
Processing characteristics of visuospatial working memory in map cognition by orienteering athletes: evidence from behavioral and EEG studies
- Published
- Accepted
- Received
- Academic Editor
- Matthew Parker
- Subject Areas
- Neuroscience, Psychiatry and Psychology
- Keywords
- Orienteering, Map cognition, Memory store, Memory retrieval, EEG
- Copyright
- © 2025 Zhu et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2025. Processing characteristics of visuospatial working memory in map cognition by orienteering athletes: evidence from behavioral and EEG studies. PeerJ 13:e19817 https://doi.org/10.7717/peerj.19817
Abstract
Through maps, the objects on the Earth’s surface are abstracted and represented on a plane, achieving the integration of physical entities and conceptual representations, thereby facilitating individuals’ rapid understanding and profound cognition of their surroundings. As a system for the storing and processing of visual and spatial information, visuospatial working memory (VSWM) provides individuals with the capacity to temporarily store and process map-related information within a spatial framework. To explore the temporal features of storing and retrieving in VSWM during map-related tasks among orienteers with varying skill levels, 22 orienteers were enrolled in the expert group, while 22 members from orienteering training teams were placed in the novice group. Employing electroencephalography (EEG) technology, this study developed a set of VSWM tasks to examine behavioral performance variations and EEG characteristics between experts and novices. The results revealed that experts exhibited enhanced rationality and superior response efficiency during the map cognition task relative to novices. Neurophysiological evidence further indicated distinct neural resource management patterns in experts, characterized by reduced θ neural oscillations in the frontal regions coupled with enhanced α oscillations in both the frontal and temporal lobes. The empirical outcomes confirm competitive advantages in expert athletes’ map processing, response speed, and optimized neural resource allocation. These findings not only deepen insights into the neural mechanisms underlying orienteers’ map cognition, but also provide strong theoretical support for orienteering training practices.
Introduction
Maps, the carriers of geospatial information, serve as a graphical representation of an individual’s spatial cognitive abilities (Dong et al., 2019). Geospatial information is presented visually through graphics, symbols, colors, etc., substantially augmenting human capacity for rapid environmental comprehension and spatial awareness. Map cognition refers to process by which an individual, while exploring unfamiliar environments, engages higher brain functions to form a comprehensive mental model including spatial layouts such as roads, directions, and distances, as well as temporal relationships and non-spatial information (Tolman, 1948). As a system for the storing and processing of visual and spatial information (Zarantonello et al., 2024), visuospatial working memory (VSWM) provides individuals with the capacity to temporarily store and process map-related information. The improvement in VSWM ability can substantially enhance an individual’s performance in map cognition tasks, allowing them to understand and remember the layout of their environment more accurately (Harris & Wolbers, 2014; Hegarty et al., 2006). This indicates that the construction of a “cognitive map” helps individuals better understand the spatial relationships between objects, and the formation and consolidation of visuospatial memory provide the foundation for spatial imagination.
In competitive sports, it is not only necessary for athletes to store and refine information from surrounding environment, but it is even more necessary to actively retrieve relevant internal representations, so that the newly acquired information can be deeply integrated to adapt to the demands of the current situation or task (Repovs & Baddeley, 2006). Research indicates that compared with novices, expert athletes demonstrate a significant advantage in memory store and retrieval. For example, in sports such as cricket and football, expert athletes demonstrate higher accuracy and efficiency when recalling the player layout on the court (Houlston & Lowes, 1993; Panchuk & Vickers, 2006). In orienteering competitions, maps are indispensable tools for athletes, especially for spatial cognition and navigation. According to the theory of map spatial cognition (Huang et al., 2024), athletes first need to visually search for key information such as checkpoints, routes, buildings, etc. from complex map information, and then mobilize the visuospatial working memory system to memorize the relevant information and match and adjust it with the field environment in real time to quickly reach each checkpoint and complete the task (Liu & He, 2016). In this process, orienteers need to have strong VSWM to carry out tasks like visual search, memory recall, and decision-making. This study has shown that specialized VSWM training can significantly enhance orienteers’ map-reading performance (Wang, 2022). The capacity of VSWM influences cognitive decision-making in orienteers differently when engaging in precise versus rough map-reading tasks (Liu, 2021). Elite orienteers typically exhibit exceptional visual memory abilities, further supporting the importance of VSWM in orienteering (Wang, 2010). Although behavioral studies have widely confirmed the memory advantages of expert athletes, the underlying neural mechanisms—particularly the specific patterns of brain activity involved in memory storage and retrieval, as well as the differences in these processes—still require further investigation.
The Baddeley–Hitch model (abbreviated as the “B&H model”) posits that when processing short-term information, the brain not only retains information, but also manipulates and prepossess it during this retention process (Baddeley & Hitch, 1977). Specifically, the memory process is delineated into two critical phases: the first is the storage process, where information is effectively encoded and initially stored in the short-term memory system; the second is the retrieval phase, involving the ability to quickly and accurately retrieve the required information from stored data. Some studies have found that there are differences in the brain areas activated by individuals during the memory storage phase and the memory extraction phase (Kuhl & Chun, 2014). Theta oscillations can coordinate neuronal activity related to human cognition, particularly memory functions (Herweg, Solomon & Kahana, 2020; Rozengurt et al., 2023), and can be enhanced in the frontal and temporal lobes during visuospatial and visual information encoding (Xie et al., 2021; Jaiswal, Ray & Slobounov, 2009). On the other hand, Alpha oscillations facilitate memory encoding by suppressing distractibility (Strunk & Duarte, 2019), and a reduction in αband activity in the frontal region indicates higher cognitive neural efficiency (Cona et al., 2019). Therefore, the study utilizes electroencephalogram (EEG) technology to monitor θ and α neural oscillations in orienteers of varying skill levels across different memory phases with the goal of revealing the cognitive features of VSWM processing in orienteers.
In summary, this study employs a recall paradigm, uses orienteering competition maps as experimental materials, designs a map cognition task, and utilizes EEG technology to explore the influence of orienteering training experience on the processing of visual short-term working memory (VSWM) of orienteers. The study hypothesizes that, during map cognition, the expert group will exhibit a motor expertise advantage, and significant differences in brain activity between the experts and novices will be observed in the frontal and temporal lobes during the memory storage and retrieval phases.
