
Bacteria associated with human saliva are major microbial components of Ecuadorian indigenous beers (chicha)
- Published
- Accepted
- Received
- Academic Editor
- Hauke Smidt
- Subject Areas
- Anthropology, Food Science and Technology, Microbiology, Infectious Diseases
- Keywords
- Lactic acid bacteria, Indigenous beer, Fermentation, Chicha, Microbiota, Artisanal fermented beverages, Streptococcus salivarius, Streptococcus mutans, Lactic acid bacteria, Fermented cassava, Ecuador, Chewed indigenous beer, Cassava, Saliva
- Copyright
- © 2016 Freire et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. Bacteria associated with human saliva are major microbial components of Ecuadorian indigenous beers (chicha) PeerJ 4:e1962 https://doi.org/10.7717/peerj.1962
Abstract
Indigenous beers (chicha) are part of the indigenous culture in Ecuador. The fermentation process of these beers probably relies on microorganisms from fermented substrates, environment and human microbiota. We analyzed the microbiota of artisanal beers (including a type of beer produced after chewing boiled cassava) using bacterial culture and 16S ribosomal RNA (rRNA) gene-based tag-encoded FLX amplicon pyrosequencing (bTEFAP). Surprisingly, we found that Streptococcus salivarius and Streptococcus mutans (part of the human oral microbiota) were among the most abundant bacteria in chewed cassava and in non-chewed cassava beers. We also demonstrated that S. salivarius and S. mutans (isolated from these beers) could proliferate in cassava mush. Lactobacillus sp. was predominantly present in most types of Ecuadorian chicha.
Introduction
The domestication of fermenting bacteria and yeast predated the domestication of animals and plants; ancestral hominids adapted to metabolize alcohol long before the Neolithic period (Carrigan et al., 2015). The organoleptic and psychotropic effects associated with the consumption of accidentally fermented fruits or cereals may have motivated early humans to replicate this process. Additionally, fermentation may have provided unintended benefits as fermenting bacteria may have reduced the risks of foodborne diseases in ancient societies (Nakamura et al., 2012; Lewus, Kaiser & Montville, 1991; Fooks & Gibson, 2002; Tesfaye, Mehari & Ashenafi, 2011); it is still unclear whether these microorganisms confer additional health benefits (McNulty et al., 2011). The use of alcoholic beverages has played a crucial role in the evolution of human societies (Joffe, 1998); nevertheless, very little is known about the process of domestication and evolution of these fermenting microorganisms (Libkind et al., 2011).
Many fermenting microorganisms have originated in the environment and food substrates (Martini, 1993) while others resemble microorganisms found in the human microbiome, suggesting human (skin or intestine) origins (Agapakis & Tolaas, 2012); in fact, some modern fermented dairy products contain intestinal bacteria (Walter, 2008).
Indigenous people from South America (such as in Ecuador) prepare a type of beer known as chicha which is made with either corn, boiled cassava or the fruit of the palm Bactris gasipaes (chonta); some cassava beers include an additional chewing step before the fermentation process. A recent report showed that bacteria present in chewed cassava beers were mainly Lactobacillus sp. (Colehour et al., 2014). We analyzed the microbial diversity (using culture dependent and culture independent techniques) in different types of Ecuadorian chicha.
Materials and Methods
Sample collection
Four samples of chicha (indigenous beer) from two geographical regions of Ecuador (Andean and Amazon regions) were collected. These samples included beer made with both chewed cassava (CC), mushed cassava (MC); mushed chonta (CB) and ground corn (CoB) (Table 1). The samples of CC and MC were purchased from the same household. All these products were obtained from rural communities. None of these beers were pasteurized, nor had they any commercial additives or preservatives. All samples were refrigerated (2–8 °C) after collection; a 2 mL aliquot of sample was stored at −20 °C, for molecular phylotyping.
| Main ingredient | Substrate scientific name | Geographical region | Site of collection | Time of fermentation |
|---|---|---|---|---|
| Chewed cassava | Manihot esculenta | Amazon | Puyo | 3 days |
| Mushed cassava | Manihot esculenta | Amazon | Puyo | 3 days |
| Chonta | Bactris gasipaes | Amazon | Tena | 2 days |
| Corn (jora) | Zea mays | Highlands | Pifo | 2 days |
Plate count of lactic acid bacteria (LAB)
A 20 mL aliquot of each sample was homogenized in 180 mL of a sodium citrate solution (10−1 dilution) and ten-fold dilutions were made in saline solution (NaCl 0.9%). One mL of each dilution was inoculated in MRS (pH 5) and M17 (pH 7, 0.5% dextrose) by pour plate method. Two incubation temperatures were used (37 and 43 °C) under aerobic and anaerobic conditions, for 3–5 days. The incubation time varied because of the different bacteria present on each product.
