
Exploring the relationship of Homalosilpha and Mimosilpha (Blattodea, Blattidae, Blattinae) from a morphological and molecular perspective, including a description of four new species
- Published
- Accepted
- Received
- Academic Editor
- Tony Robillard
- Subject Areas
- Biodiversity, Entomology, Molecular Biology, Taxonomy, Zoology
- Keywords
- Bayesian inference, Habitat, Key, Maximum likelihood, Sister group
- Copyright
- © 2021 Liao et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. Exploring the relationship of Homalosilpha and Mimosilpha (Blattodea, Blattidae, Blattinae) from a morphological and molecular perspective, including a description of four new species. PeerJ 9:e10618 https://doi.org/10.7717/peerj.10618
Abstract
This study utilized six genes (12S, 16S, 18S, 28S, COII and H3) from a total of 40 samples to construct maximum likelihood (ML) and Bayesian inference (BI) phylogenetic trees in order to infer the relationships between the morphologically similar genera Homalosilpha Stål, 1874 and Mimosilpha Bey-Bienko, 1957. The phylogenetic analysis showed the two genera have a close relationship and were recovered as sister groups based on ML and BI analyses. Four new species are described among these samples, i.e., Homalosilpha obtusangula sp. nov., Homalosilpha recta sp. nov., Homalosilpha alba sp. nov. and Homalosilpha clavellata sp. nov. based on morphological and COI data. A key to the worldwide Homalosilpha is provided.
Introduction
Genus Homalosilpha Stål, 1874 is remarkable in the family of Blattidae for its flat pronotum decorated with black or white markings (Princis, 1966a; Roth, 1999). Stål (1874) established Homalosilpha with Periplaneta ustulata Burmeister, 1838 as the type species. Then Kirby (1904) transferred three Periplaneta Burmeister, 1838 species to Homalosilpha. Homalosilpha can be easily distinguished from Periplaneta by the discoidal pronotum whose surface is scattered with various spots, sides are not deflexed and with greatest width at the middle (Shelford, 1910). Subsequently, two species were reported from Uganda and China by Shelford (1908), Shelford (1910). Later, Princis (1966a) described two species from Congo and Indonesia and provided a key for eight Homalosilpha species. In 1969, Bey-Bienko described four species, of which three species were from China. Kumar (1975) synonymized H. vicina Brunner von Wattenwyl, 1865 with H. cruralis Shelford, 1908 because the diagnostic characters used, the color of the tibiae and the anterior and posterior margin of pronotum, were variable and should be treated as intraspecific variation. After the examination of specimens from Malaysia and Indonesia by Roth (1999), Homalosilpha quadrimaculata was described on the basis of the distinct orange spots on the pronotum. Up to now, a total of twelve species of Homalosilpha had been reported worldwide (Beccaloni, 2014), of which, H. arcifera Bey-Bienko, 1969, H. gaudens Shelford, 1910, H. kryzhanovskii Bey-Bienko, 1969, H. ustulata (Burmeister, 1838), and H. valida Bey-Bienko, 1969 are distributed in China. Without professional taxonomic knowledge about Blattodea, it is challenging to identify the Homalosilpha species owing to their high similarity in appearance (Liao SR & Che YL, pers. obs., 2018–2019). Therefore, simple and accurate methods are needed to help the identification of Homalosilpha species.
Genus Mimosilpha was established with M. disticha as type species from Yunnan, China. It is strongly similar to Homalosilpha in appearance (Bey-Bienko, 1957). The main difference between Mimosilpha and Homalosilpha is that there are three rows of spines along the edge of the hind tibiae of Homalosilpha, while Mimosilpha only has two rows of spines. Only one Mimosilpha species has been recorded in the world (Beccaloni, 2014).
