Right forewing. (D) Left hind wing. (E) Right hind wing.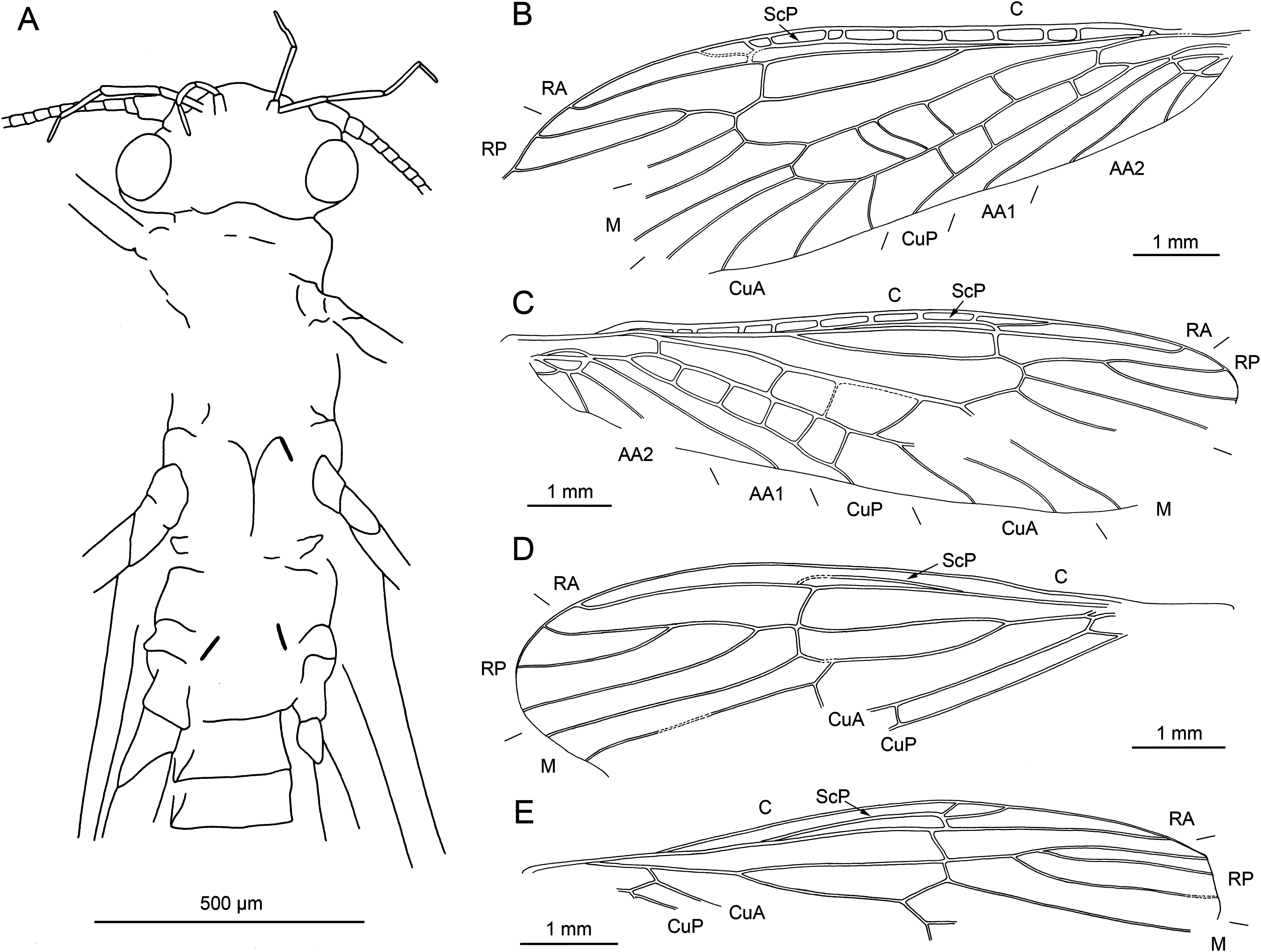{kind=link}
Material. SMNS BU-228 (Figs. 17 and 18)
Perlidae: Acroneuriinae sp. (Fig. 19)

Figure 19: Acroneuriinae spp., SMNS BU-99, photograph.
Dorsal view.{kind=link}
Material. SMNS BU-99 (Fig. 19).
Remarks to species descriptions. We refrain from associating males and females described in Largusoperla, since any association would be purely speculative. Clearly, multiple similar species lived in the common palaeohabitat.
