
Comparing cestode infections and their consequences for host fitness in two sexual branchiopods: alien Artemia franciscana and native A. salina from syntopic-populations
- Published
- Accepted
- Received
- Academic Editor
- Alex Ford
- Subject Areas
- Biodiversity, Ecology, Parasitology, Zoology
- Keywords
- Cestodes, Sexual Artemia, Syntopic populations, Invasive species, Host impact, Coevolution, Mediterranean salterns
- Copyright
- © 2015 Redón et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2015. Comparing cestode infections and their consequences for host fitness in two sexual branchiopods: alien Artemia franciscana and native A. salina from syntopic-populations. PeerJ 3:e1073 https://doi.org/10.7717/peerj.1073
Abstract
The American brine shrimp Artemia franciscana is invasive in the Mediterranean region where it has displaced native species (the sexual A. salina, and the clonal A. parthenogenetica) from many salt pond complexes. Artemia populations are parasitized by numerous avian cestodes whose effects have been studied in native species. We present a study from the Ebro Delta salterns (NE Spain), in a salt pond where both A. franciscana and native A. salina populations coexist, providing a unique opportunity to compare the parasite loads of the two sexual species in syntopy. The native species had consistently higher infection parameters, largely because the dominant cestode in A. salina adults and juveniles (Flamingolepis liguloides) was much rarer in A. franciscana. The most abundant cestodes in the alien species were Eurycestus avoceti (in adults) and Flamingolepis flamingo (in juveniles). The abundance of E. avoceti and F. liguloides was higher in the A. franciscana population syntopic with A. salina than in a population sampled at the same time in another pond where the native brine shrimp was absent, possibly because the native shrimp provides a better reservoir for parasite circulation. Infection by cestodes caused red colouration in adult and juvenile A. salina, and also led to castration in a high proportion of adult females. Both these effects were significantly stronger in the native host than in A. franciscana with the same parasite loads. However, for the first time, significant castration effects (for E. avoceti and F. liguloides) and colour change (for six cestode species) were observed in infected A. franciscana. Avian cestodes are likely to help A. franciscana outcompete native species. At the same time, they are likely to reduce the production of A. franciscana cysts in areas where they are harvested commercially.
Introduction
The American brine shrimp Artemia franciscana Kellog, 1906 is a key species for the aquaculture industry and a model organism for laboratory research (e.g., in toxicology, genetics or physiology). A. franciscana cysts, particularly originating from Great Salt Lake (USA), have been exported worldwide for aquaculture, the improvement of salt extraction in salt ponds and the pet trade market, facilitating the arrival and spread of A. franciscana outside its natural range (Amat et al., 2005; Ruebhart, Cock & Shaw, 2008; Vikas et al., 2012). In the Western Mediterranean, the introduction of this exotic species is provoking the extinction of native A. salina (Linnaeus, 1758) and A. parthenogenetica Bowen & Sterling, 1978 populations (Amat et al., 2005; Amat et al., 2007; Muñoz et al., 2014). The ability of A. franciscana to outcompete other Artemia species rapidly in the field may be largely explained by a higher reproductive rate that often allows it to eliminate native congeners within a few generations in the laboratory (Browne, 1980; Browne & Halanych, 1989; Amat et al., 2007). However, parasites can also influence biological invasions, depending on their relative impacts on native and alien species (Hatcher, Dick & Dunn, 2006; Prenter et al., 2004; Dunn et al., 2012).
Artemia spp. are intermediate hosts of avian cestodes that can have a major influence on their fitness. In the Mediterranean, native brine shrimps are parasitized by 12 species of avian tapeworms whose final hosts are flamingos, waders, grebes, ducks or gulls (Georgiev et al., 2005; Georgiev et al., 2007; Vasileva et al., 2009). To elucidate the role of parasites in an invasion it is important to study parasite infections in populations of native and alien host species co-ocurring in the same habitat (syntopic populations), but this is difficult because native Artemia have already disappeared from most sites where A. franciscana is detected. Comparisons of allopatric Artemia populations from the southern Iberian Peninsula suggest that A. franciscana populations have lower levels of infections by cestodes than the native sexual A. salina and the clonal A. parthenogenetica (Georgiev et al., 2007; Georgiev et al., 2014; Sánchez et al., 2013). In the present study, we take advantage of a unique opportunity to compare the parasitism of A. franciscana and A. salina in the only site where these two sexual species are known to coexist: the Ebro Delta salterns in north-east Spain.
Sánchez et al. (2012) compared parasitism in syntopic host populations of A. franciscana and A. parthenogenetica in southern France, and found lower cestode diversity and abundance in the invasive host. This could potentially be because the sexual invasive species can resist parasites better than the clonal native one, owing to the importance of genetic recombination in resisting parasites (Red Queen hypothesis: Van Valen, 1973; Moritz et al., 1991; Mee & Rowe, 2006). However, in the case of two sexual species, the relative effects of parasites may be more similar. Given the lack of preexisting data, comparing parasite impacts in two bisexual hosts (A. franciscana and A. salina) is of considerable interest in the context of the biological invasion.
Trophically transmitted parasites such as larval helminths with complex life cycles often induce changes in the physiology, behaviour or appearance of intermediate hosts (“host manipulation”), rendering them more vulnerable to predation and facilitating transmission to final hosts (Barber, Hoare & Krause, 2000; Moore, 2002; Poulin, 2006). In native Mediterranean Artemia populations, cestode infections are associated with a reddish colour, positive phototaxis, gigantism, longer life span, reduced fecundity and higher levels of lipids and glycogen (Thiéry, Robert & Gabrion, 1990; Amat et al., 1991; Robert & Gabrion, 1991; Sánchez et al., 2006; Sánchez, Georgiev & Green, 2007; Amarouayache, Derbal & Kara, 2009; Sánchez et al., 2009a). The pathogenic impact of endemic cestodes on the alien A. franciscana remains unclear, particularly when it co-occurs with native congeners. If A. franciscana is less affected by cestodes, this may help explain its invasion success by aiding it to outcompete native hosts.
In the present study, we compared the cestode infections between A. franciscana and A. salina in a salt pond where they coexisted. We tested the hypothesis that A. franciscana is less susceptible to native cestodes than A. salina, leading to a competitive advantage. Secondly, we compared the infections between this A. franciscana population that is coexisting with a native host (A. franciscana-syntopic population), and an A. franciscana population from a neighbouring pond where there are no congeners, in order to explore if there is a negative relationship between community diversity and disease risk (Schmidt & Ostfeld, 2001; Keesing, Holt & Ostfeld, 2006; Johnson & Thieltges, 2010). Thirdly, we analyzed the consequences of infection by different cestode species for colour change and reproductive activity in both Artemia species. We predicted fewer consequences for the alien host, owing to weaker host–parasite coevolution.
Materials and Methods
Study system and field samples
The Ebro Delta (Province of Tarragona, NE Spain) is the largest wetland area (320 km2) along the Mediterranean coast of Spain and is protected as a Natural Park, Ramsar site and an EU Special Protection Area for birds. Artemia franciscana was first detected in our study area (La Trinitat coastal salterns, 40°35′N, 00°41′E, Fig. 1) in 2007 (Amat et al., 2007). Previously, these salterns supported a tetraploid parthenogenetic population of Artemia (Amat et al., 1995), but this native taxon has not been recorded since.
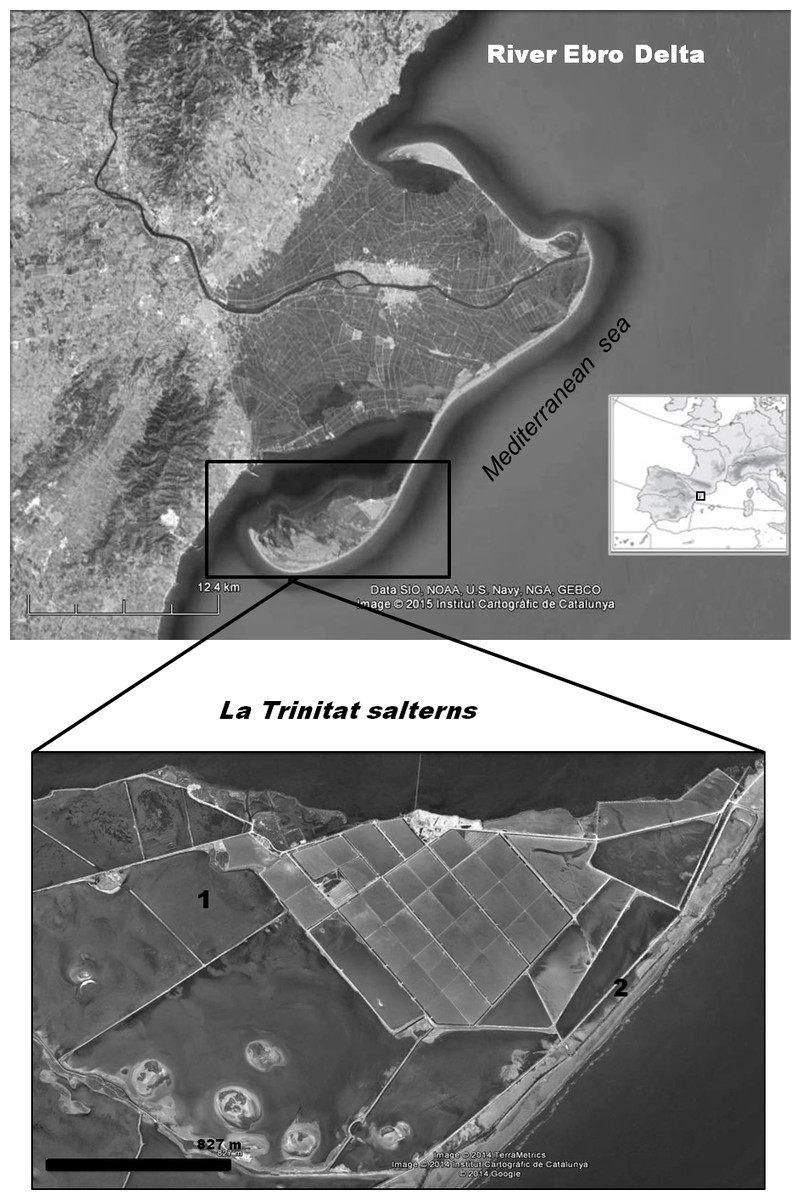
Figure 1: Geographical situation of the study area.
Location of the Ebro Delta (Province of Tarragona, NE Spain) and map of the Ebro Delta salterns La Trinitat indicating the Artemia collection sites: (1) Pond 4, (2) Pond CX.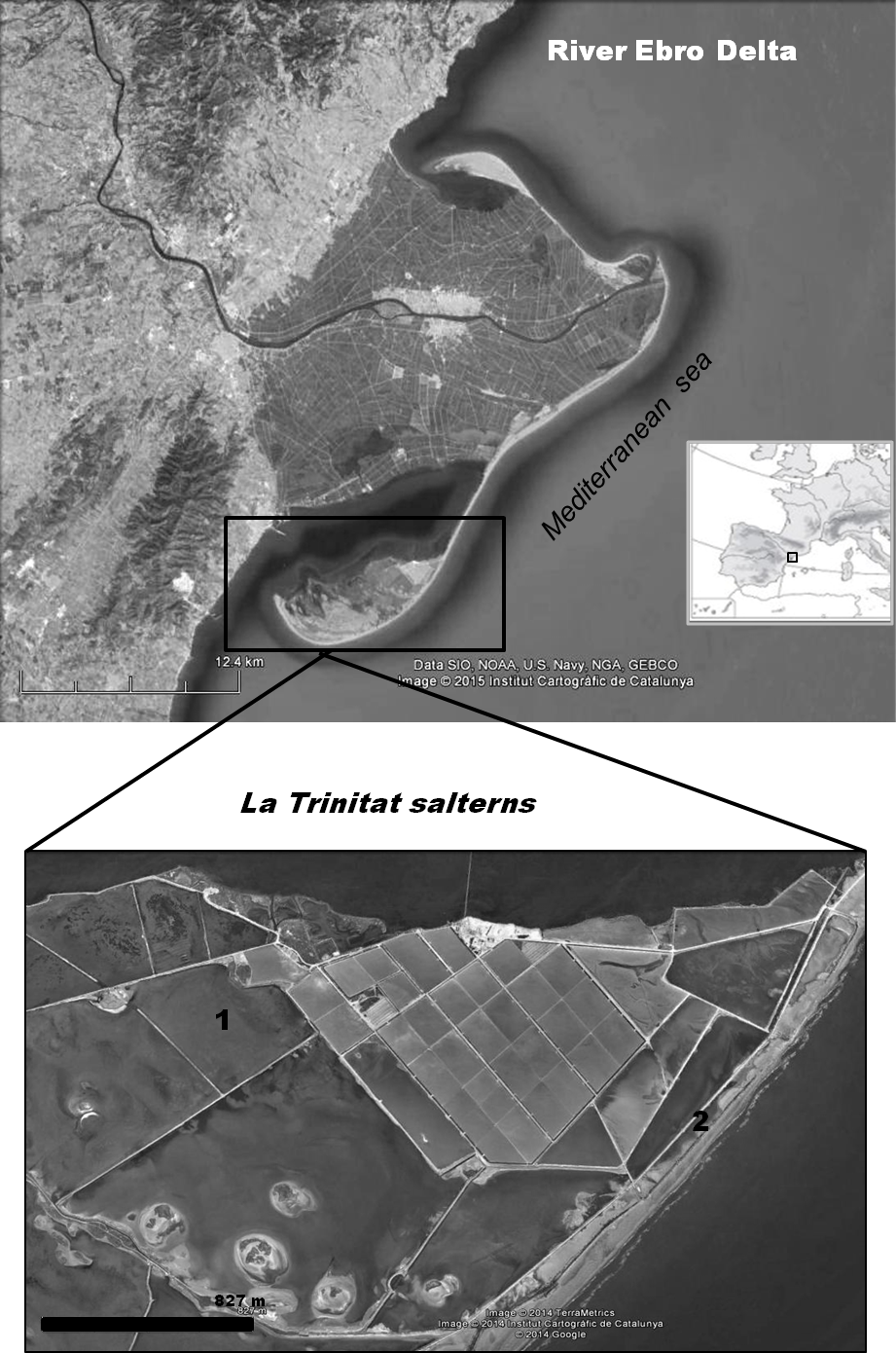{kind=link}
Repeated sampling visits were carried out from 2007 to 2010 inclusive. The present study focuses on samples collected at the following two ponds during a 12 month period from January 2009 to January 2010, when the native A. salina was in coexistence with A. franciscana. Pond CX, a large pond situated between other salt ponds and the sea, isolated from the brine circulation system (Fig. 1), was the only pond in which A. franciscana coexisted with A. salina. Samples were collected monthly from January 2009 to January 2010 (salinity S range = 45–260 g/L, mean ± s.e. = 114.6 ± 18.35; temperature T range = 5–32 °C, mean ± s.e. = 18.1 ± 2.37). Pond 4 was sampled monthly from January to March 2009, then again in January 2010 (S range = 125–150 g/L, mean ± s.e. = 132.5 ± 5.95; T range = 8.5–12 °C, mean ± s.e. = 10.9 ± 0.83). In order to increase the sample size for infected shrimps, additional samples of A. franciscana collected on other dates and in other ponds (S Redón, AJ Green, BB Georgiev, GP Vasileva, F Amat, 2015, unpublished data) were used when considering the influence of cestodes on colouration and reproductive activity of this host species.