DNA barcodes proved to be an effective tool to aid the identification of the similar species and even resolve the problem of sexual dimorphism in cockroaches (Evangelista, Buss & Ware, 2013; Che et al., 2017; Liao, Wang & Che, 2019). Multi-gene combination phylogenetic trees are increasingly applied in cockroach systematics to explore the problem of paraphyly (Inward, Beccaloni & Eggleton, 2007; Ware et al., 2008; Legendre et al., 2015), establish new taxa (Evangelista et al., 2019), infer possible sister group (Djernæs, Klass & Eggleton, 2015; Wang et al., 2017), and then revise the taxonomy (Djernæs, 2018). These studies have all been demonstrated to be informative and successful in revealing relationships among different groups via multi-gene analysis. Mimosilpha and Homalosilpha are very close in morphology with a confused taxonomy, so it is urgent to sequence six genes to infer the relationship of these two genera.
In this study, 13 COI sequences of Homalosilpha and Mimosilpha species were obtained in order to help distinguish species when combined with morphological data. We also sequenced three mitochondrial genes (12S, 16S and COII) and three nuclear genes (18S, 28S and H3) to explore the phylogenetic relationship of Homalosilpha and Mimosilpha. Moreover, we illustrate four new Homalosilpha species from China based on morphological characters and DNA barcoding.