The larva described above as Electroneuria ronwoodi sp. nov. is attributable to the subfamily Acroneuriinae, as all the adults described here in Largusoperla and possibly also adults of Pinguisoperla, described by Chen (2018b). However, we refrain from assigning Electroneuria ronwoodi sp. nov. into one of these two genera, based on the difference in the length of cerci. Usually there is a correlation between the length of cerci in larvae and adults in Systellognatha (Zwick, 1980). Electroneuria ronwoodi sp. nov. exhibits much longer cerci than Largusoperla and Pinguisoperla, therefore we prefer assigning this species to a separate genus at this time. As Electroneuria ronwoodi has an age of about 100 million years, we also exclude its placement within an extant genus of Acroneuriinae.
Discriminating species characters for males of Acroneuriinae are the mainly detailed shape of hammer and paraprocts (Stark, Froehlich & Zuñiga, 2009). For females, the crucial structure is the shape of subgenital plate. Therefore, we designate species only for the specimens with these structures well visible. We studied another male specimen (SMNS BU-228) attributable to Largusoperla, but with hammer and/or paraprocts insufficiently visible to allow detailed comparisons. Yet another investigated specimen (SMNS BU-99) is assignable to Acroneuriinae, but without visible genitalia and generally too poorly preserved to allow generic attribution.
Discussion
Phylogenetic position of Petroperlidae
The absence of crossveins in the distal half of (ScP+) RA–RP area allows the placement of Petroperlidae in the suborder Arctoperlaria (Cui et al., 2015).
Among characters of phylogenetic significance, segmentation of tarsi is well visible in the specimens. The first two tarsomeres are short, approximately equal in size and the apical tarsomere is long, approximately 3.5× longer than first tarsomere (Figs. 2B and 3D). A short basal tarsomere is either considered an autapomorphy of Perloidea (Zwick, 1973), a synapomorphy of Perloidea with Peltoperlidae + Styloperlidae (Zwick, 1980), or as a character independently derived in these lineages (Zwick, 2000; Nelson, 2009). In any case, it points to the placement of Petroperlidae in the infraorder Systellognatha. Any assignment to the infraorder Euholognatha is excluded because of the presence of euplantulae on tarsomeres 1 and 2, absent in the Euholognatha (Nelson, 2009). Further diagnostic characters of Systellognatha are the apomorphic presence of setae on the arolium (see Figs. 2B and 3D) (Nelson, 2009) and the plesiomorphic presence of numerous crossveins in the costal field, both of which also accounts for Petroperla gen. nov. and Lapisperla gen. nov. (Figs. 2E and 4A).
Moreover, in the forewing of Lapisperla keithrichardsi sp. nov., the basal crossvein in the costal field (Fig. 4B) is stronger than the remaining crossveins, short, and of opposed obliquity (in Petroperla mickjaggeri sp. nov. the view onto this character is obstructed). This character has been proposed as diagnostic for Systellognatha by Cui et al. (2015).
Within Systellognatha, a labium with glossae only slightly smaller than paraglossae and with rather stout labial palps (Figs. 1C and 2A) excludes a closer relationship of Petroperla mickjaggeri sp. nov. to Perloidea, the latter characterized by apomorphic shape of labium with glossae much shorter than paraglossae (Zwick, 2000). Therefore, based on the shape of labium, a relationship to the other systellognathan superfamily, Pteronarcyoidea, would seem more probable. However, neither Petroperla gen. nov. nor Lapisperla gen. nov. any of the wing venation characters known to represent traits of individual Pteronarcyoidea lineages (Cui et al., 2016), which are numerous crossveins between M and CuA in distal half of wing; AA2 with more than three branches; and M with more than two branches. Moreover, Petroperlidae feature several wing venation characters not present in Systellognatha. These include a vein RA almost reaching the wing apex, and a plesiomorphic origin of vein RP in the forewing close to wing base (Figs. 2E, 2F and 4A). Vein RA almost reaching the wing apex is absent in Carboniferous stem-stonefly G. carpenteri and is not known for any other stonefly taxa, both extant and fossil, except of the genus Kargaloperla Sinitshenkova, 1987 from the Upper Permian of Ural (Palaeoperlidae, see Sinitshenkova, 1987, fig. 10). It is unlikely that it represents a plesiomorphic character that was independently lost in several other basal stonefly lineages (including G. carpenteri). It is more parsimonious to assume that it is either an autapomorphy of Petroperlidae, or even a synapomorphy of Petroperlidae and Kargaloperla.
