
Combined effect of temporal inundation and aboveground-cutting on the growth performance of two emergent wetland plants, Phragmites australis and Bolboschoenus planiculmis
- Published
- Accepted
- Received
- Academic Editor
- Meredith Root-Bernstein
- Subject Areas
- Biodiversity, Conservation Biology, Ecology, Freshwater Biology, Natural Resource Management
- Keywords
- Brackish wetland, Food plant, Mesocosm experiment, Mortality, Vegetation management, Water level, Wetland restoration
- Copyright
- © 2024 Park et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Combined effect of temporal inundation and aboveground-cutting on the growth performance of two emergent wetland plants, Phragmites australis and Bolboschoenus planiculmis. PeerJ 12:e18402 https://doi.org/10.7717/peerj.18402
Abstract
Background
Phragmites australis is a common foundation species found in inland and brackish estuarine ecosystems. P. australis stands provide a wide range of habitats for wetland organisms and perform essential functions, such as nutrient cycling, pollutant filtration, wave energy reduction, and soil stabilization. However, excessive growth of P. australis can degrade the quality of wetland habitats, thereby reducing the functions of restored wetlands.
Methods
In this study, we investigated the effectiveness of vegetation management techniques, such as aboveground cutting and temporal inundation with varying depth and periodicity, in controlling the growth performance of P. australis and adjacent vegetation, Bolboschoenus planiculmis. Differences in growth responses to manipulated inundation stress between P. australis and B. planiculmis were measured.
Results
Inundation stress of 10–50 cm caused significantly greater inhibition of growth performance in B. planiculmis compared to P. australis. The combination of aboveground-cutting and inundation treatments resulted in a significant inhibitory effect on the growth performance and survival rate of P. australis. The growth performance of P. australis, including stem volume and biomass, decreased and its mortality rate increased. Our manipulated experiment suggests a combined treatment approach of moderate inundation, such as 5–10 cm for 20–30 days, and aboveground cutting to manage the overgrowth of P. australis in restored brackish wetlands.
Introduction
Over the past century, rapid land conversion, reclamation, and urban expansion have resulted in a significant loss of natural wetland habitats. Various types of wetland restoration procedures have been undertaken as mitigation strategies (Zedler & Kercher, 2005). Nevertheless, it is frequently reported that even after restoration projects are carried out to enhance the degraded function and condition of wetland ecosystems, the desired wetland condition could not be maintained for the expected periods. This site management problem can occur when restoration plans do not adequately reflect the core ecological processes required to restore wetland function (e.g., Gupta et al., 2020; Keddy, 2010; Mitsch & Gosselink, 2015; Thomaz, 2023; van Biervliet et al., 2020; Wang et al., 2019; Zarekarizi, Srikrishnan & Keller, 2020). In addition, poor management and maintenance practices after a restoration project may result in the loss of desired ecological functions and their recovery. For example, restored wetlands can degrade rapidly due to changes in habitat structure, increased sedimentation rates resulting from an altered hydrological regime, or the expansion of invasive species. Changes in the physical habitat structure and hydrological regime of restored wetlands is closely related to the level of species richness and the plant community composition (Wang et al., 2016). A deeper understanding of the key response traits of restored plant communities is critical for designing proper management measures to achieve long-term restoration goals (An et al., 2022).
Phragmites australis (common reed) is an important foundation species that provides diverse ecological roles in estuarine ecosystems (Dolinar et al., 2016). P. australis tends to establish clonal populations storing large amounts of plant biomass and providing complex microhabitats and food sources for local communities, supporting the regional food web structure (Čížková et al., 2023). However, rapid expansion of P. australis can outcompete native vegetation such as Bolboschoenus planiculmis. This can lead to a reduction in overall biodiversity and an imbalance in associated ecosystem functions. In addition, invasive genotypes of P. australis have been observed in Europe and North America. Their rapid spread and growth characteristics alter the structure and function of natural and restored wetland ecosystems (Hazelton et al., 2014; van der Putten, 1997). Uncontrolled and rapid expansion of P. australis communities can simplify wetland habitat diversity, such as vegetation type, and reduce the richness of species that can thrive in wetland vegetation (Greet & King, 2019).
The rapid expansion of P. australis including its native and invasive strains across many reed-dominated wetlands poses a global management problem that scientists have tried to tackle with various methods, including physical removal (e.g., cutting or fire; Güsewell, Le Nédic & Buttler, 2000; Kimura & Tsuyuzaki, 2011; Russell & Kraaij, 2008; Thompson & Shay, 1985), biological control (Blossey et al., 2020), chemical control (Schad et al., 2021), and water level control (Widin, Bickford & Kowalski, 2023). While some strategies, such as chemical control, have been shown to be effective in preventing further spread of P. australis (Bonello & Judd, 2020; Robichaud, Basso & Rooney, 2022; Yuckin et al., 2023), their use is sometimes controversial in the field due to concerns about safety, cost and chemical regulation in protected wetlands (see primary use of herbicide; Martin & Blossey, 2013; Quirion et al., 2018). Reports of the remaining methods that have been tested are inconsistent, and may vary depending on the wetland type of the managed site, competing plant species, and environmental characteristics. Warren et al. (2001) showed that a single treatment is ineffective, but mixed treatments (e.g., herbicide and mowing) can be more effective in the long-term perspective. Widin, Bickford & Kowalski (2023) also presented effective control of the P. australis population using a combined method of cutting and submergence, while the potential adverse effects on the other wetland plants were not tested. Given these complex outcomes, experts suggest that efficient management of P. australis requires the development of site-specific methods tailored to local environmental factors.
P. austalis beds (i.e., native strain) in restored wetlands on Eulsuk Island (Republic of Korea) provide supporting services to various wetland species, including the Oriental Great Reed Warbler (Acrocephalus orientalis), Leopard cat (Prionailurus bengalensis), Brown sesarmid crab (Chiromantes dehaani), and diverse wetland insect species (Busan Metropolitan City, 2022). However, the recent rapid expansion of P. australis is accelerating ecological succession from wetland vegetation to terrestrial vegetation, simplifying habitat types in the restored wetland. Several management trials have attempted to remove the overgrown P. australis using heavy machinery, but failed due to the rapid regrowth from the remaining rhizomes, spending a significant proportion of the management costs (Busan Metropolitan City, 2022). Continued expansion of P. australis can further threaten the loss of B. planiculmis, which is one of the primary food sources for migratory birds during the winter season. Given the current state of the site, there is an urgent need to develop a sustainable way to control the expansion of P. australis.
