
Exploring Barbronia species diversity and phylogenetic relationship within Suborder Erpobdelliformes (Clitellata: Annelida)
- Published
- Accepted
- Received
- Academic Editor
- Armando Sunny
- Subject Areas
- Biodiversity, Genetics, Genomics, Zoology, Freshwater Biology
- Keywords
- Leeches, Species delimitation, bGMYC, bPTP, Phylogenetic tree, Mitochondrial genome, rDNA
- Copyright
- © 2024 Liu et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Exploring Barbronia species diversity and phylogenetic relationship within Suborder Erpobdelliformes (Clitellata: Annelida) PeerJ 12:e17480 https://doi.org/10.7717/peerj.17480
Abstract
Background
Barbronia, a genus of freshwater macrophagous leeches, belongs to Erpobdelliformes (Salifidae: Clitellata: Annelida), and B. weberi, a well-known leech within this genus, has a worldwide distribution. However, the systematics of Barbronia have not yet been adequately investigated, primarily due to a few molecular markers, and only 20 Barbronia sequences available in the GenBank database. This gap significantly limits our understanding of the Barbronia species identification, as well as the phylogenetic placement of the genus Barbronia within Salifidae.
Methods
Next-generation sequencing (NGS) was used to simultaneously capture the entire mitochondrial genome and the full-length 18S/28S rDNA sequences. The species boundary of Barbronia species was estimated using bGMYC and bPTP methods, based on all available Barbronia COI sequences. Uncorrected COI p-distance was calculated in MEGA. A molecular data matrix consisting of four loci (COI, 12S, 18S, and 28S rDNA) for outgroups (three Haemopis leeches) and 49 erpobdellid leeches, representing eight genera within the Suborder Erpobdelliformes was aligned using MAFFT and LocARNA. This matrix was used to reconstruct the phylogenetic relationship of Barbronia via Bayesian inference (BI) and the maximum likelihood (ML) method.
Results
The full lengths of the mitochondrial genome, 18S and 28S rDNAs of B. cf. gwalagwalensis, are 14847 bp, 1876 bp 1876 bp, and 2863 bp, respectively. Both bGMYC and bPTP results based on COI data are generally congruent, suggesting that the previously proposed taxa (B. arcana, B. weberi formosana, and B. wuttkei or Erpobdella wuttkei) are synonyms of B. weberi. The specimens listed in the B. gwalagwalensis group, however, are split into at least two Primary Species Hypotheses (PSHs). The p-distance of the first PSH is less than 1.3% but increased to 4.5% when including the secondary PSH (i.e., B. cf. gwalagwalensis). In comparison, the interspecific p-distance between the B. weberi group and the B. gwalagwalensis group ranged from 6.4% to 8.7%, and the intraspecific p-distance within the B. weberi group is less than 0.8%. Considering the species delimitation results and the sufficient large p-distance, the specimen sampled in China is treated as B. cf. gwalagwalensis. The monophyly of the four Erpobdelliformes families Salifidae, Orobdellidae, Gastrostomobdellidae sensu stricto and Erpobdellidae is well supported in ML and BI analysis based on a data of four markers. Within the Salifidae, a well-supported Barbronia is closely related to a clade containing Odontobdella and Mimobdella, and these three genera are sister to a clade consisted of Salifa and Linta. According to the results of this study, the strategy of simultaneous obtaining both whole mitochondria and nuclear markers from extensively sampled Salifids species using NGS is expected to fathom both the species diversity of B. gwalagwalensis and the evolutionary relationship of Salifidae.
Introduction
Barbronia is a genus of predatory freshwater leeches belonging to the family Salifidae (Erpobdelliformes, Clitellata, Annelida) with only three species have previously been reported in China: Barbronia weberi, Barbronia zhejiangica Yang, 1996, and Barbronia yunnanensis Yang, Wang & Zhang, 1997 (Yang, 1996; Yang, Wang & Zhang, 1997). The former one is a well-known introduced leech species, however, the latter two are rarely known endemic Barbronia species. Barbronia weberi was originally described by Blanchard (1897) in Indonesia, but now it is broadly found in China (Ta-Hsiang, 1974; Zhao et al., 2020), India (Bandyopadhyay & Mandal, 2005; Chandra & Mahajan, 1971; Ghate, 1991), Myanmar (Eriksen et al., 2022), United States (Rutter & Klemm, 2001; Sawyer & Sawyer, 2018), Brazil (Pamplin & Rocha, 2000), Mexico (Garduño-Montes de Oca et al., 2016; Oceguera-Figueroa, León-Règagnon & Siddall, 2005), United Kingdom (Sawyer, 1986; Sawyer & Sawyer, 2018), Germany (Kutschera, 2004; Nehring, 2006), Hungary (Ludanyi et al., 2019), Italy (Genoni & Fazzone, 2008), Spain (Pavluk, Pavluk & Rasines, 2011), Netherlands (Van Haaren et al., 2004), Australia (Govedich et al., 2003; Govedich, Bain & Davies, 2002), New Zealand (Mason, 1976), and South Africa (Nakano & Nguyen, 2015). Barbronia weberi has achieved its current wide distribution not only through human activities (Govedich et al., 2003), but also through its ability to reproduce cocoons asexually, without the need for cross-fertilization (Sawyer, 2020). Moreover, B. weberi is the second intermediate host of the parasitic trematode Australapatemon niewiadomski (Blasco-Costa, Poulin & Presswell, 2016).
The taxonomic status of the widely distributed invasive species B. weberi needs to be evaluated on a global scale (Oceguera-Figueroa et al., 2011), which involved B. weberi formosana, B. wuttkei, B. arcana, and Barbronia sp. Molecular identification has become a valuable complement to morphological taxonomy over the past two decades. A partial mitochondrial gene COI sequence has been used to recognize Barbronia species. Barbronia wuttkei was originally described as Erpobdella wuttkei based solely on morphological traits (Kutschera, 2004), however, it was later transferred to the genus Barbronia but retained as a valid name by Grosser & Trontelj (2008) based on COI phylogenetic evidence. The genetic analysis using the COI sequence-based species delimitation approach, Poisson Tree Process (PTP), has led to the initial assumption that Barbronia wuttkei and B. arcana should be classified under the taxon B. weberi (Klass et al., 2021). One Barbronia sp. (GenBank accession: MN503261), as well as another specimen (GenBank accession: MF458701, marked as Erpobdella sp.), are recently recognized as B. gwalagwalensis on basis of COI data (Klass et al., 2021).
The phylogeny of Barbronia was initially inferred solely based on 18S ribosomal DNA sequences alone, B. weberi was found to be closely related to a clade of two erpobdellids: Dina lineata (synonym: Erpobdella lineata) and Erpobdella octoculata (Trontelj, Sket & Steinbrück, 1999). Although an attempt was made to amplify a fragment of the mitochondrial marker 12S rDNA from the B. weberi specimen by Trontelj, Sket & Steinbrück (1999), it failed using the primer pair 12S-A/12S-B (Siddall, 2002). Consequently, only the unique 18S rDNA sequence of B. weberi was used in subsequent phylogenetic study (Borda & Siddall, 2004a; Oceguera-Figueroa, León-Règagnon & Siddall, 2005). A comprehensive phylogenetic analysis was conducted using combined data from partial sequences of 18S rDNA, 28S rDNA, 12S rDNA, and cytochrome c oxidase subunit I (COI) published by Borda & Siddall (2004b). This study confirmed that the Barbronia species (B. weberi, B. weberi formosana, B. gwalagwalensis, and an undescribed Barbronia species) formed a monophyletic group closely related to Linta be. Another phylogenetic result supported that B. weberi and B. arcana were part of the Salifidae clade, a sister group of Erpobdellidae (Oceguera-Figueroa, León-Règagnon & Siddall, 2005). An expanded dataset including six Barbronia specimens represented all mentioned Barbronia species and B. wuttkei, forming a monophyletic group that was a sister group to a clade of Salifa and Linta species (Oceguera-Figueroa et al., 2011). Recently, Barbronia represented by three taxa (Anderson, Braoudakis & Kvist, 2020), and Barbronia weberi only (Tessler, Siddall & Oceguera-Figueroa, 2018), are closely related to a clade that includes Odontobdella blancharidi and Mimobdella japonica.