An origin of RP close to the wing base presumably represents a plecopteran ground plan character known already from Paleozoic stoneflies. It is also present in G. carpenteri, and according to Béthoux et al. (2011) it is a diagnostic character of early stoneflies, also supporting the affinity of G. carpenteri to Plecoptera. It is also present in Permian Palaeoperla exacta Sharov, 1961 (Palaeoperlidae) and Perlopsis filicornis Martynov, 1940 (Perlopseidae) and can be observed in Euholognatha as well, e.g. recent Leuctridae or fossil Perlariopseidae (Sinitshenkova, 1992, fig. 2). Probably the origin of RP in the forewing situated close to the wing base is also retained in Petroperlidae and lost in all other Systellognatha.
Based on available evidence, we propose the phylogenetic position of Petroperlidae as a distinct stem lineage of Systellognatha. The presence of a short first tarsomere is evaluated here as potential apomorphic character of Systellognatha and was already early present in the stemline of Systellognatha. Further support for a placement in Systellognathan stemline is the apomorphic presence of an arolium with setae. Tarsal euplantulae and numerous crossveins in the costal field are plesiomorphies shared with remaining Systellognatha. The plesiomorphic proximal origin of RP excludes the position of Petroperlidae in the crown group Systellognatha, and RA reaching the wing apex might be an autapomorphic character of Petroperlidae. Vein RA almost reaching the wing apex is not completely preserved in Lapisperla keithrichardsi sp. nov. due to the missing wing apices. However, from the course of RA in right hind wing approximating the wing apex (Figs. 4D and 4E), we assume a similar pattern in Petroperla mickjaggeri sp. nov. Therefore, as Lapisperla keithrichardsi sp. nov. and Petroperla mickjaggeri sp. nov. share the same unique combination of characters, we assign them to the same family Petroperlidae.
Relationships with several fossil groups considered as stem-Systellognatha (Illies, 1965; Stark & Gaufin, 1976) cannot be reliably analysed. Permian Tshekardoperlidae Sinitshenkova, 1987 and Jurassic Platyperlidae Sinitshenkova, 1982 are known in the larval stage only (Sinitshenkova, 1987; Carpenter, 1992). The family Palaeoperlidae Sharov, 1961, whose members sometimes exhibit aforementioned wing venation characters, represent a very vaguely defined Permian group known only from larvae and isolated forewings and thus many other characters (tarsi, mouthparts) cannot be compared. Association of larvae and adults described as Palaeoperlidae is also uncertain. In Sinitshenkova (2002), Palaeoperlidae was considered as a sister group of Perlina (containing Zwick’s Systellognatha + Antarctoperlaria), in other words one of the most basal branches within Plecoptera. However, wing venation indicates at least some members of Palaeoperlidae (genus Kargaloperla) might be related to Petroperlidae, and Palaeoperlidae might not constitute a monophyletic group.
Systematic position of Electroneuria ronwoodi gen. nov. sp. nov
Predaceous mouthparts with long and slender palps are diagnostic for larvae of the superfamily Perloidea within the arctoperlariid group Systellognatha (Zwick, 1980).
Of the three families of Perloidea (Perlidae, Perlodidae, and Chloroperlidae), the wing pad arrangement of Electroneuria ronwoodi sp. nov. rather resembles Perlidae and Chloroperlidae, since the ‘rounded shape of larval wing-pads whose edges meet medially, without leaving a separate notal contour’ is mentioned as a possible synapomorphy of these two families within Perloidea by Zwick (2000).
Electroneuria ronwoodi sp. nov. can be excluded from Chloroperlidae, as larvae of this family exhibit a slender body with usually an oval pronotum and cerci distinctly shorter than the abdomen (see examples in Brittain & Saltveit, 1996; Stewart & Stark, 2002), presumably apomorphic features of the family (Zwick, 2000). On the contrary, the body of Electroneuria ronwoodi sp. nov. is relatively robust bearing long cerci. Another apomorphic character of Chloroperlidae (except for subfamily Paraperlinae) is a thin, asymmetrically inserted, terminal maxillary palpomere (Zwick, 1973, 1980, 2000; Surdick, 1985), which is not present in Electroneuria ronwoodi sp. nov.