Phenotypic characterization
Ten colonies (showing different morphology) were randomly picked from MRS plates from each sample. A subset of colonies showing characteristics of lactic acid bacteria (oxidase negative, catalase negative, Gram positive rods or cocci) was selected for molecular characterization (Table 2); 5 from CC, 6 from MC, 6 from CB and 8 from CoB. Strains were stored at −20 °C in MRS or M17 broth with 20% of glycerol.
| Sample | Isolate ID | Culture media | Growth condition | Identification (16S ribosomal RNA gene) |
|---|---|---|---|---|
| Chewed cassava beer | 25 A2 | MRS | Anaerobic | Leuconostoc mesenteroides |
| 25 C2 | MRS | Aerobic | Lactobacillus fermentum | |
| 25 E2 | M17 | Anaerobic | Streptococcus mutans | |
| 25 F1 | M17 | Aerobic | Lactococcus lactis | |
| 25H1 | M17 | Aerobic | Streptococcus salivarius | |
| Mushed cassava beer | 26 A1 | MRS | Anaerobic | Lactobacillus fermentum |
| 26 B1 | MRS | Anaerobic | Lactobacillus fermentum | |
| 26 C2 | MRS | Aerobic | Lactobacillus fermentum | |
| 26 E2 | M17 | Anaerobic | Streptococcus salivarius | |
| 26 F2 | M17 | Anaerobic | Streptococcus salivarius | |
| 26 G1 | M17 | Aerobic | Streptococcus salivarius | |
| Chonta beer | 27 A1 | MRS | Anaerobic | Lactobacillus plantarum |
| 27 B1 | MRS | Anaerobic | Weissella confusa | |
| 27 C1 | MRS | Aerobic | Weissella confusa | |
| 27 E1 | M17 | Aerobic | Lactococcus lactis | |
| 27 F2 | M17 | Anaerobic | Lactococcus lactis | |
| 27 G2 | M17 | Aerobic | Lactococcus lactis | |
| Corn beer | 61 B2 | MRS | Anaerobic | Lactobacillus casei |
| 61 G1 | M17 | Anaerobic | Leuconostoc mesenteroides | |
| 61 G2 | M17 | Anaerobic | Lactobacillus plantarum | |
| 61 H1 | MRS | Anaerobic | Lactobacillus parabuchneri | |
| 61 I1 | MRS | Anaerobic | Lactobacillus paracasei | |
| 61 J1 | MRS | Anaerobic | Lactobacillus pantheris | |
| 61 K1 | M17 | Anaerobic | Leuconostoc mesenteroides | |
| 61 L1 | M17 | Anaerobic | Leuconostoc mesenteroides |
Genotypic characterization of bacterial colonies
DNA was extracted from one colony using DNAzol Reagent (Life Technologies, Carlsbad, CA, USA) following manufacturer instructions and the DNA was stored at −20 °C until used. The 16S ribosomal RNA gene was amplified in 25 uL containing: 1X PCR buffer, 2.5 mM MgCl2, 0.25 mM dNTP’s, 0.2 uM 27F primer (5′-AGAGTTTGATCCTGGCTCAG-3′), 0.2 uM 1492R primer (5′-GGTTACCTTGTTACGACTT-3′) (Martin-Laurent et al., 2001), 0.5 U GoTaq Flexi DNA polymerase (Promega, Madison, WI, USA), 5 uL of sample DNA and Milli-Q water. The times and temperatures used for the amplification were: melting (94 °C, 1 min), annealing (56 °C, 30 s), elongation (72 °C, 30 s), this routine was repeated for 30 cycles, and final extension (72 °C, 10 min). Amplicons were subjected to gel electrophoresis (1% agarose gel), sequenced at Functional Biosciences (Madison, WI, USA) and DNA sequences analyzed using Seqmatch (Ribosomal Database Project: http://rdp.cme.msu.edu/) and submitted to GenBank; the accession numbers are KT722809–KT722833).
High throughput sequencing analysis
In order to complement the culture-based protocols, we investigated the microbial diversity using FLX amplicon pyrosequencing. DNA was extracted from all beer samples using DNeasy Plant Mini kit (Qiagen) following manufacturer’s protocols, but instead of using AE buffer for elution, we used same volume of PCR Milli-Q water. DNA samples from four types of beer were sent to CD Genomics (Shirley, NY, USA), for 16S-based phylotyping. DNA was subjected to bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP) using primers 939F-5′TTGACGGGGGCCCGCAC3′ and 1492R-5′TACCTTGTTACGACTT3′. For fungal sequences we used ITSF-5′CTTGGTCATTTAGAGGAAGTAA3′. Resulting sequences (minimum length = 250 nucleotides) were trimmed and quality scored using USearch (http://drive5.com/); chimeras were detected using UCHIIME (http://drive5.com/) in de novo mode and were compared using BLASTn to a ribosomal database. Identity values were used to make assignments to the appropriate taxonomic levels: greater than 97% identity were resolved at the species level and between 95 and 97% at the genus level. The number of bacterial sequences we obtained were: 2,965 readings for CC, 3,320 for MC, 3,046 for B and 15,623 for CoB. For fungi we obtained 6,763 readings from CC, 6,925 from MC and 6,558 from CB. We did not carry out fungi analysis of CoB. All sequences were submitted to Sequence Read Archive and accession numbers are: SRP070493, SRS1299611, SRX1612367, SRR3202831, SRS1299612, SRX1612366, SRR3202830, SRS1299613, SRX1612365, SRR3202829, SRS1310202, SRX1600290, SRR3187397, SRS1310203, SRX1600289, SRR3187396, SRS1310204, SRX1600288, SRR3187395, SRS1310207, SRX1612364, SRR3202828, SRS1310208, SRX1600292, and SRR3202832.
Streptococcus salivarius and Streptococcus mutans growth in cassava solution
To rule out the possibility of S. salivarius or S. mutans contamination, one colony of a pure culture of each bacteria (obtained from beers) was diluted in 25 mL of sodium citrate (2%) separately. Subsequently, 1 mL of this cell suspension was used to inoculate tubes containing 9 mL of sterile (autoclaved) chewed cassava solution (10%) and incubated at 37 °C under anaerobic conditions. A 100 μL aliquot from each incubated tube was extracted and plated in M17 (this was done by triplicate) at 0, 24, 48 and 72 h of inoculation. Results from each day were compared to determine the ability of these bacteria to grow in chewed cassava solution.