The implementation of next-generation sequencing (NGS) technologies has led to a substantial increase in the number of whole mitochondrial genomes in the GenBank database (Kuang & Yu, 2019), and has gradually become a mainstay of species identification and phylogeny research (Franco-Sierra & Diaz-Nieto, 2020; Kortsinoglou et al., 2020). To date, however, only about 20 sequences of various gene fragments from Barbronia specimens are available in the GenBank database. Eight of these sequences are the mitochondrial COI barcodes, six partial 18S rDNA sequences, four short 28S rDNA sequences, and only two 12S rDNA sequences, representatively. Neither a complete mitochondrial genome of Barbronia species nor even the family Salifidae has been published. This gap significantly limits our understanding of either the species identification or phylogenetic relationships of Barbronia within Salifidae.
In the present study, the whole mitochondrial genome, as well as the 18S rDNA and 28S rDNA of Barbronia cf. gwalagwalensis, are assembled and annotated; the species boundary of Barbronia species is evaluated using two kinds of single-locus COI based species delimitation methods; the phylogenetic relationships of Barbronia within the Suborder Erpobdelliformes are conducted based on both mitochondrial and nuclear data.
Materials and Methods
DNA extraction and assembly of the mitochondrial genome, and rDNAs
The specimen later identified as Barbronia cf. gwalagwalensis was sampled using a hydrobiological Peterson dredge during a general benthic sampling expedition in Yunlong Lake (Xuzhou, Jiangsu province, China), and morphological identification was performed with reference to Yang (1996). The total DNA material was extracted using Ezup Column Animal Genomic DNA Purification Kit (Sangon Biotech Company, Shanghai, China) following the manufacturer’s protocols. The quality of DNA was evaluated via gel electrophoresis and visualized on a ChemiDoc XRS + (Bio-Rad, Hercules, CA, USA). The quantification of DNA was checked via the Spectrophotometer NanoDrop 2000. DNA was eluted in ddH2O and stored at −20 °C until used for next-generation sequencing (NGS). A sequencing library was constructed using a Nextera XT DNA Library Preparation Kit (Illumina, San Diego, CA, USA), and sequencing reads were generated on a MiSeq System (Illumina, San Diego, CA, USA) to generate 150 bp paired-end reads. Low-quality bases in raw FastQ reads were trimmed using Trimmomatic v.0.36 with default setting, the remaining reads were assembled as contig and used to reconstruct the mitochondrial genome via GetOrganelle V1.1.7 with recommended setting (Jin et al., 2020). The annotation of mitochondrial protein-coding genes, Transfer RNAs, and ribosomal DNAs of B. cf. gwalagwalensis was performed by selecting the Invertebrate genetic code and referencing the RefSeq63 Metazoa dataset via MITOS2 Webserver. Additionally, manual checks were conducted by comparing nucleotide sequences with those from the published available closely related leech species to refine these annotations when possible using Geneious 2021.
Single-locus based species delimitation
Species delimitation of Barbronia species should be assessed using various molecular species delimitation approaches with data from single-locus or multiple loci. Only 20 Barbronia sequences are available in GenBank, and half of these sequences are partial mitochondrial gene COI sequences (Table 1). In addition, two specimens (accession number: DQ009666 and MF458701) labeled as Erpobdella sp. are also included, because both are used in previous studies of Barbronia (Anderson, Braoudakis & Kvist, 2020; Grosser & Trontelj, 2008; Klass et al., 2021; Oceguera-Figueroa et al., 2011). The species delimitation of Barbronia was conducted solely based on a single-locus dataset of COI, using both a Bayesian implementation of the general mixed yule-coalescent model (bGMYC) and an updated version of the maximum likelihood Poisson Tree Processes model (bPTP) in the current study. The bGMYC model is capable of inferring the transition between the Yule model (species level) and coalescent model (population level within species) (Fujisawa & Barraclough, 2013), with posterior probabilities accounting for phylogenetic uncertainty (Reid & Carstens, 2012). The bPTP analysis required a phylogenetic gene tree, and putative species were estimated with Bayesian support through a simulation of Poisson Tree Processes (Zhang et al., 2013).
| Groups | Taxon | Country | COI accession | COI size (bp) | 12S accession |
18S accession |
28S accession |
|---|---|---|---|---|---|---|---|
|
Barbronia weberi |
B. weberi | Costa Rica | HQ336339 | 649 | – | AF099951 | HQ336356 |
| B. weberi formosana | United States | AY786456 | 600 | – | AY786461 | AY786448 | |
| “B. arcana” | Mexico | DQ235598 | 649 | DQ235588 | DQ235608 | – | |
| B. weberi | Mexico | KU553102 | 645 | – | – | – | |
| B. weberi | Mexico | KU553103 | 645 | – | – | – | |
| “Erpobdella wuttkei” | Germany | DQ009666 | 584 | – | – | – | |
| Barbronia sp | South Africa | AY786457 | 360 | – | AY786463 | AY786450 | |
|
Barbronia gwalagwalensis |
B. cf. gwalagwalensis | China | OQ339201* | 1,534 | OQ339201* | OQ269483* | OQ269482* |
| B. gwalagwalensis | South Africa | AY786455 | 480 | – | AY786462 | AY786449 | |
| Barbronia sp | Myanmar | MN295405 | 660 | – | – | – | |
| Barbronia sp | South Korea | MN503261 | 710 | – | MT010330 | – | |
| “Erpobdella. sp” | France | MF458701 | 634 | – | – | – | |
| Outgroups | Mimobedlla japonica | Japan | AB679658 | 1,267 | – | – | – |
| Odontobdella blancharidi | Japan | AB938004 | 1,267 | – | – | – |
Note:
The groups referred to the clade A, clade B and outgroups of the COI tree in Fig. 3, respectively. Taxon names enclosed in quotations indicate that these taxa may have been misidentified. The accession numbers highlighted with asterisks in bold indicate newly added data., and a dash (“−”) indicates that no sequences are available.
For species delimitation analyses in bGMYC and bPTP, the input COI phylogenetic tree was performed with BEAST Version 1.10.4, using HKY + I + G substitution model with two partitions (one partition comprised the 1st and 2nd codon positions of COI, the other one was the 3rd codon position) with a yule speciation prior and a strict clock. A total of ten million MCMC generations were performed, with samples taken every 1,000 generations. After burn-in, a maximum clade credibility tree was built using TreeAnnotator. The bGMYC analyses were conducted following the approach proposed by Liu et al. (2017). The bGMYC analyses consisted of the total 50,000 MCMC generations, with a thinning interval of every 100 generations, discarding the first 2,000 generations as burn-in, and setting the upper and lower bounds on the threshold parameter from 1 (the minimum number of species) to 11 (the maximum number of tips in COI tree). For bPTP analyses were performed with default setting via an online server (https://species.h-its.org/). In addition, it is worth noting that GMYC models may not provide accurate delineate species properly in data sets composed of only one or two species (Dellicour & Flot, 2015), therefore, Mimobedlla japonica and Odontobdella blancharidi were included in the analyses of species delimitation.
Phylogenetic analysis using multiple loci
To corroborate previous overarching phylogenetic frameworks, 261 sequences of four molecular markers (COI, 12S rDNA, 18S rDNA, and 28S rDNA) representing 49 erpobdellid leeches and three Haemopis leeches were retrieved from the GenBank database (Table 2). The genera Dina and Mooreobdella are synonyms of the genus Erpobdella (Siddall, 2002), consequently, both Dina and Mooreobdella are referred as Erpobdella in the current study. COI sequences were aligned using MAFFT 7.45 with the most accurate LINSI option (Katoh & Standley, 2014), and sequences of rDNA were aligned via LocARNA with default settings (Will et al., 2012). The individual alignments of four markers were concatenated in PhyloSuite (Zhang et al., 2020) as a matrix used for flowing analyses. Phylogeny analyses were performed separately using Bayesian inference (BI) and the maximum likelihood (ML) method via MrBayes V3.2.7 (Altekar et al., 2004) and IQ-TREE (Nguyen et al., 2015). Haemopis caeca, Haemopis kingi, and Haemopis marmorata belonging to Haemopidae (Hirudiniformes) were used as outgroups. The best-fit evolutionary model of each molecular marker (COI, 12S rDNA, 18S rDNA and 28S rDNA) data was determined using PartitionFinder 2 with the ‘greedy’algorithm based on the AICc score (Lanfear et al., 2017). For Bayesian analysis, two independent runs, with four Markov Chain Monte Carlo (MCMC) chains each, were simultaneously carried out for four million generations and sampled every 10,000 generations. The analysis was assumed to have reached stationarity when the potential scale reduction factor value (PSRF) approached 1.0 and the effective sample size value >100. After discarding the 25% samples as burn-in, the 50% majority-rule consensus tree was built. For the ML analysis, the reliability of bootstrap values and tree topology was assessed by ultrafast bootstrap using 1,000 replicates.