Materials and Methods
Participants
The required sample size for this study was calculated using G*Power 3.1.9 (Faul et al., 2009). A two-tailed test was specified with an effect size of Cohen’s f = 0.25 and a statistical power of 1-β = 0.95. Repeated measures analysis of variance (ANOVA) was employed as the statistical method, resulting in a required sample size of 36 participants. To account for potential data quality issues such as attrition or technical errors, the sample size was increased to 44 participants. Following the criteria for expert athletes established by Swann, Moran & Piggott (2015), the 44 participants were categorized into expert (22 participants) and novice groups (22 participants), as detailed in Table 1.
| Group | Sport grade | Age/Year | Training years/ Year | Training frequencytimes/week |
|---|---|---|---|---|
| Experts | National level 2 or higher | 21.23 ± 2.25 | 5.35 ± 1.12 | ≥3 |
| Novices | NO | 20.17 ± 1.34 | 2.00 ± 0.63 | 1–2 |
Inclusion criteria comprised: (1) normal or corrected-to-normal binocular visual acuity, with no neurological disorders; (2) right-handedness; (3) absence of sleep deprivation (e.g., insomnia or staying up late) within 24 h prior to experimentation, and optimal alertness during testing; (4) no prior participation in cognitively analogous studies.
All participants provided signed informed consent forms. The protocol adhered to ethical principles outlined in the Declaration of Helsinki and was approved by the Ethics Committee of Shaanxi Normal University (Approval number: 202416006).
Measuring instruments
Experiments were performed using the eye-tracker aSee pro (7 Invensun, Beijing, China) to present the stimuli. The eye-tracker was in a local area network (LAN) with the EEG acquisition device, and the eye-tracking device sent signals to the EEG device via a wireless synchronization box at the beginning of each stimulation to ensure the accuracy of the data.
EEG data were acquired using a wireless multichannel EEG acquisition system (ZhenTec NT1, ZhenTec Intelligence Technologies Co., Ltd., Xi’an, China) , equipped with 32 active electrodes. The system specifications included an input impedance of 1 GΩ, input noise <0.4 µV, common-mode rejection ratio (CMRR) of −120 dB, electrode-scalp impedance maintained below 10 kΩ, and a sampling rate of 500 Hz.
As shown in Fig. 1, 32 EEG electrodes were positioned according to the 10-10 system on a custom elastic fabric cap, with the following regional distribution:
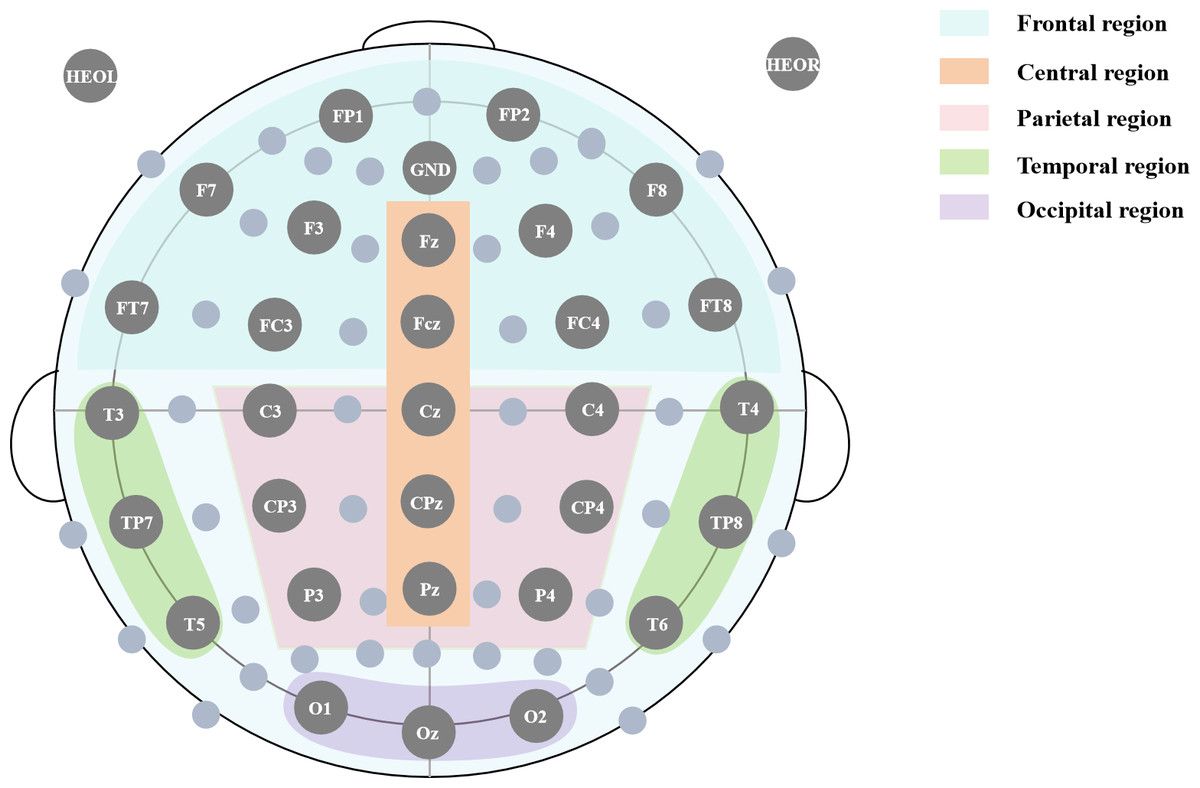
Figure 1: 10-10 International electrode placement system.
{kind=link}
Positioning electrode: GND, Cz
Non-functional electrodes: HEOL, HEOR
Frontal region: FP1, FP2, F3, F4, FC3, FC4, F7, F8, FT7, FT8
Central region: Fz, FCz, Cz, CPz, Pz
Parietal region: C3, C4, CP3, CP4, P3, P4
Temporal region: T3, T4, TP7, TP8, T5, T6
Occipital region: Oz, O1, O2
Experimental design and materials
A 2 (Group: Expert, Novice) × 2 (Memory Process: storage, retrieval) mixed-design experiment was conducted, with the behavioral dependent variable being the participants’ task completion rate and the neural dependent variable being the participants’ neural oscillations in the θ and α frequency bands.
