
Changes of Tamarix austromongolica forests with embankment dams along the Laizhou bay
- Published
- Accepted
- Received
- Academic Editor
- Mykola Karabiniuk
- Subject Areas
- Biodiversity, Ecology, Plant Science, Environmental Impacts, Forestry
- Keywords
- Disaster relief, Embankment dam, Storm surge, Coastal conservation, Natural vegetation, Shrub
- Copyright
- © 2024 Yang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Changes of Tamarix austromongolica forests with embankment dams along the Laizhou bay. PeerJ 12:e17934 https://doi.org/10.7717/peerj.17934
Abstract
Background
Embankment dams were built south of the Laizhou bay in China for controlling storm surge disasters, but they are not enough to replace coastal forests in protecting the land. This study was designed to evaluate the effects of embankment dams on natural forests dominated by Tamarix austromongolica and test whether the dam-shrub system is a preferable updated defense.
Methods
Coastal forests on two typical flats, one before and one behind embankment dams, were investigated using quadrats and lines. Land bareness, vegetation composition and species co-occurrence were assessed; structures of T. austromongolica populations were evaluated; and spatial patterns of the populations were analyzed using Ripley’s K and K1,2 functions.
Results
In the area before embankment dams, 84.8% of T. austromongolica were juveniles (basal diameter ≤ 3 cm), and 15.2% were adults (basal diameter > 3 cm); behind the dams, 52.9% were juveniles, and 47.1 were adults. In the area before the dams, the land bareness was 13.7%, four species occurred, and they all were ready to co-occur with T. austromongolica; behind the dams, the land bareness was 0%, and 16 species occurred whereas they somewhat resisted co-occurrence with T. austromongolica. In the area before the dams, the T. austromongolica population was aggregated in heterogeneous patches, and the juveniles tended to co-occur with the adults; behind the dams, they were over-dispersed as nearly uniform distributions, while the juveniles could recruit and were primarily independent of the adults. These results indicate that the T. austromongolica species did not suffer from the unnatural dams, but benefited somehow in population expansion and development. Overall, the T. austromongolica species can adapt to artificial embankment dams to create a synthetic defense against storm surges.
Introduction
Coastal areas worldwide have far denser average populations and industries than inland areas, and are normally closely tied to inlands through commercial activities (Kyzar et al., 2021). Unfortunately, storms and hurricanes always impact coasts more seriously than inland areas, by surging seawater and damaging airflow (Wang et al., 2021; Zhang et al., 2023). In China, coastal areas near the Laizhou bay, which is south of the Bohai Sea, suffer the worst from storms (Liu et al., 2020). Every winter and spring, prevailing north-west winds repeatedly push seawater into the southern part of the semi-enclosed Laizhou bay, and the water floods the plain coastal area. About every five years, this area experiences a serious disaster (Yu et al., 1987). In 2007, a strong storm surge incurred a direct economic loss of 2.1 billion Chinese Yuan in this area, a total that did not include the loss in ecosystem services such as soil and biodiversity, primary production, and carbon sequestration (Mehvar et al., 2019; Lin & Egerer, 2020; Yu, 2021). Given the trends of global warming and raising sea-levels, storm surges worldwide may become more frequent and severe (Hinkel et al., 2021; Fang et al., 2022; Tay et al., 2022). In that case, studies must be conducted in advance to take precautions against the disasters, and the Laizhou Bay is a common location for these types of studies.
Coastal forests are effective barriers against storms and seawater surges, preventing forelands from seawater erosion and protecting the backlands from storm blowing (MacFarlane, Kinzer & Banks, 2015; Torita, Igarashi & Tanaka, 2022; van Hespen et al., 2023). Natural forests have self-restoration ability, so do not require expensive and labor-intensive human interventions for maintenance. These forests also offer many benefits, such as atmospheric carbon sequestration, useful bio-products, beautiful scenery, and clean air (Urbis, Povilanskas & Newton, 2019; Bell-James, Boardman & Foster, 2020; Zhang et al., 2021). The dominant native plants around the Laizhou bay are Tamarix austromongolica, Phragmites australis and Suaeda salsa. Phragmites australis is a grass species that normally grows in wetlands or marshes, and S. salsa is an annual grass species that occurs only in tidal or moist flats. The non-native Spartina alterniflora was once planted in a few estuaries, but now is viewed a challenge to ecological health (Wei et al., 2023). Tamarix austromongolica is a shrub species that is widely distributed in the Laizhou bay area, and is present in both tidal flats and broad terrestrial backlands tens of kilometers from the seashore. Tamarix austromongolica shrubs thus play a prominent role in making coastal forests and protecting the lands. The T. austromongolica species originated from the riparian lands in the Ganshu Province, China and is dispersed along the Yellow River into the estuarine delta, subsequently flourishing around the Laizhou Bay (Sun et al., 2016; Liang et al., 2019; Wen et al., 2020). This species is similar to Tamarix chinensis, a sister species in the same genus, but can be identified clearly in blossom through soft drooping inflorescences (Liang et al., 2019; Wen et al., 2020; Fang et al., 2021).