In this study, we conducted manipulated plant growth experiments near the restored wetland site on Eulsuk Island. The pot experiments setup in the restored wetland evaluated the effects of two types of management measures (inundation and aboveground cutting) on two emergent wetland species, P. australis and B. planiculmis. We included B. planiculmis in the control experiment to check for any negative effects of management measures on other native vegetation that plays an important role in the restored wetland system. Wetland inundation (e.g., water level control) is a relatively easy management measure to apply in restored wetlands with floodgates. Aboveground removal cannot be applied over large areas due to limited access to restored wetlands and higher costs, but it can limit the expansion of plant communities in a short term. Our main objectives were (1) to evaluate the growth performance of P. australis and B. planiculmis with different levels of inundation and aboveground cutting, and (2) to examine changes in tuber production of B. planiculmis under different inundation stress used to control P. australis. Based on the manipulated experiments, we try to propose an optimal management plan for the most effective use of the two methods (i.e., inundation and aboveground cutting) to control P. australis expansion without negatively affecting other vegetation in the restored wetlands.
Materials and Methods
Study site
The pot experiment and plant collection were conducted in the Nakdong River estuary (NRE, N35°06′13.81″E128°56′46.81″), Republic of Korea (Figs. 1A, 1B). The study site has a humid subtropical climate based on the Köppen climate classification (Busan Metropolitan City, 2022). The annual average temperature is 14.6 °C. The annual rainfall is approximately 1,354 mm. Over half of the annual precipitation falls during the summer monsoon season (i.e., June to August). The NRE has been designated as Natural Monument (No. 179), Wetland Protection Area, and Nature Reserve due to its ecological significance as a wintering habitat for migratory waterfowl and its distinctive biodiversity.

Figure 1: Map of the experiment site (A, B) and mesocosm tanks for inundation experiment (C, D).
(C) Constant water level tank, (D) mesocosm tank during the inundation experiment. Different numbers indicate the adjusted water depth in different tank sections. Map source credit in (A): Positron basemap (https://github.com/CartoDB/basemap-styles?tab=readme-ov-file) provided by ©CARTO and ©OpenStreetMap contributors. Map source credit: © Google maps.{kind=link}
Plant materials
Two dominant emergent plant species, P. australis and B. planiculmis, were used in the NRE pot experiments. P. australis, also known as common reed, is a perennial wetland plant of the Poaceae family that grows to 1–3 m. P. australis is found in a wide range of habitat from inland to brackish coastal environments. In the Nakdong River Estuary, the P. australis community is established along the mid-high tide line of the tidal marsh, sandbars, and elevated sand dunes (Busan Metropolitan City, 2022). Recent studies in other continents have reported aggressive behavior of invasive P. australis (e.g., haplotype M in North America; Lindsay et al., 2023; Saltonstall, 2002) by expanding its distribution or replacing native subspecies. Majority haplotypes of P. australis in the Republic of Korea were identified as the P types that is the dominant form in East Asia and Australia (Chu et al., 2011).
B. planiculmis is a tuberous wetland plant belonging to the perennial Cyperaceae family and can grow to a height of 0.4–0.9 m. The B. planiculmis community is also distributed along the mid to high tide line of the tidal marsh, partly overlapping with P. australis. Both plant species are clonal plants with a guerrilla architecture that has a strong ability to spread laterally due to its greater inter-ramet distance. The guerrilla architecture provides advantages for foraging in heterogeneous environments (Xue et al., 2018). The annual production of tuberous roots of B. planiculmis is regionally important in the NRE and the East Asian Australasian Flyway, as many waterfowl species, such as Cygnus cygnus (Whooper swan) and Anser fabalis (Bean goose), use it as a carbohydrate source during the winter season at stopover sites (Kim et al., 2013, 2016; Kim & Kim, 2021).
We collected all plant material from a nearby population established near the restored wetland in the NRE. To establish plant clones for our manipulation experiments, we first collected the rhizomes of juvenile P. austalis plants from the restored wetland in April 2010 (Fig. 1B). We cut the collected rhizomes to a length of 10 cm in with a similar wet weight. Each rhizome had a single node producing stem and roots. Prepared rhizomes were planted in near-surface sediment (2~3 cm deep) in plastic pots that were 10 cm in diameter and 15 cm in height. Each pot was filled with sediment that we collected from the tidal marsh at NRE. The collected sediment was sieved through a 2 mm filter to remove other plant material prior to use. The texture of the collected soil was classified as sandy loam. P. austalis rhizomes were grown in soil saturated with water from the restored wetland. In October 2009, B. planiculmis tubers were collected from the NRE and stored in wet condition inside a refrigerator (4 °C). As the growth rate of juvenile plants can be influenced by different tuber sizes, we screened tubers of the same size (i.e., length: 2 cm). In April of the following year, we planted selected tubers of B. planiculmis in plastic pots and kept them under saturated soil conditions for 2 weeks to facilitate tuber sprouting. All plant clones of the wetland species were grown under the same condition for 2 months before the manipulation experiments.
Pot experiments
Two mesocosm tanks measuring 6 m in length, 4 m in width, and 1 m in depth, were placed on the shore of a restored wetland in Eulsuk Island, Nakdong River Estuary (coordinates: N 35.1039°, E 128.9464°; EPSG 4326). The aforementioned restored wetland is located near the estuarine barrage and is connected to the primary river channel by two sluice gates. Therefore, the average salinity of the restored wetland remains brackish except during prolonged periods of flooding after the summer monsoon. The mesocosm tanks were filled with water from the restored wetland. The salinity of the tank water was monitored, and maintained at a brackish condition between 4–8 PSU throughout the experiment periods. Tank water was changed once at the end of inundation treatment to prevent algal blooms in the tank.