Stimuli consisted of orienteering competition maps (Fig. 2), developed by five certified cartographers (National Senior Cartographer Level). A total of twenty prototype maps were initially created, and the difficulty of the maps was assessed by 100 online volunteers who used a 10-point Likert scale (mean score: 4.55 ± 0.61, indicating moderate difficulty). Ten maps were selected for formal testing, supplemented by two practice maps.
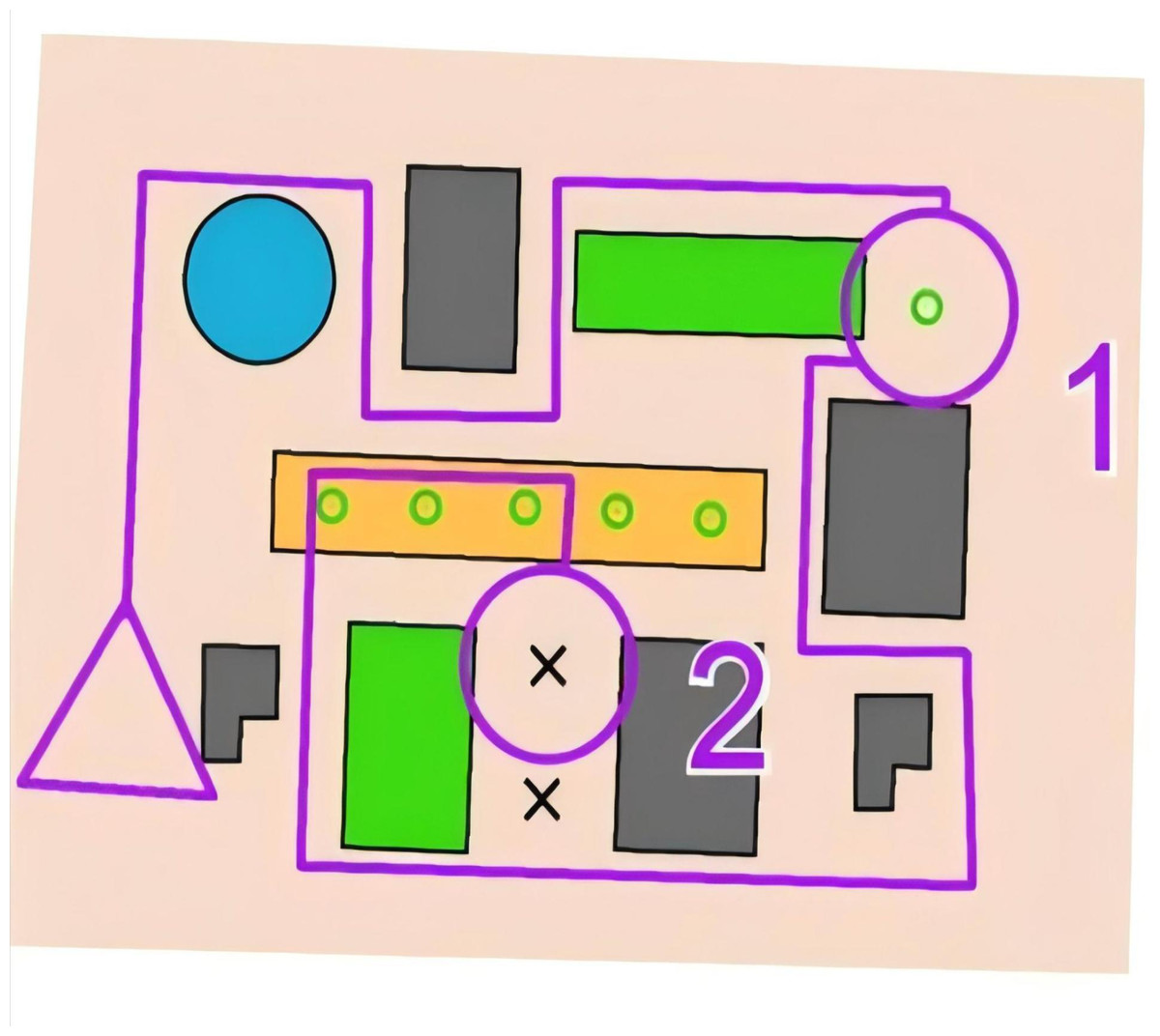
Figure 2: Example of experiment stimulus materials.
{kind=link}
Procedure
Prior to the experiment, participants were asked to complete a self-designed questionnaire documenting their gender, age, years of training, sport grade. During testing, participants were seated 60 cm away from the monitor. They were required to refrain from moving any part of their body throughout the data recording process.
The experiment comprised a practice phase and a formal experimental phase. Participants first completed practice trials (2 test maps) to familiarize themselves with experimental procedure so that they could better control the amplitude of movements and reduce muscle activity in the formal experiment. All materials presented during practice phase were not used in the formal phase, and no data were collected during the practice phase.