Artemia samples were collected from each pond with 160 µm and 500 µm mesh hand-nets and transported alive to the laboratory. Living Artemia individuals, anaesthetized with a few drops of distilled water saturated with chloroform, were examined under a stereomicroscope and juveniles and adults were separated. Juveniles are immature specimens with sexual segments (ovisac or hemipenis) that are not completely developed. Juveniles and adults were identified to species after Amat (1985), Hontoria & Amat (1992a) and Hontoria & Amat (1992b). The proportion of juveniles varied over time (Fig. S1) confirming that both species had multiple generations per year (Amat et al., 2007). Both juveniles and adults were sexed (Amat, 1985). Adult females with empty ovisac and no signs of functional ovaries were classified as castrated and those with embryos, naupliae, or cysts filling the ovisac, or ovulating (oocites moving along the ovaries or filling the oviducts) were classified as ovigerous (Fig. S3). The colour of adults and juveniles was assigned to three categories: dark-red, light-red and not-red (Fig. S2). This research was conducted under a permit from the Ebro Delta Natural Park office provided to FA.
Parasite identification
Juvenile and adults were examined, while lightly anaesthetized, under a stereomicroscope for cestode cysticercoids, until a total of approximately 400 individual shrimps (when available) was reached, including juveniles and adults. After observations of the cysticercoids in situ, each infected specimen was prepared in a temporary glycerol mount and examined under a compound microscope. Identification was based on Georgiev et al. (2005) and Vasileva et al. (2009). More details of sampling protocols are provided by S Redón, AJ Green, BB Georgiev, GP Vasileva, F Amat (2015, unpublished data).
Quantitative analysis of cestode infection and statistics
Several descriptors were applied to the cestode infections in Artemia. Prevalence (P%: proportion of infected individuals in the host population), abundance (MA: mean number of cysticercoids for the total number of shrimps examined) and intensity (MI: mean number of cysticercoids in infected shrimps) were calculated for the overall infection and for each parasite species (terminology following Bush et al., 1997). Also, we quantified species richness (SR: mean number of cestode species present in each Artemia individual) and the species richness in infected specimens (SRinfected).
(i) Host species: invader vs. native
To compare cestode infections between host species, we used monthly samples from pond CX in which a total of 2,150 shrimps were examined (1,252 A. franciscana and 898 A. salina). On a month by month basis, Mann–Whitney U tests were employed to compare the abundance of parasites, the infection intensity and SR between host species, separately for adults and juveniles. Differences in the prevalence of cestodes between host species were analyzed with Fisher Exact tests.
(ii) Influence of coexistence with A. salina on parasitism in A. franciscana
We compared the parasite infections in A. franciscana in a syntopic population (presence of A. salina, pond CX) with those in a single species population sampled simultaneously (pond 4). Using Fisher Exact and Mann–Whitney U tests, we compared the cestode infections in A. franciscana between ponds on the same sampling day during three months (January, February and March 2009), separately for adults and juveniles.
Parasite castration effects
Taking advantage of all A. franciscana samples available (including those from other dates and ponds; S. Redón, AJ Green, BB Georgiev, GP Vasileva, F Amat, 2015, unpublished data), we compared the proportion of infected and uninfected females that were castrated. The castration effect of cestodes in A. franciscana was evaluated by Wilcoxon tests for paired samples, comparing the proportions of castrated shrimps within a given sample, thus avoiding any non-independence of observations of different individuals within the same sample. In contrast, Fisher Exact tests were applied for A. salina, because the small number of samples with this species obliged us to pool them and treat each individual shrimp as an independent observation. Since castration was never recorded in uninfected female A. salina, this pooling was unlikely to bias the observed levels of castration.
Effects of parasites on colour
In order to assess the effects of parasites on the colour of Artemia individuals, we compared the proportion of red individuals (summing the “dark-red” and “light-red” categories, Fig. S2) between infected and non-infected specimens with Fisher Exact tests. The same test was employed to analyze colour-effects between host sexes and host species for both adults and juveniles. In addition, to test the influence of infection intensity on colour, Mann–Whitney U tests were applied to compare the cestode intensity between infected individuals that were red and those that were not. All statistical analyses were carried out using SPSS 15.0 for Windows (SPSS Inc., Chicago, IL, USA).
Results
Comparing parasite loads in A. franciscana and A. salina in syntopic populations
In pond CX where both Artemia species coexisted, A. salina was detected only in colder months, from January to May 2009, and September 2009 to January 2010 (Fig. 2), whereas A. franciscana was present all year long. Nine cestode species were detected in A. franciscana, and seven of these in A. salina (Table 1). These included four species whose adults are parasitic in flamingos (Flamingolepis liguloides, F. flamingo, Gynandrotaenia stammeri and Gynandrotaenia sp.), three in shorebirds (Anomotaenia tringae, A. microphallos and Eurycestus avoceti), one in gulls (Wardium stellorae) and one in shelducks (Fimbriarioides tadornae).
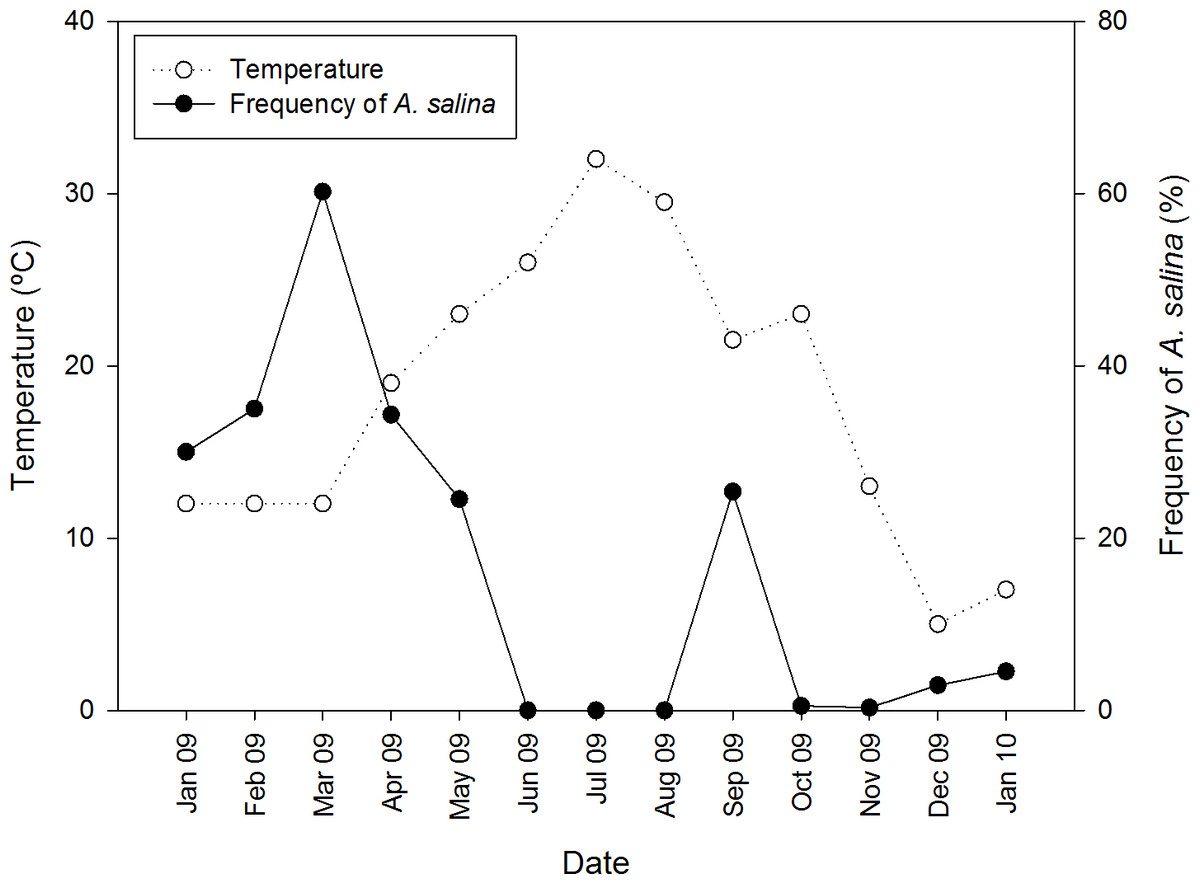
Figure 2: Seasonal variation of temperature and proportion of native A. salina in the brine shrimp community from pond CX throughout an annual cycle.