| Family | Species | COI | 12S | 18S | 28S |
|---|---|---|---|---|---|
| Salifidae | Barbronia cf. gwalagwalensis | OQ339201* | OQ339201* | OQ269483* | OQ269482* |
| Salifidae | Barbronia gwalagwalensis | AY786455 | – | AY786462 | – |
| Salifidae | Barbronia weberi | HQ336339 | DQ235588 | AF099951 | HQ336356 |
| Salifidae | Odontobdella blanchardi | AB675016 | AB675017 | AB663651 | AB663671 |
| Salifidae | Mimobdella japonica | AB675014 | AB675015 | AB663650 | AB663670 |
| Salifidae | Linta be | AY786460 | – | AY786466 | AY786453 |
| Salifidae | Salifa motokawai | LC029431 | LC029432 | LC029434 | LC274548 |
| Salifidae | Salifa perspicax | HQ336341 | HQ336349 | HQ336375 | HQ336359 |
| Salifidae | Salifa perspicax | HQ336343 | HQ336351 | HQ336377 | HQ336360 |
| Orobdellidae | Orobdella angustata | LC323139 | LC323141 | LC323140 | LC431606 |
| Orobdellidae | Orobdella brachyepididymis | LC106320 | LC106318 | LC106319 | LC274535 |
| Orobdellidae | Orobdella dolichopharynx | AB675028 | AB675029 | AB663665 | AB663666 |
| Orobdellidae | Orobdella esulcata | AB675020 | AB675021 | AB663655 | AB663656 |
| Orobdellidae | Orobdella ghilarovi | LC431609 | LC431616 | LC431608 | LC431607 |
| Orobdellidae | Orobdella ijimai | AB675030 | AB675031 | AB663659 | AB663660 |
| Orobdellidae | Orobdella kanaekoikeae | LC184548 | LC184547 | LC184551 | LC274533 |
| Orobdellidae | Orobdella kawakatsuorum | AB675032 | AB675033 | AB663661 | AB663662 |
| Orobdellidae | Orobdella ketagalan | AB704787 | AB704788 | AB704785 | LC274546 |
| Orobdellidae | Orobdella koikei | AB679688 | AB679689 | AB698883 | LC274543 |
| Orobdellidae | Orobdella masaakikuroiwai | AB938006 | AB937997 | AB938003 | LC274530 |
| Orobdellidae | Orobdella meisai | LC314424 | LC314422 | LC314423 | LC431605 |
| Orobdellidae | Orobdella mononoke | AB698866 | AB698867 | AB698868 | LC274547 |
| Orobdellidae | Orobdella montipumila | LC616663 | LC616667 | LC616674 | LC616673 |
| Orobdellidae | Orobdella nakahamai | LC106331 | LC106329 | LC106330 | LC274534 |
| Orobdellidae | Orobdella naraharaetmagarum | LC087144 | LC087142 | LC087143 | LC274531 |
| Orobdellidae | Orobdella octonaria | AB675024 | AB675025 | AB663667 | AB663668 |
| Orobdellidae | Orobdella octonaria | HQ336338 | HQ336348 | HQ336372 | HQ336355 |
| Orobdellidae | Orobdella okanoi | LC106342 | LC106340 | LC106341 | LC274532 |
| Orobdellidae | Orobdella shimadae | AB675026 | AB675027 | AB663663 | AB663664 |
| Orobdellidae | Orobdella tsushimensis | AB675018 | AB675019 | AB663653 | AB663654 |
| Orobdellidae | Orobdella whitmani | AB675022 | AB675023 | AB663657 | AB663658 |
| Orobdellidae | Orobdella yamaneae | LC106350 | LC106348 | LC106349 | LC274536 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella ampunganensis | LC274551 | LC274564 | LC274517 | LC274516 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella ampunganensis | LC274559 | LC274568 | LC274525 | LC274524 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella extenta | LC274553 | LC274565 | LC274519 | LC274518 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella extenta | LC274555 | LC274566 | LC274521 | LC274520 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella extenta | LC274557 | LC274567 | LC274523 | LC274522 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella monticola | AB675011 | AB675010 | AB663649 | AB663669 |
| Gastrostomobdellidae sensu stricto | Gastrostomobdella monticola | LC274549 | LC274563 | LC274514 | LC274513 |
| Erpobdellidae | Dina lineata | – | AF099952 | AF099950 | – |
| Erpobdellidae | Erpobdella adani | MG745144 | MG745141 | MG745138 | – |
| Erpobdellidae | Erpobdella adani | MG745145 | MG745142 | MG745139 | – |
| Erpobdellidae | Erpobdella adani | MG745146 | MG745143 | MG745140 | – |
| Erpobdellidae | Erpobdella annulata | HQ336345 | – | HQ336379 | HQ336362 |
| Erpobdellidae | Erpobdella bucera | MN612829 | MN613043 | MN613063 | MN613084 |
| Erpobdellidae | Erpobdella costata | AY425460 | AY425442 | AY425478 | AY425406 |
| Erpobdellidae | Erpobdella dubia | AF116023 | AF462022 | AF115997 | AY425365 |
| Erpobdellidae | Erpobdella japonica | AB675012 | AB675013 | AB663648 | AB663652 |
| Erpobdellidae | Erpobdella japonica | AF116026 | AF462023 | AF116000 | AY425366 |
| Erpobdellidae | Erpobdella mexicana | DQ235595 | DQ235585 | DQ235605 | HQ336364 |
| Erpobdellidae | Erpobdella mexicana | DQ235597 | DQ235587 | DQ235607 | HQ336365 |
| Erpobdellidae | Erpobdella microstoma | MN612934 | MN613044 | MN613065 | MN613086 |
| Erpobdellidae | Erpobdella montezuma | GQ368760 | GQ368820 | GQ368802 | – |
| Erpobdellidae | Erpobdella obscura | MN613005 | MN613045 | MN613066 | MN613087 |
| Erpobdellidae | Erpobdella obscura | MN612911 | MN613046 | MN613067 | MN613088 |
| Erpobdellidae | Erpobdella ochoterenai | DQ235596 | DQ235586 | DQ235606 | HQ336370 |
| Erpobdellidae | Erpobdella ochoterenai | DQ235599 | DQ235590 | DQ235609 | HQ336371 |
| Erpobdellidae | Erpobdella octoculata | HQ336344 | – | HQ336378 | HQ336361 |
| Erpobdellidae | Erpobdella parva | MN612997 | MN613052 | MN613073 | MN613094 |
| Erpobdellidae | Erpobdella parva | MN612930 | MN613053 | MN613074 | MN613095 |
| Erpobdellidae | Erpobdella punctata | HQ336346 | HQ336352 | HQ336380 | HQ336363 |
| Erpobdellidae | Erpobdella punctata | MN612994 | MN613056 | MN613077 | MN613098 |
| Erpobdellidae | Erpobdella testacea | AF116027 | AF462025 | AF116003 | AY425370 |
| Erpobdellidae | Erpobdella triannulata | DQ235602 | DQ235592 | DQ235612 | HQ336366 |
| Erpobdellidae | Erpobdella triannulata | HQ336347 | HQ336353 | DQ235614 | HQ336367 |
| Erpobdellidae | Mooreobdella melanostoma | AF116025 | AF462027 | AF115999 | AY425395 |
| Haemopidae | Haemopis caeca | AY040702 | AY425419 | AY040687 | AY425376 |
| Haemopidae | Haemopis kingi | OK447797 | OK489304 | OK489319 | OK486532 |
| Haemopidae | Haemopis marmorata | OK447840 | OK489305 | OK489313 | OK486525 |
Note:
The accession numbers highlighted with asterisks in bold indicate newly added data, and a dash (“−”) indicates that no sequences are available.