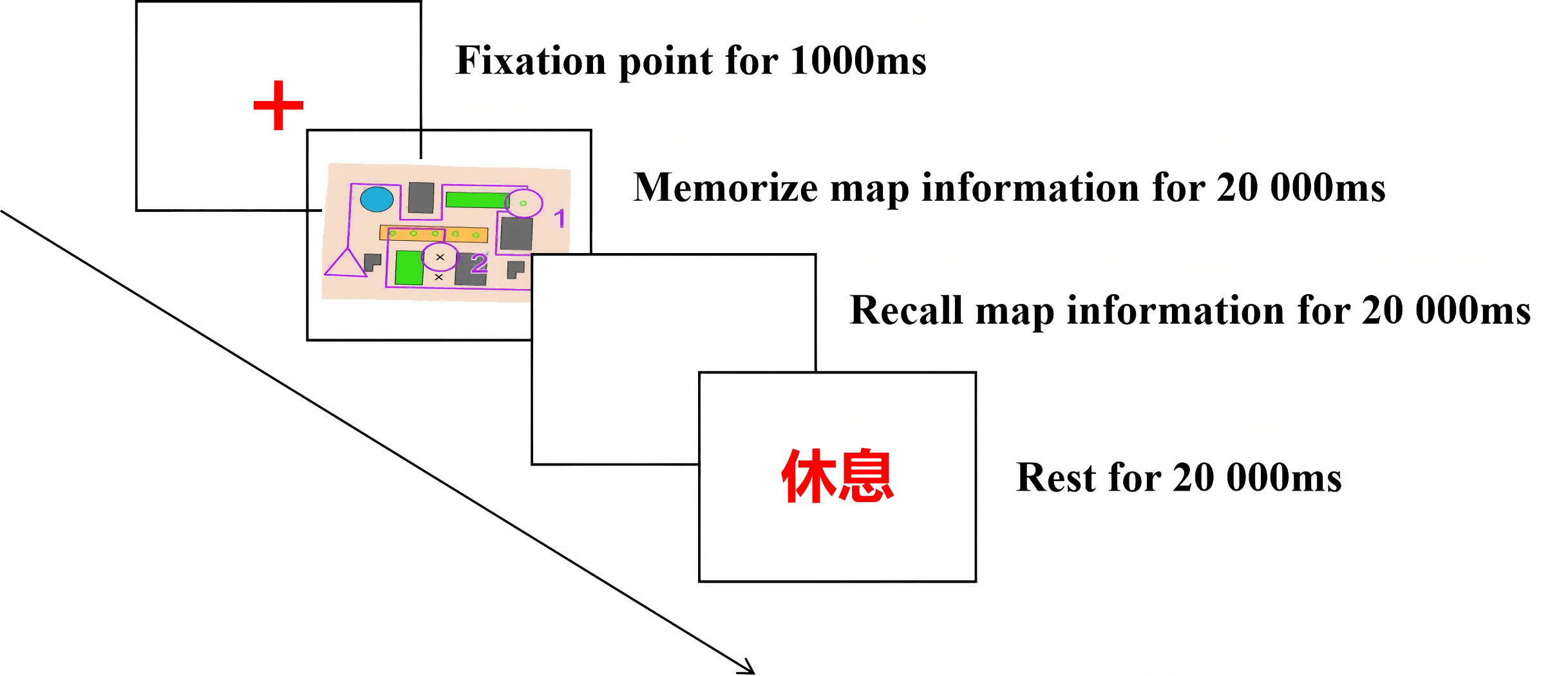
Prior to the formal experiment, three minutes of resting-state EEG data were collected. The formal phase consisted of 10 trials, each comprising two sequential tasks: memory storage and memory retrieval, with maps presented sequentially. As illustrated in Fig. 3, each trial commenced with a central fixation cross (“+” symbol) displayed in the center of the screen. Once the participant’s gaze was fixed on the central point, the experimenter initiated the presentation of the experimental map by pressing a button.Participants were regularly prompted to relax their bodies during the experiment, and the EEG data were monitored in synchronized real-time, and timely guidance was given to adjust the posture or speed of movement when electromyography (EMG) artifacts were detected in order to safeguard the signal quality.
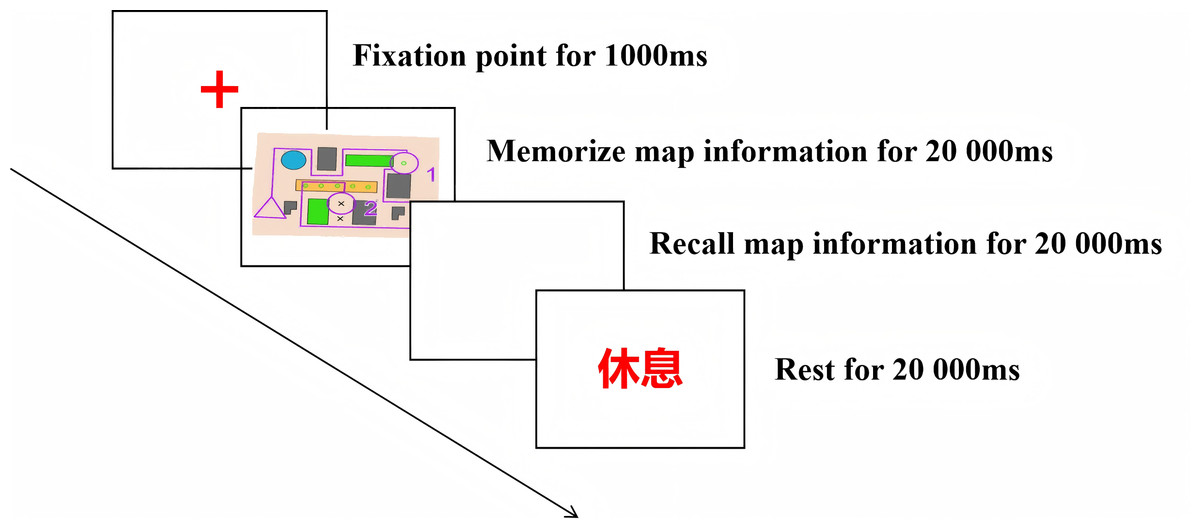
Figure 3: Flowchart of a single trial.
{kind=link}
During the memory storage task, participants observed and memorized map information for 20 s. Following a 20-second intermission, the memory retrieval task required participants to reproduce the previously viewed map on paper, including map features and routes, within 20 s. The trial ended when participants pressed a key upon completion. If no button response was recorded within 20 s, the corresponding data were excluded from subsequent statistical analyses. Each trial concluded with a 20-second inter-trial interval before automatic progression to the next trial, totaling 10 complete trials.
Data analysis
Behavioral data processing and analysis
Three orienteering experts independently assessed participants’ behavioral performance using a 10-point scoring system (5 points for map features and 5 points for routes). The arithmetic mean of the three experts’ scores was calculated as the final behavioral outcome. Behavioral data were systematically organized using Microsoft Excel, and the mean values of all performance metrics across participants were computed as behavioral results.
EEG data preprocessing
EEG data preprocessing was performed using the EEGLAB 2013.1 toolbox (available at: https://eeglab.org/download/). The workflow included the following steps: (1) format conversion, data visualization, and quality assessment; (2) electrode localization and removal of non-functional electrodes (HEOL, HEOR) , retaining 30 electrodes; (3) bandpass filtering (1–40 Hz) combined with 50 Hz notch filtering to eliminate powerline interference; (4) taking into account the sampling rate, the length of the experiment and the reliability of the data analysis results, the data were divided into segments of 2 s in length; (5) interpolated bad channel; (6) re-referencing the data, with the average reference selected for this study; (7) use the ADJUST plug-in to assist in the elimination of artifactual components such as ocular, cardiac, and EMG that are not related to neural activity; (8) artifact removal by rejecting EEG segments exceeding ±70 µV amplitude thresholds; (9) manual visual inspection to ensure data quality. (10) The Welch method is employed for power spectral analysis, where the signal is divided into segments with 50% overlap between consecutive segments. A window function ω is applied to each segment, and the power spectra of all segments are averaged to obtain the overall power spectrum of the signal. Power spectral density (PSD) values were calculated for each channel in the theta band (4–7 Hz) and alpha band (8–13 Hz) for participants during the memory storage and retrieval phases. The electrodes were categorized into frontal, central, parietal, temporal, and occipital regions, and the within-group values were derived by calculating the average power spectral density (PSD) values for each group of electrodes.