Statistical analysis
We used Mann-Whitney U test to test whether S. salivarius and S. mutans were able to grow in cassava solution. Shannon indices were calculated using the formula H = −Σpilog(pi), pi being the relative frequency of the abundance of each species found. Principal component analysis (PCA) of the bacterial species and abundance of the four beverages was performed using the software SPSS v21 (IBM Corp, Armonk, NY, USA).
Results
Characterization of bacterial isolates
Twenty-five bacterial isolates (cultured from the four beer types) were characterized by 16S rDNA sequencing showing 99–100% identity when compared with GenBank sequences (Table 2). The predominant bacterial species in all beers were Lactobacillus fermentum (16%), Lactococcus lactis (16%), Leuconostoc mesenteroides (16%), and Streptococcus salivarius (16%); followed by Lactobacillus plantarum (8%), Weissella confusa (8%), Lactobacillus casei (4%), Lactobacillus pantheris (4%), Lactobacillus parabuchneri (4%), Lactobacillus paracasei (4%) and Streptococcus mutans (4%). The most diverse bacterial composition (using culture-dependent techniques) found in CoB (6 bacterial species), followed by the CC (5 bacterial species), CB (3 bacterial species) and MC (2 bacterial species). Intriguingly, cassava beers contained human salivary bacteria: both CC and MC had Streptococcus salivarius while CC had also S. mutans (Table 2).
High throughput sequencing analysis
The beer with greater diversity was CC (31 bacterial species), followed by CoB (26 bacterial species), CB (21 bacterial species), MC (20 bacterial species). The predominant bacterial species in CC were Lactobacillus spp. (40.9%) followed by human microbiota bacteria: Streptococcus salivarius (31.94%), Streptococcus parasanguinis (5.41%), Streptococcus pneumoniae (3.65%). The most prevalent bacteria in MC were Streptococcus spp. (83%) followed by Lactococcus sp. (9.32%); the majority of streptococci have been described as part of the human microbiota: Streptococcus salivarius (65%), Streptococcus pasteurianus (7.74%), and Streptococcus parasanguinis (3.47%). The most prevalent bacteria in CB were Weissella confusa (46%), Weissella sp. (20%), and Lactococcus lactis (9%). The dominant bacteria in CoB were Weissella sp. (19%) and Lactobacillus plantarum (12.5%), Lactococcus garviae (2.76%) Lactobacillus brevis (2.5 %) (Table 3). The dominant fungal species present in different beers analyzed was very similar; Saccharomyces cerevisiae was the most abundant comprising 92% of all the taxa detected (Table 4).
| Bacterial species | CC | MC | CB | CoB | Cultured | Possible origins |
|---|---|---|---|---|---|---|
| Bacillus amyloliquefaciens | 0.0 | 0.5 | 0.0 | 0.00 | – | Environment |
| Carnobacterium maltaromaticum | 0.0 | 0.0 | 1.0 | 0.1 | – | Environment |
| Enterobacter asburiae | 0.5 | 0.0 | 0.0 | 0.0 | – | Environment |
| Enterobacter cancerogenus | 0.5 | 0.0 | 0.0 | 0.0 | – | Environment |
| Enterobacter sp | 1.3 | 0.0 | 0.1 | 0.0 | – | Environment |
| Fructobacillus sp | 0.0 | 0.0 | 0.0 | 3.8 | – | Vegetables |
| Gluconacetobacter intermedius | 0.0 | 0.0 | 0.0 | 0.6 | – | Fermented food |
| Kluyvera ascorbate | 0.4 | 0.0 | 0.0 | 0.0 | – | Human gut, food |
| Lactobacillus brevis | 8.4 | 0.1 | 2.5 | 0.6 | – | Environment, gut |
| Lactobacillus camelliae | 0.0 | 0.0 | 0.0 | 7.3 | – | Environment, gut |
| Lactobacillus casei | 0.0 | 0.0 | 0.0 | 3.1 | + | Environment, gut |
| Lactobacillus delbrueckii | 8.0 | 0.0 | 0.0 | 0.0 | – | Environment, gut |
| Lactobacillus fermentum | 6.5 | 3.8 | 0.0 | 0.0 | + | Environment, gut |