Results
General features of mitochondrial genomes and nuclear rDNAs
The newly assembled mitochondrial genome of B. cf. gwalagwalensis is AT-rich (71.9%) circular mapping molecule (total length, 14,847 bp), with an average coverage of 209-fold. The components of B. cf. gwalagwalensis mitogenome consist of 13 protein-coding genes (PCGs), 22 transfer RNAs (tRNAs), 2 rDNAs (12S and 16S rDNA), and a possible control region (451 bp, the longest non-coding reign is located between tRNA R and tRNA H). All mitochondrial genes of Barbronia are encoded on the same strand, and gene order are generally consistent with previously published erpobdellid mitogenomes (Fig. 1 and Table 3). Among 13 Barbronia PCGs, both NAD2 and NAD5 genes are inferred to use ATT as an initiation codon, the COX3 gene is initiated with ATA, and the remaining genes used ATG as a start codon. The predicted secondary structures of 22 tRNAs had a similar clover leaf shape (Fig. 2), and these tRNAs range in size from 60 to 69 bp, with the shortest tRNA gene being tRNA L and the longest one being tRNA Q. The complete length of nuclear 18S rDNA and 28S rDNAs are 1,876 and 2,863 bp, respectively, the former one is identical but longer than the partial 18S rDNA sequence (AY786462) of B. gwalagwalensis.

Figure 1: Comparison of the newly assembled mitochondrial genome (OQ339201) of Barbronia cf. gwalagwalensis with six previously published erpobdellids using BLASTn.
The outermost slot (in black) displays 22 tRNAs, the 12S and 16S rDNA, and 13 coding gene regions on the mitochondrial genome (14,847 base pairs in length). Each of the six inner slots represents regions of the mitochondrial genome (OQ339201) where a BLAST hit occurred in one of the six erpobdellid specimens. Empty regions indicate where there were no BLAST hits between the newly assembled mitochondrial genome and the six previously published erpobdellid specimens using an expect value cutoff of 1e-10, alignment length cutoff of 100, and percent identity cutoff of 80%.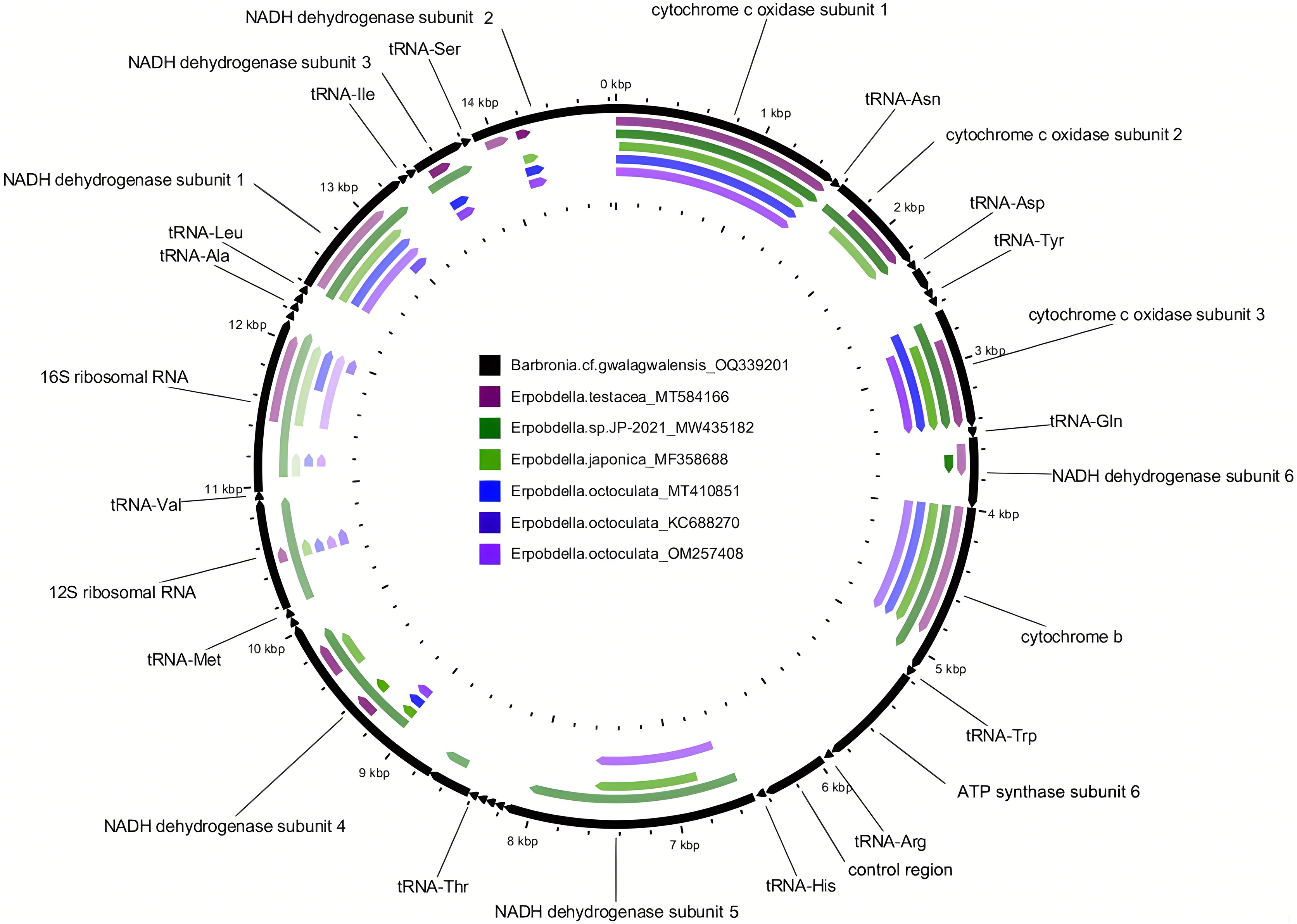{kind=link}
| Taxa | Size (bp) | %GC | CDSs | tRNAs | rRNAs | Accession |
|---|---|---|---|---|---|---|
| B. cf. gwalagwalensis | 14,847 | 28.1 | 13 | 22 | 2 | OQ339201* |
| Erpobdella testacea | 14,495 | 26.9 | 13 | 20 | 2 | MT584166 |
| Erpobdella japonica | 14,725 | 27.9 | 13 | 22 | 2 | MF358688 |
| Erpobdella octoculata | 13,035 | 27.9 | 12 | 20 | 2 | MT410851 |
| Erpobdella octoculata | 15,580 | 27.5 | 13 | 22 | 2 | OM257408 |
| Erpobdellidae sp. | 14,746 | 30.2 | 13 | 22 | 2 | MW43152 |
| “Erpobdella octoculata” | 14,407 | 28.4 | 13 | 22 | 2 | KC688270 |
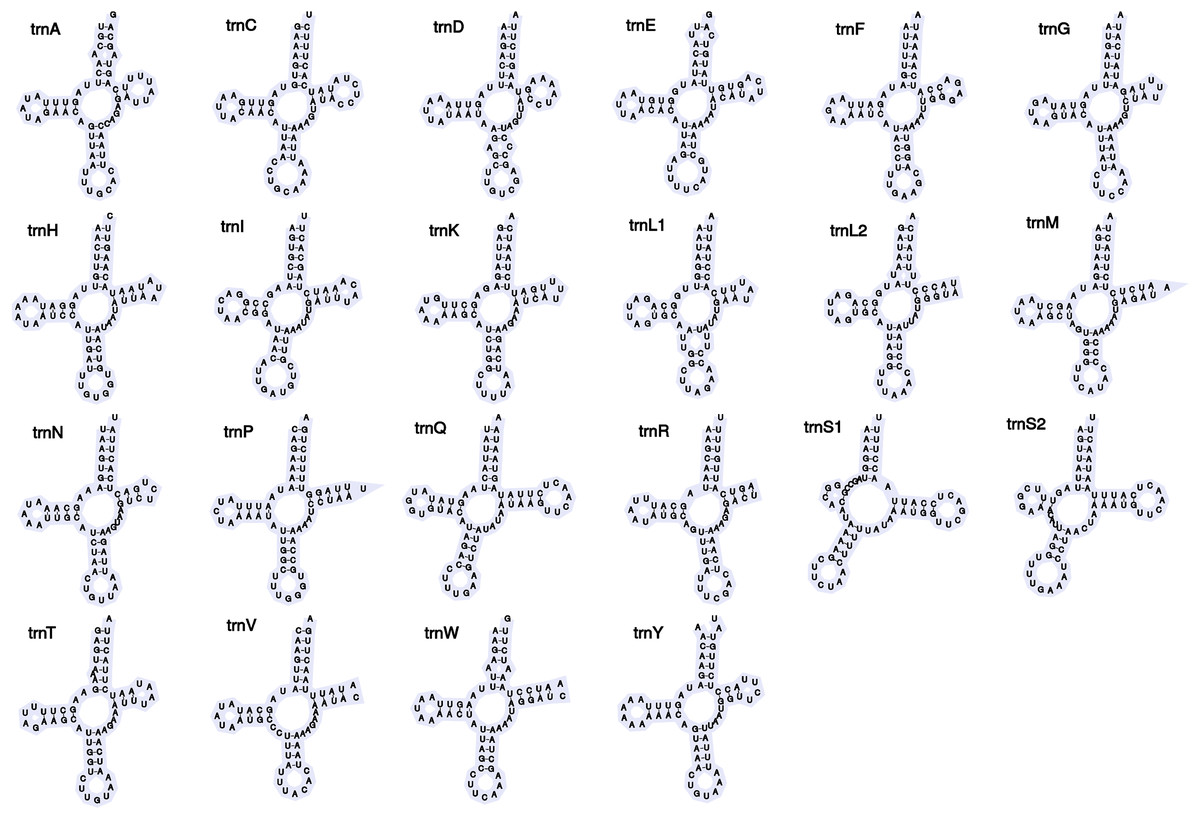
Figure 2: Secondary structures of tRNA genes in the mitogenome of Barbronia cf. gwalagwalensis.