Statistical analysis
The processed data were analyzed using SPSS 26.0 statistical software. The normality of the data in each group was assessed using the Shapiro–Wilk test, and all groups were found to conform to a normal distribution (P > 0.05). Outliers were determined by checking if the studentized residuals exceeded ±3 standard deviations, with no outliers detected in any group. Independent samples t-tests were conducted to analyze the behavioral indicators of orienteers with different skill levels. A repeated-measures ANOVA was employed to evaluate the differences in EEG characteristics between expert and novice orienteers during the memory phase. Sphericity assumptions were assessed using Mauchly’s test, with Greenhouse–Geisser correction applied when violations occurred (P < 0.05). When a significant interaction effect was observed, simple effect analyses for each factor were performed using Bonferroni post hoc tests. A p-value < 0.05 in the repeated-measures ANOVA was considered statistically significant.
Results
Behavioral results
An independent samples t-test was used to analyze the behavioral outcomes of orienteers with different skill levels in the map cognition task. As shown in Fig. 4, the expert group demonstrated significantly higher map recall scores than the novice group (t(42) = 7.006, P = 0.000 < 0.001, Cohen’s d = 1.66), and the expert group also exhibited significantly shorter map recall times compared to the novice group (t(42) = −7.629, P = 0.000 < 0.001, Cohen’s d = 2.30). Descriptive statistics are summarized in Table 2.
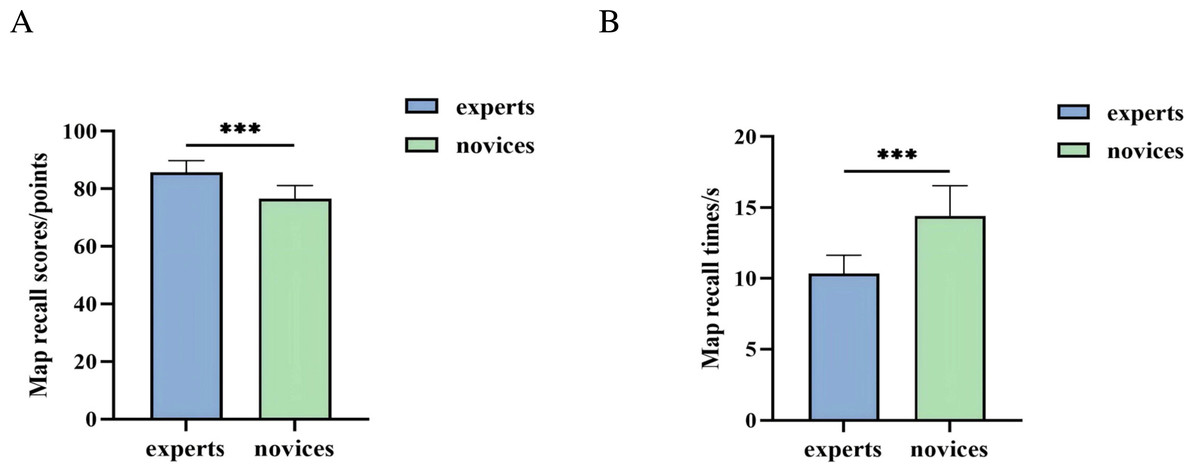
Figure 4: Behavioral performance of orienteers with different skill levels in map cognition task.
(A) Map recall score. (B) Map recall times. (*** represents P < 0.001.)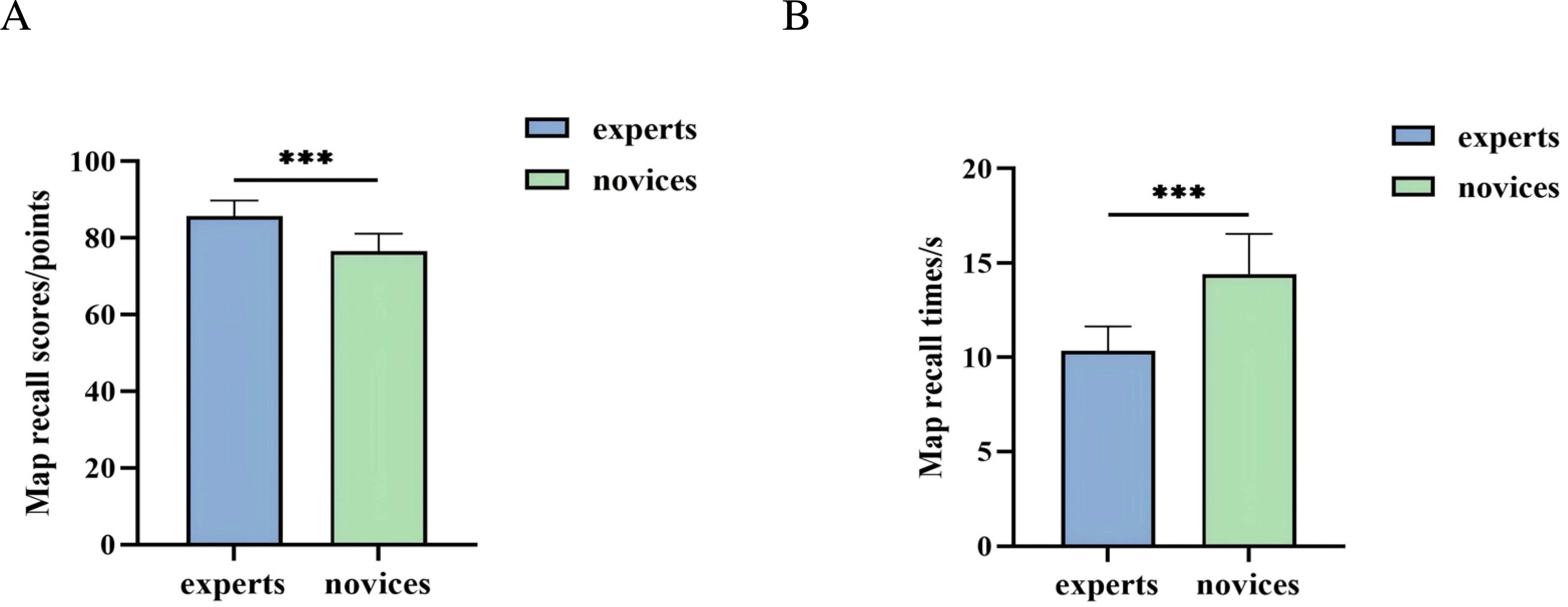{kind=link}
| Expert group | Novice group | |
|---|---|---|
| Map recall scores | 8.57 ± 0.41 | 7.66 ± 0.46 |
| Map recall times | 10.34 ± 1.30 | 14.42 ± 2.14 |
Results of neural oscillations
A repeated-measures ANOVA was conducted to analyze the θ-band neural oscillations during the memory storage and retrieval phases among orienteers of different skill levels. The results, as shown in Fig. 5, revealed a significant main effect of group in the frontal region (F(1, 42) = 4.694, P = 0.036, η2 = 0.101), with the expert group exhibiting significantly lower θ-band neural oscillations in the frontal region during both the memory storage and retrieval phases compared to the novice group. Descriptive statistics are presented in Table 3.