| Lactobacillus harbinensis | 0.0 | 0.0 | 0.0 | 2.1 | – | Vegetables |
| Lactobacillus manihotivorans | 1.8 | 0.0 | 0.0 | 0.0 | – | Vegetables |
| Lactobacillus parabuchneri | 0.0 | 0.0 | 0.0 | 1.4 | + | Oral microbiota |
| Lactobacillus paracasei | 0.0 | 0.0 | 0.0 | 8.6 | + | Environment, gut |
| Lactobacillus paracollinoides | 0.0 | 0.0 | 0.0 | 16.0 | – | Environment, gut |
| Lactobacillus plantarum | 10.8 | 0.0 | 12.4 | 0.1 | + | Environment, gut |
| Lactobacillus sp | 3.4 | 0.0 | 0.7 | 1.3 | – | Environment, gut |
| Lactobacillus vaccinostercus | 1.2 | 0.0 | 0.2 | 0.0 | – | Environment, gut |
| Lactococcus garviae | 0.0 | 0.0 | 2.8 | 0.0 | – | Fermented food |
| Lactococcus lactis | 2.1 | 0.0 | 8.9 | 0.0 | + | Environment, gut |
| Lactococcus sp | 0.2 | 9.3 | 1.0 | 0.2 | – | Gut |
| Leuconostoc citreum | 0.0 | 1.5 | 1.2 | 0.0 | – | Fermented food |
| Leuconostoc lactis | 1.7 | 0.1 | 0.2 | 0.8 | – | Environment |
| Leuconostoc sp | 0.0 | 0.0 | 0.1 | 4.6 | – | Vegetables |
| Oenococcus kitaharae | 0.0 | 0.0 | 0.0 | 1.2 | – | Vegetables |
| Serratia sp | 1.0 | 0.0 | 0.0 | 0.0 | – | Environment |
| Streptococcus gallolyticus | 0.0 | 0.5 | 0.0 | 0.0 | – | Oral microbiota |
| Streptococcus oralis | 1.4 | 0.2 | 0.0 | 0.0 | – | Oral microbiota |
| Streptococcus parasanguinis | 5.4 | 3.5 | 0.0 | 0.0 | – | Oral microbiota |
| Streptococcus pasteurianus | 0.0 | 7.7 | 0.0 | 0.0 | – | Human gut |
| Streptococcus pneumoniae | 3.6 | 0.5 | 0.0 | 0.0 | – | Human nasopharynx |
| S. pseudopneumoniae | 0.5 | 0.0 | 0.0 | 0.0 | – | Human nasopharynx |
| Streptococcus salivarius | 32.0 | 65.0 | 0.0 | 0.0 | + | Oral microbiota |
| Streptococcus sp | 2.5 | 2.3 | 0.1 | 0.0 | – | Human microbiota |
| Streptococcus thermophilus | 1.2 | 2.59 | 0.0 | 0.0 | – | Vegetables |
| Streptococcus vestibularis | 0.4 | 0.8 | 0.0 | 0.0 | – | Oral microbiota |
| Weissella cibaria | 0.1 | 0.0 | 0.9 | 0.9 | – | Vegetables |
| Wbconeissella confusa | 0.5 | 0.1 | 45.9 | 25.3 | + | Vegetables |
| Weissella paramesenteroides | 0.5 | 0.0 | 0.1 | 0.0 | – | Environment |
| Weissella sp | 0.2 | 0.3 | 19.8 | 19.4 | – | Vegetables |
Note:
Chewed cassava, CC; mushed cassava, MC; chonta, CB; corn, CoB. Numbers indicate percentages and “+” indicates that bacterium recovered in culture.
| Fungal species | CC | MC | CB | Possible origins |
|---|---|---|---|---|
| Saccharomyces cerevisiae | 92.533 | 92.023 | 92.033 | Vegetables |
| Penicillium citrinum | 0.03 | 0.021 | 0.062 | Soil |
| Debaryomyces hansenii | 0.636 | 0.547 | 0.549 | Sea water |
| Hanseniaspora uvarum | 0.044 | 0.056 | 0.075 | Vegetables |
| Wallemia muriae | 0.118 | 0.115 | 0.137 | Salty water |
| Wallemia sp | 1.316 | 1.701 | 1.602 | Salty water |
| Aspergillus sp | 0.089 | 0.047 | 0.032 | Soil |
| Pichia kudriavzevii | 1.05 | 1.5 | 1.32 | Vegetables |
| Aspergillus versicolor | 0.104 | 0.138 | 0.135 | Soil |
| Pichia burtonii | 0.118 | 0.123 | 0.107 | Vegetables |
| Hyphopichia burtonii | 0.089 | 0.067 | 0.073 | Starch substrates |
| Cyberlindnera sp | 0.532 | 0.54 | 0.545 | Waste deposits |
| Pichia sp | 0.044 | 0.04 | 0.054 | Soil |
| Saccharomyces bayanus | 0.104 | 0.132 | 0.096 | Vegetables |
| Galactomyces sp | 3.149 | 2.908 | 3.133 | Rumen, fermented food |
| Pichia fermentans | 0.044 | 0.042 | 0.047 | Vegetables |
Note:
Chewed cassava, CC; mushed cassava, MC; chonta, CB. The numbers indicate percentages.
Growth of S. salivarius and S. mutans in cassava solution
Streptococcus salivarius (Fig. 1) and S. mutans (Fig. 2) grew in chewed cassava solution. After 48 h of culture (S. salivarius) and 72 h (S. mutans), the bacterial counts went down.