{kind=link}
The outcome of single-locus-based species delimitation
The B. weberi groups and the B. gwalagwalensis group listed in Table 1 correspond to two main well-supported (PP > 0.99) clades in COI phylogenetic analysis (Fig. 3). All members of the B. weberi group are consistently recognized as a well-supported PSH, including the previous described taxa B. weberi, B. weberi formosana, B. arcana, Barbronia wuttkei (clade A in Fig. 3). The primary species hypothesis (PSH) of B. weberi groups proposed by bPTP are consistent with bGMYC, but the difference between two results is the species delimitation of individuals listed in the B. gwalagwalensis group (clade B in Fig. 3). In the bPTP results, the five individuals of the B. gwalagwalensis group in clade B are divided into two PSHs. One well supported PSH comprise only B. cf. gwalagwalensis, and the other moderately supported PSH encompass the paratype of B. gwalagwalensis (GenBank accession number: AY786455) and the remaining specimens (MF458701, MN295405 and MN503261, Fig. 3 and Table 1). However, the five individuals of the B. gwalagwalensis group are either split into four well-supported PSHs or collectively classified as one moderate PSH in the bGMYC analysis (Fig. 3). In the four well-supported PSHs, one PSH contains MF458701 and AY786455, and each of the remaining PSHs is formed separately from MN295405, MN503261and OQ339201. Barbronia cf. gwalagwalensis (OQ339201) alone is consistently recognized as a separately well-supported PSH in both bGMYC and bPTP analyses (threshold 0.95–1), although it is also somewhat classified with the paratype B. gwalagwalensis (AY786455) in a moderately supported PSH (threshold 0.5–0.9) by bGMYC. Moreover, the uncorrected p-distance of COI sequences between B. cf. gwalagwalensis (OQ339201) and the paratype B. gwalagwalensis (AY786455) is 4.4% (Fig. 4), which is substantially higher compared to the p-distance (ranged from 0.2% to 1.2%) among four individuals in clade B (Fig. 4) collected from Myanmar (MN295405), South Korea (MN503261), South Africa (AY786455) and France (MF458701) (Fig. 4 and Table 1). The p-distance between above four individuals and B. cf. gwalagwalensis ranged from 3.8% to 4.5%. In contrast, the p-distances of seven B. weberi individuals in clade A (Fig. 3) sampled from South Africa, the United States, Mexico, Costa Rica, and Germany are less than 0.8%, and the interspecific uncorrected p-distance between the B. weberi group and the B. gwalagwalensis group (clade A and clade B, respectively, Fig. 3) ranged from 6.4% to 8.7%. Considering the result of species delimitation and the sufficiently large p-distance mention above, the current specimen (OQ339201) is treated as B. cf. gwalagwalensis.
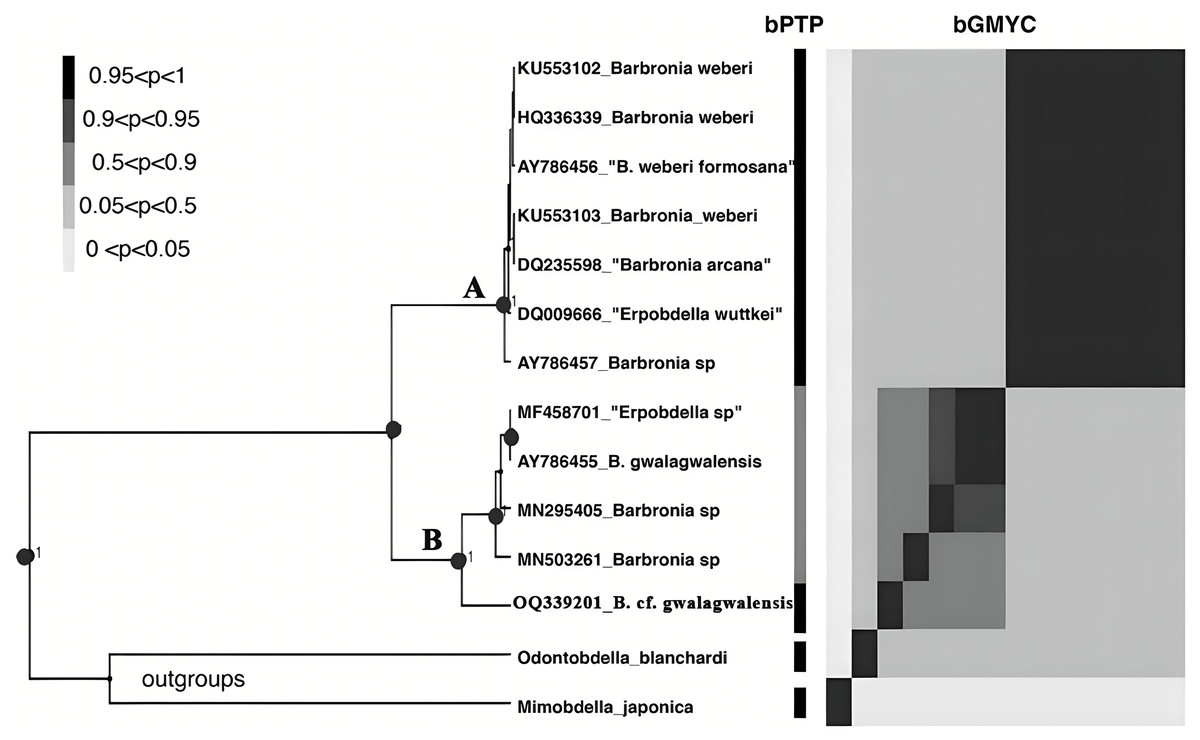
Figure 3: The results of the primary species hypothesis (PSHs) based on COI data.