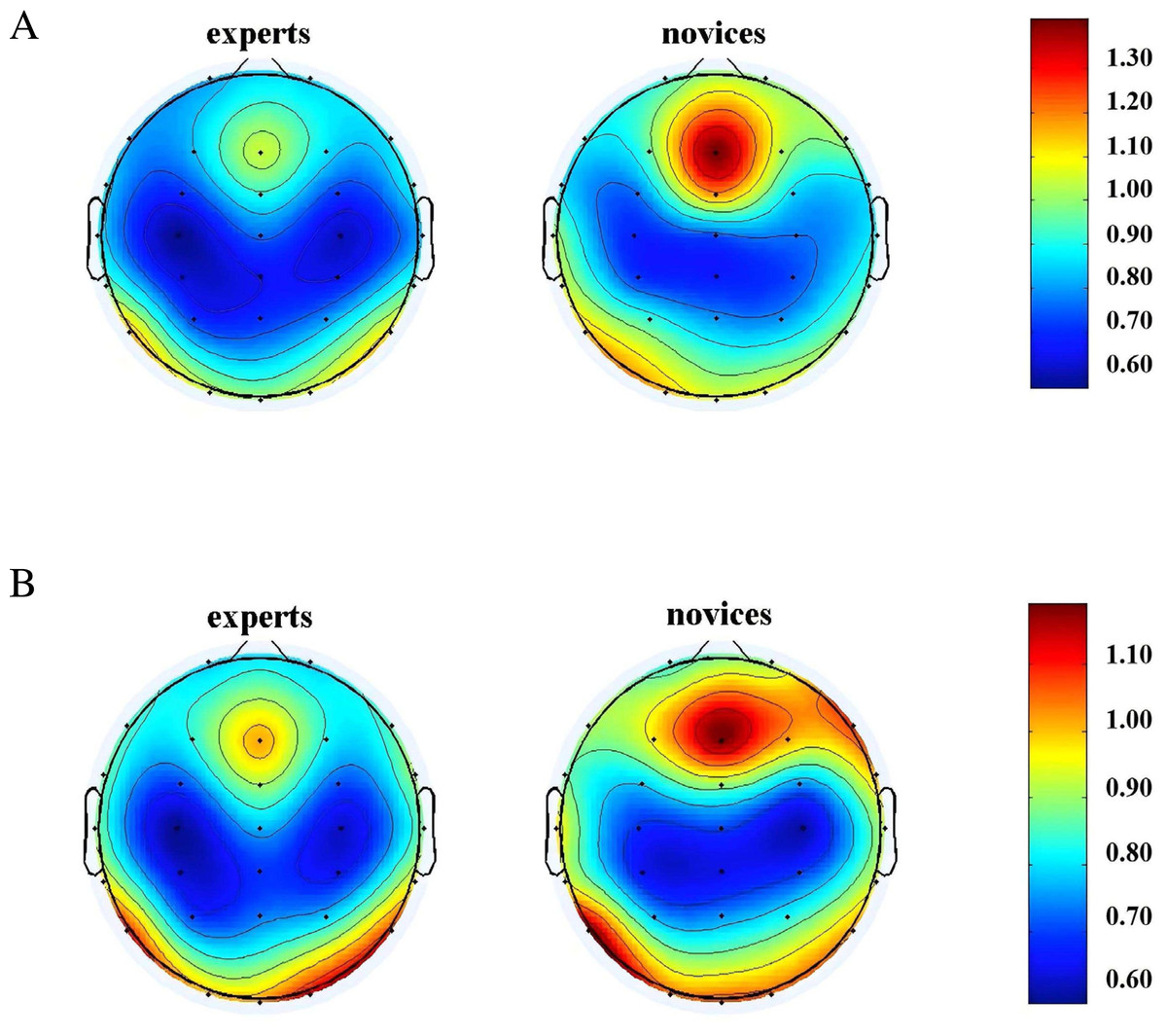
Figure 5: θ-band neural oscillations of orienteers with different skill levels in map cognition task.