One experimental tank was used to maintain the constant water level condition during the initial growth period and the last 90-days before harvest (Fig. 1C). The other tank was used to manipulate four different water level conditions (i.e., 0, 10, 30, 50 cm depth from the soil surface) using aluminium blocks of different heights (Figs. 1D, 2). The manipulated water level conditions did not fully submerge the stem of P. australis. After 60 days of initial growth period, pots of P. australis and B. planiculmis were then moved to four different water level conditions mixed with four different inundation periods (i.e., 0, 10, 20, 30 days; Figs. 2A, 2B). To compare the growth performance of P. australis under combined stressors, we placed P. australis pots, one of which used stem-cutting as a mechanical management option, in the restoration wetlands. There were eight replicate pots for P. australis, including a stem-cutting group, and 10 replicate pots for B. planiculmis in each manipulation experiment group. In total, the P. australis experiment had 160 pots (i.e., control group: 16 pots, manipulated group: cut [2 levels] × depth [3 levels] × inundation period [3 levels] × 8 replicates) and B. planiculmis experiment had 100 pots (i.e., control group: 10 pots, manipulated group: depth [3 levels] × inundation period [3 levels] × 10 replicates). We randomly placed the replicate pots within the manipulated depth ranges in the tanks. At the end of inundation treatments, all pots were moved to the initial tank and the water level was adjusted to sediment saturation level as in the control group pots. Our experimental procedure mimics raising and maintaining the water level for 30 days to make plant inundation condition in the restored wetland, and then lowering the water level to average condition for the remainder of the growing season. To minimize the potential differences from pot position, we randomly rotated the pot position every 2 months.
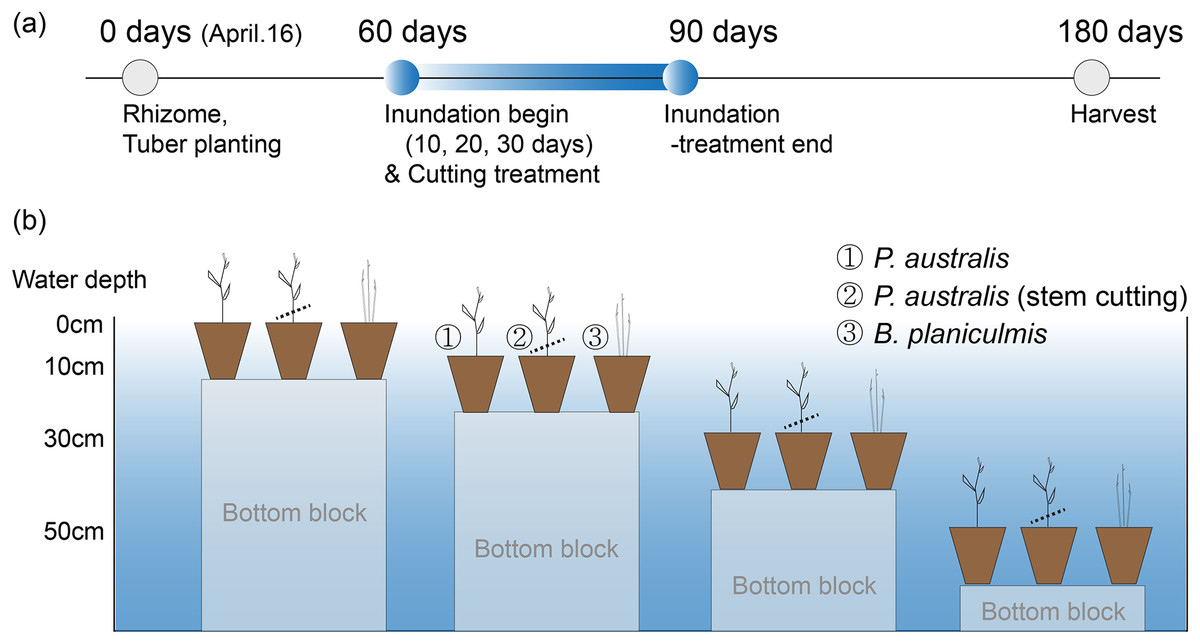
Figure 2: Experimental design to test the inundation effects on the growth of two emergent species, Pragmites australis and Bolboschoenus planiculmis.
(A) Experimental design, (B) illustrated experimental mesocosm established in the restored wetland.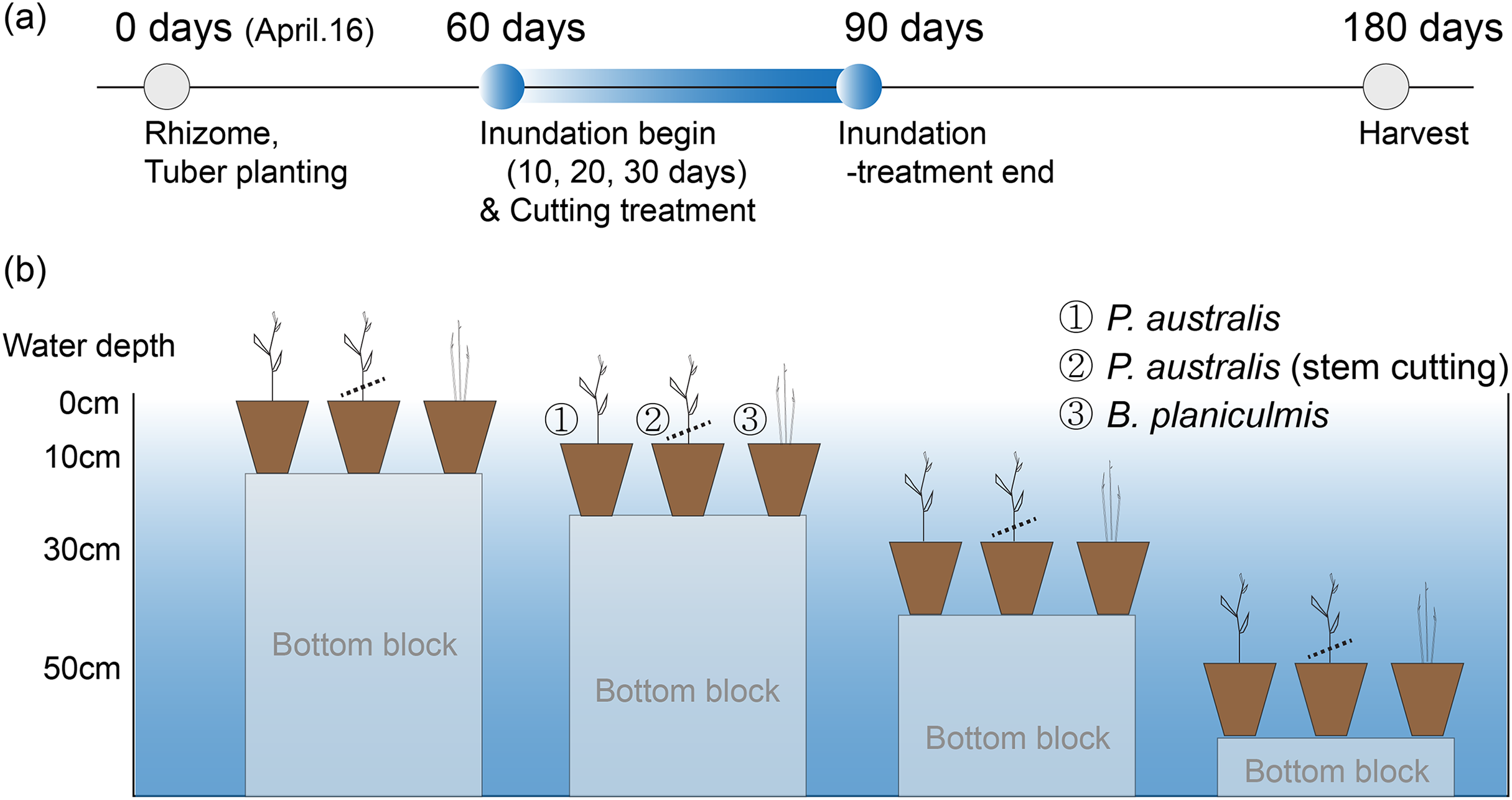{kind=link}
We harvested all plants at the end of the 90 days growth period after the flooding manipulation experiment. We measured stem length (cm), stem diameter (cm), stem density in each pot (stems per pot), and dry weight of the entire plant for each pot. Stem volume was calculated assuming the stem was a volumetric cone by using the following equation.