The right vertical axis indicates the proportion of all Artemia present (whether adults or juveniles) that were A. salina. A. franciscana were present in all months.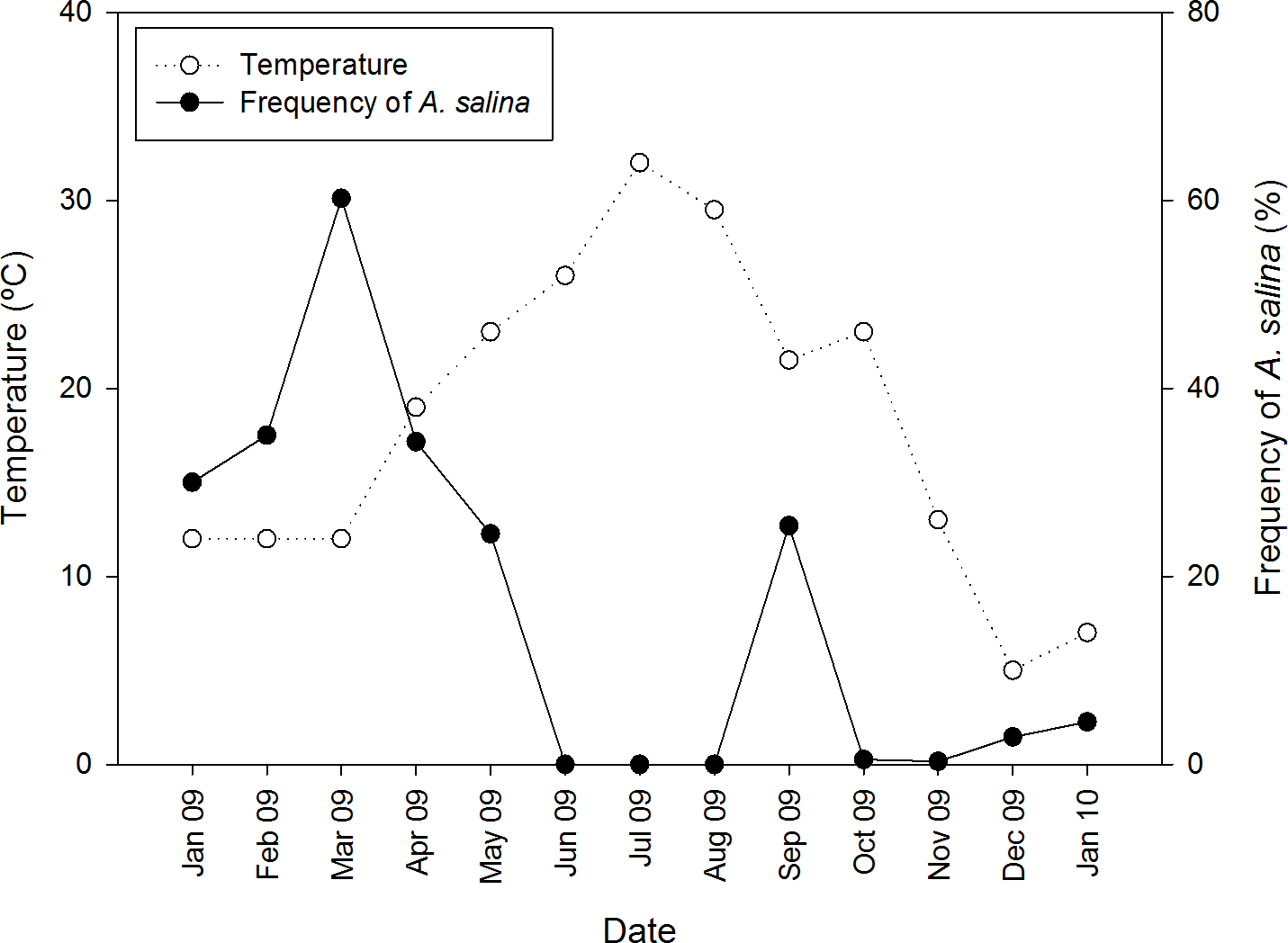{kind=link}
| Host—adults | Host—juveniles | ||||
|---|---|---|---|---|---|
| Cestode species | A. franciscana N = 487 | A. salina N = 381 | A. franciscana N = 765 | A. salina N = 517 | |
| Flamingolepis liguloides (FL) | P% | 0.8 | 17.6 | 1.4 | 8.3 |
| MI | 1.00 ± 0.00 | 1.25 ± 0.07 | 1.18 ± 0.18 | 1.21 ± 0.11 | |
| MA | 0.008 ± 0.004 | 0.221 ± 0.03 | 0.017 ± 0.01 | 0.101 ± 0.02 | |
| Flamingolepis flamingo (FF) | P% | 2.3 | 1.3 | 2.2 | 1.2 |
| MI | 1.09 ± 0.09 | 1.20 ± 0.20 | 1.00 ± 0.00 | 1.17 ± 0.17 | |
| MA | 0.025 ± 0.01 | 0.016 ± 0.01 | 0.022 ± 0.01 | 0.014 ± 0.01 | |
| Wardium stellorae (WS) | P% | 1.4 | 1.0 | 0.0 | 0.0 |
| MI | 1.14 ± 0.14 | 1.00 ± 0.00 | 0.00 | 0.00 | |
| MA | 0.016 ± 0.01 | 0.011 ± 0.01 | 0.00 | 0.00 | |
| Fimbriarioides tadornae (FT) | P% | 1.4 | 0.0 | 0.4 | 0.4 |
| MI | 1.00 ± 0.00 | 0.00 | 1.00 ± 0.00 | 1.00 ± 0.00 | |
| MA | 0.014 ± 0.01 | 0.00 | 0.004 ± 0.002 | 0.004 ± 0.003 | |
| Eurycestus avoceti (EA) | P% | 8.6 | 3.9 | 0.7 | 0.2 |
| MI | 1.02 ± 0.02 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 | |
| MA | 0.088 ± 0.01 | 0.039 ± 0.01 | 0.007 ± 0.003 | 0.002 ± 0.002 | |
| Anomotaenia tringae (AT) | P% | 4.9 | 7.3 | 0.3 | 1.2 |
| MI | 1.08 ± 0.08 | 1.14 ± 0.09 | 1.50 ± 0.50 | 1.17 ± 0.17 | |
| MA | 0.053 ± 0.01 | 0.084 ± 0.02 | 0.004 ± 0.003 | 0.014 ± 0.01 | |
| Anomotaenia microphallos (AM) | P% | 0.8 | 0.0 | 0.0 | 0.0 |
| MI | 1.00 ± 0.00 | 0.00 | 0.00 | 0.00 | |
| MA | 0.008 ± 0.00 | 0.00 | 0.00 | 0.00 | |
| Gynandrotaenia stammeri (GS) | P% | 0.2 | 0.00 | 0.0 | 0.0 |
| MI | 1.00 | 0.00 | 0.00 | 0.00 | |
| MA | 0.002 ± 0.002 | 0.00 | 0.00 | 0.00 | |
| Gynandrotaenia sp. (GSP) | P% | 0.4 | 0.8 | 0.1 | 0.0 |
| MI | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.00 | 0.00 | |
| MA | 0.004 ± 0.003 | 0.008 ± 0.005 | 0.001 ± 0.001 | 0.00 | |
| Total infection | P% | 19.1 | 24.9 | 3.9 | 9.5 |
| MI | 1.15 ± 0.04 | 1.52 ± 0.08 | 1.40 ± 0.13 | 1.41 ± 0.14 | |
| MA | 0.220 ± 0.02 | 0.378 ± 0.04 | 0.055 ± 0.01 | 0.134 ± 0.02 | |
| Species richness ± SE | 0.21 ± 0.02 | 0.32 ± 0.03 | 0.05 ± 0.01 | 0.11 ± 0.02 | |
| SR infected | 1.10 ± 0.03 | 1.28 ± 0.06 | 1.30 ± 0.09 | 1.18 ± 0.06 | |
Notes:
- P%
-
prevalence
- MI
-
mean intensity
- MA
-
mean abundance
- ±SE
-
standard error
- SR infected
-
species richness of infected individuals
Adult brine shrimps
A total of 868 adults were examined in samples with both species present. Of these, 24.9% of A. salina and 19.1% of A. franciscana were parasitized by cysticercoids (Table 1). Cestodes were detected in all five months of co-existence (Fig. 3). For overall infection, A. salina had a higher prevalence than A. franciscana (Table 1), with statistically significant differences in April and May (Fig. 3A).
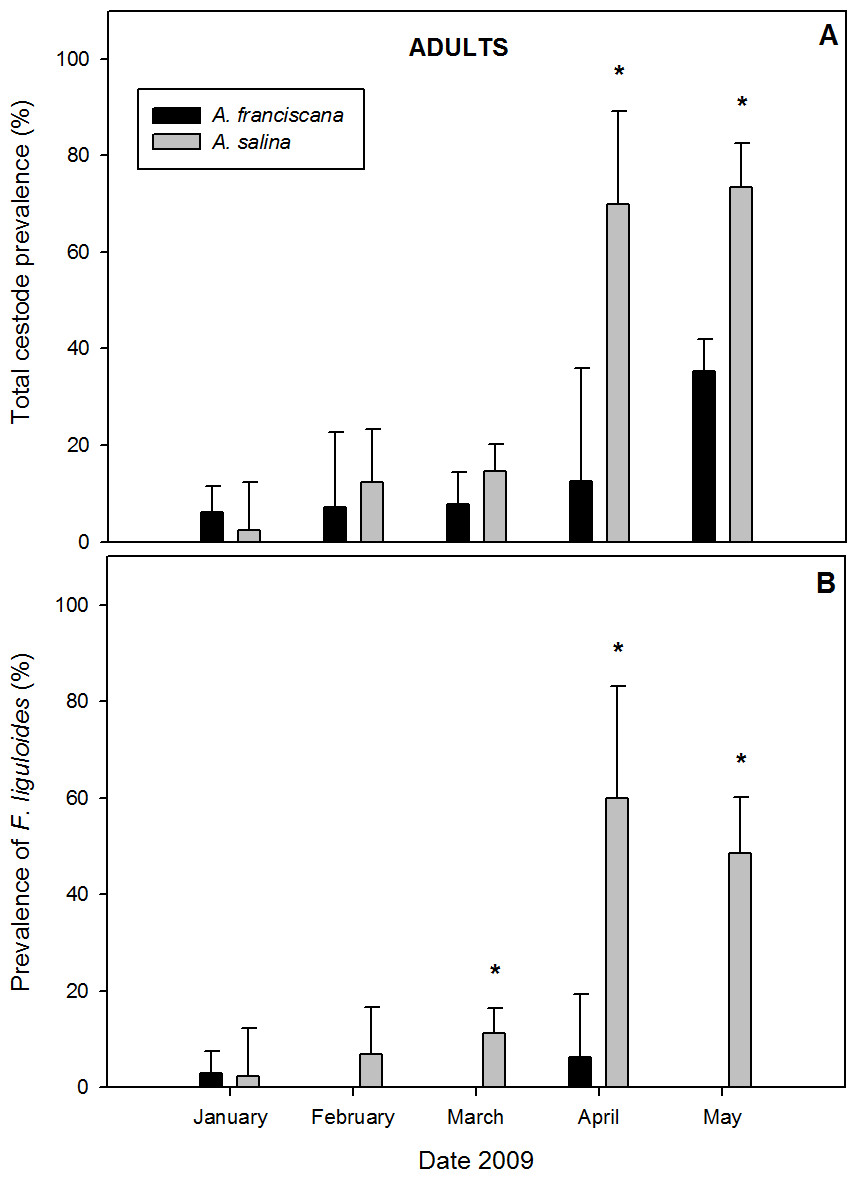
Figure 3: Comparative infection in adults of syntopic brine shrimp populations: A. franciscana and A. salina from pond CX, during months when they were co-existing.
(A) Total cestode prevalence, (B) Prevalence of Flamingolepis liguloides. Bars show upper 95% confidence intervals. * significant differences at p < 0.05 according to Fisher Exact tests.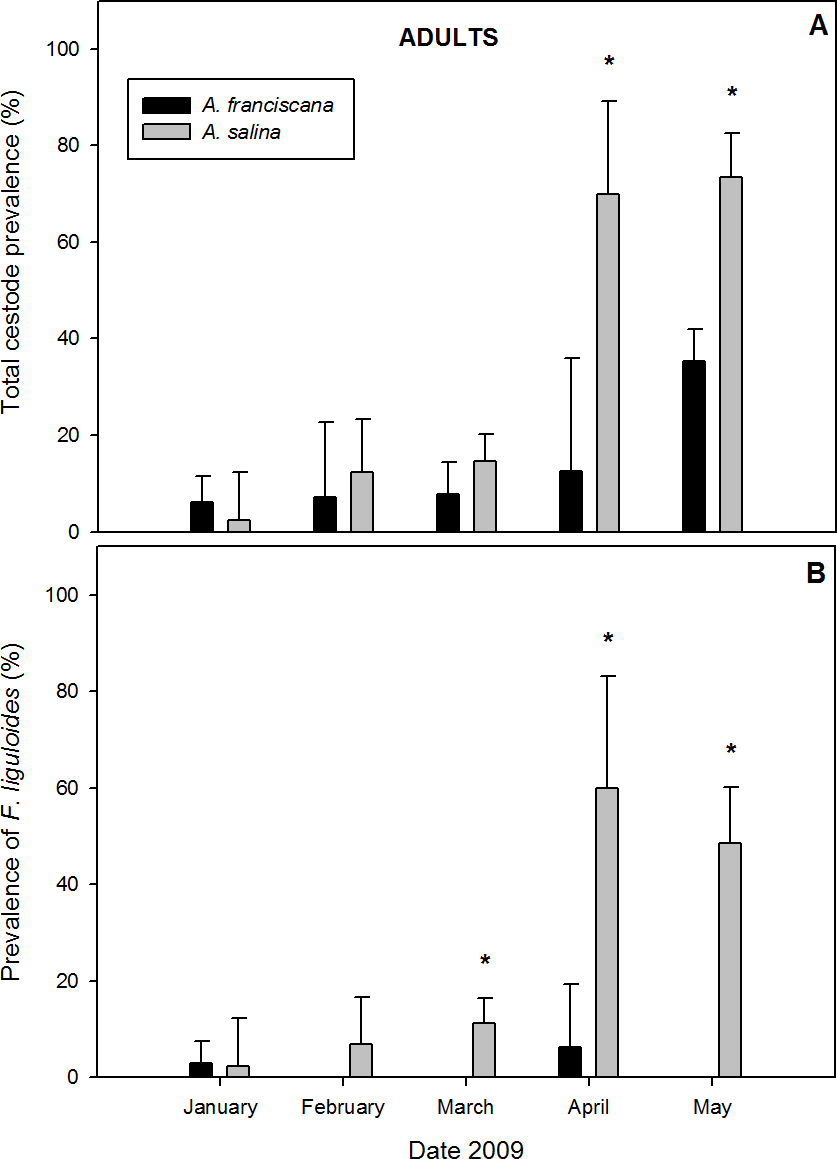{kind=link}
Mean total cestode abundance varied from 0.02 to 1.35 in A. salina and from 0.06 to 0.42 in A. franciscana, and was significantly higher in A. salina in April and May (Mann–Whitney U tests, p < 0.05). Total cestode intensity ranged from 1 to 1.84 in A. salina and 1 to 1.19 in A. franciscana, and was significantly higher in the former in May (p < 0.001). The relative abundance of cestode species varied between hosts. F. liguloides was the most prevalent and abundant parasite in A. salina (Table 1), and its prevalence and abundance were significantly lower in A. franciscana in March, April and May (Fig. 3B). In May, the prevalence and abundance of A. tringae were also significantly higher in A. salina (41.2% and 0.471 ± 0.08, respectively) than A. franciscana (11.6% and 0.126 ± 0.03). Except in January, E. avoceti was the most prevalent and abundant parasite in A. franciscana (Table 1). No significant differences in the intensity of individual cestode species were recorded (Table 1).