(A) Memory storage phase. (B) Memory retrieval phase.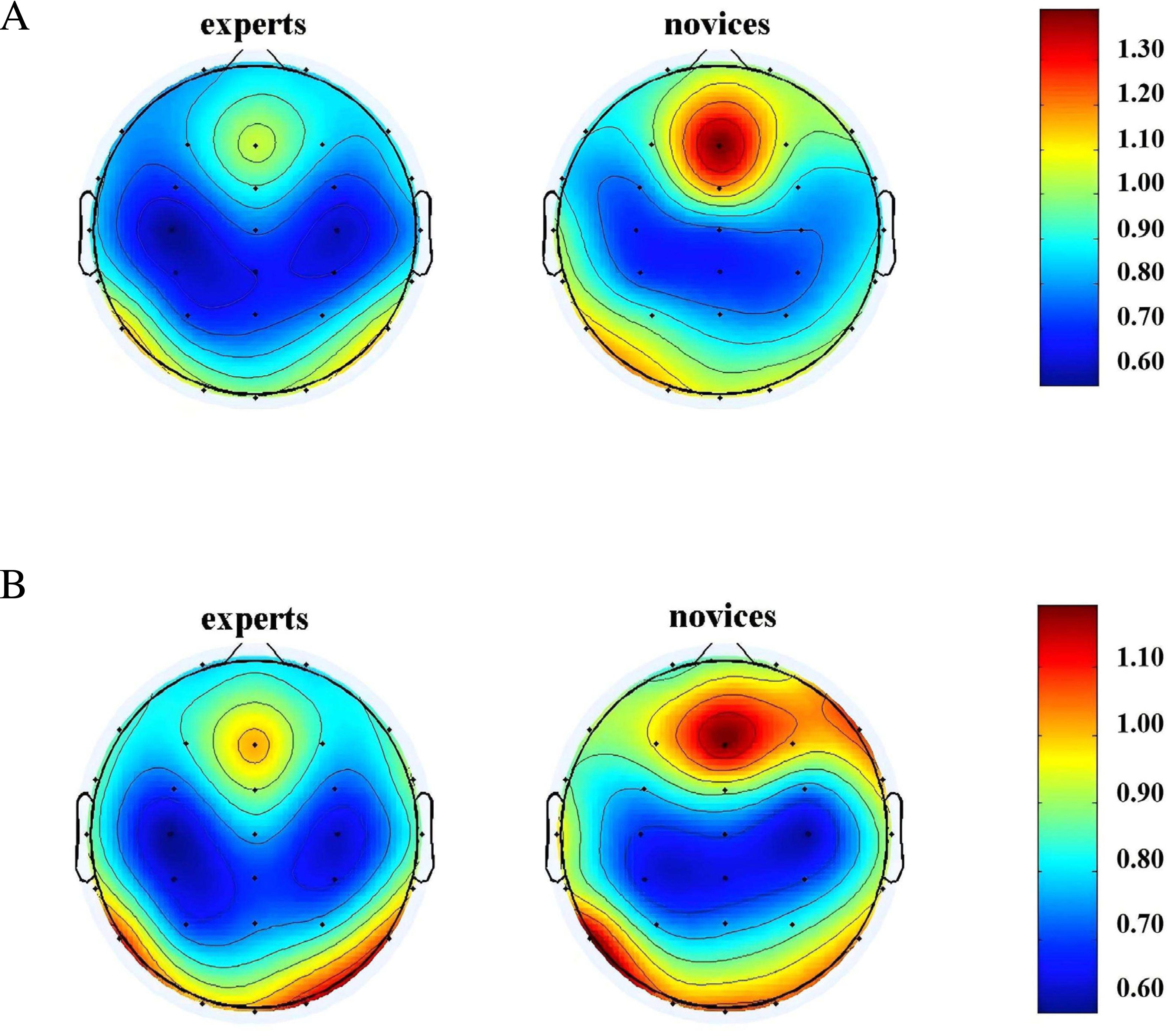{kind=link}
| Expert group | Novice group | |||
|---|---|---|---|---|
| Memory storage (Hz) | Memory retrieval (Hz) | Memory storage (Hz) | Memory retrieval (Hz) | |
| Frontal region | 0.71 ± 0.12 | 0.74 ± 0.14 | 0.79 ± 0.28 | 0.82 ± 0.21 |
| Central region | 0.66 ± 0.13 | 0.66 ± 0.15 | 0.75 ± 0.30 | 0.68 ± 0.17 |
| Parietal region | 0.63 ± 0.13 | 0.67 ± 0.15 | 0.67 ± 0.26 | 0.64 ± 0.10 |
| Temporal region | 0.96 ± 0.21 | 0.99 ± 0.19 | 0.97 ± 0.36 | 0.95 ± 0.20 |
| Occipital region | 0.97 ± 0.35 | 0.91 ± 0.27 | 1.01 ± 0.49 | 0.98 ± 0.36 |
A repeated-measures ANOVA was conducted to analyze the α-band neural oscillations during the memory storage and retrieval phases among orienteers of different skill levels. The results, illustrated in Fig. 6, showed significant main effects of group in the frontal (F(1, 42) = 5.004, P = 0.031 < 0.05, η2 = 0.106) and temporal regions (F(1, 42) = 33.243, P = 0.000 < 0.001, η2 = 0.442). The expert group demonstrated significantly higher α-band neural oscillations in these regions during both the memory storage and retrieval phases compared to the novice group. In the parietal region (F(1, 42) = 9.938, P = 0.003 < 0.01, η2 = 0.191), a significant main effect of memory phase was observed, with both expert and novice groups exhibiting higher α-band neural oscillations in the parietal region during the memory storage phase than during the retrieval phase. Descriptive statistics are summarized in Table 4.
Figure 6: α-band neural oscillations of orienteers with different skill levels in map cognition task.
(A) Memory storage phase. (B) Memory retrieval phase.{kind=link}
Discussion
The ability to rapidly store and retrieve information enables orienteers to respond promptly to dynamic competition environments, swiftly adjust tactical strategies, and gain a competitive edge through agile decision-making. The results of this study demonstrate that expert group performs significantly better than the novice group in map cognition tasks, exhibiting higher levels of rationality and shorter response times. Consistent with findings from Smith et al. (2021), the expert group demonstrates superior memory abilities when processing novel, domain-general stimuli and monitoring spatial changes. In rapid changing sports scenarios, athletes must integrate their motor skills with cognitive and perceptual processes to make rapid and accurate decisions. Expert athletes accomplish visuospatial working memory tasks by combining perceptual information with chunks stored in long-term memory, and this strategic encoding ability is crucial to their enhanced memory performance compared to novice group (Whitaker et al., 2020). Furthermore, in line with the theory of the deliberate practice theory, extensive and focused training allows expert athletes to develop significant cognitive advantages over novices (Macnamara & Maitra, 2019). In orienteering competitions, athletes must rapidly integrate environmental information and make decisions based on prior knowledge and experience. Expert orienteers combine VSWM with spatial cognition and information integration skills to form an efficient and flexible competitive system. Consequently, they demonstrate a marked superiority over novice orienteers in map cognition tasks.