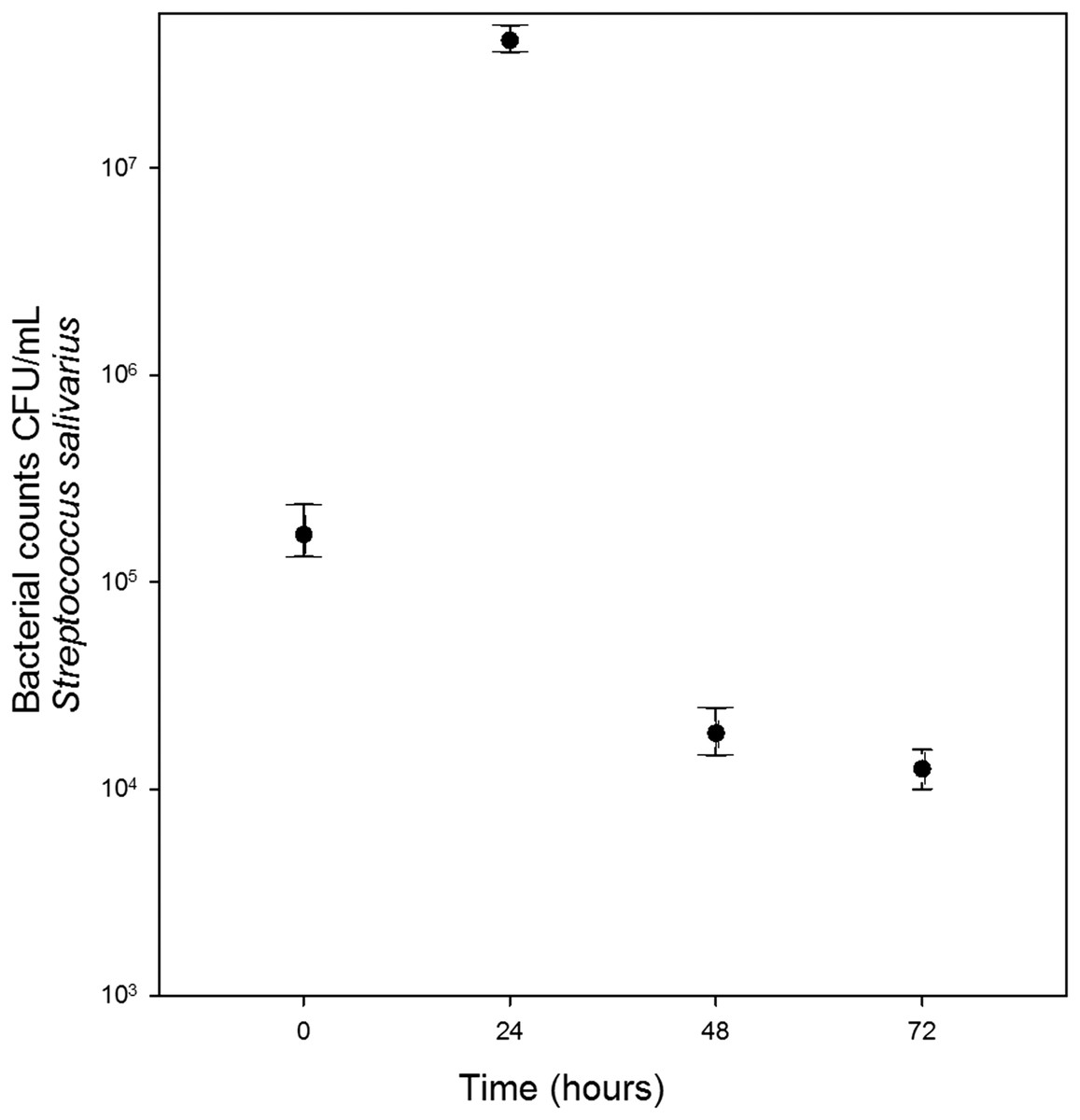
Figure 1: Growth of S. salivarius in sterile chewed cassava solution.
There is a significant increase in CFU (Mann-Whitney U test) at the 24 h of incubation compared with those at inoculation time (0 h).{kind=link}
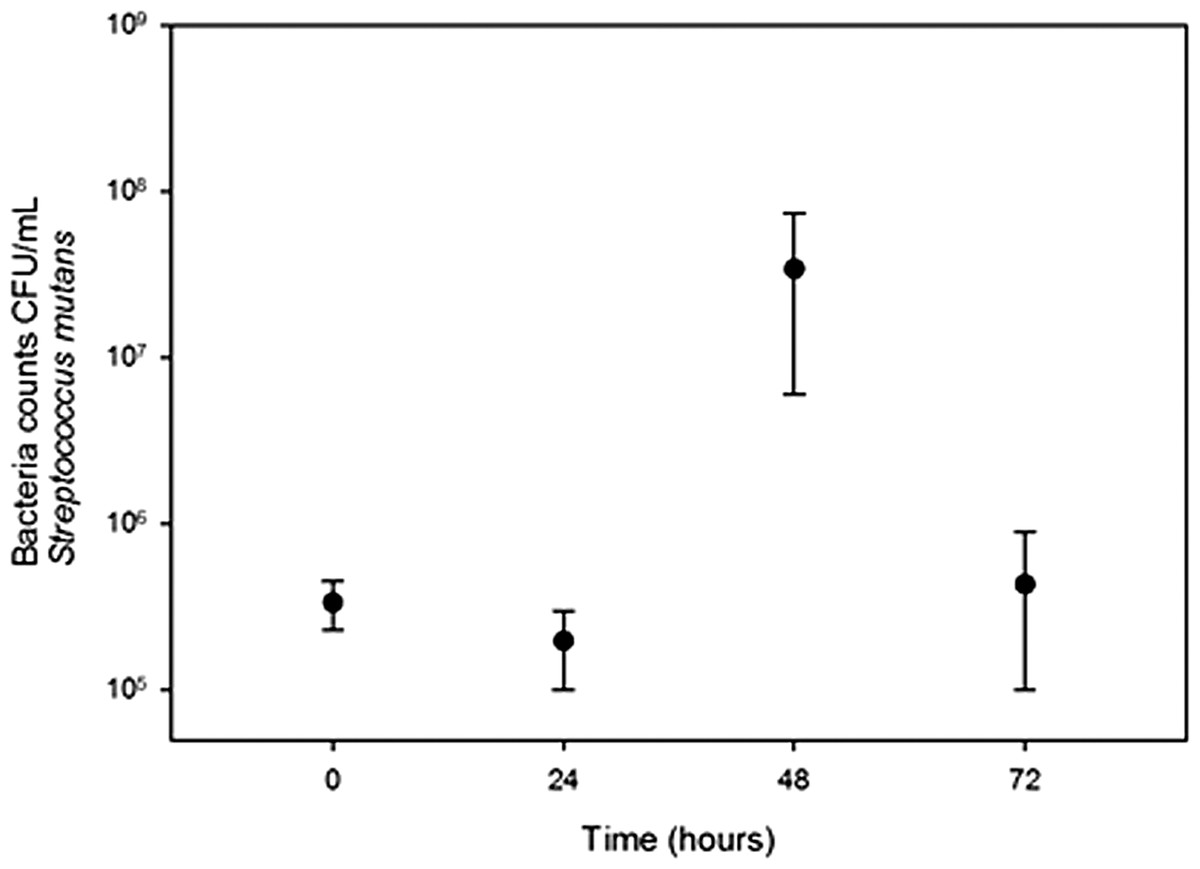
Figure 2: Growth of S. mutans in chewed cassava solution.