Thus more likely is a placement within Perlidae, also supported by the presence of thoracic gills (Zwick, 1980). Thoracic gills are difficult to identify in Electroneuria ronwoodi sp. nov. due to its poor preservation. The specimen most probably is an exuvia, as only the exoskeleton is preserved. Furthermore, respective body regions corresponding with the insertion points of thoracic gills are obscured by mineral particles, but the remains of thoracic gills might be visible at least on the left side of the body, between meso- and metathorax. Its affinity to Perlidae is further confirmed by the presence of an occipital row of short spinules (Stark & Gaufin, 1976; Sivec, Stark & Uchida, 1988; Zwick, 2000). The arrangement of this row represents the larval key character distinguishing Perlidae subfamilies Perlinae and Acroneuriinae (Stark & Gaufin, 1976). In Electroneuria ronwoodi sp. nov., the row is slightly sinuate, regular medially, in lateral parts disintegrate into scattered goups of longer spinules (Figs. 5D and 6E), an arrangement present in Acroneuriinae, in contrast to the straight and complete occipital row of Perlinae (Stark & Gaufin, 1976; Sivec, Stark & Uchida, 1988).
There are also two Permian families, Palaeoperlidae and Tshekardoperlidae, which might be candidates for a closer relationship to Electroneuria gen. nov., but both of them are rather poorly defined. Diagnoses are mostly based on the length ratios of body parts (Sinitshenkova, 1987). Both Palaeoperlidae and Tshekardoperlidae differ from Electroneuria ronwoodi (apart from the marked difference in geological layer) in the shape of thoracic segments with clearly visible notal contour between wingpads (figs. 8, 25–27 in Sinitshenkova, 1987). Furthermore, Tshekardoperlidae are characterized by prolonged antennal segments, 2–3 times longer than wide in the middle third of the antenna (Sinitshenkova, 1987). Electroneuria ronwoodi sp. nov. exhibits much shorter antennal segments (Fig. 5C). Jurassic Platyperlidae most notably differ by the presence of a median projection on the posterior margin of terminal abdominal segment (Carpenter, 1992; Sinitshenkova, 1987) which is missing in Electroneuria ronwoodi sp. nov.
Based on the combination of characters as presented above, we attribute Electroneuria ronwoodi to the Perlidae subfamily Acroneuriinae. There are no larval diagnostic characters for the three Acroneuriinae tribes (Stark & Gaufin, 1976; Murányi & Li, 2016), but from all described genera of Acroneuriini, Anacroneuriini, and Kiotinini, the larval specimen Electroneuria ronwoodi sp. nov. can be diagnosed by the combination of characters as follows: (1) occipital spinule row complete medially, (2) fringe of long thin setae laterally on pronotum (most of them broken, but bases clearly visible), (3) long hair-like setae on surface of wing pads and abdominal terga, (4) posterior margin of abdominal terga with numerous very long thin setae, and (5) long setae on cerci absent.
Palaeodiversity, Palaeoecology, and Palaeobiogeography of Perlidae
The family Perlidae, despite being very abundant and diversified today, was so far only scarcely found in the fossil record. Several specimens of the extant genus Perla are known from Eocene Baltic amber (Pictet & Hagen, 1856; Carpenter, 1992). A single immature larva of Perla cf. burmeisteriana Claassen, 1936 was described by Prokop (2002) from Lower Miocene deposits of the Czech Republic. An adult specimen of Dominiperla antigua Stark & Lenz, 1992 was described from Dominican amber (dated between upper Eocene to lower Miocene) and attributed to Acroneuriinae (subfamily ‘Anacroneuriinae,’ as stated in Stark & Lenz, 1992 probably represents a spelling error). Affinities were evaluated based on the course of M1 vein, only slightly divergent from R until the origin of Rs (present in some Perlidae), and small, spindle-shaped eggs similar to the Neotropical genus Anacroneuria. A young larva is also known from Oligocene of SW Montana, attributed to ?Acroneuria (Lewis & Gundersen, 1987). Another larva was described as Euperlida parvicercifera Cifuentes-Ruiz, 2007 from Oligocene of Mexico (Cifuentes-Ruiz et al., 2007) and tentatively attributed to Perlidae subfamily Acroneuriinae based on general body resemblance (the shape and size of the head, wing pads, body pattern, foreleg, and pronotum shape).