(1)
The number of pots with surviving plants was recorded to calculate the mortality rate for different inundation depths and periods. At the end of inundation treatment after 90 days, all experimental pots were checked for the survival of plants to the end of the experiment. Plant mortality was recorded according to the following criteria; (1) no new shoot or leaf growth after the inundation treatment, (2) decomposition of stem or rhizome with losing its original shape and color. Mortality rate was calculated as the relative percentage of pots with dead plants to the number of replicate pots under the same experimental condition. All measured plant material was moved to the laboratory and cleaned with tap water to remove any attached sediment or algae. The dry weight of all plant material was measured after drying in a 70 °C convection oven for 3 days. In addition, the number of B. planiculmis tubers was counted, and their dry weight was also recorded.
Data analysis
Differences in measured response variables between P. australis and B. planiculmis were tested using an aligned-rank transform (ART) ANOVA method. The ART ANOVA is a non-parametric analysis of variance that allows testing of the combined treatment effects of two factors (Wobbrock et al., 2011). Where the main effect was significant, post-hoc comparisons for the interaction in a two-way model were carried out using ART contrasts (ART-C; Elkin et al., 2021). All statistical analysis was performed in the R environment (version 4.3.0; R Core Team, 2023).
Results
Inundation effect on the mortality of P. australis and B. planiculmis
Mortality of two emergent plants was notably affected by different inundation depths and lengths in pot experiments (Figs. 3A–3C). All plants in the control groups survived under inundation treatments, with a mortality rate of 0%, except those in the stem cutting experiment, which had a mortality rate of 12.5%. Inundation treatments slightly increased the mortality rate of P. australis to a range of 0–37.8%, as shown in Fig. 3A. When combined with mechanical stress (i.e., stem cutting) prior to inundation treatment, these inundation effects on mortality rate were further enhanced (Fig. 3B). Mortality rate of P. australis increased to 50–87.5% with both inundation and stem cutting. Mortality rate of P. australis was higher in groups exposed to inundation for 20–30 days when compared to those exposed to short-term inundation for 10 days. The mortality rate of B. planiculmis increased with deeper inundation depths and longer inundation periods (see Fig. 3C). All pots planted with B. planiculmis survived the 10cm inundation treatment. The mortality rate of B. planiculmis increased when exposed to 30–50 cm inundation for 20–30 days.
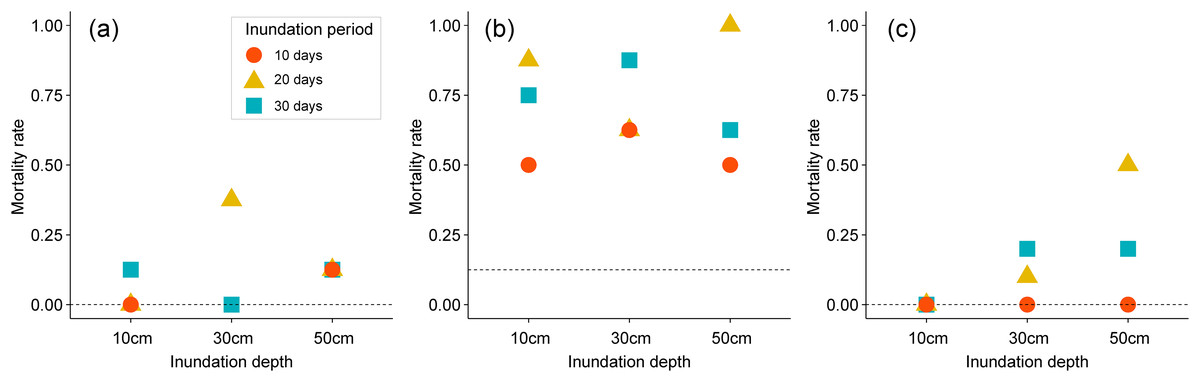
Figure 3: Mortality rate of different inundation conditions. (A) Phragmites australis, (B) P. australis after stem cutting treatment, (C) Bolboschoenus planiculmis.