Embankment dams are currently the main physical framework used to stop storm surges (Gittman et al., 2015; Winterwerp et al., 2020). However, as unnatural objects, there are concerns that they may disrupt the functions of natural ecosystems to maintain themselves and serve the societies (Ezcurra et al., 2019; Miao et al., 2022). In the past few decades, stone embankment dams have been built along the seashore of the Laizhou bay, changing the coastal landscape that was initially made of natural vegetation and civilian facilities for salt-extraction, fishery, and tourism (Fig. 1A). The embankment dams were built in former supratidal flats and shallow waters. They are mainly the structures of stones and cement, with the height of about 3∼6 m to prevent usual surges. The dams were expensive to build, but greatly reduced the destructive power of surging water, and some of them have been incidentally used as roads or community squares by locals. These embankment dams cannot completely replace coastal forests as a perfect defense against storms because storms can still reach beyond the dams into the backlands (Rutledge et al., 2021). Even if they could, coastal forests should not be ignored because of pleasant services for humans. This study addresses the following questions: Do embankment dams seriously threaten recruitment and persistence of T. austromongolica populations? How do T. austromongolica forests change with the dams? Is the updated embankment-dam-shrub system a preferable defense for shielding the disastrous coasts?

Figure 1: Study vegetation on the coast beside the Laizhou bay.
(A) Typical coastal landscape with embankment dams and shallow sea; (B) supratidal flat and pioneer plants before embankment dams; (C) typical vegetation with T. austromongolica behind embankment dams; (D) fruits of T. austromongolica; (E) measurement of T. austromongolica fruits, where the length between numbers 1 and 2 is 1 cm. These photos were taken by Hongxiao Yang on south coast of the Laizhou bay with the DJI Mini 3 drone (http://www.dji.com) and the Xiaomi 12X camera (http://www.mi.com).{kind=link}
Materials and Methods
Study area and materials
Weifang City, China lies on the southern coast of the Laizhou bay. North of this city, the terrains are alluvial plain, alluvial marine plain, and marine depositional plain, with a mean slope of approximately 1/3000. The average annual temperature is 12 °C−14 °C, and the annual precipitation is approximately 560 mm. More than 15 days per year experience strong winds (>17.2 m/s) in this area (Liu et al., 2010). Storm surges that cause a sudden rise in sea level >1 m occur about 3 times annually, while those causing a sudden rise in sea level >3 m occur about once every ten years (Zhang et al., 2002). When an extreme storm surge happens to co-occur with an astronomic tide, the sea may surge into the inner land >10 km away. In areas close the sea (<20 km away), the salinity of the soil is often >3‰, making cropping agriculture infeasible. The main industries of this area are fisheries and salt refineries, and with these industries, embankment dams, roads, wharfs and saltpans were built or renewed in the past few decades. Some T. austromongolica shrubs have been planted within or beside these facilities, while native plants grow naturally away from them, forming coastal wild vegetation. In wetlands or salt-marshes, the dominant wild plants are S. salsa and P. australis. In terrestrial highlands, the characteristic vegetation is mainly T. austromongolica-dominated forests with understory grasses or herbs such as Setaria viridis, Artemisia scoparia and Artemisia hedinii. Adult T. austromongolica shrubs are often taller (3–5 m) than other species (<1 or 2 m), so they play a critical role in weakening winds and protecting ground surface (Figs. 1B, 1C). The fruits of T. austromongolica are ready for anemochory, with a 1000-grain weight of 0.045 g, a length of about 0.5 mm, and many thin pappi >2 mm long (Figs. 1D, 1E).