The bPTP and bGMYC results were summarized and visualized, the left side was the maximum clade credibility tree from BEAST analyses, and the species delimitation results, using bPTP and bGMYC methods based on COI data, were showed on the right side, with colors corresponding to the posterior probability of same primary species hypotheses (PSHs) under a specific threshold (at the upper left). The accession numbers and taxon names in GenBank presented besides underlines at the tip of the tree. Clade A and clade B referred to the B. weberi group and the B. gwalagwalensis group, respectively.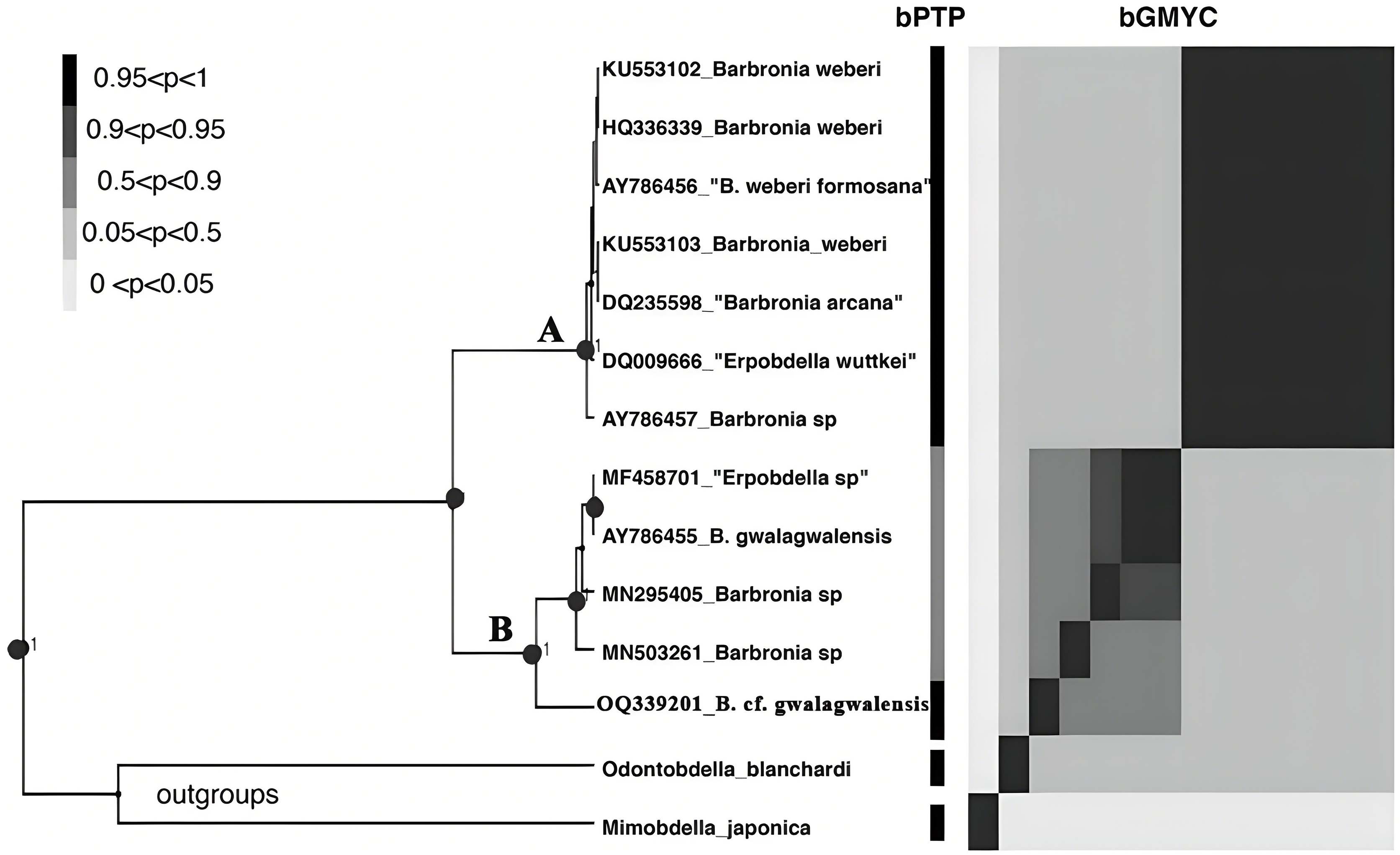{kind=link}

Figure 4: Summary of the bGMYC and bPTP species delimitation methods using the mitochondrial data set.
The uncorrected p-distance matrix corresponding to the alignment of the Barbronia COI sequences. The uncorrected p-distance was calculated in MEGA X using the pairwise deletion option, the uncorrected p-distances were visualized in the upper triangular portion of this matrix, with a color bar (0~10% uncorrected p-distance). The intraspecific and interspecific uncorrected p-distances were represented by the red and blue circles, and the size of circles indicate the value of the corresponding uncorrected p-distances which were listed in triangular portion of this matrix. At the left, the accession numbers and taxon names in GenBank presented besides underlines, corresponding to the numbers from 1 to 12 at the top.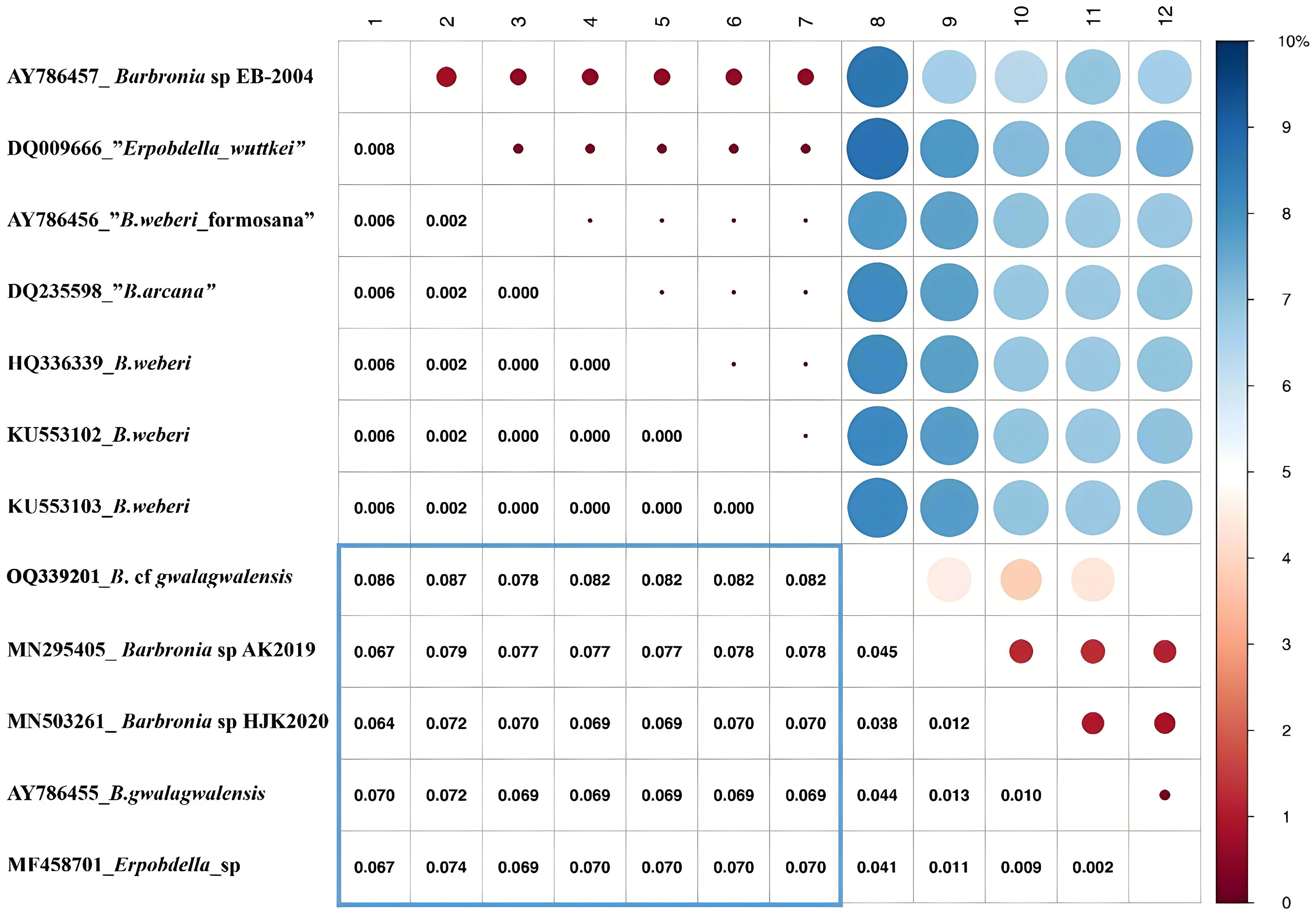{kind=link}
Phylogenetic relationships within Erpobdelliformes
The best-fit model for each partition, GTR + I + G, was selected using PartitionFinder 2. Three Haemopis leeches belonging to Haemopidae (Hirudiniformes) were used as outgroups both in Bayesian Inference (BI) analysis and the maximum likelihood (ML) analysis. Both the BI tree and ML tree topologies show generally well-supported for four main monophyletic groups (PP = 1.00, and BS > 95%), despite differences in the placement of the four clades (Fig. 5). The first clade (PP = 1.00, and BS = 99%) is Salifidae represented by nine specimens from five genera (Barbronia, Mimobdella, Odontobdella, Linta, and Salifa), the second clade (PP = 1.00, and BS = 99%) is Orobdellidae consisted of 18 Orobdella species, the third one (PP = 1.00, and BS = 95%) is Gastrostomobdellidae sensu stricto on behalf of three Gastrostomobdella species, and the last one is a well-supported Erpobdellidae clade (PP = 1.00, and BS = 100%) of 18 Erpobdella species. Barbronia is a well-supported (PP = 1.00, BS = 100%) monophyletic group, and its closely related clade consisted of Mimobdella and Odontobdella. The genus Salifa is recovered as paraphyletic with the inclusion of Linta be. Gastrostomobdellidae sensu stricto (containing genus Orobdella only), but not Gastrostomobdellidae sensu lato (Orobdella + Gastrostomobdella), is a well-supported monophyletic group. The monophyly of Erpobdelliformes was well supported in Bayesian Inference (BI) analysis, with a posterior probability (PP) of 1, but it was not corroborated by the maximum likelihood (ML) analysis.