Materials and Methods
Morphological study
Morphological terminology used in this paper mainly follows McKittrick (1964), Roth (2003) and Li et al. (2018).
The maculae on the pronotum provided effective information for the identification of Homalosilpha species, which could be grouped into three types (Shelford, 1910; Princis, 1966a; Bey-Bienko, 1969; Roth, 1999): (1) white spots scattered on the black pronotum, represented by H. decorata and H. quadrimaculata, (2) one large central dark spot on the pale brown pronotum, represented by H. hanni, H. nigricans and H. gaudens, and (3) the symmetrical multiple spots and stripes on the disk of the pale brown pronotum, represented by the remaining Homalosilpha species. The third type is more complex than the former two, so we herein address the specific part of the maculae for description as shown in Fig. 1.
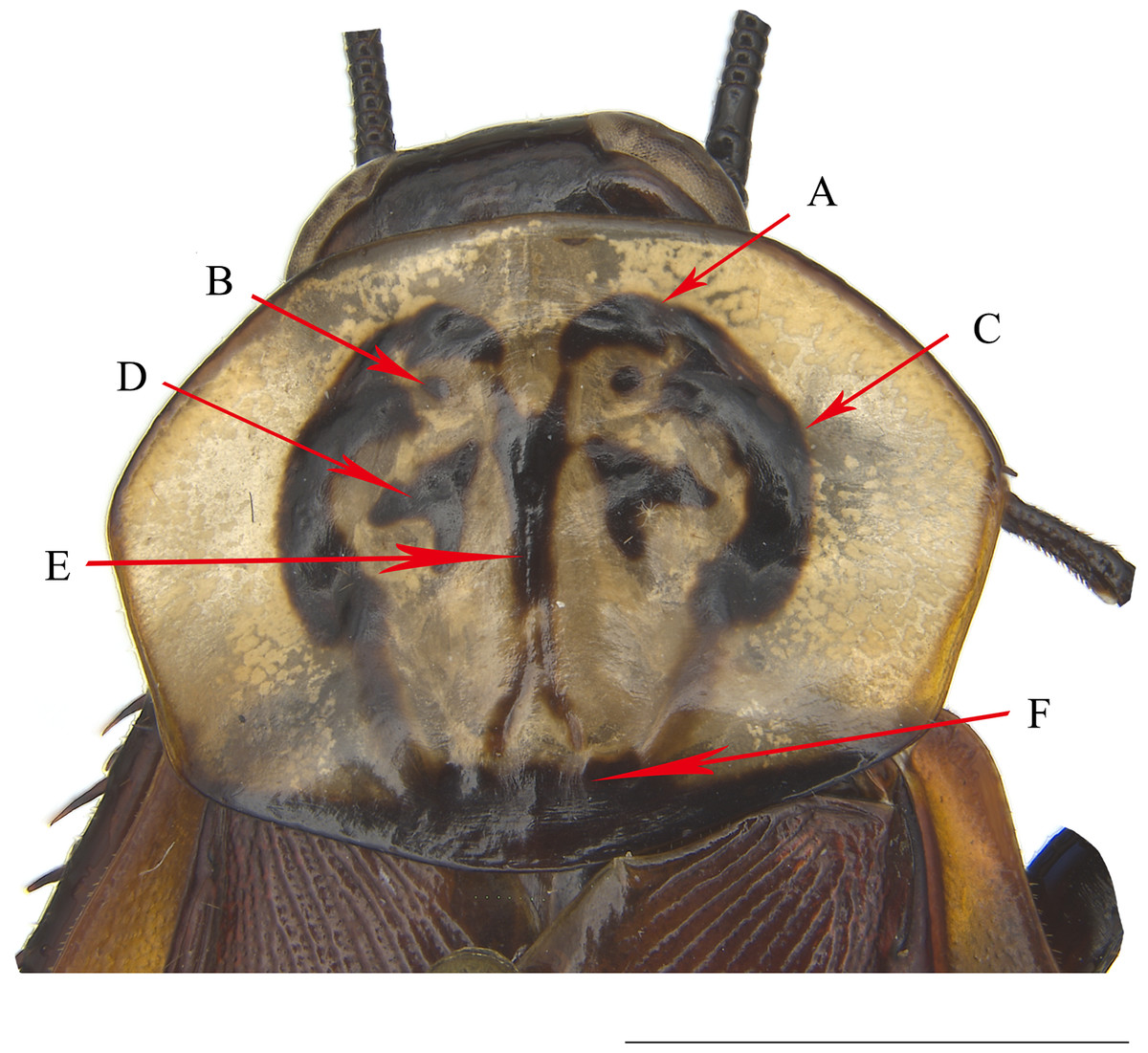
Figure 1: The pronotum of H. obtusangula sp. nov.
(A) Anterior pattern. (B) Small spot. (C) Lateral pattern. (D) Central pattern. (E) Vertical pattern. (F) Posterior band. Scale bars = 5 mm.{kind=link}
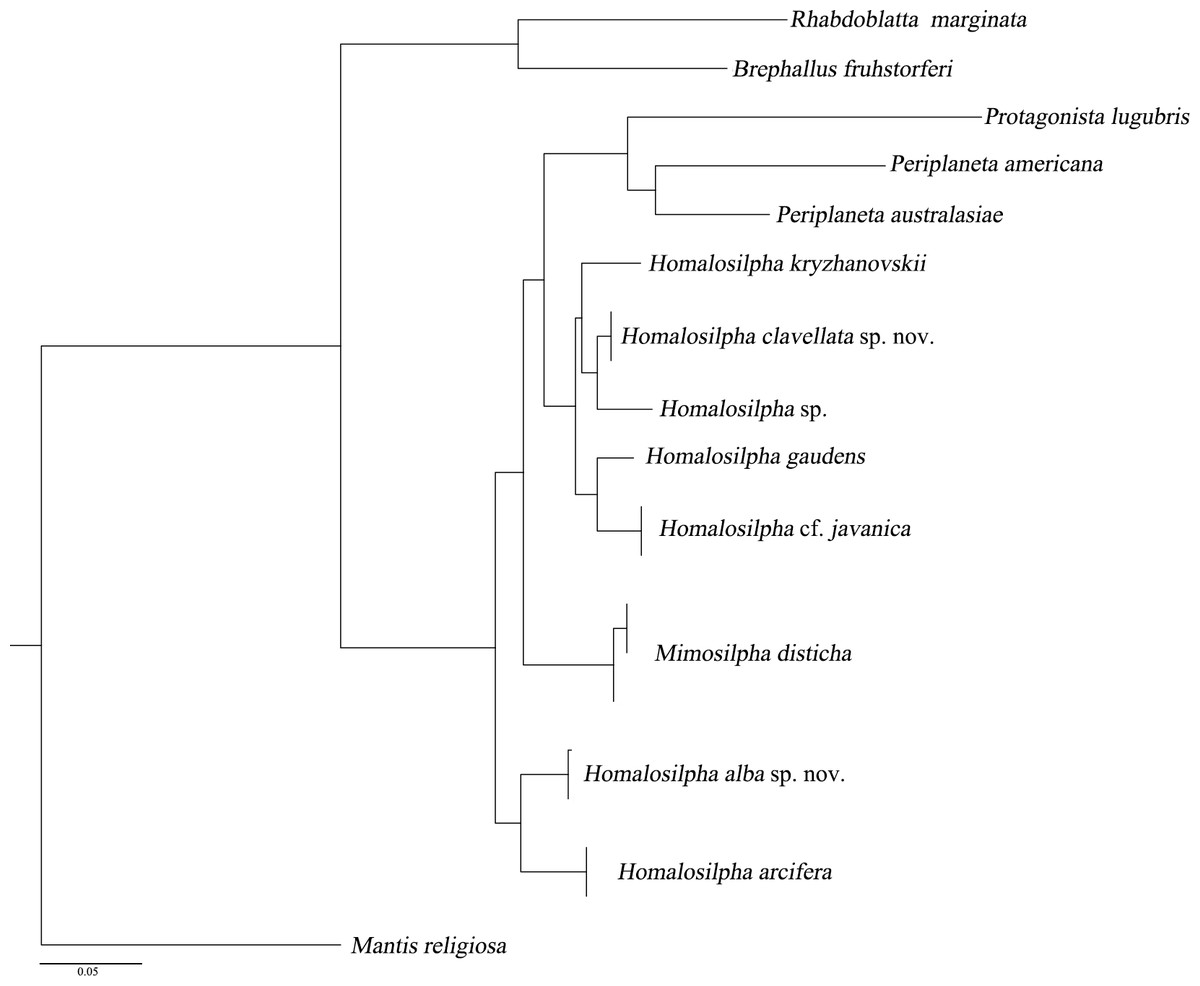