There is a significate increase in CFU (Mann-Whitney U test) at the 48 h of incubation compared with those at the inoculation time (0 h).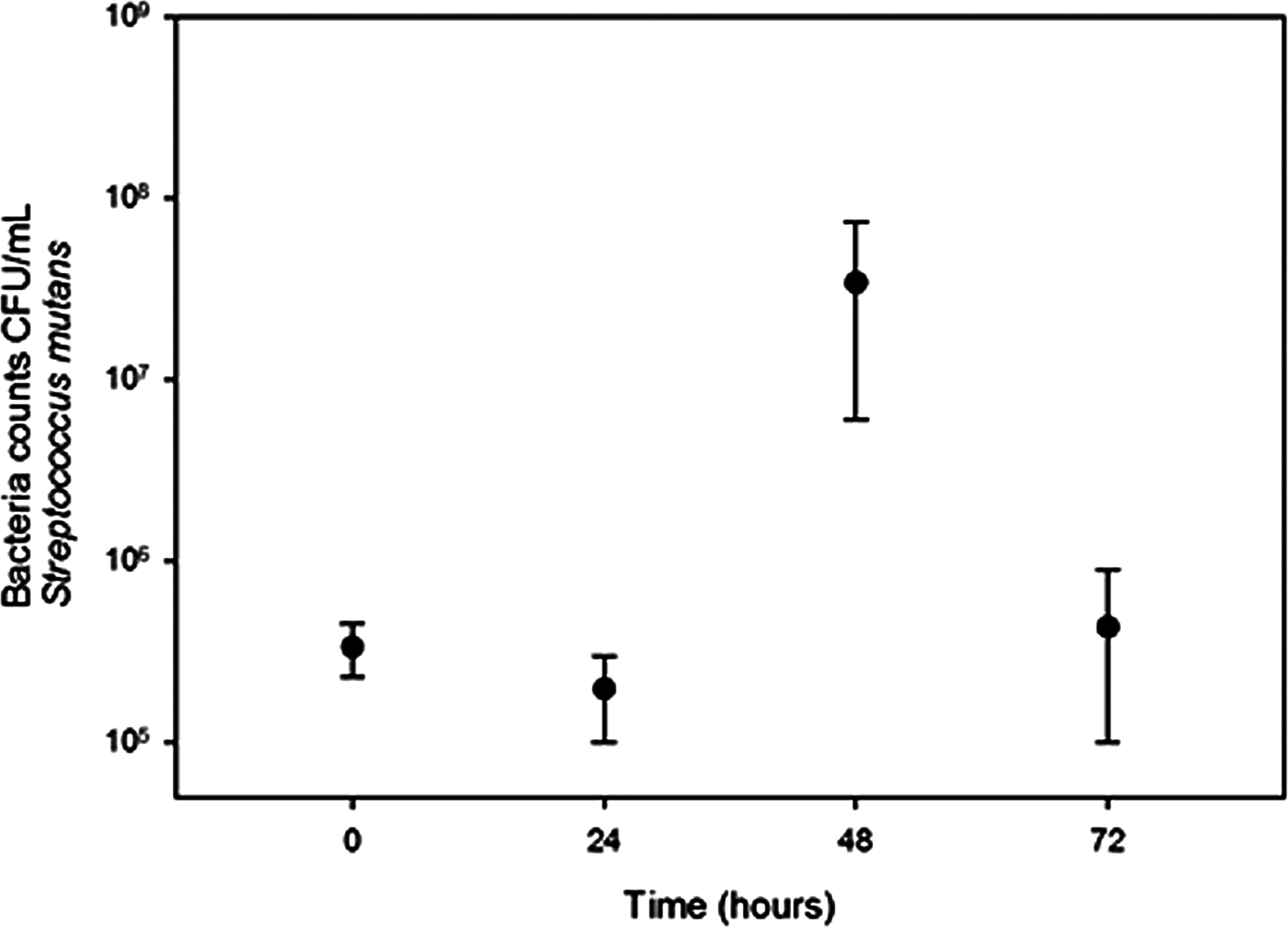{kind=link}
Diversity estimations
CC was the beverage with the most species diversity (H = 1.06, E = 0.71), followed by CoB (H = 0.94, E = 0.66), CB (H = 0.71, E = 0.54), and MC (H = 0.59, E = 0.45). The evenness values followed the same pattern and suggest that CC is also the most heterogeneous in terms of species (Hayek & Buzas, 2010; Pielou, 1966).
Principal component analysis
The type of beer (fermented substrate) accounted for 90.4% of the bacterial species variability and cassava beers had more similar bacterial composition and abundance than the other types of beer; interestingly CB and CoB also showed similarity (Fig. 3).