Field survey
Two typical sites with natural vegetation were sampled from the coast in different locations. The first site (N37.2004°, E119.2338°) is a supratidal flat with a width >200 m (vertical to the sea-land boundary) and a length >2 km (horizontal to the above boundary) and situated just before an embankment dam built more than 20 years ago (Fig. 1B). When a storm surge occurs, this site can be inundated for a short time in seawater waves. The coverage of T. austromongolica at this site was about 5–30%, and was relatively dense near the embankment dams. The second site (N37.1205°, E119.2103°) is a piece of typical land behind an embankment dam that was built more than 20 years ago. This site used to be a typical supratidal flat similar to the first site, as elder locals remembered. This land is now waterlogged only during or after summer rainstorms, because the embankment dams are effective at stopping storm surges. The coverage of T. austromongolica at this site was approximately 30–50%.
Survey one: In June 2022, 300 small (1 m ×1 m) quadrats were taken at each site for surveying the growing vegetation, and all living plants in the quadrats were identified and recorded. The quadrats were in 10 haphazard sampling lines and in every line, 30 quadrats were sampled in the distance of at least 5 m. If no plants were found, the quadrat was recorded as bare or blank.
Survey two: In August 2022, 20 large (10 m ×10 m) quadrats were used at each site for surveying T. austromongolica populations in blossom. The T. austromongolica individuals (both juveniles and adults) are mostly <5 m high, and an adult may comprise 3-10 or more gathered branches that were connected on the ground to a common taproot. Basal (ground) diameter of the thickest branch of every T. austromongolica shrub was measured and recorded, by digging until a common taproot was found.
Survey three: The T. austromongolica individuals at each site were measured with a random 200 m sampling-line, where all branches shooting from the same taproot were viewed as an individual. A measuring rope in the precision scale of 0.1 m was used in this survey; all T. austromongolica individuals appearing on, below, or near (<1 m away) the rope were included and their basal diameters of the main stems and relative positions, namely, projections of the main stems of the individuals to the 200 m line, were recorded.
Data analyses
Most of the basal diameters of the T. austromongolica plants were thinner than 8 cm. The plants were classified into juveniles and adults by basal diameters: plants with basal diameters <= 3 cm were considered juveniles, while those with basal diameters >3 cm were classified as adults. The average basal diameter of all the T. austromongolica individuals sampled from the large quadrats at the same site was then calculated as well as the ratios of juveniles and adults to the total number of T. austromongolica plants. The basal diameters were compared between the two sites using the T test in the R4.2.1 software (R Core Team, 2022).
The deviation coefficient of the T. austromongolica population at each site was calculated, being the ratio of variance to mean (V/ ), that is, the degree of T. austromongolica population deviating from the theoretical Poisson random distribution (Zhang, 2011). The V/ value can be checked with the T test method. If V/ >1, the T. austromongolica population followed an aggregation distribution, indicating it was under-dispersed; if V/ = 1, the T. austromongolica population followed the Poisson random distribution; if V/ <1, the population conformed to uniform distribution, indicating it was over-dispersed. The deviation coefficient can be calculated using Eqs. (1)–(4):
(1) (2) (3) (4)
where X is the number of T. austromongolica plants in a certain quadrat, N is the number of quadrats, and t follows the T-distribution with N-1 degrees of freedom.
One-dimensional point pattern analyses were performed with the PASSaGE2 software, using the relative positions of the T. austromongolica plants on the sampling lines (Rosenberg & Anderson, 2011; Miao et al., 2014; Muvengwi et al., 2018).