Horizontal dotted line indicates the mortality rate of the control plot (i.e., no inundation during the experimental period) of each species.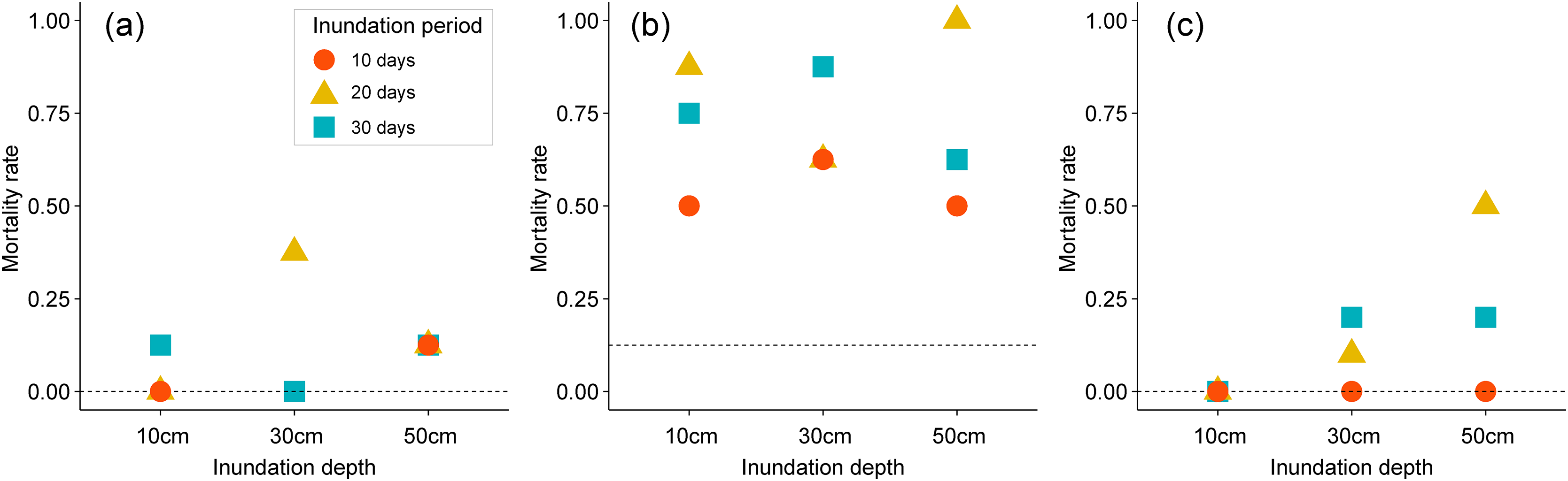{kind=link}
Change of stem volume and biomass
We found that changes in inundation depth significantly decreased the stem volume and total biomass of P. australis (Table 1 and Figs. 4A, 4B). However, the growth performance of P. australis remained unchanged with increasing inundation periods (stem volume: F = 1.13, p = 0.329; total biomass: F = 0.33, p = 0.718). When combined with stem cutting treatment, stem volume of P. australis showed a significant decrease with changing inundation depths (F = 5.99, p = 0.004) and periods (F = 6.03, p = 0.004; Figs. 4C and 4D). After stem cutting, all combinations of different inundation treatments showed a decrease in total biomass compared to the control group. Therefore, no difference was observed between the different inundation treatment groups (p > 0.38).
| Traits | Factors | Species | |||||
|---|---|---|---|---|---|---|---|
| P. australis | P. australis (cut) | B. planiculmis | |||||
| F | p | F | P | F | p | ||
| Stem volume | Depth | 6.74 | 0.002** | 5.99 | 0.004** | 49.95 | <0.001*** |
| Period | 1.13 | 0.329 | 6.03 | 0.004** | 18.01 | <0.001*** | |
| Depth × period | 0.95 | 0.443 | 1.05 | 0.387 | 22.62 | <0.001*** | |
| Total biomass | Depth | 8.38 | <0.001*** | 0.98 | 0.380 | 66.27 | <0.001*** |
| Period | 0.33 | 0.718 | 0.45 | 0.640 | 0.87 | 0.423 | |
| Depth × period | 1.01 | 0.409 | 2.14 | 0.086 | 28.04 | <0.001*** | |
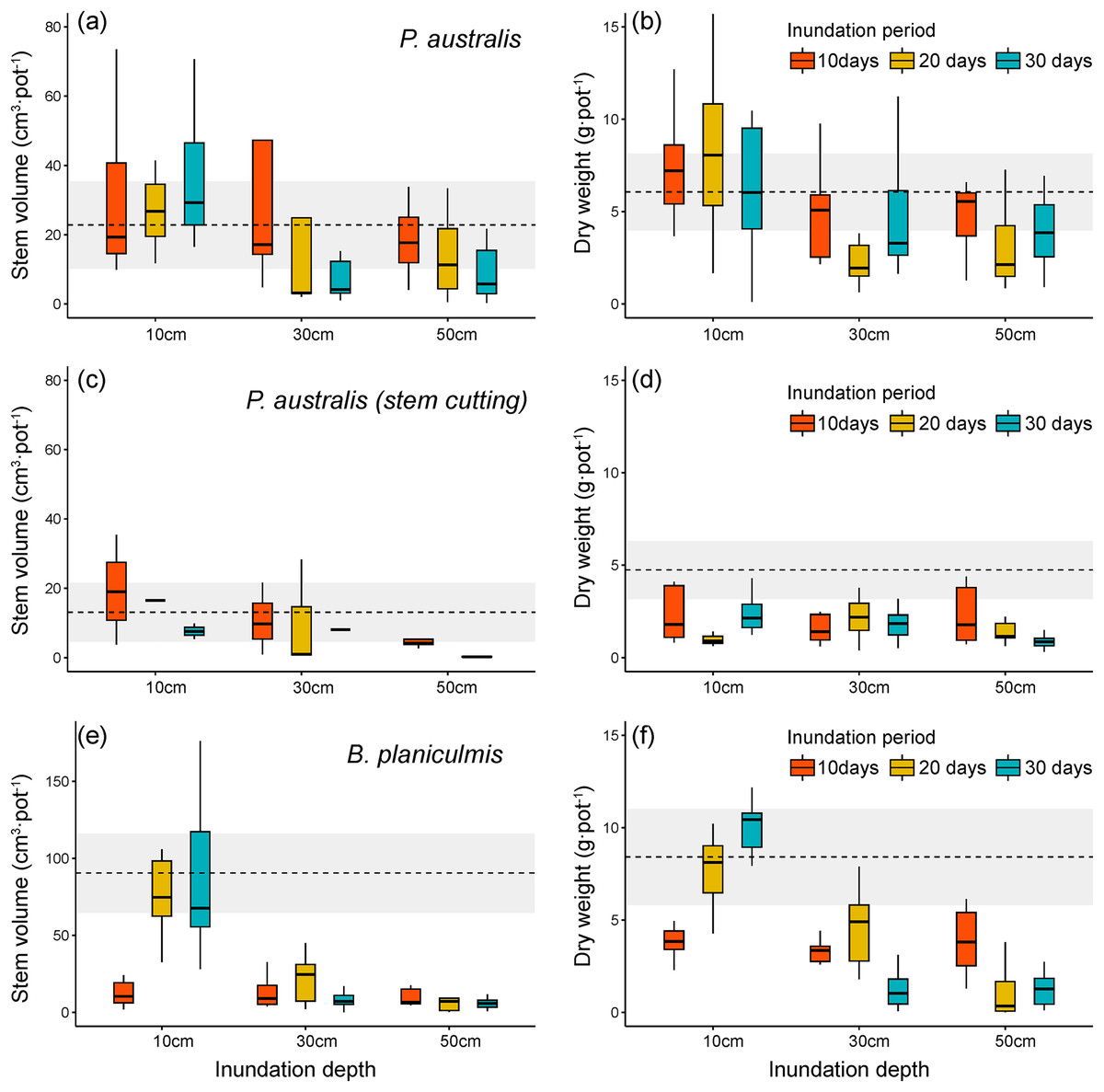
Figure 4: Stem volume and total dry weight after different inundation treatments. (A, B) Phragmites australis, (C, D) P. australis after stem cutting treatment, (E, F) Bolboschoenus planiculmis.