Figure 2: Maximum likelihood (ML) tree of the cockroaches based on COI.
ML tree derived from COI gene analysis following GTRGAMMA model with 1,000 bootstrap replicates.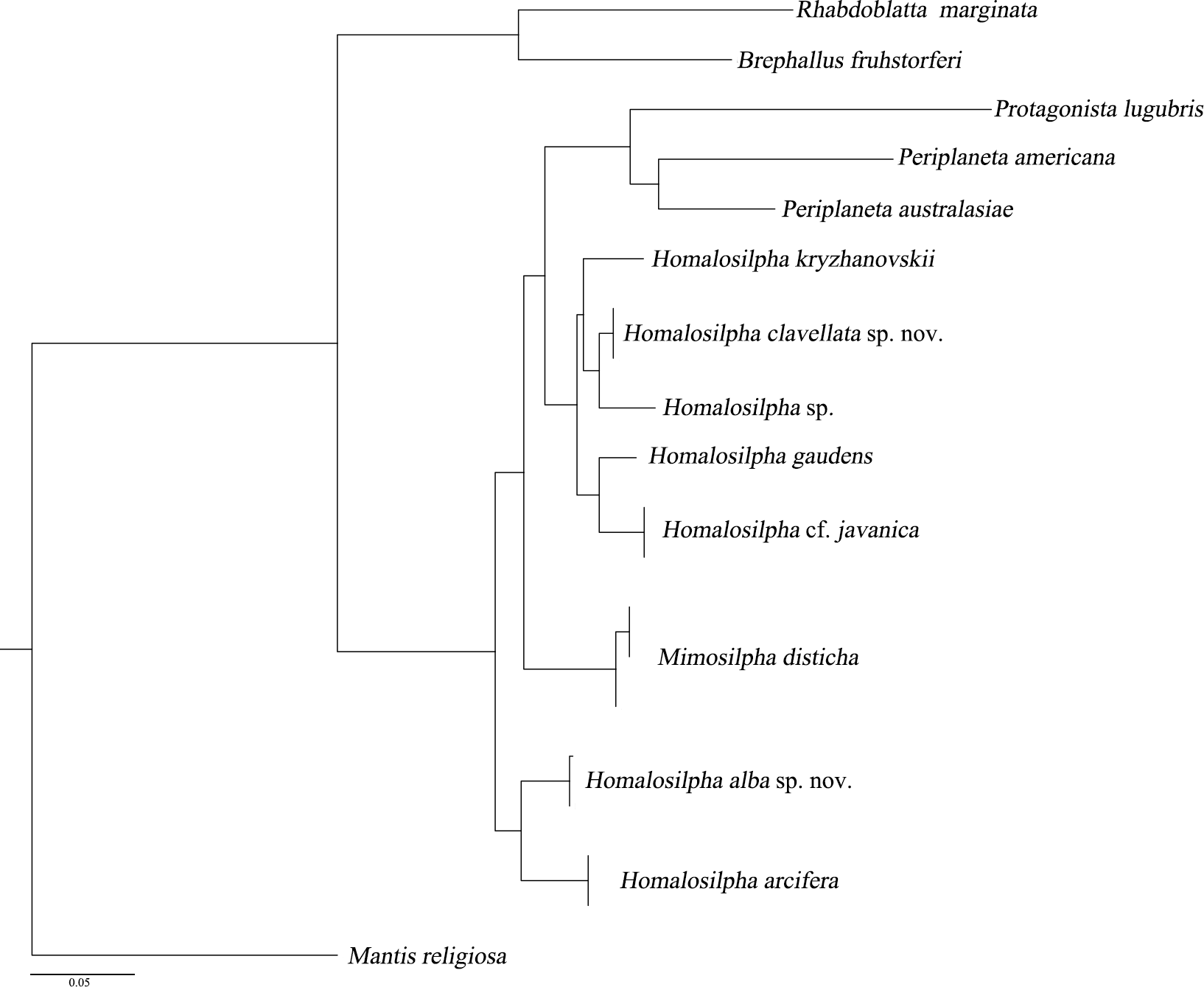{kind=link}
All specimens were measured by vernier caliper for the body length from the anterior to the posterior, for the body length including tegmina from the anterior to the tip of tegmina, and pronotum length × width at the longest and the widest part. Genital segments of the examined specimens were macerated and photographs (Figs. 1–9) of the specimens were taken as previously described in Liao, Wang & Che (2019). The type materials are deposited in the Institute of Entomology, College of Plant Protection, Southwest University, Chongqing, China (SWU).
DNA extraction, PCR, and sequencing
We sampled seven genes of 13 species (Tables S1 and S2) from Homalosilpha in this study: mitochondrial 12S, 16S, cytochrome c oxidase subunit I (COI) and subunit II (COII), and nuclear 18S, 28S, and histone H3. Total DNA was extracted from the leg tissue of samples according to the Hipure Tissue DNA Mini Kit (Tsingke Biological Technology, Beijing, China). All fragments were amplified using PCR; primers for amplifications are given in Table 1. Reactions were carried out