Population clumping: The juvenile and adult plants are treated as identical entities in these analyses. Ripley’s K functions on a series of distance scales were calculated to show the average number of T. austromongolica plants occurring within specific distances on both sides of a T. austromongolica plant. Thus, it can be known that how spatial distances affected the occurrence of T. austromongolica plants. In this software, the maximum distance that can be analyzed is 1/4 of the length of the lines, i.e., 50 m. Tamarix austromongolica individuals were hypothesized to follow a Poisson random distribution and were randomly permuted for 499 times for obtaining 95% confidence intervals of the K functions (between the upper 97.5% and lower 2.5% confidence values) (Dale, 1999; Vaz, Honrado & Lomba, 2019; Hu et al., 2022). If actual K is higher than the upper 97.5% line, the T. austromongolica plants were aggregated, meaning that the individual number exceeded the hypothesized Poisson distribution. If the actual K was below the lower 2.5% line, the individuals were over-dispersed or scattered in uniform distribution, meaning that the T. austromongolica plants were significantly fewer than the hypothesized Poisson distribution. If the K value was between the upper 97.5% and lower 2.5% lines, the plants followed the Poisson distribution. The K can be calculated using the following Equation (5): (5)
where A is the length of the sampling line, n is the number of T. austromongolica plants, and wij is the weight for edge effect. If the distance between points i and j is less than or equal to d, kij is 1; otherwise, kij is 0.
Juvenile-adult association: The juvenile and adult plants are treated as different entities, and Ripley’s K1,2 functions in different distance scales were calculated to show average numbers of entities of another class appearing beside the juveniles or adults. Thus, their co-occurrence (or spatial association) within a certain distance can be demonstrated. The juveniles and the adults were hypothesized to follow two independent Poisson random distributions, and 499 random permutations were performed to obtain the upper 97.5% and lower 2.5% envelope lines that define the 95% confidence interval of the K1,2 function (Dale, 1999). If actual K1,2 value is higher than the upper 97.5% line, the juveniles were positively associated with the adults, meaning that they co-occur with the adults. If actual K1,2 value was below the lower 2.5% line, the juveniles were negatively associated with the adults, indicating that they were over-dispersed from the adults. If actual K1,2 value was between the upper 97.5% and lower 2.5% lines, the juveniles were independent of the adults. The K1,2 values can be calculated using the following Equation (6): (6)
where A is the length of the sampling line, n1 is the number of juveniles, n2 is the number of adults, and wij is a weight for edge effect. If the distance between points i and j is less than or equal to d, kij is 1; otherwise, kij is 0.
Finally, the rate of bare small quadrats at each site was calculated, being the ratio of the bare small quadrats to all small quadrats. The species richness, or number of species, was measured in each quadrat, and the Pearson correlation coefficient between species richness and T. austromongolica occurrence (1 = present, 0 = absent) was calculated. The Jaccard co-occurrence rate (I) of T. austromongolica with another species at each site was calculated using the following Equation (7) (Jaccard, 1901; Zhang, 2011). (7)
where a is the number of quadrats that included both species 1 (T. austromongolica) and species 2 (another one), b is the number of quadrats with only species 1, and c is the number of quadrats with only species 2.
Results
Basal diameters of T. austromongolica stems at the two sites
On the supratidal flat, 197 T. austromongolica plants were found in the 20 large quadrats, with basal diameters ranging from 0.3 cm to 5.5 cm, averaging 2.22 ± 0.97 cm. Of the 197 plants, 167 (84.8%) were juveniles and 30 (15.2%) were adults. At the site behind the embankment dams, 51 T. austromongolica plants were recorded in the 20 comparative quadrats, with basal diameters ranging from 1.5 cm to 8.5 cm, averaging 3.41 ± 1.24 cm. Of the 51 plants, 27 (52.9%) were juveniles and 24 (47.1%) were adults. There was a significant difference in the diameters of the T. austromongolica plants between the two sites (T-test, t = 6.39, d.f. =66.2, p < 0.001). The ratio of juveniles on the tidal flat was significantly higher than behind the embankment dams, indicating that T. austromongolica could easily reach and establish in supratidal flats, but experienced self-thinning behind the embankment dams with a smaller number of juveniles.
Spatial aggregation of T. austromongolica at the two sites
On the supratidal flat, the density of T. austromongolica plants was 9.85 ± 7.71 individuals/100 m2, and the deviation coefficient V/ was 6.04, which is significantly greater than 1 (t19 = 49.90, p < 0.01), indicating that the plants followed the aggregation distribution, instead of the Poisson random distribution. In the area behind the embankment dams, the density was 2.55 ± 1.00 individuals/100 m2, and the deviation coefficient was 0.39, which is significantly less than 1 (t19 =2.68, p < 0.05), indicating they were over-dispersion in uniform distribution, instead of following the Poisson random distribution.