Horizontal dotted line indicates mean of the control plot (i.e., no inundation during the experimental period) of each species. Grey shaded area around dotted line indicates 95% confidence interval. NA, not available.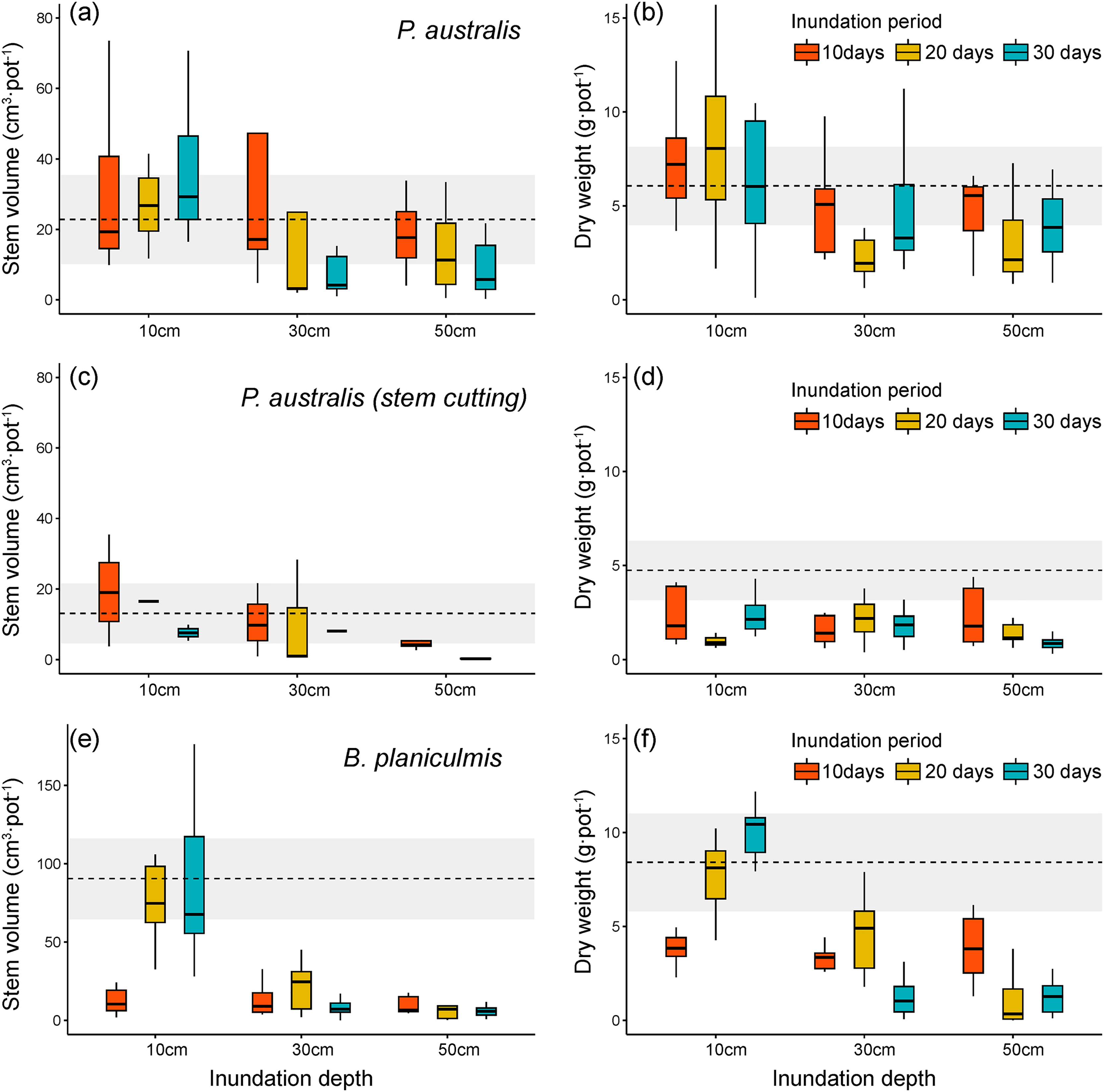{kind=link}
In comparison, B. planiculmis was found to be more sensitive to different inundation conditions than P. australis (Table 1 and Figs. 4E, 4F). Stem volume and total biomass of B. planiculmis decreased with increasing inundation depth and duration. The combined effect of inundation depth and period resulted in significant changes in the growth performance of B. planiculmis across different inundation environments (stem volume: F = 22.62, p < 0.001; total biomass: F = 28.04, p < 0.001). In particular, the combined effect of inundation depth and period showed distinct growth response patterns of B. planiculmis (Figs. 4E, 4F). At inundation depths below 10 cm, stem volume and total biomass of B. planiculmis increased with longer inundation periods. Under deeper inundation conditions, between 30 and 50 cm, longer inundation periods tended to reduce stem volume and biomass of B. planiculmis. This suggests that B. planiculmis thrives in shallow habitat conditions between 10 and 30 cm.