A. salina had a higher SR of cestodes (Table 1), being significantly higher in April (0.70 ± 0.15 for A. salina vs. 0.13 ± 0.09 for A. franciscana, p < 0.05) and May (1.12 ± 0.11 vs. 0.40 ± 0.04, p < 0.001). In May, the SR of infected specimens was also significantly higher for A. salina (1.52 ± 0.09 vs. 1.12 ± 0.04, p < 0.001).
Juvenile brine shrimps
A total of 1,282 juveniles were examined in samples with both species present. Of these, 9.5% of A. salina and 3.9% of A. franciscana were parasitized by cysticercoids (Table 1). Cestodes were detected in all months of co-existence except January (Fig. 4). For the overall infection, prevalence and abundance were significantly higher in A. salina from March to May (Fig. 4A). No significant differences were detected in intensity between host species (Table 1).
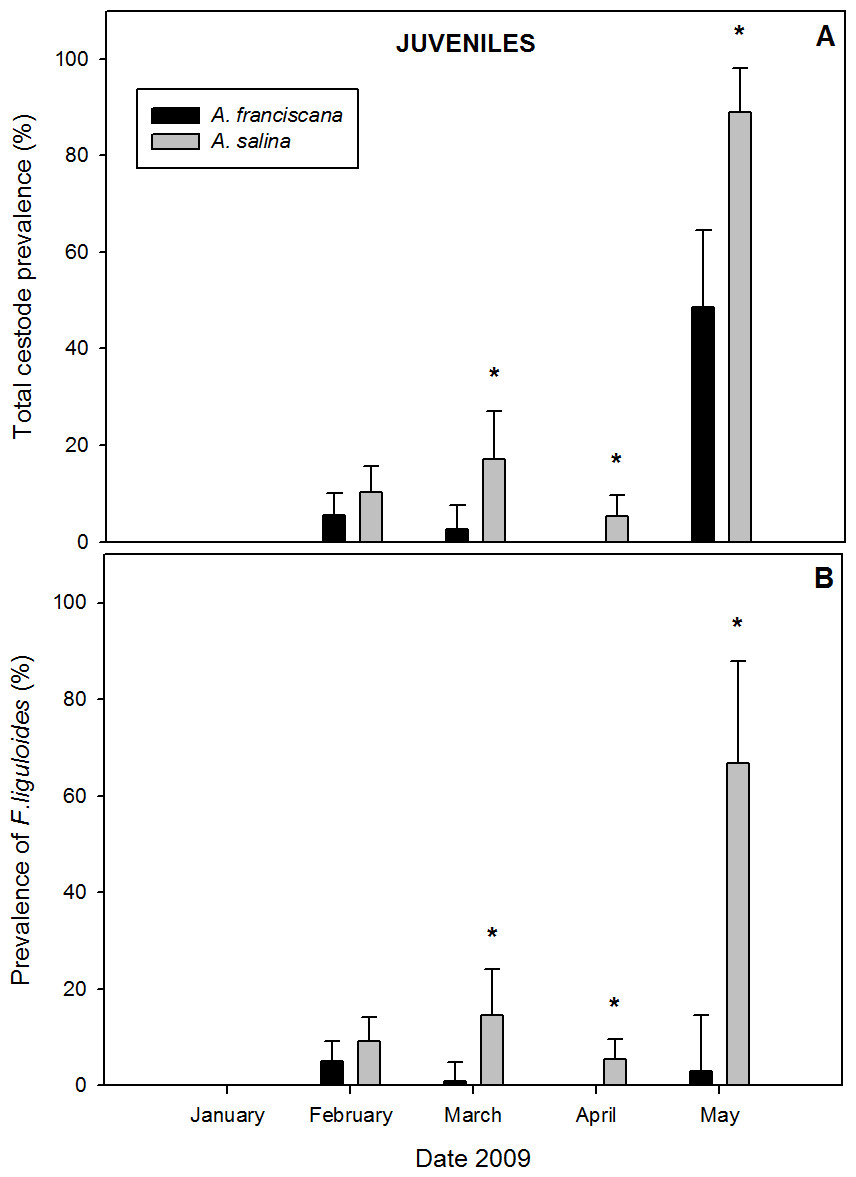
Figure 4: Comparative infection in juveniles of syntopic brine shrimp populations: A. franciscana and A. salina from pond CX, during months when they were co-existing.
(A) Total cestode prevalence, (B) Prevalence of Flamingolepis liguloides. Bars show upper 95% confidence intervals. * significant differences at p < 0.05 according to Fisher Exact tests. No cestodes were recorded in January.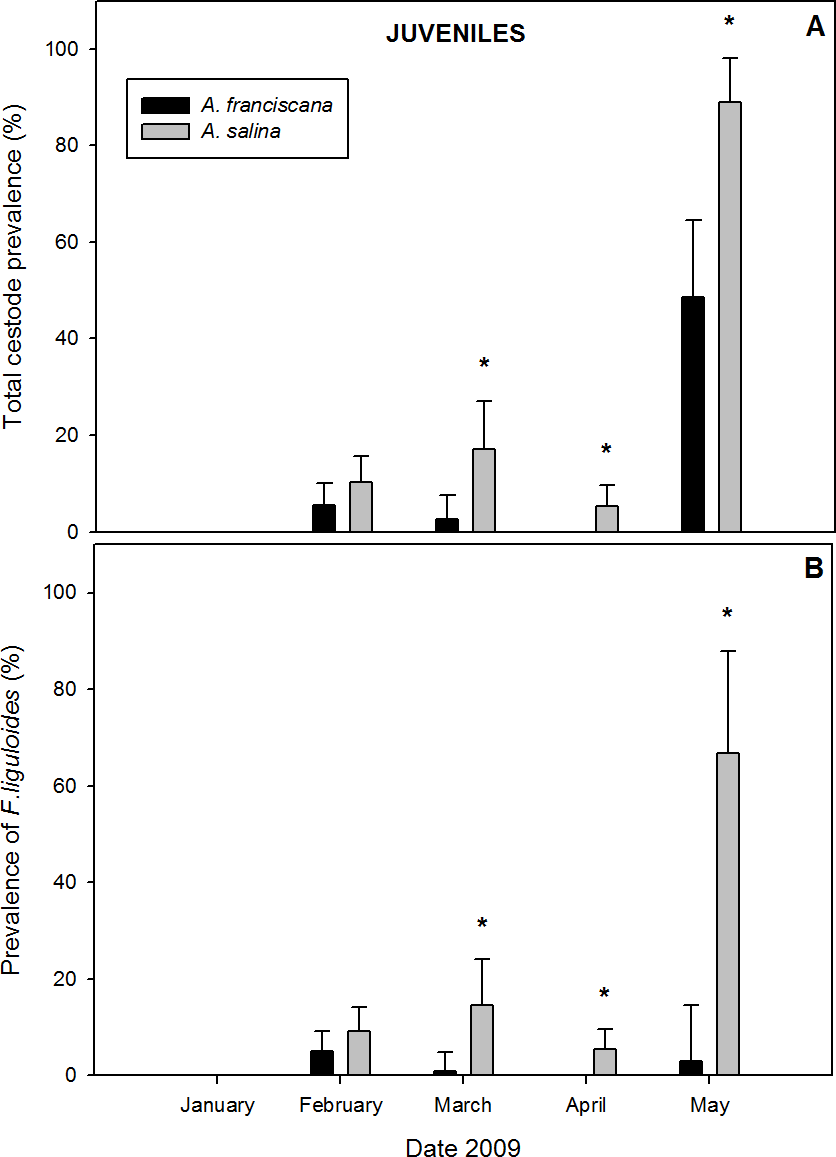{kind=link}
Prevalence and abundance of F. liguloides were significantly higher in A. salina from March to May (Fig. 4B). Prevalence and abundance of A. tringae were also significantly higher in A. salina in May (66.7% vs. 5.7%, and 0.778 ± 0.22 vs. 0.086 ± 0.06). SR was significantly higher for A. salina in March (0.18 vs. 0.04; p = 0.001), April (0.05 vs. 0.00; p < 0.001) and May (1.56 vs. 0.63; p = 0.003). However, no significant differences were detected for SR of infected specimens.
The influence of coexistence with native A. salina on parasitism in A. franciscana
For adult A. franciscana, higher total prevalence and abundance of cestodes were recorded in each of three months in pond CX where it was coexisting with A. salina (AF-syntopic) than in pond 4 where it was the only Artemia species present (AF-single), with statistically significant differences in March (Table 2). Prevalence was significantly higher in AF-syntopic for F. liguloides and E. avoceti in January and March, respectively (Table 2). SR was significantly higher in AF-syntopic in March (Table 2). No significant differences were detected for infection intensity.
| January 2009 | February 2009 | March 2009 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cestode species | AF-single N = 963, S = 125 g/l | AF-syntopic (75.9%) N = 132, S = 56 g/l | p value | AF-single N = 244, S = 130 g/l | AF-syntopic (32.9%) N = 28, S = 65 g/l | p value | AF-single N = 186, S = 150 g/l | AF-syntopic (33.8%) N = 104, S = 70 g/l | p value | |
| FL | P% | 0.3 | 3.0 * | 0.005 | 0.8 | 0.0 | 1.000 | 0.0 | 0.0 | – |
| MI | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.000 | 1.00 ± 0.00 | 0.00 | – | 0.00 | 0.00 | – | |
| MA | 0.0031 ± 0.00 | 0.0303 ± 0.01** | 0.000 | 0.0082 ± 0.01 | 0.00 | 0.631 | 0.00 | 0.00 | 1.000 | |
| FF | P% | 0.7 | 2.3 | 0.011 | 2.5 | 0.0 | 1.000 | 0.0 | 1.0 | 0.359 |
| MI | 1.14 ± 0.14 | 1.00 ± 0.00 | 0.833 | 1.00 ± 0.00 | 0.00 | – | 0.00 | 1.00 | – | |
| MA | 0.0083 ± 0.00 | 0.0303 ± 0.01 | 0.081 | 0.0246 ± 0.01 | 0.00 | 0.402 | 0.00 | 0.0096 ± 0.01 | 0.181 | |
| WS | P% | 0.0 | 0.0 | – | 0.0 | 0.0 | – | 0.5 | 0.0 | 1.000 |
| MI | 0.00 | 0.00 | – | 0.00 | 0.00 | – | 1.00 | 0.00 | – | |
| MA | 0.00 | 0.00 | 1.000 | 0.00 | 0.00 | 1.000 | 0.0054 ± 0.01 | 0.00 | 0.455 | |
| FT | P% | 0.2 | 0.0 | 1.000 | 0.8 | 3.6 | 0.279 | 0.0 | 0.0 | – |
| MI | 1.00 ± 0.00 | 0.00 | – | 1.00 ± 0.00 | 1.00 | 1.000 | 0.00 | 0.00 | – | |
| MA | 0.0021 ± 0.00 | 0.00 | 0.600 | 0.0082 ± 0.01 | 0.0357 ± 0.04 | 0.187 | 0.00 | 0.00 | 1.000 | |
| EA | P% | 1.7 | 0.8 | 0.710 | 0.8 | 3.6 | 0.279 | 0.0 | 4.8 * | 0.006 |
| MI | 1.00 ± 0.00 | 1.00 | 1.000 | 1.00 ± 0.00 | 1.00 | 1.000 | 0.00 | 1.00 ± 0.00 | – | |
| MA | 0.0166 ± 0.00 | 0.0076 ± 0.01 | 0.431 | 0.0082 ± 0.01 | 0.0357 ± 0.04 | 0.187 | 0.00 | 0.0481 ± 0.02* | 0.003 | |
| GS | P% | 0.3 | 0.0 | 1.000 | 0.4 | 0.0 | 1.000 | 0.0 | 0.0 | – |
| MI | 1.00 | 0.00 | – | 1.00 ± 0.00 | 0.00 | – | 0.00 | 0.00 | – | |
| MA | 0.0041 ± 0.00 | 0.00 | 0.521 | 0.0031 ± 0.00 | 0.00 | 0.735 | 000 | 0.00 | 1.000 | |
| GSP | P% | 0.5 | 0.0 | 1.000 | 0.0 | 0.0 | – | 0.0 | 1.9 | 0.128 |
| MI | 1.00 ± 0.00 | 0.00 | – | 0.00 | 0.00 | – | 0.00 | 1.00 ± 0.00 | – | |
| MA | 0.0052 ± 0.00 | 0.00 | 0.407 | 0.00 | 0.00 | 1.000 | 0.00 | 0.0192 ± 0.01 | 0.058 | |
| Total infection | ||||||||||
| P% | 3.7 | 6.1 | 0.232 | 5.3 | 7.1 | 0.658 | 0.5 | 7.7 * | 0.001 | |
| MI | 1.03 ± 0.03 | 1.00 ± 0.00 | 0.917 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.000 | 1.00 | 1.00 ± 0.00 | 1.000 | |
| MA | 0.0384 ± 0.01 | 0.0606 ± 0.02 | 0.204 | 0.0533 ± 0.01 | 0.0714 ± 0.05 | 0.691 | 0.0054 ± 0.01 | 0.0865 ± 0.03* | 0.001 | |
| Species Richness | 0.04 ± 0.01 | 0.06 ± 0.02 | 0.203 | 0.05 ± 0.01 | 0.07 ± 0.05 | 0.691 | 0.01 ± 0.01 | 0.08 ± 0.03 * | 0.001 | |
| SR infected | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.000 | 1.00 ± 0.00 | 1.00 ± 0.00 | 1.000 | 1.00 | 1.00 ± 0.00 | 1.000 | |
Amongst juvenile A. franciscana, total prevalence, abundance and species richness were significantly higher in AF-syntopic in March (as for adults), but significantly lower in AF-syntopic in January (when no infected juveniles of either Artemia species were recorded in the syntopic population). The abundance of F. flamingo in March was also significantly higher for AF-syntopic (Table 3).