From the 200 m sampling lines, Ripley’s K at the supratidal site was always higher than the 97.5% envelope line in distance scales (d) < 50 m (Fig. 2A), indicating T. austromongolica aggregated or clumped as patches with radii of several meters instead of following the Poisson random distribution. In contrast, Ripley’s K in the area behind the embankment dams was near and at times slightly lower than the 2.5% envelope line, especially on the fine scales <30 m (Fig. 2B), suggesting that the T. austromongolica plants followed a nearly uniform distribution, being almost segregated instead of aggregated.
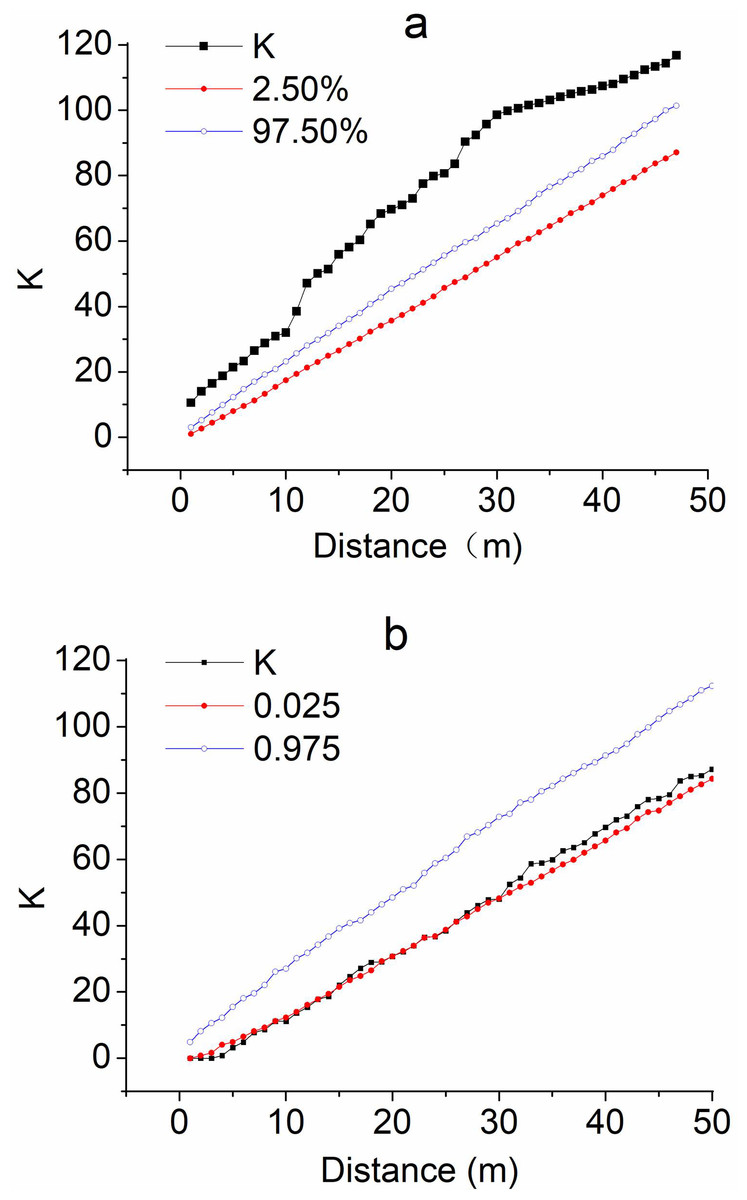
Figure 2: Ripley’s K (spatial aggregation) of T. austromongolica plants at two comparative sites.
(A) Supratidal flat before an embankment dam; (B) sandy land behind embankment dams.{kind=link}
Spatial associations of T. austromongolica juveniles with adults at the two sites
On the supratidal flat, where the distances were <6 m, Ripley’s K1,2 of the T. austromongolica juveniles with adults was higher than the upper 97.5% envelope line, showing a tendency of co-occurrence or aggregation (Fig. 3A); where the distances were >6 and <40 m, K1,2 varied from the upper 97.5% to the lower 2.5% envelope lines, indicating the juveniles were basically independent of the adults in their occurrences; where the distances were >40 m, K1,2 fell below the lower 2.5% envelope line, indicating juveniles were very scarce over 40 m away from T. austromongolica adults.