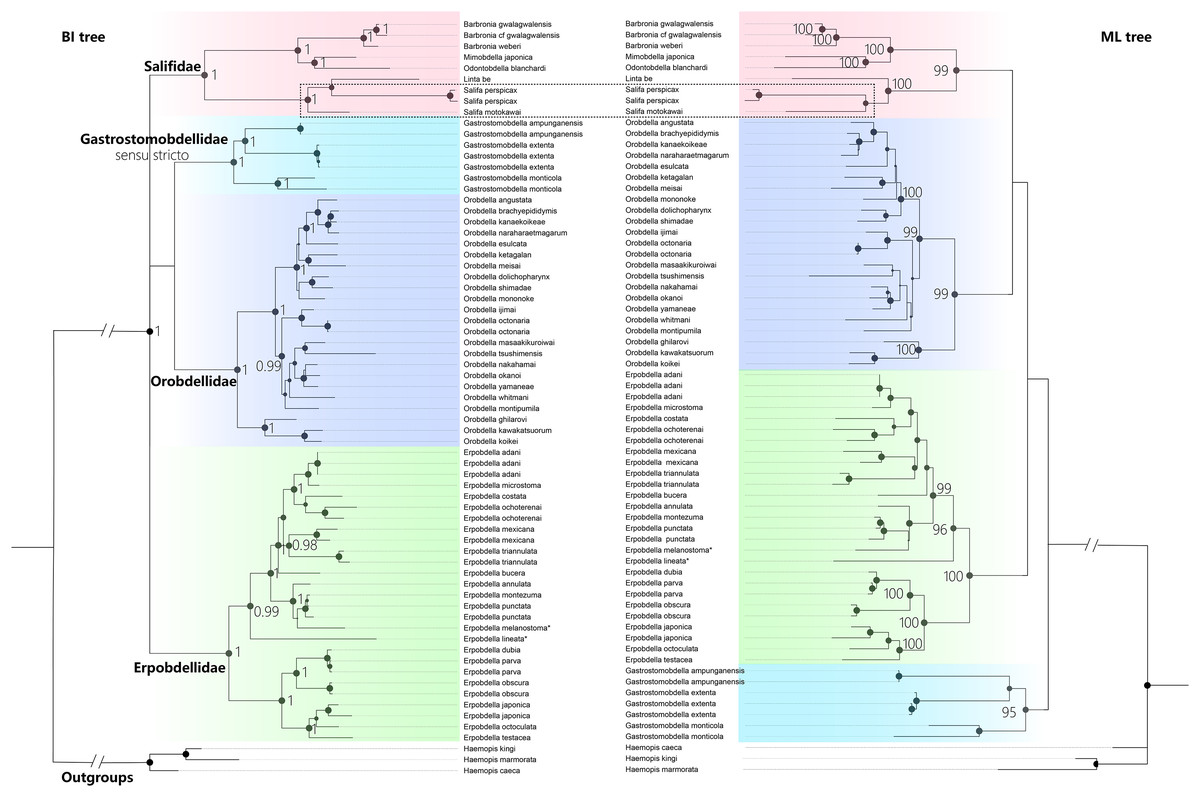
Figure 5 : The Comparison of BI and ML trees constructed using a concatenated data of both mitochondrial genes (COI, 12S rDNA) and nuclear markers (18S and 28S rDNA), respectively.
The monophyly of the genus Barbronia and four Erpobdelliformes families (Salifidae represented by five genus, Orobdellidae, Gastrostomobdellidae sensu stricto and Erpobdellidae) were well supported. Two Erpobdella species name with an asterisk (*) were the synonym of Dina lineata and Mooreobdella melanostoma, respectively. The size of circles indicates that either posterior probabilities or the bootstrap values of corresponding nodes were estimated in BI or ML analyses. The 28S rDNA of B. gwalagwalensis (AY786449) was not included in the current analyses, since it was a partial conserved region of 28S rDNA but significantly different from other 28S rDNA sequences of Barbronia listed in the Table 1.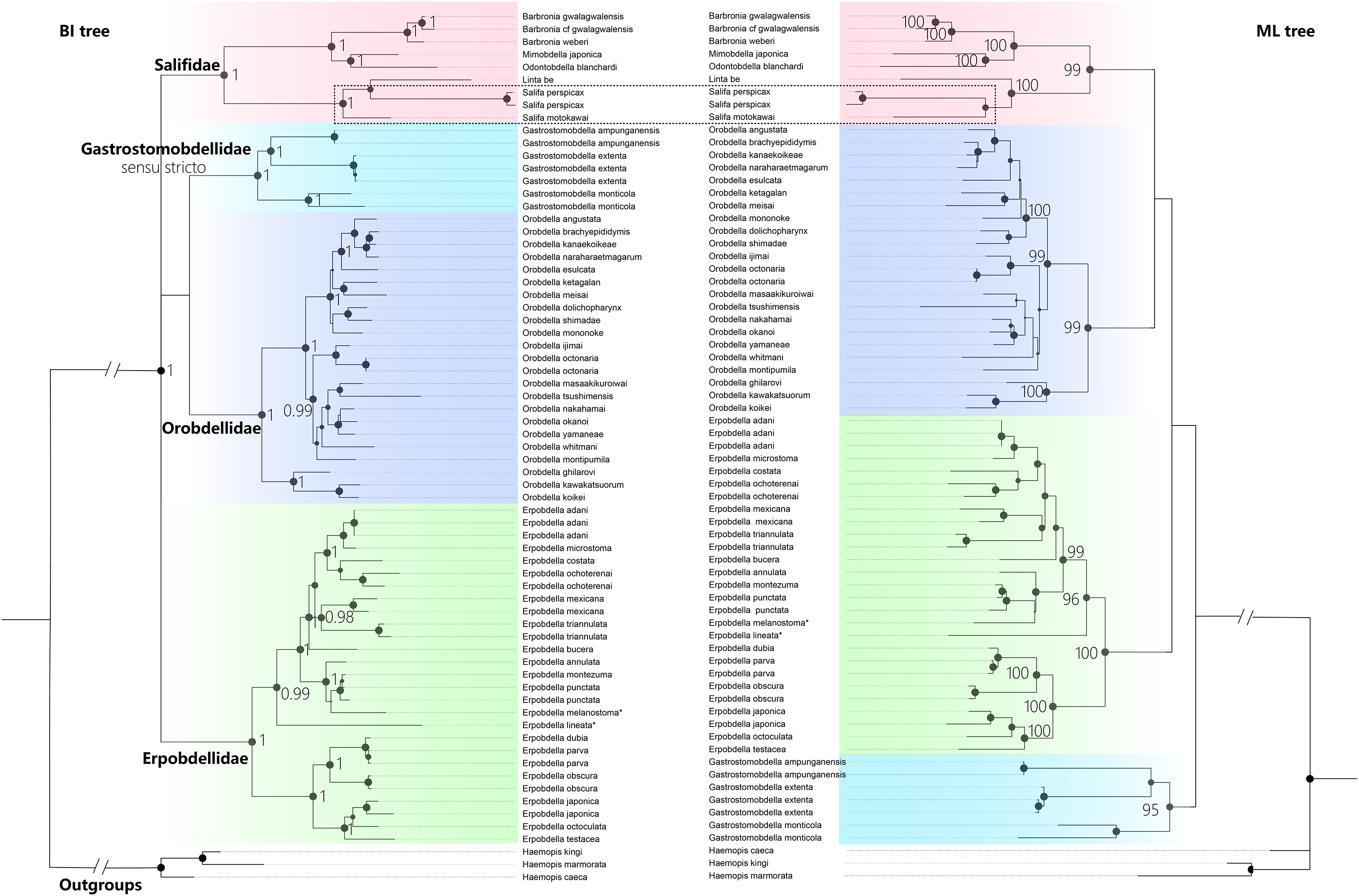{kind=link}
Discussion
To classify Barbronia cf. gwalagwalensis as a distinct species, additional evidence is needed since cryptic speciation is common among clitellates (Erséus & Gustafsson, 2009; Liu et al., 2017; Martinsson & Erséus, 2021). Van Haaren et al. (2004) identified one Dutch specimen as belonging to the B. assiuti/weberi complex, acknowledging that distinguishing the species or identifying it based solely on the morphological characteristics of this single specimen is also not feasible, which highlights the need for careful distinction between individual sample identities and broader species classification. The evidence for new species should encompass both multiple loci-based genetic approaches and morphological analyses from numerous specimens (Dellicour & Flot, 2015; Jorger & Schrodl, 2013; Stengel et al., 2022; Sukumaran, Holder & Knowles, 2021). Initially, this specimen was morphologically identified as the common species B. weberi due to the observation of accessory gonopores (the anterior and posterior ones separately close to the male and female gonophores). The presence of two accessory gonopores in this specimen rules out the possibility of it being B. zhejiangica or B. yunnanensis (Yang, Wang & Zhang, 1997). Molecular results show that this specimen is more closely related to B. gwalagwalensis and the Korean Barbronia specimen (KF966549), the latter of which has been tentatively assigned to Barbronia cf. zhejiangica by Klass et al. (2021). Interestingly, accessory gonopores have also been observed in B. gwalagwalensis (Westergren & Siddall, 2004). However, there are no previous records of B. gwalagwalensis in China. Unfortunately, a thorough morphological examination of this small specimen is unfeasible due to the complete utilization of its body for DNA extraction, a necessary step for constructing a high-quality library in next-generation sequencing. Therefore, the current classification for this specimen is as the taxon B. cf. gwalagwalensis, rather than describing it as a new species. This situation highlights the potential for B. gwalagwalensis to have a broader distribution than previously thought. Initially identified in South Africa, this species has also been found in Myanmar, as noted by Klass et al. (2021). Further studies, involving more samples from across Asia and Europe and detailed morphological analysis, are required to draw definitive conclusions.