| January 2009 | February 2009 | March 2009 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Cestode species | AF-single N = 168 | AF-syntopic (65.7%) N = 134 | p value | AF-single N = 67 | AF-syntopic (50.4%) N = 179 | p value | AF-single N = 248 | AF-syntopic (59.4%) N = 111 | p value | |
| FL | P% | 1.2 | 0.0 | 0.505 | 0.0 | 5.0 | 0.119 | 0.0 | 0.9 | 0.309 |
| MI | 1.00 ± 0.00 | 0.00 | – | 0.00 | 1.00 ± 0.00 | – | 0.00 | 1.00 | – | |
| MA | 0.0119 ± 0.01 | 0.00 | 0.206 | 0.00 | 0.0503 ± 0.02 | 0.062 | 0.00 | 0.0090 ± 0.01 | 0.135 | |
| FF | P% | 1.8 | 0.0 | 0.257 | 1.5 | 1.7 | 1.000 | 0.0 | 1.8 | 0.095 |
| MI | 1.33 ± 0.33 | 0.00 | – | 1.00 | 1.00 ± 0.00 | 1.000 | 0.00 | 1.00 ± 0.00 | – | |
| MA | 0.0238 ± 0.01 | 0.00 | 0.121 | 0.0149 ± 0.01 | 0.0168 ± 0.01 | 0.919 | 0.00 | 0.0181 ± 0.01* | 0.034 | |
| EA | P% | 1.8 | 0.0 | 0.257 | 0.0 | 0.0 | – | 0 | 0.9 | 0.309 |
| MI | 1.00 ± 0.00 | 0.00 | – | 0.00 | 0.00 | – | 0.00 | 1.00 | – | |
| MA | 0.0179 ± 0.01 | 0.00 | 0.121 | 0.00 | 0.00 | 1.000 | 0.00 | 0.0090 ± 0.01 | 0.135 | |
| GS | P% | 0.6 | 0.0 | 1.000 | 0.0 | 0.0 | – | 0.0 | 0.0 | – |
| MI | 1.00 | 0.00 | – | 0.00 | 0.00 | – | 0.00 | 0.00 | – | |
| MA | 0.0060 ± 0.01 | 0.00 | 0.372 | 0.00 | 0.00 | 1.00 | 000 | 0.00 | 1.000 | |
| GSP | P% | 0.6 | 0.0 | 1.000 | 0.0 | 0.6 | 1.00 | 0.0 | 0.0 | – |
| MI | 1.00 | 0.00 | – | 0.00 | 1.00 | – | 0.00 | 0.00 | – | |
| MA | 0.0060 ± 0.01 | 0.00 | 0.372 | 0.00 | 0.0056 ± 0.01 | 0.541 | 0.00 | 0.00 | 1.000 | |
| Total infection | ||||||||||
| P% | 6.0 * | 0.0 | 0.003 | 1.5 | 5.6 | 0.298 | 0.0 | 2.7 * | 0.029 | |
| MI | 1.10 ± 0.10 | 0.00 | – | 1.00 | 1.30 ± 0.15 | 0.727 | 0.00 | 1.33 ± 0.33 | – | |
| MA | 0.0655 ± 0.02* | 0.00 | 0.004 | 0.0149 ± 0.01 | 0.0726 ± 0.02 | 0.165 | 0.00 | 0.0360 ± 0.02* | 0.009 | |
| Species richness | 0.06 ± 0.02* | 0.00 | 0.004 | 0.01 ± 0.01 | 0.07 ± 0.02 | 0.165 | 0.00 | 0.04 ± 0.02* | 0.009 | |
| SR infected | 1.00 ± 0.00 | – | 1.00 | 1.30 ± 0.15 | 1.33 ± 0.36 | – | ||||
Influence of parasites on host colouration
Red colouration was strongly associated with presence of cestodes in both Artemia species, and in both adults and juveniles (Fig. 5 and Table 4). Among infected adults, red colouration was significantly more frequent in A. salina than A. franciscana (Table 4). There were no differences between sexes for either host species in the probability of redness when infected.
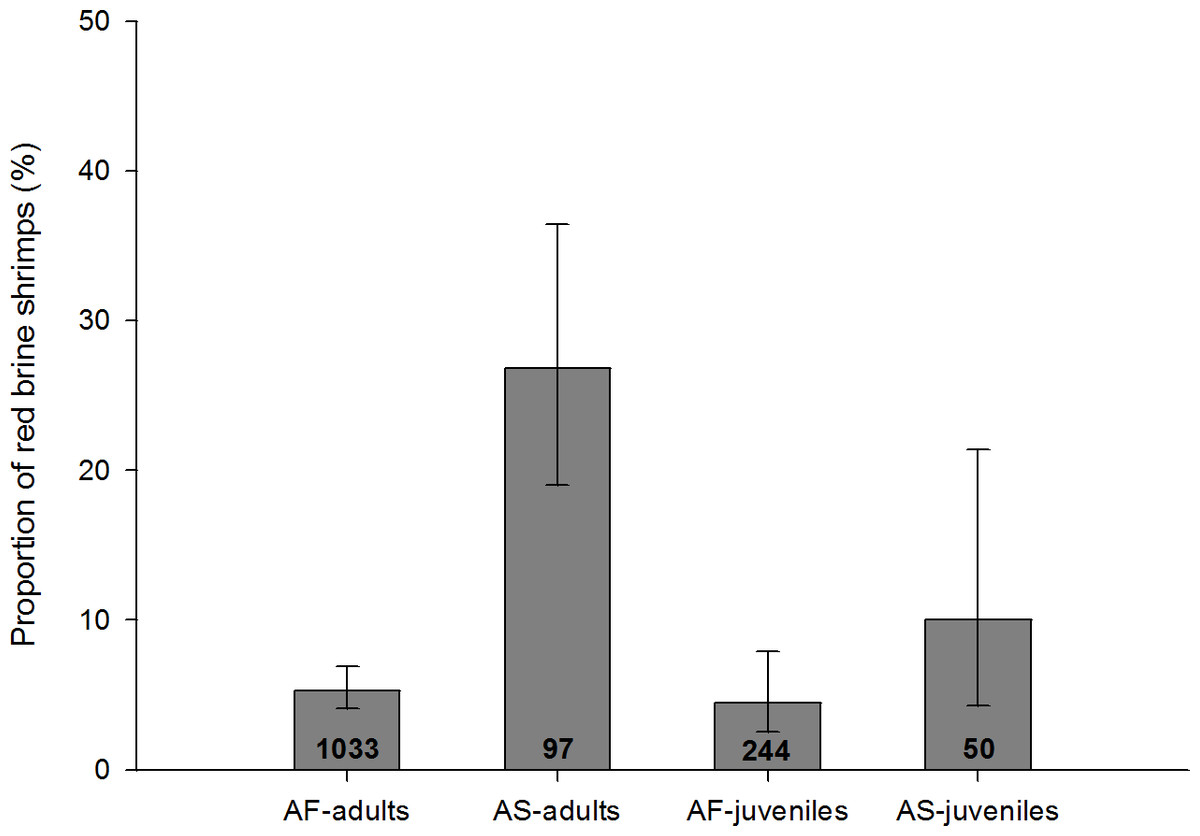
Figure 5: Proportion of infected adults and juveniles with red colouration for A. salina (AS) and A. franciscana (AF).
Bars show 95% confidence intervals. The total number of infected specimens in each group is shown within the columns. Red colouration was not recorded in uninfected A. salina, and was very rare in uninfected A. franciscana (absent in juveniles, 0.02% in adults). For all four categories the proportion of red shrimps is significantly higher in infected individuals (Fisher Exact tests, p < 0.001).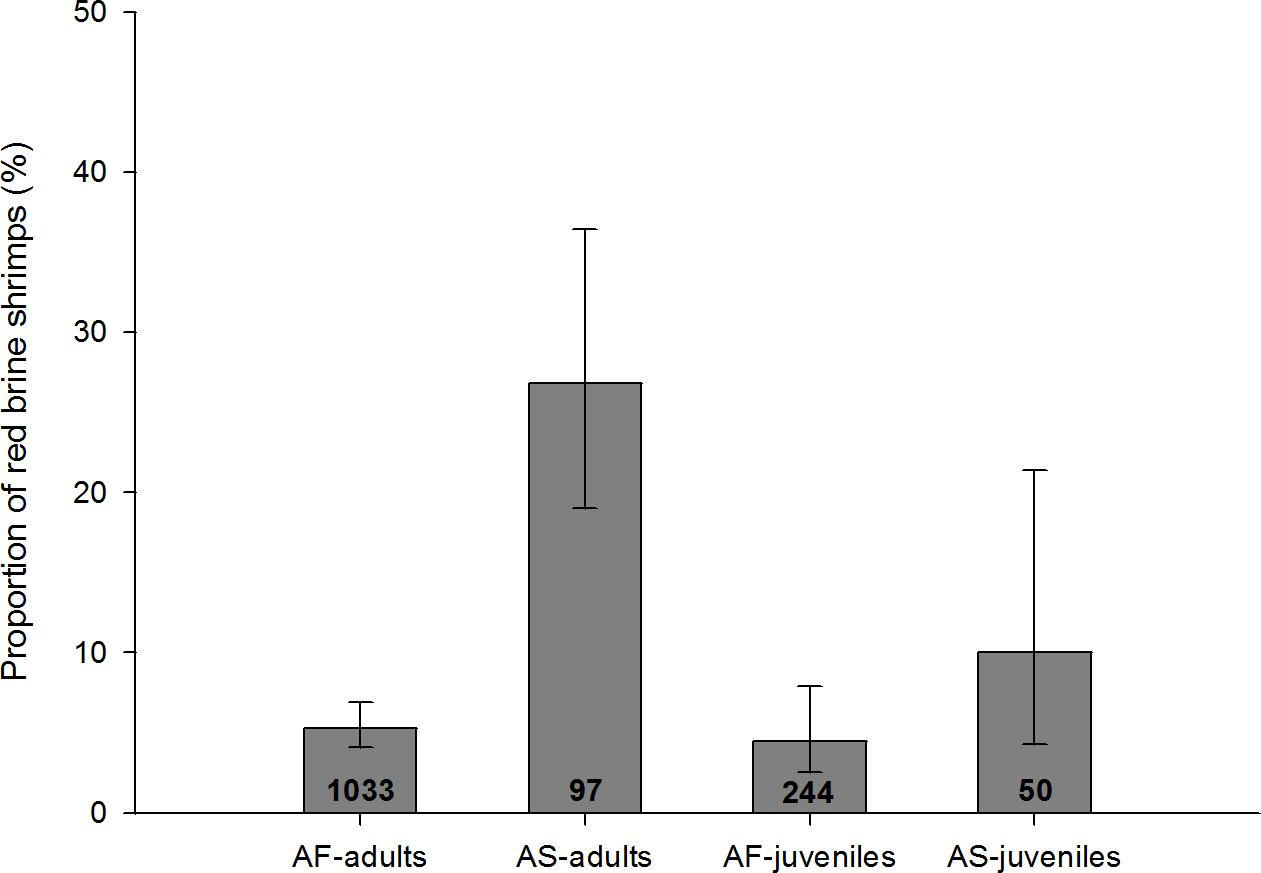{kind=link}
| Adults | Juveniles | |||
|---|---|---|---|---|
| A. franciscana | A. salina | A. franciscana | A. salina | |
| % Red | % Red | % Red | % Red | |
| Uninfected individuals | 0.02% (8,260) | 0% (325) | 0% (8,658) | 0% (507) |
| Infected individuals | 5.3%**a (1,033) | 26.8%**b (97) | 4.5%** (244) | 10%** (50) |
| Infection status | ||||
| Single infections (1 species)—Total | 4.2%**a (931) | 12.2%**b (74) | 4.1%** (221) | 0% (41) |
| Infected only by FL | 2.3%* (86) | 0% (47) | 0% (32) | 0% (35) |
| Infected only by FF | 1.3%* (79) | 0% (2) | 5.3%** (114)a | 0% (5) |
| Infected only by WS | 0% (26) | 0% (3) | 0% (2) | – |
| Infected only by FT | 0% (12) | – | 0% (7) | – |
| Infected only by EA | 4.2%** (620)a | 12.5%* (8)a | 2.3%* (42)a | 0% (1) |
| Infected only by AT | 13.6%**a (22)a | 72.7%**b (11)a | – | – |
| Infected only by AM | 18.2%** (33)a | – | 66.7%** (3)a | – |
| Infected only by GS | 2.3%* (43)a | – | 0% (9) | – |
| Infected only by GSP | 0% (7) | 0% (3) | 0% (2) | – |
| Multiple infections (≥2 species) | 15.7%**a (102) | 73.9%**b (23) | 8.7%**a (23) | 55.6%**b (9) |
For both Artemia species, there was a positive relationship between infection level (in terms of species richness and/or intensity of infection) and the likelihood of red colour.