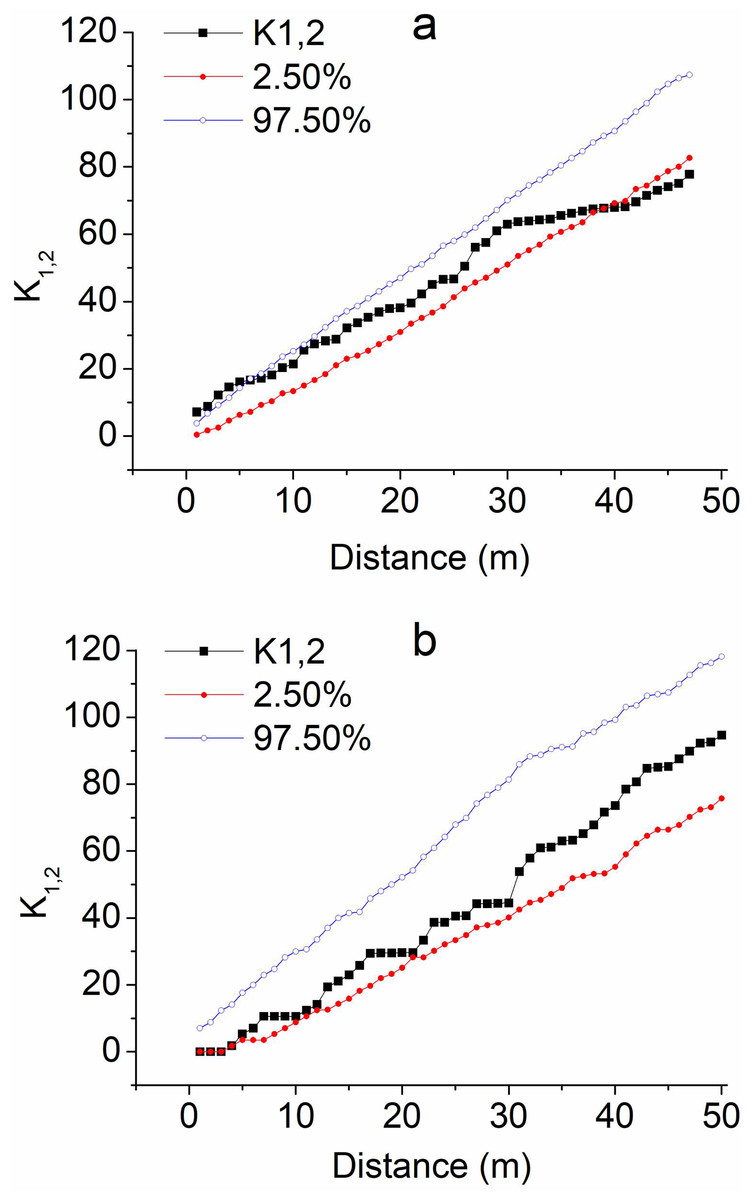
Figure 3: Ripley’s K 1,2 (spatial associations) of T. austromongolica juveniles with adults at two comparative sites.
(A) Supratidal flat just before a dam; (B) sandy land behind dams.{kind=link}
Behind the embankment dams, Ripley’s K1,2 values were significantly different. On fine distance scales <4 m, K1,2 was near the lower 2.5% envelope line, and on other larger scales, it ranged from the upper 97.5% to the lower 2.5% envelope lines (Fig. 3B). These results suggest that the juveniles were independent of the adults on most distance scales >4 m, being neither positively nor negatively associated with the adults. Overall, before the embankment dams, the juveniles co-occurred near the adults, especially at distances less than 6 m; behind the embankment dams, they occurred independently on scales >4 m.
Plant communities at the two sites
In the supratidal flat, the rate of bare small quadrats was 13.7%, and four species were found: T. austromongolica, P. australis, S. salsa and Mulgedium tataricum. Tamarix austromongolica tended to co-occur with other species (Pearson correlation coefficient of T. austromongolica occurrence with species richness: r298 =0.382, p < 0.001). The average Jaccard co-occurrence rate of T. austromongolica with other species was 0.162 ± 0.130.