Tuber production of B. planiculmis
We also compared how inundation treatments affected tuber production in B. planiculmis. Similar to the results for stem volume and total biomass, the density and total weight of B. planiculmis tubers exhibited a negative response to shifting inundation conditions (Table 2 and Figs. 5A, 5B). Tuber density was higher in the 10 cm inundation group and the control group than in plants that grew in deeper inundation depths (F = 52.40, p < 0.001). Tuber density increased with inundation periods in the 10 cm inundation treatment. Under deeper inundation conditions, tuber density decreased with increasing inundation periods (F = 20.50, p < 0.001). Total tuber weight exhibited a similar response pattern to the changes in tuber density under different inundation conditions.
| Traits | Factors | F | p |
|---|---|---|---|
| Tuber number | Depth | 52.40 | <0.001*** |
| Period | 3.88 | 0.025* | |
| Depth × period | 20.50 | <0.001*** | |
| Tuber weight | Depth | 49.23 | <0.001*** |
| Period | 0.97 | 0.383 | |
| Depth × period | 21.64 | <0.001*** |
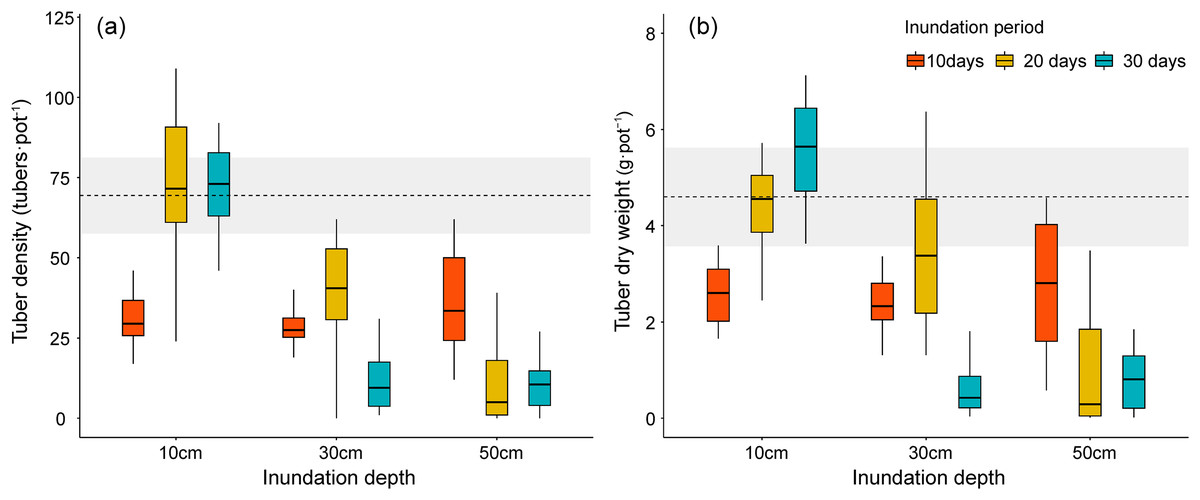
Figure 5: Tuber production of Bolboschoenus planiculmis under different inundation conditions. (A) tuber density, (B) tuber dry weight.
Horizontal dotted line indicates mean of the control plot (i.e., no inundation during the experimental period). Grey shaded area around dotted line indicates 95% confidence interval.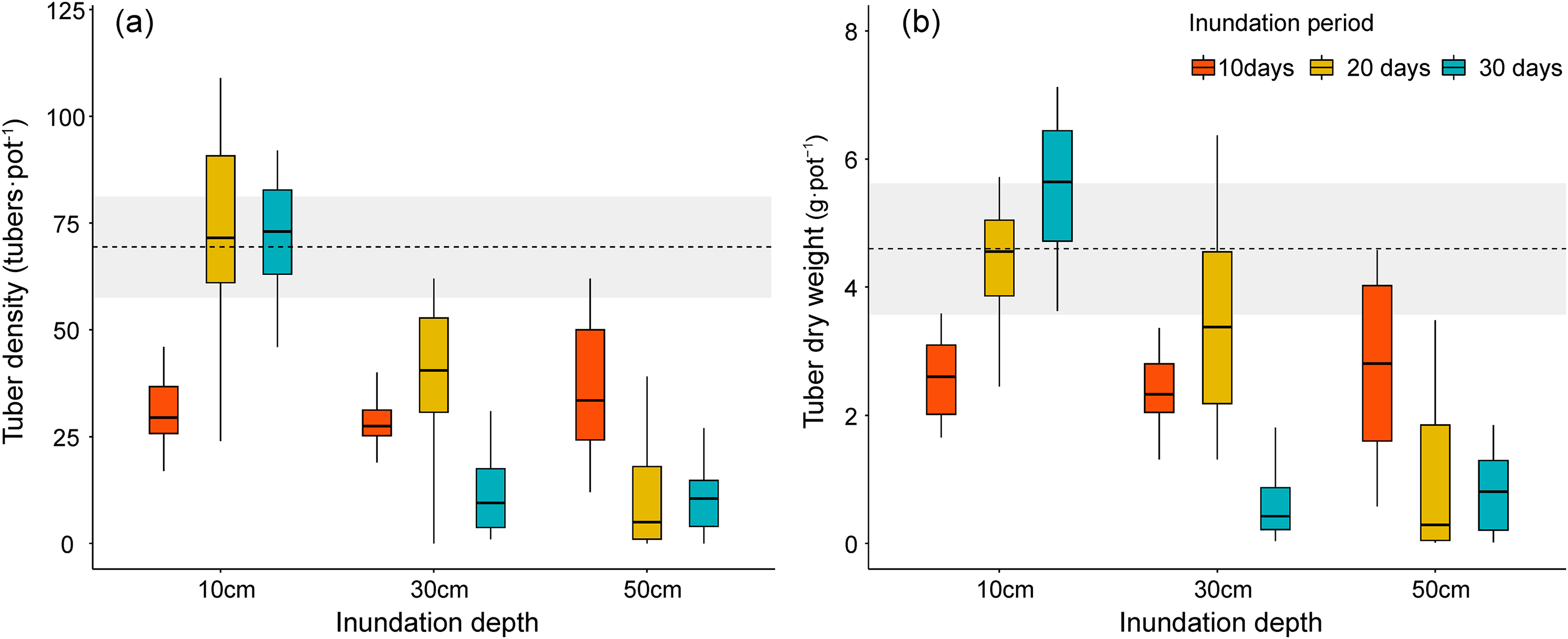{kind=link}
Discussion
Effect of temporal inundation in restored wetland
We tested the growth performance of P. australis and B. planiculmis for the effect of temporal inundation, aboveground cutting, and the combined effect of both. Previous studies on freshwater P. australis populations have reported that increasing inundation period, depth, or frequency can rapidly reduce the growth rate and biomass of P. australis (Tóth, 2016; Yi et al., 2020). In this study, we observed that independent short-term inundation treatments (i.e., 10–30 days, 10–50 cm depth) did not significantly reduce the growth performance of P. australis compared to the control group (i.e., saturated surface sediment). The inconsistent effects of inundation treatment observed in this study may be due to the local environmental conditions to which the experimental P. austalis has been adapted. Song et al. (2021) identified the distinct local adaptations of P. australis populations from inland and coastal wetlands. Due to periodic environmental variations associated with the hydrological regime (such as mean water level, fluctuation frequency, and tidal range), the growth characteristics of P. australis populations in coastal wetlands may experience reduced physiological stress via water level manipulation.