For infected adults, the total number of cysticercoids was higher in red individuals both for A. franciscana (mean ± s.e. = 2.4 ± 0.18 when red, 1.23 ± 0.02 when not; Mann–Whitney U test, U = 11,280, p < 0.001) and A. salina (2.08 ± 0.20 when red, 1.31 ± 0.07 when not; U = 487, p < 0.001). A similar result was obtained for infected juvenile A. salina (3.8 ± 0.58 when red, 1.16 ± 0.06 when not; U = 4, p < 0.001). Owing to the larger sample size, the positive correlation between infection intensity and probability of red colouration was particularly clear in A. franciscana adults (Fig. 6), for which the same pattern was apparent for the dominant cestode species, E. avoceti (results not shown).
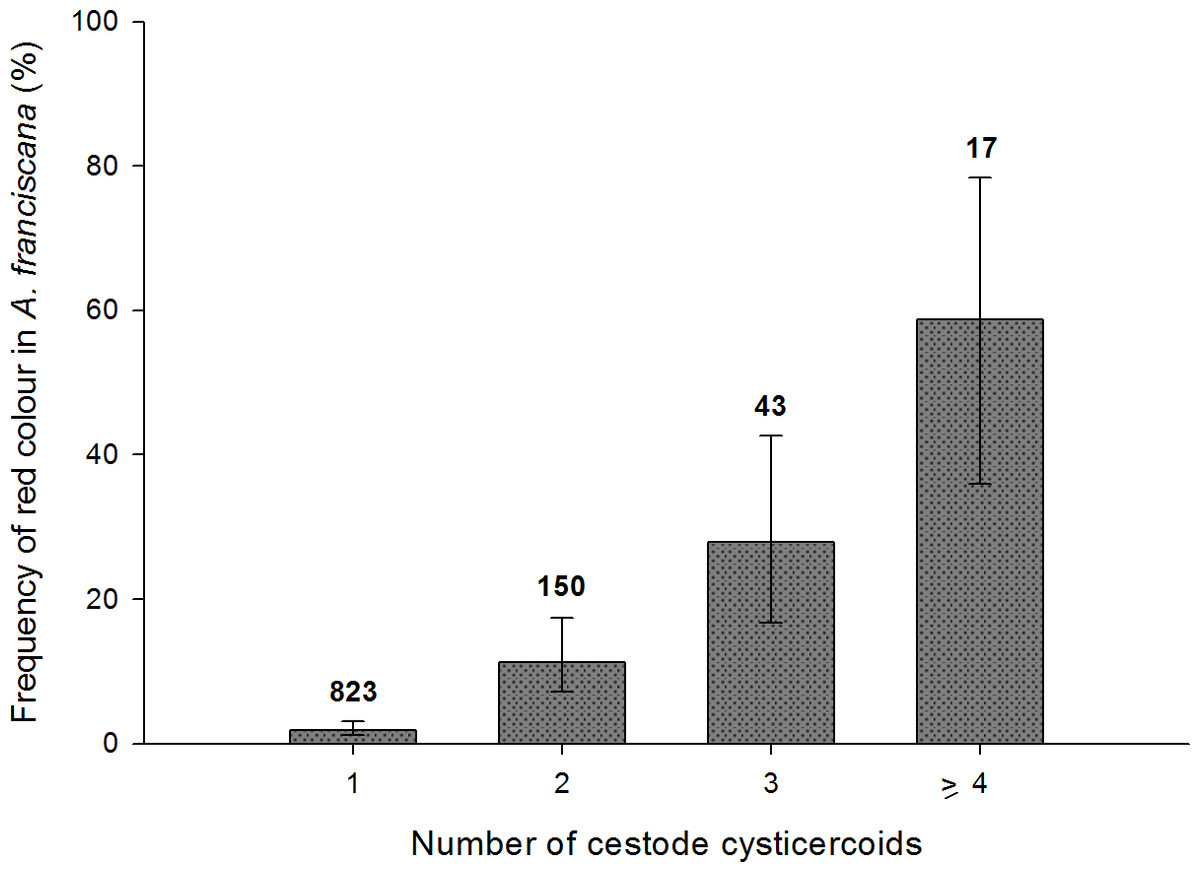
Figure 6: Colour pattern in relation to intensity of infection in adult A. franciscana.
Bars show 95% confidence intervals. The total number of infected specimens in each group is indicated.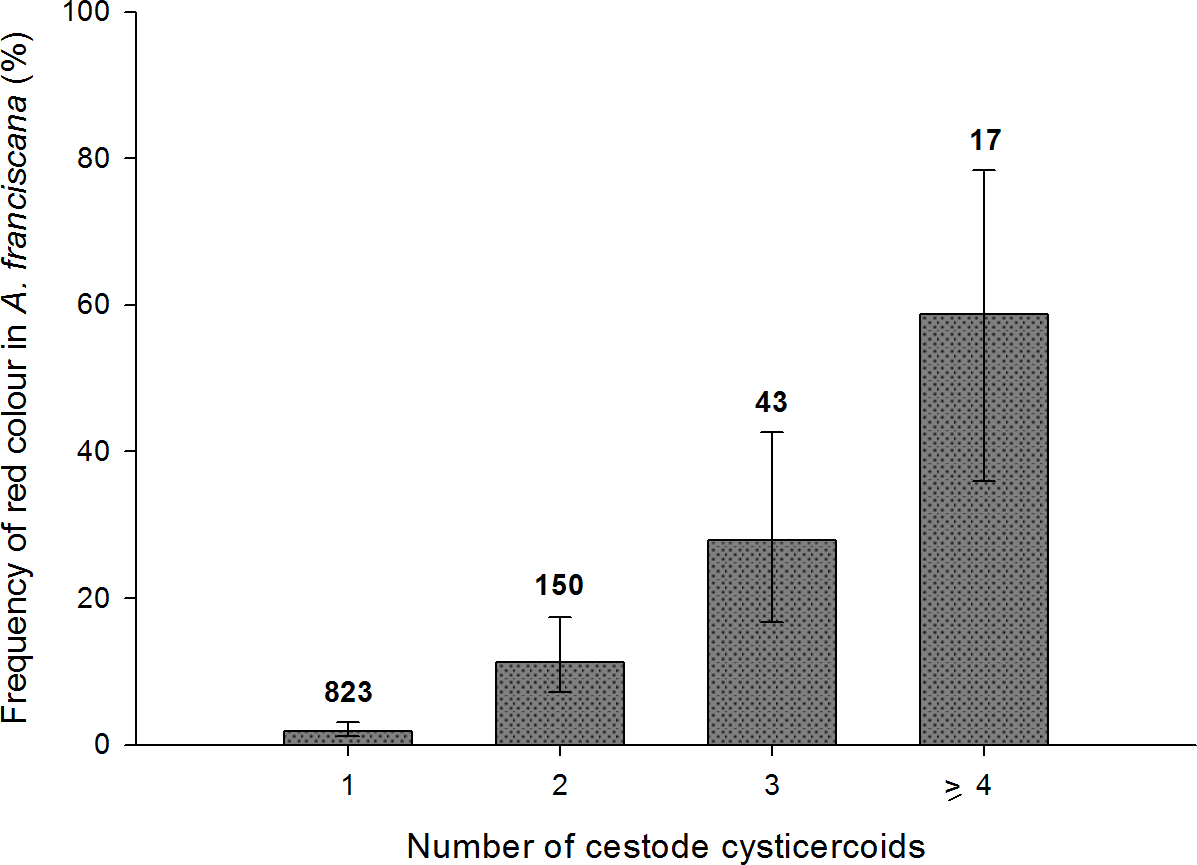{kind=link}
Where only a single species of cestode was present (single infections), there was a significant redness effect in the case of six species for A. franciscana adults, three species for A. franciscana juveniles and two species for A. salina adults (Table 4). In most of these cases, the presence of a single cysticercoid was enough to cause a significant effect (Table 4). For A. salina, no colour-effects were observed for single infections of the most abundant cestode, F. liguloides (Table 4).
Simple infection by a single A. tringae cysticercoid was significantly more likely to cause redness in adult A. salina (N = 10, of which 70% were red) than in adult A. franciscana (N = 21, with 9.5% red) (Fisher Exact test, p = 0.001). No significant differences between host species in colour-effects were detected for other cestode species, although sample sizes for A. salina were small (Table 4).
Cestodes and host castration
In A. franciscana adults, a higher proportion of infected females were castrated (i.e., with empty ovisac and no signs of functional ovaries, Fig. S3) than females which were not infected (Fig. 7 and Table 5).

Figure 7: Castration effects in adult female A. franciscana and A. salina.