Behind the embankment dams, the rate of bare small quadrats was 0%, and 16 species were observed, including T. austromongolica, Artemisia scoparia, Conyza Canadensis, Cynanchum chinense, M. tataricum, Setaria viridis, P. australis, Salsola collina, Poa annua, Chenopodium album, Suaeda glauca, Bromus japonicas, Descurainia Sophia, Coreopsis grandiflora, Kochia scoparia, and Lepidium apetalum. However, T. austromongolica somewhat resisted co-occurring with other species (Pearson correlation coefficient of T. austromongolica occurrence with species richness, r298 = −0.103, p = 0.07). The average Jaccard co-occurrence rate with other species was only 0.058 ± 0.084, which was less than that in the supratidal flat.
Discussion
Living of Tamarix austromongolica in coastal environments
Seawater naturally ebbs and flows on the coastline, as usual tides twice a day, astronomical tides several times every month, and occasional surges with extreme storms. In the intertidal zone, terrestrial plants have no chance to establish because of frequent seawater inundation. In the supratidal zone, waves rush in only when storms overlap spring tides. Frequency of the surges may be as low as once every few years or decades, thus allowing a few terrestrial plants to temporarily enter for heterogeneous establishment. Around the Laizhou bay, the main plants are T. austromongolica, P. australis and S. salsa, as observed on the supratidal flat. Of the species, only T. austromongolica is a widespread woody species with strong wind-stopping stems, superior in this aspect to the grassy and habitat-limited P. australis, S. salsa and S. alterniflora (Zhang et al., 2021; Wei et al., 2023).
On the supratidal flat, T. austromongolica clumped in places where seawater receded relatively early and the micro-terrain was high and coarse. On the flats, the soils are salty, water logging is usual for rain and seawater penetration, and erratic winds are common. Tamarix austromongolica, as a species of the Tamarix genus, must be suited to these conditions, since it has successfully colonized the flat (Natale et al., 2010; Wang et al., 2018). The fruits of T. austromongolica are adapted to anemochory (Figs. 1D, 1E). After the fruits mature each year, they fly with available winds, with some falling on bare spots, such as the supratidal flats, where soil salinity may be high, but the fruits can bear it. If the ground surface is adequately moist, seeds wrapped in the fruits germinate within several days and start to grow. In this way, T. austromongolica plants successfully covered more and more supratidal flats and other available bare lands, thus created a natural defense against storms and seawater threats.
State of T. austromongolica in flats before the embankment dams
In flats before the embankment dams, the T. austromongolica population was in development since it was averagely younger than that behind the embankment dams, and the rate of juveniles to adults was 167: 30. The deviation coefficient (V/ ) was greater than 1, and Ripley’s K was higher than the upper 97.5% envelope line on all distance scales. The Ripley’s K1,2 indicated that the juveniles tended to co-occur near the adults in distances <5 m or 6 m. These results suggest that T. austromongolica plants, including juveniles and adults, were clumped as patches in radii of several meters. Of the small quadrats before the embankment dams, 13.7% had no plants, indicating that this site was not crowded with vegetation. Because of this, T. austromongolica plants were conditioned to co-occur with other species, instead of inhibiting each other.
This state is believed to result from the embankment dams. Before the embankment dams were constructed, this site was a shallow sea without any terrestrial plants. After the embankment dams were constructed, marine sediment deposited before the embankment dams and the areas gradually emerged as a flat. The flat was initially free of terrestrial plants because of intolerant daily seawater inundation. After the emergence of the supratidal flat, some land was spared from frequent seawater inundation, and terrestrial plants were able to enter for establishment. With convenient winds, tiny fruits of T. austromongolica easily flew into the flat. Some dropped in water and drifted away with water flows, while some rolled on the ground with winds to stay beside the embankment dams, haphazard jetsam, or existing plants. Those stopping near embankment dams, initial jetsam or existing plants might germinate and grow into new juvenile plants, then joining in a heterogeneous or clumped pattern of T. austromongolica plants, as observed in this study. However, this site was still subject to extreme storm surges and might be inundated once more. In this case, time and chance were far from infinite for them to grow enough as a saturated crowded state. Then, some bare land and spare resources were left, just as the 13.7% bare small quadrats observed in this study.