Likewise, populations of P. australis distributed on intertidal mudflats in the Nakdong R. Estuary have adapted to periodic tidal changes. The range of water level manipulation (i.e., 0–50 cm) used in our inundation experiments may not be a sufficient to induce physiological changes in P. australis growth. This finding indicates that a greater degree of inundation or water level manipulation is necessary to manage the growth performance of P. australis in coastal areas, where local plant populations are continuously exposed to inundation, decreased soil iron oxides (Ding et al., 2021) and water level fluctuations. This can serve as an applicable management measure to control the spread of P. australis, if the objective is only to control the expansion of P. australis. However, there may be potential differences in plant growth responses due to environmental settings between restored wetland sites and controlled mesocosms, so further field experiments are needed for in-depth understanding.
As described in our experimental site, the potential range of inundation treatments in restored wetlands may be constrained when the management objective deals with multiple species or management targets. We found that different growth responses to manipulated inundation stress were observed between P. australis and B. planiculmis. In our experiment, under 10–50 cm of inundation stress, the growth inhibition of B. planiculmis was significantly greater than that of P. australis. This suggests that the use of inundation for vegetation control may have negative impact on other endemic wetland plants that have a narrower range of adaptations to water level fluctuations or inundation periods. P. australis has a higher density of aerenchyma in its stems, rhizomes, and roots compared to B. planiculmis (Mal & Narine, 2004; Ding et al., 2021; Tshapa, Naidoo & Naidoo, 2021). The enhanced gas pathway of P. australis ensures oxygen supply to the belowground system during flood conditions. These morphological characteristics explain the different growth responses of the two emergent plants in our experiments. Our experiment result provides empirical evidence that deeper inundation cannot efficiently manage to maintain higher biomass of B. planiculmis, an important food plant resource at the study site.
In our experiment, we observed that biomass and tuber production of B. planiculmis tended to increase under shallow inundation of 0–20 cm and short-term inundation of 10–20 days. Similar experiments manipulating water levels have also shown that tuber production of B. planiculmis was higher under shallow inundation environments (less than 5–10 cm), and decreased under deeper water conditions (Liu et al., 2016; An, Gao & Tong, 2018; Ding et al., 2021). In controlled plot experiments, An et al. (2022) reported that the optimum growth range for B. planiculmis in freshwater environments is between 11–36 cm depth. They also suggested that shallow inundation (0–10 cm) during the early growing season and deeper inundation (10–25 cm) during later growing seasons can assist in B. planiculmis growth and enable it to outcompete other plant species (An et al., 2022). This could be attributed to the local adaptation of B. planiculmis communities, which are often distributed in environments with fluctuating water levels (Hroudová, Zákravský & Flegrová, 2014).
Aboveground-cutting effect on the growth performance of P. australis
Previous restoration experiments reported that cutting and leaving the above-ground part of P. australis resulted in rapid expansion of the remaining belowground plant portion within a year, while aboveground cutting had no significant effect (Greet & King, 2019). A study using various mowing regimes, such as stem cutting, on P. australis plot experiments in Central Europe found that mowing had only a short-term effect on reducing shoot size, even with various cutting trials (Güsewell, Le Nédic & Buttler, 2000). There seems to be little practical evidence supporting the long-term effect of a single mowing event. Other studies have emphasized the importance of nutrient availability in controlling excessive colonization of P. australis in wetland ecosystems (Uddin & Robinson, 2018).
In our short-term experiment, we found that combination of aboveground-cutting and inundation treatments had a significant inhibitory effect on the growth performance and survival rate of P. australis. The growth traits such as stem volume and biomass of P. australis, and the associated survival rate, were both decreased. It is noteworthy that, despite increased level of inundation after cutting, there was no increase in growth inhibition effect. A similar application in the Laurentian Great Lakes watershed showed that a cut-to-drown management strategy for P. australis greatly reduced belowground biomass production and rhizome non-structural carbohydrate content (Widin, Bickford & Kowalski, 2023). Aboveground cutting of P. australis not only affects the total biomass of P. australis patches, but also has a significant management effect in reducing the adaptive capacity of P. australis in wetland ecosystems. For example, the remaining dry shoots of P. australis (usually produced in the previous year) play an important role in aerating the belowground system through the Venturi effect (Björn et al., 2022). In addition, convective flow of gases (e.g., oxygen, carbon dioxide) is initiated in leaf sheaths of the living P. australis stem, then the gases are transmitted through culm, rhizome, and vented to old stems (Armstrong & Armstrong, 1991). This interconnected air-flow network of P. australis provide a resistant and escape mechanism against partial inundation of the entire stem-rhizome network consisting of new and old stems. In this respect, a cut-to-drown strategy or combination of cutting and inundation treatment can inhibit the supply of air through the stem-rhizome network into the drown plant part of P. australis, suppress the diffusion of oxygen, and decrease the essential metabolic processes in the stem-root network.
Conclusions
Drawing upon two core findings from our manipulated experiment, we recommend a combined treatment approach of moderate inundation (e.g., 5–10 cm, for 20–30 days) and aboveground cutting to manage the excessive expansion of P. australis in restored brackish wetlands. Proposed measures can reduce unintended harm to other native wetland plant species, including B. planiculmis, which is a food source for migratory waterfowl, by controlling the growth performance of P. australis using moderate inundation management techniques. A low range of water level adjustment offers management benefits that minimize the compositional change of the hydrological regime, which can also affect wetland species zonation and their community composition. In cases where the P. australis patch is substantially large, the area that can be reasonably managed may be restricted due to the high resource requirement for physically removing P. australis. Since each restoration wetland is unique and requires tailored management approaches, further advancements in the management of large-scale P. australis beds are necessary. And wetland restoration design should be firmly based on the ecological processes to minimize management cost and maintain the desired ecological state.
Supplemental Information
Raw data from the mesocosm experiment of two wetland plant species.
Please refer to the methods section for the specific unit of each variable.
Raw data from the mesocosm experiment on belowground tuber characteristics of Bolboschoenus planiculmis.
Please refer to the methods section for the specific unit of each variable.