Bars show 95% confidence intervals.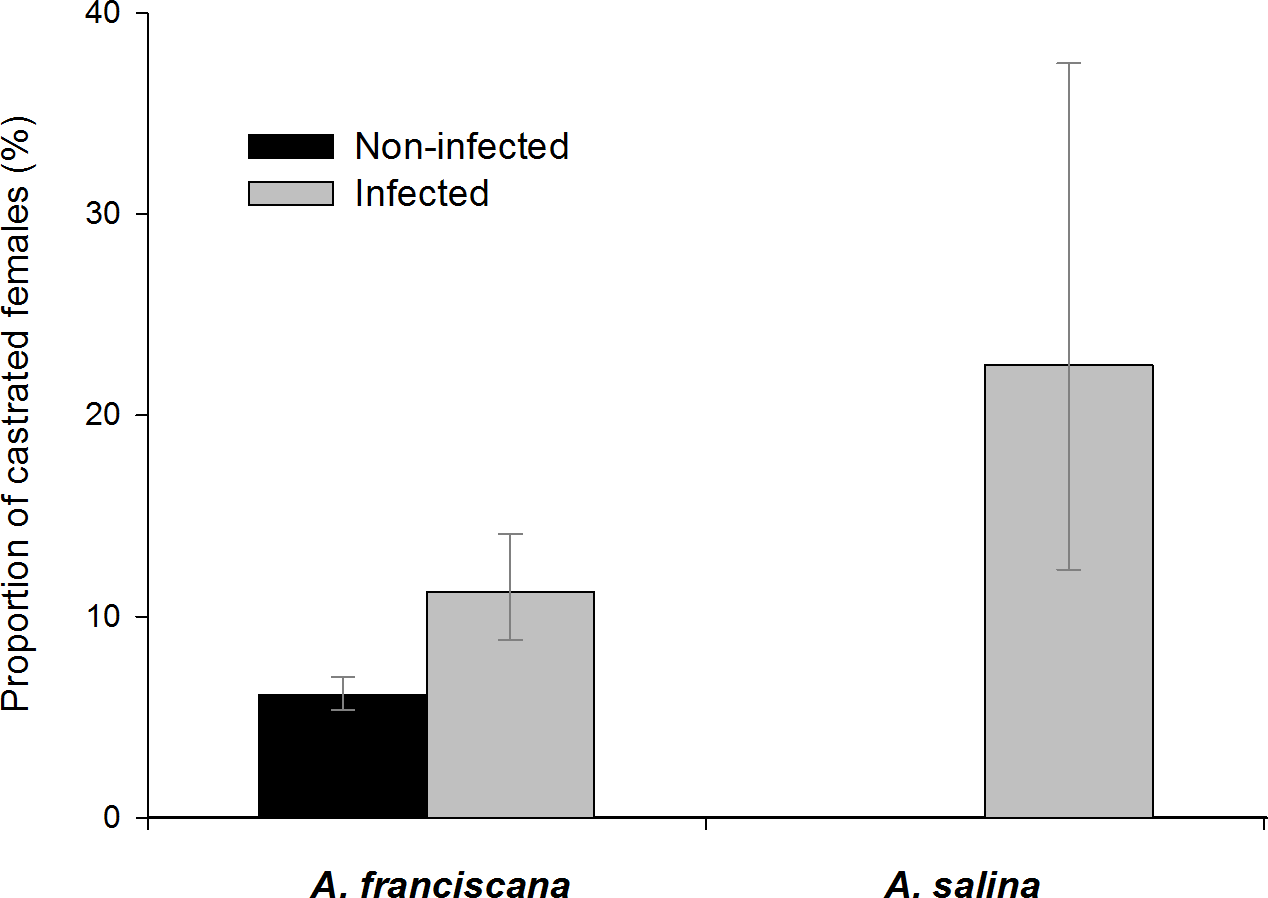{kind=link}
| A. franciscana | A. salina | |||
|---|---|---|---|---|
| N total | % castration | N total | % castration | |
| Uninfected females | 3,267 | 6.1% | 111 | 0% |
| Infected females—Total | 553 | 11.2%* | 40 | 22.5%** |
| Single infections (1 species)—Total | 496 | 8.5%* | 29 | 20.7%** |
| Infected only by FL | 32 | 25%* | 14 | 35.7%** |
| Infected only by FF | 28 | 7.1% | 1 | 0% |
| Infected only by WS | 14 | 0% | 1 | 0% |
| Infected only by EA | 362 | 7.5%* | 5 | 0% |
| Infected only by AT | 9 | 0% | 5 | 20%* |
| Infected only by AM | 22 | 4.5% | 0 | – |
| Infected only by GS | 21 | 14.3% | 0 | – |
| Infected only by GSP | 1 | 100% | 3 | 0% |
Among infected, castrated female A. franciscana, 42 (68%) were infected by only one parasite species (i.e., single infections, Table 5), and of these most were infected with E. avoceti (64.3%) or F. liguloides (19%). Castration rates in females infected only by E. avoceti or only by F. liguloides were significantly higher than those in uninfected females (Table 5). For E. avoceti, the probability of castration clearly increased with the intensity of infection, and 35.7% of females with ≥3 cysticercoids were castrated (p = 0.001). Unlike E. avoceti, infection with a single F. liguloides cysticercoid had a significant castration effect (p = 0.004). Of female A. franciscana infected by more than one cestode species (co-infections) (N = 57), 35.1% were castrated. Among these castrated females (N = 20), E. avoceti was present in 18 individuals and F. flamingo in seven.
For A. salina, castration was only recorded in infected females (Fig. 7 and Table 5). Castration effects were significant for single infections by F. liguloides or by A. tringae (Table 5). If we include females with small broods (<10 eggs in the ovisac) and no oocytes migrating in the ovaries, the proportion of female A. salina with limited fecundity when infected with F. liguloides increases from 35.7% to 57.1%.
Discussion
We present a unique study comparing the cestode infections and their consequences for two closely related branchiopod crustaceans. The native A. salina generally had more cestodes than the alien A. franciscana. When coexisting with the native, the alien had higher rates of infection than when alone. Although the cestodes were shown to have important effects on the fecundity and colour of both host species, these effects were stronger in the native species.
Comparing infections in syntopic A. franciscana and A. salina: how important is co-evolution?
Comparative studies of parasite infections in native and alien host species are essential to understand the role of parasites in biological invasions (Dunn, 2009; Kelly et al., 2009; Dunn et al., 2012). The present work sheds light on the role of endemic cestodes in competitive interactions between native and alien sexual Artemia. Cestode parasitism differed clearly between invasive and native brine shrimps under the same environmental conditions and at the same time. Cestode species richness and the prevalence of several species were lower in A. franciscana, and the difference was especially striking for F. liguloides (Figs. 3 and 4). These results are consistent with studies of populations in the southern Iberian Peninsula which have found A. franciscana to have low infection rates throughout the annual cycle. In summer, Georgiev et al. (2007) found lower levels of cestode infections in three A. franciscana populations than in an allopatric A. salina population. Studies throughout the annual cycle of A. franciscana in Cadiz Bay and of A. salina in Almería also found that the invasive species had fewer infections (Sánchez et al., 2013; Georgiev et al., 2014). Our results for syntopic populations indicate that these earlier results are not explained by other differences between these allopatric populations (e.g., in environmental conditions, or in the density of birds). A previous study in syntopic populations in Aigües-Mortes in France showed that A. franciscana had fewer cestodes than native, clonal A. parthenogenetica (Sánchez et al., 2012), although this might be related to the advantages of sex in resisting parasites. In any case, the present study provides strong evidence that the introduced A. franciscana is a less susceptible host for cestodes than the native sexual A. salina.
The lower burden of parasites in introduced hosts suggests A. franciscana is resistant to several native cestode species such as F. liguloides. Different capacities to infect the new host may be the result of varying host–parasite co-evolutionary history. The lower infection of F. liguloides in A. franciscana may be explained by a strong immune response of the host against this parasite, given the restricted distribution of flamingos in North America which barely overlaps with the natural range of A. franciscana (Muñoz et al., 2013). There are no flamingos in the western USA, from where A. franciscana has been exported around the world. Observations of dead cysticercoids of F. liguloides in A. franciscana support a strong immune response in this host (Georgiev et al., 2014). In contrast, the high prevalence of E. avoceti recorded in A. franciscana in our study may reflect a shared coevolutionary history, since E. avoceti occurs in shorebirds in North America (Clark, 1954). A. franciscana does not appear to be especially resistant to cestodes in its native range, since the total prevalence of cestodes in Great Salt Lake, USA was much higher than in the Ebro Delta and other parts of the invasive range (Redón et al., 2015). Our results are consistent with previous studies that have found parasites of native hosts to have no or a limited capacity to spread to non-indigenous congeners (Dunn & Dick, 1998; Torchin, Byers & Todd, 2005; Genner, Michel & Todd, 2008).
How does coexistence of both species influence infection parameters in Artemia franciscana?
When a non-indigenous species arrives in a new range, it can affect native communities through different processes: “spillover” of introduced parasites, “spillback” of native parasites, and “dilution” of parasitism (Kelly et al., 2009). A. franciscana was introduced into Europe as imported cysts (dormant eggs) free of native cestode parasites, so “spillover” is not expected in this system. The low levels of parasitism recorded in A. franciscana indicate that it is a poor or incompetent reservoir for native cestodes, making “spillback” to native Artemia unlikely. We might expect a parasite dilution effect of the invader to the benefit of the native host which should be subjected to lower disease transmission when mixed with incompetent, alien hosts (Keesing, Holt & Ostfeld, 2006; Hall et al., 2009; Johnson & Thieltges, 2010). A reduced infection rate by native trematodes was demonstrated experimentally in native freshwater snails Potamopyrgus antipodarum exposed together with the invader Lymnaea stagnalis (Kopp & Jokela, 2007). Given the strong effects of cestodes on A. salina, such a “dilution effect” could increase the possibility of local persistence of A. salina in the presence of the invader (Hatcher, Dick & Dunn, 2006).
Because our study area did not include ponds holding only A. salina, we cannot reliably assess the strength of any dilution effect on this species. However, the levels of cestode prevalence we recorded in the presence of A. franciscana were not unusually low compared to studies of A. salina elsewhere in the absence of the invader (Table S1). Likewise, Sánchez et al. (2012) found no evidence of a dilution effect on A. parthenogenetica when coexisting with A. franciscana, since the total prevalence in A. parthenogenetica was high (c. 70%).
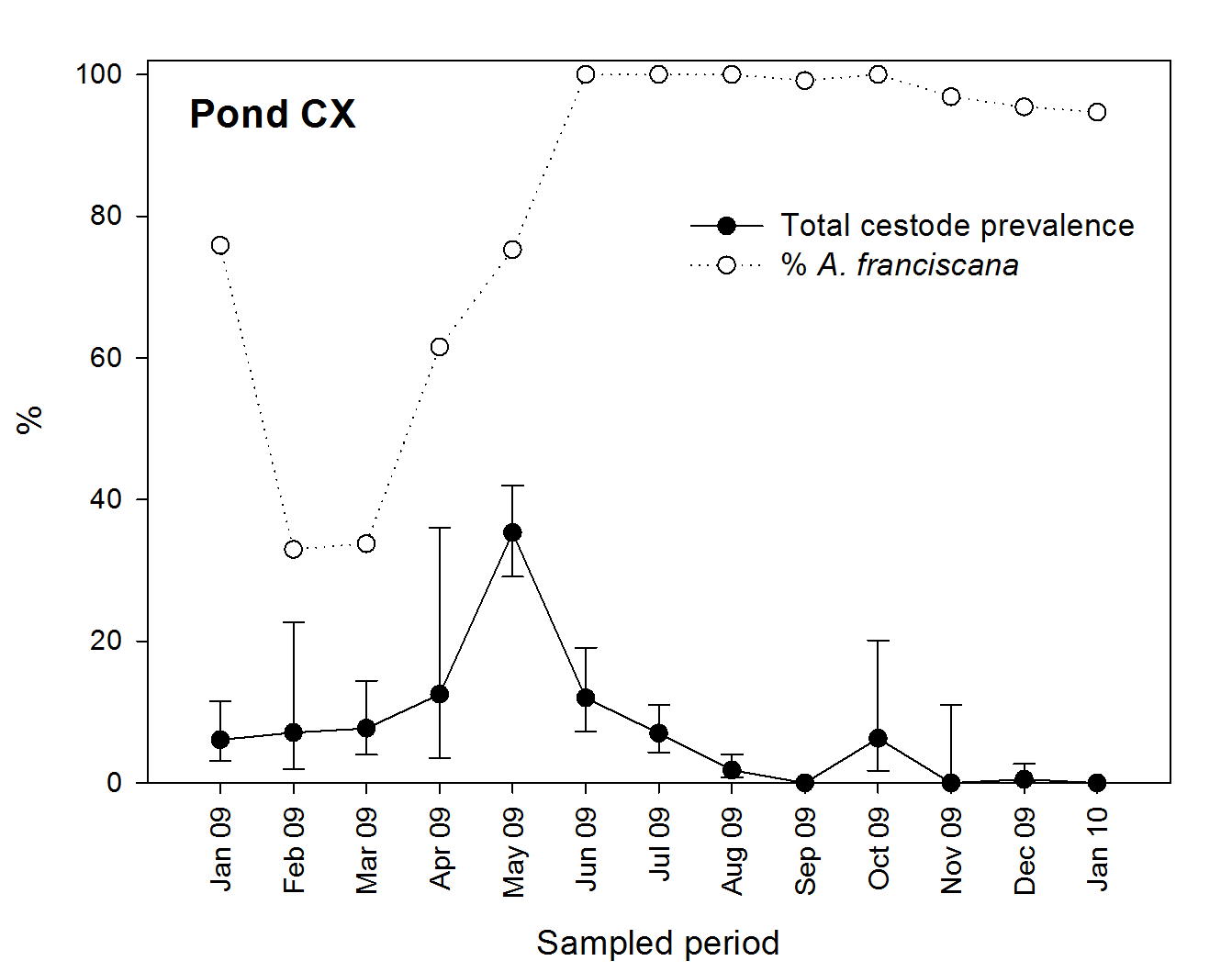
Our study allowed a good test of the “dilution effect” in the other direction, i.e., whether the native host reduced parasitism in the alien host. We found the opposite (an “amplification effect”), with higher infection in A. franciscana when coexisting with A. salina. Indeed, the temporal dynamics of infection in A. franciscana in pond CX (with both Artemia species) seem to depend on the proportion of native hosts in the Artemia population. Thus, total prevalence increased from January to May 2009 (coinciding with the abundance of A. salina), then declined from June to December when A. franciscana was the dominant species (Fig. S4). In contrast, in pond 4 (without A. salina), higher prevalences were recorded in July and August during 2007 and 2008 (S Redón, AJ Green, BB Georgiev, GP Vasileva, F Amat, 2015, unpublished data). Since A. franciscana is a poor reservoir (i.e., the cestodes may not circulate effectively in them), adding A. salina to the community might increase the prevalence of the infection in the alien host because the added host is a better reservoir (Keesing, Holt & Ostfeld, 2006; Hatcher, Dick & Dunn, 2006). Since Artemia are only intermediate hosts, such an explanation would require final avian hosts to be relatively faithful to individual ponds, so that birds that become infected in the pond with A. salina are more likely to release cestode eggs there than in other ponds. Another possible and not mutually exclusive explanation for our results is that the final avian hosts are more attracted to ponds holding A. salina because this is their preferred prey, leading to greater parasite circulation in these ponds. Owing to weak manipulation by native parasites, A. franciscana is much less abundant at the water surface where they are accessible to birds, and as shown in our study, they are less likely to have the red colour preferred by birds (Sánchez et al., 2009b). In turn, this suggests that the generally low infection rates in A. franciscana may be accounted for not only by a greater resistance to parasites than native species, but also by a tendency for final hosts (birds) to avoid invaded salt ponds, reducing levels of parasite circulation. A study of how waterbird use differs between salt ponds with native or alien shrimp is required.