State of T. austromongolica behind the embankment dams
Behind the embankment dams, the T. austromongolica population has developed into an over-dispersed state. The individuals were older, on average, than those growing before the embankment dams, and the rate of juveniles to adults was 27:24. The deviation coefficient (V/) was less than 1, and Ripley’s K was near or below the lower 2.5% envelope line on all distance scales. The juvenile-adult K1,2 indicated that the occurrences of juvenile plants were basically independent of the adult plants, except at distances <4 m where the juveniles resisted co-occurrence with the adult plants. These results suggest that T. austromongolica individuals behind the embankment dams experienced resource competition and self-thinning. At this site, 0% small quadrats were free of plants, meaning that living plants were much more crowded than in the areas before the embankment dams. In this case, T. austromongolica had to compete for limited soil nutrients and water with other plants, and were precluded from co-occurring with other plants in a fine distance.
The embankment dams are believed to account for this. After the embankment dams were constructed, seawater could no longer inundate the land behind the embankment dams, such that plants were allowed to unremittingly enter for establishment. After many years, the area became crowded with plants, and there were nearly no bare land. Meantime, resource competition among them became quite tense. On one hand, the T. austromongolica population was dominated by the process of self-thinning and had to present an over-dispersed pattern (Lara-Romero et al., 2016; Forrester et al., 2021). On another hand, co-occurrence rate of T. austromongolica with other species was likewise decreased. Nevertheless, T. austromongolica still recruited because of the adequate supply of juveniles. Behind the embankment dams, the occurrences of juveniles were primarily independent of the adults, except at distances <4 m. At this site, fruits of T. austromongolica still flew in with winds and dropped on the ground. Those dropping very near (<4 m) the T. austromongolica adults could scarcely grow due to unendurable competition with growing adults, but those dropping farther from the adult plants had more chances to germinate and grow. For the latter, their establishment was dependent of whether they just dropped beside a dead plant that could no longer compete for living resources but released their ever possessed ones (Thompson et al., 2020; Li et al., 2022). The death and decay of existing plants was largely uncertain; correspondingly, the occurrence of T. austromongolica juveniles was primarily stochastic and independent, rather than closely associated with the T. austromongolica adults (Kyzar et al., 2021; Cannon, Piovesan & Munne-Bosch, 2022; Knapp, Attinger & Huth, 2022).
Changes of T. austromongolica forests with the embankment dams
The embankment dams facilitated the expansion and development of T. austromongolica populations. Before the embankment dams, sediments were stopped and piled up as new land where the T. austromongolica population could easily colonize. With gradual broadening of the new land, the T. austromongolica forests then expanded seaward. Behind the embankment dams, the absence of storm surges facilitated the development of T. austromongolica population as well, resulting in a saturated pattern with over-dispersed T. austromongolica shrubs, where T. austromongolica recruited with necessary juveniles. Once bare spots are available, T. austromongolica fruits enter for possible germination and establishment. In this way, T. austromongolica are able to keep dispersing and persisting along the coast even with the intervention of unnatural dams.
The embankment dams and T. austromongolica forests were united as an effective defense against storm surges. This study indicates that the embankment dams did not set back the recruitment of T. austromongolica, which is dispersed by winds instead of seawater. After T. austromongolica forests were established, they could, in turn, protect to some extent the embankment dams from the damage of storm surges. Thus, the forests collaborated with the embankment dams as a tenacious and self-maintaining defense against storms, and coastal people were exempted from huge cost and labor for maintaining the defense. This study demonstrates that coastline hardening may greatly change primitive ecosystems, but not necessarily cause an unpleasant degradation of the ecosystems, just as the T. austromongolica forests experienced (Gittman et al., 2015; Powell et al., 2019).
Conclusion
This study shows that the shrub species T. austromongolica did not suffer from the building of embankment dams. Fruits of T. austromongolica readily flew with winds and fell in spare flat that emerged with the embankment dams, thus creating a patch-like barrier to resist seawater surges. Behind the embankment dams, T. austromongolica populations developed smoothly to a steady state where resource competition among plants became intensified, and grown-up T. austromongolica shrubs were over-dispersed as nearly uniform distribution. These findings suggest that building embankment dams is preferred and the embankment dams can collaborate with native T. austromongolica forests as ideal self-persistent defense to protect calamitous coasts around the temperate Laizhou bay. This study is informative for coastal areas not only along the Laizhou Bay, but also others in similar troubles as long as they are suitable for the growth of T. austromongolica.
Supplemental Information
Coast forests data
Data is sampled from the south coast of the Laizhou Bay.