Over last two decades, species identification based on molecular data has indeed provided a valuable complement to morphological taxonomy (Mahadani et al., 2022), aided by the increasing availability of genetic techniques (Dellicour & Flot, 2015). In the current study, seven specimens of B. weberi constituted a well-supported clade in the COI phylogenetic results, this clade is also recognized as one valid species both in bGMYC and bPTP analyses (Fig. 3). The current species delimitation results support that the previously proposed taxa (B. arcana, B. weberi formosana and B. wuttkei) are synonyms of B. weberi rather than valid species, which is generally consistent with previous studies (Klass et al., 2021; Oceguera-Figueroa et al., 2011). The COI sequence similarity among the seven individuals within the B. weberi group (clade A in Fig. 3), collected from South Africa, the United States, Mexico, Costa Rica, and Germany are nearly identical. The lack of molecular differentiation (small p-distance) between them can be explained by the large effective population size but low rate of mutation, caused by both the relatively large distribution of invasive B. weberi and the potentially low speciation rate (i.e., reproduce cocoon without cross-fertilization as mentioned in the introduction). However, it is important to note that a gene tree is not always consistent with a species tree when population sizes are large and speciation rates are high (Dellicour & Flot, 2015), single locus-based methods may not be sufficient for delimiting specimens listed in the group of B. gwalagwalensis, especially considering the relatively high genetic diversity between specimens sampled from southern Africa, France, Korea, and China (Fig. 4). On the basis of all available Barbronia data listed in Table 1, all Barbronia specimens tend to lump into one taxon by applying multiple loci-based species delimitation method. This is likely due to insufficient data. In this scenario, utilizing data from both the mitochondrial genome and nuclear loci collected from each specific B. gwalagwalensis specimen (listed in Table 1) is expected to provide a more promising solution for resolving the species delimitation issue in B. gwalagwalensis, rather than relying solely on the partial mitochondrial gene COI.
Mitochondrial genomes and full-length nuclear rDNA have been now routinely applied to assess species boundaries and deep relationships in many phylogenetic studies due to obtaining sufficient data efficiently through NGS (Jia et al., 2023; Moreno-Carmona et al., 2023; Prada et al., 2023). However, a complete mitochondrial genome of any Barbronia species or of the family Salifidae remains unpublished, highlighting a significant gap in our understanding of their systematics. This lack of such data presents a challenge in accurately determining species delimitation and phylogenetic relationships within these groups, underscoring the need for more comprehensive genomic research. In the current study, the first complete mitochondrial genome of salifid leeches is assembled, and compared with whole or incomplete mitochondrial genome sequences of erpobdellids (Fig. 1 and Table 3), representing three nominal species and one unrecognized species in the GenBank. The specimen named Erpobdella octoculata (KC688270 and NC_023927) is likely to be a misidentified specimen or the outcome of sequencing contamination (Oceguera-Figueroa et al., 2016). Clearly, it is far from insufficient using these data to estimate either the species delimitation of Barbronia species or the phylogenetic relationships of Barbronia within Erpobdelliformes (Arhynchobdellida, Hirudinea). Therefore, four conventional molecular markers used in many previous studies of Erpobdelliformes are collected and analyzed, including full-length 18S and 28S rDNA sequences and COI and 12S rDNA sequences extracted from the newly acquired mitochondrial genome. The four well-supported clades, i.e., Salifidae, Orobdellidae, Gastrostomobdellidae sensu stricto, and Erpobdella, are clearly distinguished in the current molecular phylogenetic results. The Barbronia clade is sister to a clade consisting of Odontobdella blanchardi and Mimobdella japonica with well supported in the current study (Fig. 5), and the monophyly of the Barbronia is congruent with previous research (Anderson, Braoudakis & Kvist, 2020; Klass et al., 2021; Nakano et al., 2018; Nakano & Nguyen, 2015; Oceguera-Figueroa et al., 2011). However, the Salifa genus represented by two species is not monophyletic in the current study and in Nakano et al. (2018) result, which is not consistent with previous studies (Klass et al., 2021; Nakano & Nguyen, 2015). The Orobdellidae clade is well supported, which is congruent with previous studies (Nakano, 2016a, 2016b, 2022; Nakano et al., 2018; Nakano & Nguyen, 2015; Nakano, Ramlah & Hikida, 2012). Additionally, previous studies have shown that Erpobdella forms a well-supported clade within the Erpobdellidae family. However, incorporating more species from Gastrostomobdella, rather than just one, provides stronger support for a closer relationship between the Erpobdella clade and the Gastrostomobdellidae family (Nakano et al., 2018; Nakano & Nguyen, 2015). Despite these insights, the robust phylogenetic positions of four families within Erpobdellidae remains uncertain in the current study, these findings underscore the intricate evolutionary relationships within the Erpobdellidae family, indicating the need for further research using more data to fully elucidate the phylogeny of these leech genera. To solve these problems, further studies involving more sampled specimens, or at least simultaneously obtaining more molecular data (i.e., mitochondrial genomes and rDNAs) from a few valuable specimens using NGS are urgently needed. The forthcoming systematics study of B. gwalagwalensis and related species is expected to benefit significantly from the methods employed in our current research, particularly in acquiring mitochondrial genomes and full-length rDNA sequence. This approach effectively bridges gaps between sequences amplified with different primers, notably improving results in cases where conventional primers are less effective (Siddall, 2002; Trontelj, Sket & Steinbrück, 1999).
Conclusions
The first complete mitochondrial genome, full-length 18S and 28S rDNAs of salifid leeches within the family Salifidae are provided. The species delimitation results based on COI data supported the view that the previously proposed taxa (Barbronia arcana, B. weberi formosana, and B. wuttkei) are synonyms of B. weberi. However, the taxonomic status of B. gwalagwalensis and B. cf. gwalagwalensis need to be further studied, necessitating extensive fieldwork across Asia and Europe. Barbronia formed a well-supported clade, including B. weberi, B. gwalagwalensis, and B. cf. gwalagwalensis, and Barbronia is sister to a well-supported clade comprising Odontobdella and Mimobdella. They constitute a strongly supported monophyletic group and are incorporated into the Salifidae family. Additionally, the families Salifidae, Gastrostomobdellidae sensu stricto (containing only the genus Orobdella) and Orobdellidae are all well-supported monophyletic clades. According to the results of this study, the strategy of obtaining both whole mitochondria and nuclear markers from extensively sampled salifid species using NGS is expected to elucidate the species diversity of B. gwalagwalensis and the evolutionary relationship of Salifidae.