Castration and colour effects in native and alien hosts
In many trophically transmitted parasites, larval parasite stages alter host characteristics such as longevity, size, colour or behaviour to increase the risk of predation by the final host (Moore, 2002; Thomas, Adamo & Moore, 2005; Médoc, Bollache & Beisel, 2006). Parasitic castrators benefit by diverting host resources away from reproduction, allowing faster growth and earlier arrival at the infective stage, or an increase in host lifespan, thus increasing opportunities for transmission (Hurd, Warr & Polwart, 2001). A. parthenogenetica infected with F. liguloides live longer than uninfected ones (Amat et al., 1991), and this increased host longevity may ensure the complete development of the infective stage from oncosphere to ripe cysticercoid (see Redón et al., 2011), ready to be transmitted to final hosts.
We found significant but mild castrating effects of cestodes in A. franciscana, and much stronger effects in native A. salina. Despite the smaller sample size, for A. salina we could confirm a castrating effect for A. tringae and F. liguloides. The latter species was already known to have a strong castrating effect on A. salina (Amarouayache, Derbal & Kara, 2009) and A. parthenogenetica (Amat et al., 1991; Sánchez et al., 2012). In A. franciscana, we found significant castration effects for F. liguloides and E. avoceti, with a stronger effect by the former (a single cysticercoid of F. liguloides is able to castrate, whereas two or more are needed for E. avoceti). This is the first time that a castration effect has been demonstrated for dilepidid cestodes in Artemia, and suggests these effects are widespread and not limited to F. liguloides. Our results have important implications for the use of A. franciscana in aquaculture, since they show that cestode infections reduce host fecundity, even in the introduced range. This suggests that cyst production in sites within the introduced range where cysts are harvested for aquaculture (e.g., Bohai Bay in China) is likely to be reduced by cestode infections.
This is the first study of the colour-effects of cestodes in A. franciscana in the invasive range, and the first to consider the effects in A. salina in detail. The association of red colouration with cestode infection is well known for native A. parthenogenetica, especially for F. liguloides, and appears to be due to parasite manipulation associated with the preference that final hosts have for predating red Artemia (Sánchez et al., 2009b; Sánchez et al., 2012). However, it is also possible that this altered pigmentation is caused by a host immune response against infection (e.g., given the antioxidant function and immunostimulating properties of carotenoids) rather than a specific parasitic strategy to ensure transmission. The red colour of cestode-infected A. parthenogenetica is partly due to carotenoids (Amat et al., 1991; Sánchez et al., 2006) and carotenoids seem to play an important role in immune defence by providing free radical scavengers against cestode infection (Van der Veen, 2005). Cornet, Biard & Moret (2007) found a positive relationship between carotenoid concentration in haemolyymph and immune parameters (those evolved in the prophenoloxidase cascade) in the crustacean Gammarus pulex, suggesting that carotenoids can help to reduce the costs of immunity by limiting self-harming. A positive relationship between carotenoid concentration and the abundance of cestodes (but not of nematodes) was also observed in greylag geese Anser anser (Figuerola et al., 2005), suggesting there may be a similar host response to cestode infection across a broad taxonomic range.
The proportion of infected shrimps with a red colour was higher for A. salina than for A. franciscana, whether considering co-infections or only single infections. This further suggests that the invasive species has greater resistance to cestode infections. Red colouration in A. salina was significantly associated with infections by A. tringae and E. avoceti, and the redness effect of A. tringae was stronger than in A. franciscana. Surprisingly, we could not detect any effect of F. liguloides on A. salina colour, suggesting that this native sexual host is more resistent to this parasite than the asexual A. parthenogenetica, as predicted by the Red Queen hypothesis. However, red colouration may be of less value to increase transmission to the filter-feeding flamingos that are final hosts of F. liguloides, than to shorebirds which are visual predators and final hosts of E. avoceti and Anomotaenia spp.
Red pigmentation in A. franciscana is linked with cestode infection, particularly with dilepidids (E. avoceti, A. tringae, and A. microphallos), and to a lesser extent with flamingo parasites (F. liguloides, F. flamingo and G. stammeri), a finding in line with the higher prevalence and castration effects we recorded for dilepidids. There was a strong positive correlation between red-colour and overall infection-intensity, as previously observed for A. parthenogenetica (Sánchez et al., 2006). Ours is the first study to compare redness effects separately in juvenile and adult Artemia. For A. franciscana juveniles, we found a redness effect for three cestode species, indicating that cestodes manipulate host colour even in early stages of host development. There is a need for detailed research into the nature and function of the pigments causing red colouration in different Artemia species and life stages parasitized by different cestodes.
Previous studies in other host–parasite systems have also found native parasites to have less pathological effects in alien hosts than in native hosts (Bauer et al., 2000; Cornet, Sorci & Moret, 2010). For example, in an acantocephalan-gammarid system, Cornet, Sorci & Moret (2010) found that a local parasite Pomphorhynchus laevis had less ability to infect and induce pathogenic effects in invasive than in native gammarids.
Consequences of the invasion of A. franciscana for native parasites
Our results demonstrate that A. franciscana has accumulated novel parasites following its introduction. However, it is not clear that all the cestodes parasitizing native Artemia in the Mediterranean region will be able to persist if all the native populations are replaced by the American species. Coextinctions (i.e., the loss of one especies as a result of the extinction of the species it depends on; Dunn et al., 2009) seem likely in our host–parasite system, especially at a local scale of individual ponds. Cysticercoids have to be able to establish, survive and grow until reaching an infective-stage in brine shrimps, and then reach avian final hosts to complete the parasite’s cycle. It is possible that some cestodes will not be able to infect this new host at a high enough rate or to reach their final hosts with a sufficiently high frequency (according to the very low prevalences and the weaker manipulation observed) to ensure the survival of a viable metapopulation. When introduced species become hosts for native parasites it does not necessarily mean that they play a major role in the life cycles and transmission of these parasites. The reduced levels of infection and reduced castration effects in A. franciscana suggest that cestodes may not be able to infect or manipulate the alien host to a sufficient extent to ensure viability. The reduced redness suggests that cestodes are less likely to reach final hosts (through bird predation) when infecting alien Artemia, and in turn that the value of salterns for waterbird conservation may be reduced by the invasion. Parasites are an important component of food webs (Lafferty, Dobson & Kuris, 2006; Lafferty et al., 2008) and ecosystem functioning (Hudson, Dobson & Lafferty, 2006), so loss of cestodes may have an impact on the stability of hypersaline ecosystems.
Conclusions
Cestodes have a greater impact on native A. salina than on invasive A. franciscana in several ways, suggesting that native cestodes are poorly adapted to these novel hosts. They cause infections of higher prevalence and species richness in the native host. For a given infection intensity, they cause a greater impact on host fecundity, and a greater change in colouration, likely to translate into a higher predation rate by birds. Therefore, cestodes can influence competition between Artemia species and are likely to help A. franciscana to outcompete native species in the Mediterranean region and other parts of the world. In turn, the viability of these cestode populations across broad scales, where waterbird populations interact with different hypersaline ecosystems, is threatened by the loss of native Artemia which may act as a “source” for cestodes whereas A. franciscana populations may act as a “sink”.
Supplemental Information
Proportion of juvenile brine shrimps in each sample examined
(A) pond CX, (B) pond 4. Proportions are shown for both Artemia species when present.
{kind=link}
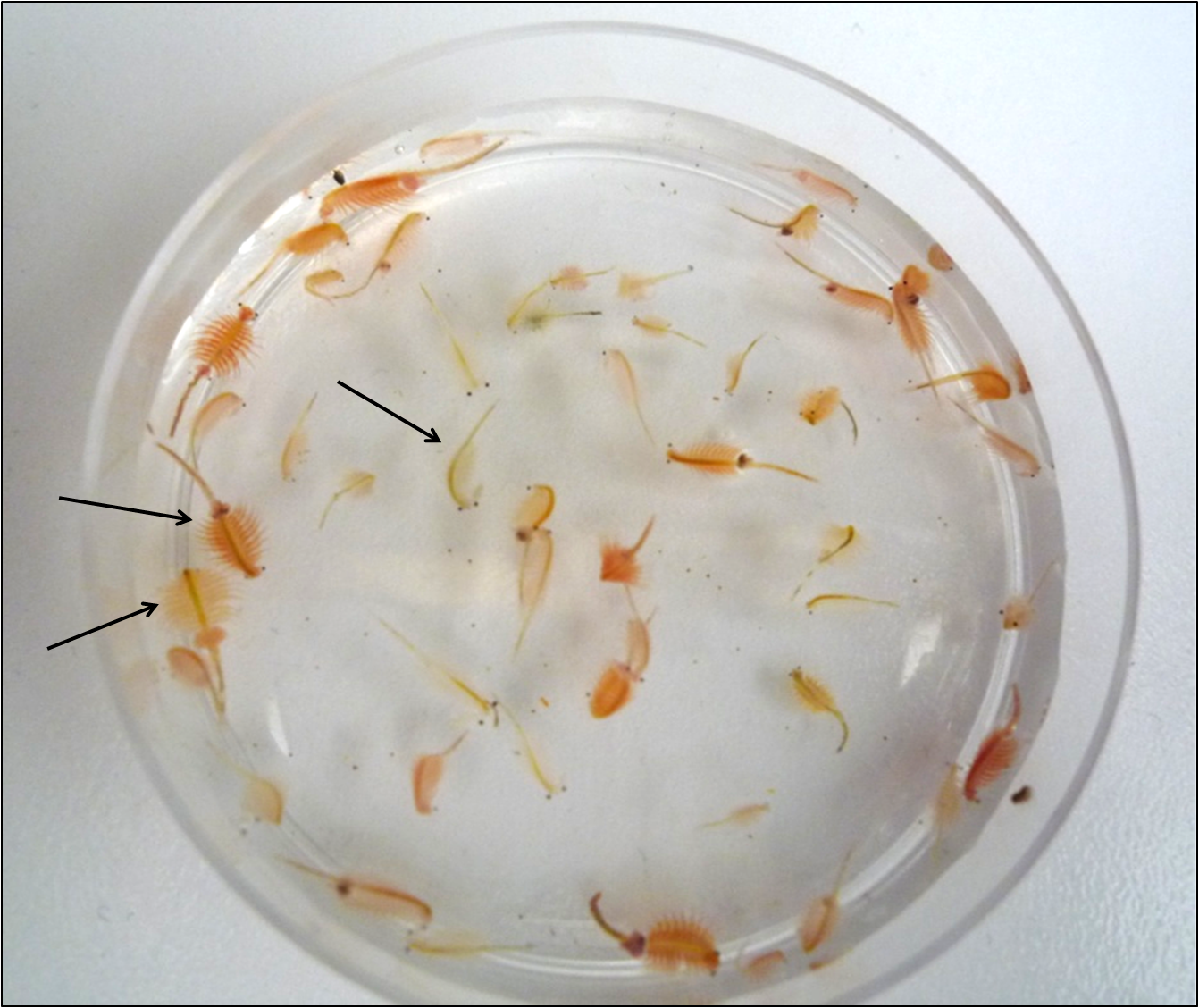
Brine shrimp Artemia individuals of different colouration
Examples highlighted with arrows from left to right are: light-red, dark-red, and not-red.
{kind=link}
General aspect of adult brine shrimp females with different reproductive status
(A) ovigerous, (B) castrated. Females are infected by larval cestodes (indicated by arrows).
{kind=link}
Temporal dynamics of cestode infection in adult A. franciscana from pond CX and proportion of this species in the community, through an annual cycle (January 2009–January 2010)
Bars show 95% confidence intervals.
{kind=link}
Comparative data on total cestode prevalence in relation to host community diversity.
Data are presented for the native A. salina (AS) and the invader A. franciscana (AF) when coexisting (syntopic-populations, present study) and from populations without co-ocurrence of other congeneric species or those with small numbers of diploid A. parthenogenetica (PD). When many data are available (e.g., for several months, years), the mean value of total prevalence and the range (in parentheses) are given. * mean prevalence of cestode infection was recalculated excluding individuals infected with nematodes recorded in September 2007.
Data for parasite castration effects
Raw data used for the analyses of parasite castration effects in adult females of both Artemia spp. including all samples available from Ebro delta salterns.
Data for parasite colouration effects
Raw data used for the analyses of parasite colour effects in both Artemia spp. including all samples available from Ebro delta salterns.
Data from pond CX
Raw data for quantitative analyses of infections between alien and native Artemia from pond CX during months of coexistence (January to May 2009).
Data A. franciscana populations
Raw data for comparative infections between A. franciscana from syntopic (pond CX) and single (pond 4) populations.