The oldest Perlidae species Archaeoperla rarissimus Liu, Ren & Sinitshenkova, 2008 described by Liu, Sinitshenkova & Ren (2008) from Jurassic/Cretaceous of China, was later synonymized by Cui et al. (2015) with Sinosharaperla zhaoi Liu et al., 2007, which probably represents stem-Systellognatha, although attribution to Perlidae was not excluded by Cui et al. (2015). If we do not consider S. zhaoi as a genuine Perlidae, the genera Largusoperla, Pinguisoperla, and Electroneuria from the mid-Cretaceous represent the oldest record of the family Perlidae. At the same time, they represent fossils attributed to Perlidae without doubt and based on multiple characters. Previously published Perlidae records were assigned to this family mostly based on a limited number of characters, often only on a superficial resemblance.
Three tribes were proposed within Perlidae subfamily Acroneuriinae based on adult male characters (Acroneuriini, Anacroneuriini, and Kiotinini, see Stark & Gaufin, 1976; Stark, Froehlich & Zuñiga, 2009; Murányi & Li, 2016). Regarding the position of Largusoperla, we concur with Chen, Wang & Du (2018) that the form of the hammer, an elevated knob rather than a low callus, suggests affinity to Anacroneuriini, whereas the well-developed anterior ocellus and short cerci represent plesiomorphies within Perlidae and suggest a possible position of Largusoperla in the stemline of Acroneuriini + Anacroneuriini.
Out of 10 stonefly specimens described in the present study, eight are attributed to the subfamily Acroneuriinae and, apart from one larval specimen, belong to the single genus Largusoperla, as the four species described recently by Chen, Wang & Du (2018) and Chen (2018c). A single specimen described by Chen (2018b) in the genus Pinguisoperla probably also belongs to the same subfamily. From such a pattern (even taking into consideration a fragmentary nature of the fossil record), we might assume that the stonefly community in the Burmese amber forest streams was dominated by representatives of a limited number of higher taxa. This might be caused by the nature of the original habitat, which has been assumed to be a tropical Araucaria forest (Grimaldi, Engel & Nascimbene, 2002). Present stonefly diversity is concentrated in the temperate regions, a pattern probably consistently followed throughout their history (Sinitshenkova, 1997). However, as demonstrated by our data, some groups like Acroneuriinae had been adapted to the warmer streams of the Cretaceous Burmese amber palaeohabitat. Although the adults might be speculated to represent allochthonous material in the inclusions, our finding of a larva of the same taxonomic placement rather corroborates the occurrence of Acroneuriinae in the Burmese amber forest streams.
With more than 1,000 described species, Perlidae is the most diverse extant family. The most diverse perlid areas are Asia and Central and South America. At present, the Chinese perlid species represent about 25% of the world Perlidae, making the Oriental region one of the most important areas of the world for perlid diversity (Fochetti & De Figueroa, 2008). We assume placement of all Acroneuriinae adult material dealt with in the present study into the tribe Anacroneuriini sensu Stark & Gaufin (1976). At present, all extant Anacroneuriini are restricted to the Neotropics and southern portion of the Nearctic (Murányi & Li, 2016). Stark & Gaufin (1976) hypothesized recent Anacroneuriini arose from a now extinct Oriental group which dispersed across Northern Africa in Early Cretaceous and became established in South America. Largusoperla and Electroneuria gen. nov. might actually represent such a stem-Anacroneuriini group, still to be found in the Oriental region during the late Cretaceous and subsequently becoming extinct.
Close relations between Neotropic and Oriental fauna dating back to the Cretaceous were already known in other groups of aquatic and terrestrial insects, e.g. the Burmese amber damselfly family Mesomegaloprepidae, related to extant Neotropical taxa (Huang et al., 2017) or the psocid family Compsocidae, known from Burmese amber and the extant fauna of Central America (Azar, Hakim & Huang, 2016; Sroka & Nel, 2017). Our study thus provides support for the hypothesis of Stark & Gaufin (1976) claiming oriental origin of Anacroneuriini.
Conclusions
Our findings report with the fossil family Petroperlidae the presence of a new taxon in the stem lineage of Systellognatha in the Cretaceous. It reveals new insights into the character evolution within this group: Petroperlidae have shortened tarsomeres 1 and 2, which consequently must now be assumed as ground plan character of Systellognatha. Otherwise, the Burmese Cretaceous stonefly fauna was dominated by species of the Anacroneuriini (Perlidae: Acroneuriinae), a taxon, which today is restricted to the Nearctic realm. This points to an oriental origin of Anacroneuriini, thus confirming the respective hypothesis of Stark & Gaufin (1976).