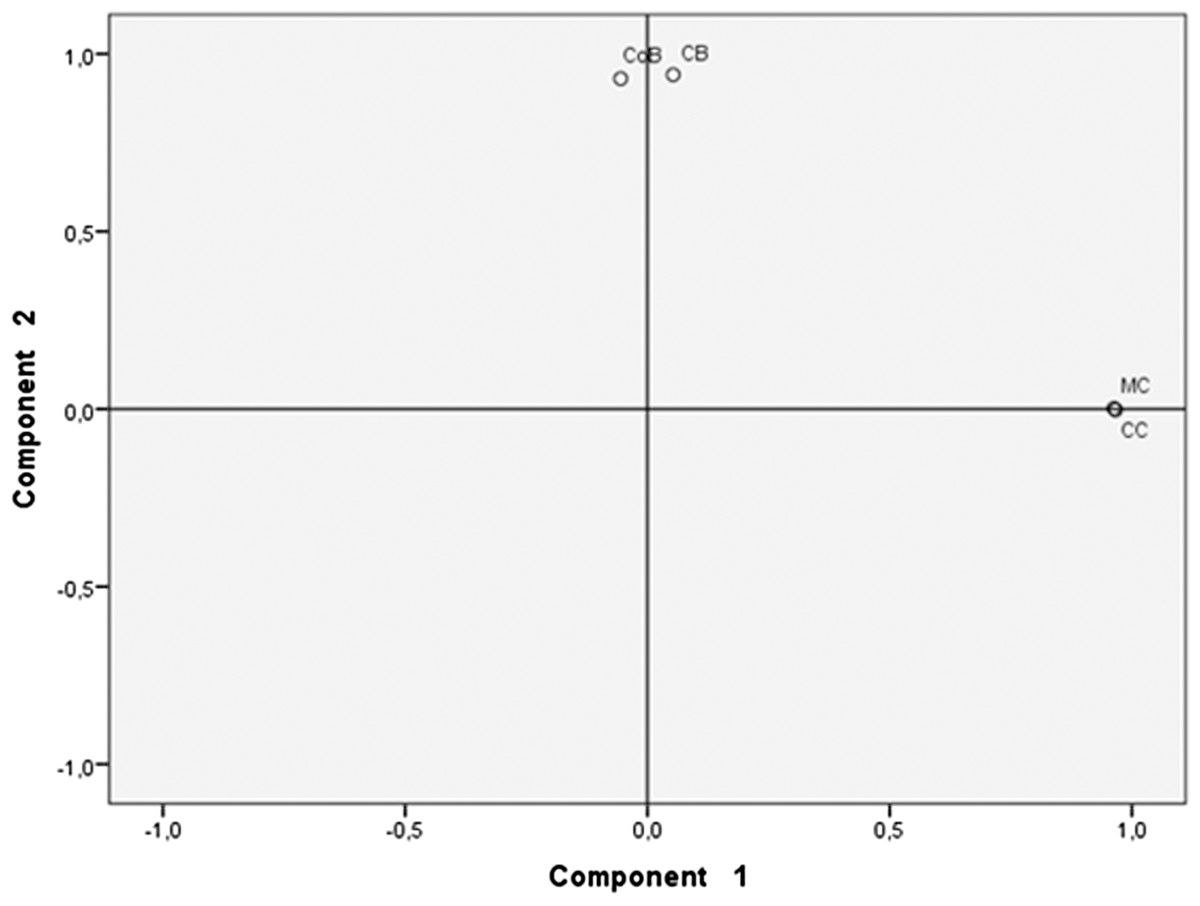
Figure 3: Principal component analysis of beers’ microbiota.
Beers made with cassava (MC and CC) formed a cluster different from the cluster formed by beers made with either chonta (CB) or corn (CoB). Each pair of beverages that form a group share a similar bacterial species profiles and abundance.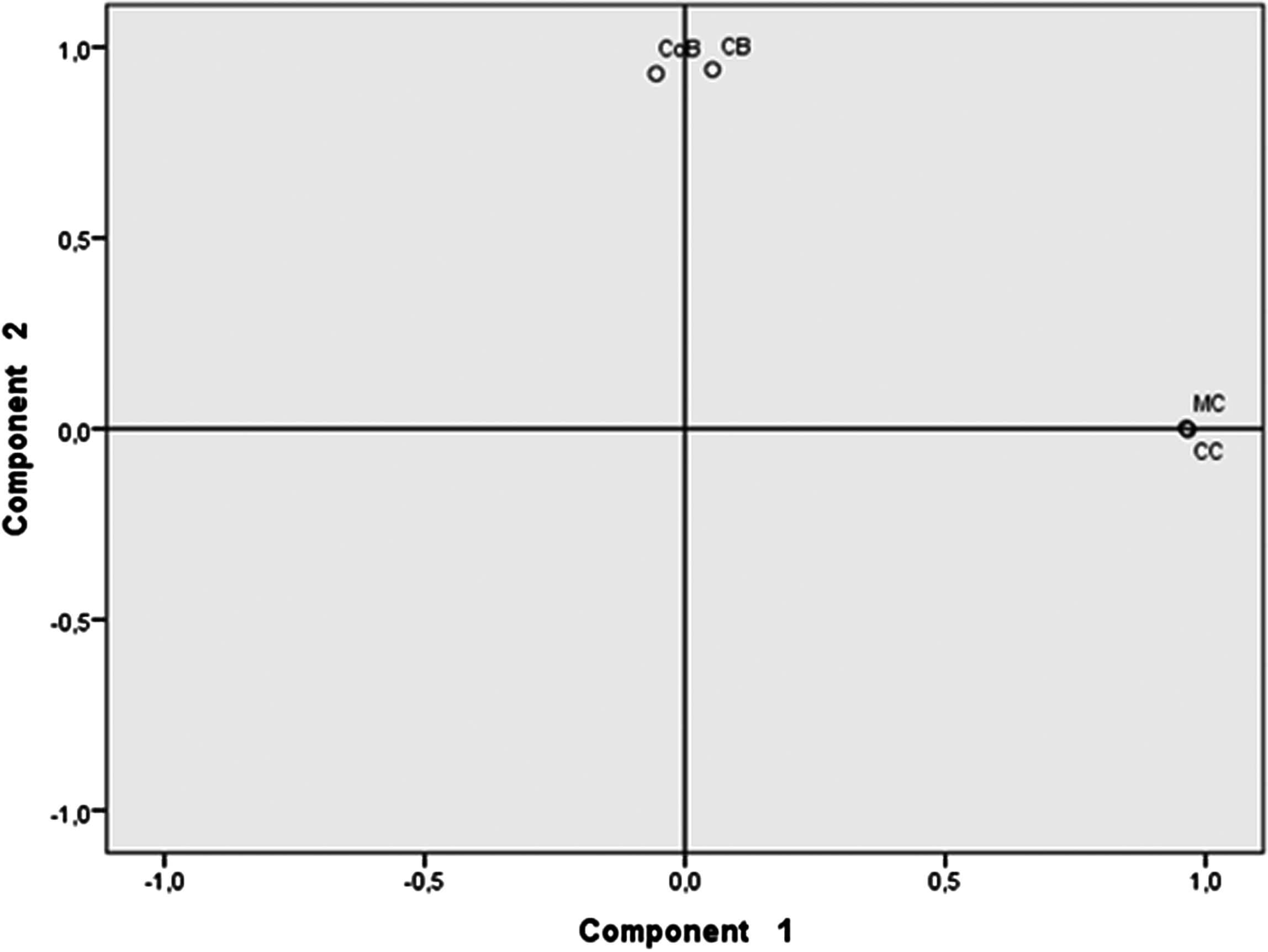{kind=link}
Discussion
Our study found higher bacterial diversity in beer that contained human saliva (Tables 2 and 3); therefore, saliva may not only speed up the fermentation process (by providing amylases as suggested by Henkel (2005)) but may also offer an additional bacterial inoculum which may favor this process. This finding may provide additional explanation for the adoption of such a peculiar process in the beer’s manufacture.
Our study also demonstrated the presence of oral streptococci such as S. salivarius, S. mutans, S. parasanguinis in cassava beers; these bacteria may thrive on carbohydrates present in the oral cavity after starchy meals (Moye, Zeng & Burne, 2014; Burne, 1998). Oral bacteria S. salivarius and S. mutans were cultured from cassava chicha (with saliva and without saliva) in large numbers and were shown to grow in mushed cassava under laboratory conditions. Oral bacteria in beer without human saliva may indicate contamination of fermenting containers (or utensils). Fermenting bacteria are known to produce biofilm in containers (Kebede et al., 2007) and both types of cassava beers were obtained from the same household, and most likely they use the same pots for both type of beers. It is possible that some strains of S. salivarius from these beers may be adapting to the fermentation process; Streptococcus thermophilus, a bacteria used as starter in yogurt (Burton et al., 2006) may have evolved from S. salivarius (Hols et al., 2005). Future studies should investigate the prevalence of S. salivarius in larger number of cassava chichas from other locations and find out whether the strains of S. salivarius isolated from beers are different from those isolated from human saliva.