in volumes of 25 µL, containing 22 µL of 1 × 1 T3 supper mix (Tsingke Biological Technology, Beijing, China), 1 µL of each primer and 1 µL of DNA template, except for COII containing 12.5 µL T2 mix (Tsingke Biological Technology, Beijing, China), 8.5 µL of ultrapure water, 1 µL of each primer and 2 µL of DNA template. The amplification conditions were: initial denaturation at 98 °C for 2 min, followed by 35 cycles for 10 s at 98 °C, 10 s at 43−55 °C, and 15 s at 72 °C, with a final extension of 2 min at 72 °C; however for COII: initial denaturation at 94 °C for 5 min, followed by 35 cycles for 45 s at 94 °C, 45 s at 50 °C, and 45 s at 72 °C, with a final extension of 10 min at 72 °C. All sequences were deposited in GenBank (accession numbers in Tables S1 and S2).
Sequence processing and phylogenetic analyses
In this study, a total of 14 COI sequences, whose lengths were 658 bp, were combined with one Protagonista, two Periplaneta, one Rhabdoblatta, one Brephallus and one mantid sequence to infer species delimitation analysis for Homalosilpha and Mimosilpha (Table S1). Intraspecific and interspecific genetic divergence values are quantified based on the Kimura 2-parameter (K2P) distance model (Kimura, 1980), using MEGA 7 (Kumar, Stecher & Tamura, 2016).