| Expert group | Novice group | |||
|---|---|---|---|---|
| Memory storage (Hz) | Memory retrieval (Hz) | Memory storage (Hz) | Memory retrieval (Hz) | |
| Frontal region | 0.72 ± 0.13 | 0.68 ± 0.09 | 0.63 ± 0.13 | 0.64 ± 0.10 |
| Central region | 0.64 ± 0.14 | 0.63 ± 0.14 | 0.59 ± 0.13 | 0.56 ± 0.06 |
| Parietal region | 0.64 ± 0.12 | 0.59 ± 0.11 | 0.58 ± 0.15 | 0.53 ± 0.08 |
| Temporal region | 0.91 ± 0.14 | 0.91 ± 0.20 | 0.71 ± 0.11 | 0.69 ± 0.10 |
| Occipital region | 0.73 ± 0.15 | 0.76 ± 0.14 | 0.67 ± 0.14 | 0.70 ± 0.14 |
The EEG results revealed that the expert group exhibited significantly lower θ-band neural oscillations in the frontal region compared to the novice group. As demonstrated by Badre & Wagner (2007), the frontal cortex plays a key role in supporting cognitive control and executive functions, helps individuals reactivate and retrieve relevant memory information. According to the neural efficiency hypothesis, experts typically display higher neural efficiency than novices when performing the same tasks, as reflected by reduced activation in task-related brain regions (Ge et al., 2021). θ-wave activity in the cerebral cortex, particularly reduced fluctuations in the frontal region, is linked to optimal athletic performance (Filho, Dobersek & Husselman, 2021). Through prolonged training, expert athletes develop exceptional spatial cognition abilities, enabling them to establish a more efficient spatial information processing mechanism, consistent with these findings. In the process of memory storage, they rapidly identify critical map features including: terrain characteristics and landmark symbols, and effectively match this information to the real-world environment. During memory retrieval, they rapidly retrieve task-relevant details such as terrain features and landmark locations, from memory, which reduces the demand on the frontal lobe.
Additionally, the expert group displayed significantly higher α-band neural oscillations in the central and temporal regions compared to the novice group. In both the expert and novice groups, α-band neural oscillations were higher in the parietal region during the memory storage phase compared to the retrieval phase. This α-band neural activity is associated with specific attentional and spatial perceptual processes (Andrew, Steven & Paul, 2024). Increased α-band oscillations are related to the suppression of irrelevant information (Bastos et al., 2018; Borghini et al., 2018) and tend to increase with higher cognitive load (Paul et al., 2010; Tuladhar et al., 2007). During memory storage, the continuous transfer of information necessitates frequent memory refreshing, leading to heightened activation in brain regions responsible for memory updating and encoding (Liang et al., 2018). During memory retrieval, the memory information stored in the brain prior to neural changes will be activated. The individual needs to focus attention on the retrieved information and collaborate with other brain regions to successfully retrieve relevant memories (Chen et al., 2022). Long-term physical activity can reshape how the brain encodes memories (Jin, Ding & Zhou, 2022; Zhou & Jin, 2021). Compared to novice athletes, expert ones demonstrate significantly enhanced cognitive functions, such as information processing and motor planning, through long-term training. In map cognitive tasks, expert athletes are better at filtering relevant stimuli from irrelevant information, responding to them, and retaining them in memory. This is associated with increased α-band neural oscillations in the central region. The temporal lobe, a key hub for memory processing, plays a vital role in classifying and representing spatial scenes and spatial-related dimensional information, ensuring perceptual accuracy and mitigating interference from similar information (Zeng et al., 2024). Merim, Peter & Fernand (2008) employed functional magnetic resonance imaging (fMRI) to investigate expert-novice differences between experts and novices in chess, and found that chess experts exhibited more pronounced activation in the temporal lobe. Consistent with the findings of this study, expert athletes undergo brain adaptations driven by specialized knowledge and skills, leading to the collaborative engagement of the temporal lobe, which influences memory task performance and ultimately improves decision-making. The parietal cortex plays a role in the processing of perceptual information and the construction of spatial cognition, and can help individuals match current perceptual information with information in long-term memory, thereby triggering memory extraction (Rizza et al., 2024). Alpha-band neural oscillations in the parietal lobe were significantly higher in the memory storage phase than in the memory extraction phase for both the expert and novice groups. The cognitive processing of specialized scenario maps is closely tied to athletes’ unique professional knowledge structure. During memory storage tasks, athletes rapidly retrieve and process critical map features such as shapes, symbols, colors, and legend annotations on maps, with both speed and accuracy. In contrast, during memory retrieval tasks, pre-existing memory information, shaped by long-term training, is activated. Athletes must mobilize cognitive resources to regulate distracting interference information (Fan & Zheng, 2019), which results in an increased cognitive load and elevated α-band neural oscillations in the parietal cortex.
Distinct neural optimizations and adaptations are reflected in the differences in brain activity among orienteers of varying skill levels during map cognition.These differences not only highlight variations in their map cognition abilities but also offer valuable insights for future training and interventions.Through targeted training approaches, the map cognition performance of novice orienteers can be improved.
Conclusion
This study, based on the B&H model and the expert-novice paradigm, examined the cognitive features of VSWM processing in orienteers of different skill levels. Key findings include: expert orienteers demonstrated superior map cognition with higher rationality and shorter response times; the expert group were able to efficiently utilize neural resource, characterized by decreased θ-band oscillations in the frontal lobe and enhanced α-band processing in the frontal and temporal regions. These results not just deepen our understanding of the neural mechanisms underlying expert performance in orienteering but also provide robust theoretical support for practical application of orienteering training.
Limitations and Future Directions
This study investigated the cognitive features of VSWM processing in orienteers of different skill levels; however, several limitations should be acknowledged. The specific constraints of EEG experimental paradigms may compromise ecological validity, and the exclusive use of sport-specific map scenarios as experimental stimuli potentially limits the generalizability of findings. In the future, virtual simulation technologies to develop more ecologically valid VSWM tasks still need to be explored. Expanding the experimental design to include both sport-specific and non-sport-specific contexts would enable a more comprehensive understanding of the temporal dynamics underlying VSWM processing in orientees. Such methodological improvements could provide deeper insights into the cognitive mechanisms specific to this athletic population while enhancing the practical applications of findings in training program development.