A recent study failed to detect S. mutans and S. salivarius in chicha prepared with chewed cassava in Ecuador (Colehour et al., 2014). The disagreement between both studies may result from differences in samples in both studies; Colehour et al. (2014) collected beers that were fermenting for four days while we collected samples that were fermenting for three days. Beer microbiota changes overtime (Steinkraus, 2002) and in the case of S. mutans and S. salivarius we observed a sharp increase and decline in bacterial populations in 24 h (Figs. 1 and 2). Unlike Colehour et al. (2014), we also carried out bacterial cultures.
Reduction on streptococci populations may be due to the consumption of all the nutrients, accumulation of toxic metabolites, autolysis (Dufour & Lévesque, 2013). Also, these bacteria are known to form biofilm (Ajdić et al., 2002; Li et al., 2002) which may change bacterial location and reduction of planktonic cells. Additionally, unlike our study Colehour et al. (2014) found predominance of L. reuteri which is known to antagonize S. salivarius (Nikawa et al., 2004). Similar to previous studies (Colehour et al., 2014; Elizaquível et al., 2015; Puerari, Magalhães-Guedes & Schwan, 2015), Lactobacillus was a dominant genus of lactic bacteria in chicha found in both culture dependent and independent assessments.
Our study complements previous microbiological analyses carried out in chicha and shows for the first time the potential adaptation of S. salivarius, S. mutants (and possibly other streptococci from the human upper respiratory tract) to grow in cassava mush. The study not only shows how bacteria from human microbiota may adapt to artisanal fermentative processes but also shows that chewed chicha may potentially transmit human pathogens such as S. mutans, one of the causative agents of dental plaque and cavities (Loesche, 1986); Streptococcus mutans can be transmitted person-to-person, most likely through saliva (Baca et al., 2012). This is especially relevant because these types of beers are consumed as early as two or three days after preparation.
The main limitation of our study was the low number of samples analyzed of each beer. However this limitation does not invalidate the main findings of this study. Additionally, the culture medium (MRS) is not suitable to culture Lactobacillus from cereals (Minervini et al., 2012), therefore we may have underestimated the bacterial diversity in these beers.