To infer the relationship of Homalosilpha and Mimosilpha, we included sequence data from 22 Blattidae taxa (ingroup including seven Homalosilpha and one Mimosilpha species) and 12 outgroup taxa (Table S2). These analyses were performed based on six genes (12S, 16S, 18S, 28S, COII and H3). These data was aligned by online Mafft 7 (https://mafft.cbrc.jp/alignment/server/), and the methods were the same as Wang et al. (2017). Specifically, the Q-INS-i algorithm was selected for non-coding protein genes (12S, 16S, 18S, 28S), the G-INS-i algorithm was selected for coding protein genes (COII, H3) and used with other parameters at their default values. Alignments of sequences were inspected visually and manually adjusted in MEGA 7.0. Poorly aligned characters were removed but these were limited. The remaining bases are as following: 424nt for 12S, 447nt for 16S, 1814nt for 18S, 596nt for 28S, 665nt for COII, 328nt for H3.
The molecular data set was divided into eight partitions (partitioned by gene: 12S, 16S, 18S, 28S, COII_pos1, COII_pos2, H3_pos1, and H3 _pos2). We excluded the third codon positions of the protein-coding genes (COII and H3) because of the high level of mutational saturation. The third codon position (pos3) (ISS = 0.977) was much more saturated than the first and second codon position (pos12) (ISS = 0.493) after using Xia’s method implemented in DAMBE 5.0 (Xia, 2013). Phylogenetic analyses were constructed using maximum likelihood (ML) and Bayesian inference methods. ML analyses were performed using RaxML 7.7.1 (Stamatakis, Hoover & Rougemont, 2008), and BI analyses were performed using MrBayes 3.2 (Ronquist et al., 2012). ML analyses was performed using RAxML v.7.7.1 (Stamatakis, Hoover & Rougemont, 2008). We used the GTRGAMMA model for the combined datasets with 1,000 bootstrap replicates. For BI analyses, PartitionFinder v.1.1.1 (Lanfear et al., 2012) was utilized to choose models and model selection was based on BIC. For eight partitions, the best-fitting substitution models are as follows: GTR+I+G: 12S, 16S; TIMef+I+G: 18S, H3_pos1, H3_pos 2; TrN+I+G: 28S, COII_pos1, COII_pos2. Two independent sets of Markov chains were run, each with one cold and three heated chains for 5,000,000 generations, and every 1,000th generation was sampled. Convergence was inferred when a standard deviation of split frequencies < 0.01 was completed. The ESS values are all more than 200.
Results
Species delimitation based onCOIand morphological data
In this study, we acquired 14 COI sequences representing seven Homalosilpha and 1 Mimosilpha species. All of the new sequences have been deposited in GenBank with accession numbers MW201581–MW201594 (Table S1). The COI sequences we acquired had a relatively high AT content (67.2%), with an average nucleotide composition of A = 31%, T = 36.2%, C = 16.5%, and G = 16.3%. Sequence analysis revealed that 109 (16.57%) sites were variable, of which 79 (12%) sites were parsimoniously informative. ML analysis revealed that clades from the same species, constituted monophyletic groups with high support values (Fig. 2). In particular, the male and female of H. clavellata sp. nov., which have different markings on the pronotum, could be matched by COI data.
We observed the lowest and largest K2P interspecies genetic distance (Table 2) among these species. The lowest distance 0.031 was between Homalosilpha sp. and H. clavellata sp. nov., the largest distance 0.084 between H. nigricans and H. arcifera, H. arcifera and H. kryzhanovskii, as well as M. disticha and H. kryzhanovskii. Morphologically, although having a low genetic distance, Homalosilpha sp. and H. clavellata sp. nov. show distinct differences in the maculae on the pronotum, the band in the interocular space (H. clavellata sp. nov.: yellow and straight transverse band, Homalosilpha sp.: yellow and tortuous transverse band) and even in the female genitalia (H. clavellata: anterior of basivalvula with spots, the shape of first valvule uniform, Homalosilpha sp.: surface of basivalvula with fold, the end of the first valvula swollen). Finally, a total of 8 Homalosilpha and Mimosilpha species were recovered after combining the results of molecular data with morphological data, of which 4 species are new to science: H. obtusangula sp. nov., H. recta sp. nov., H. alba sp. nov., and H. clavellata sp. nov. More details about these new Homalosilpha species are provided below.
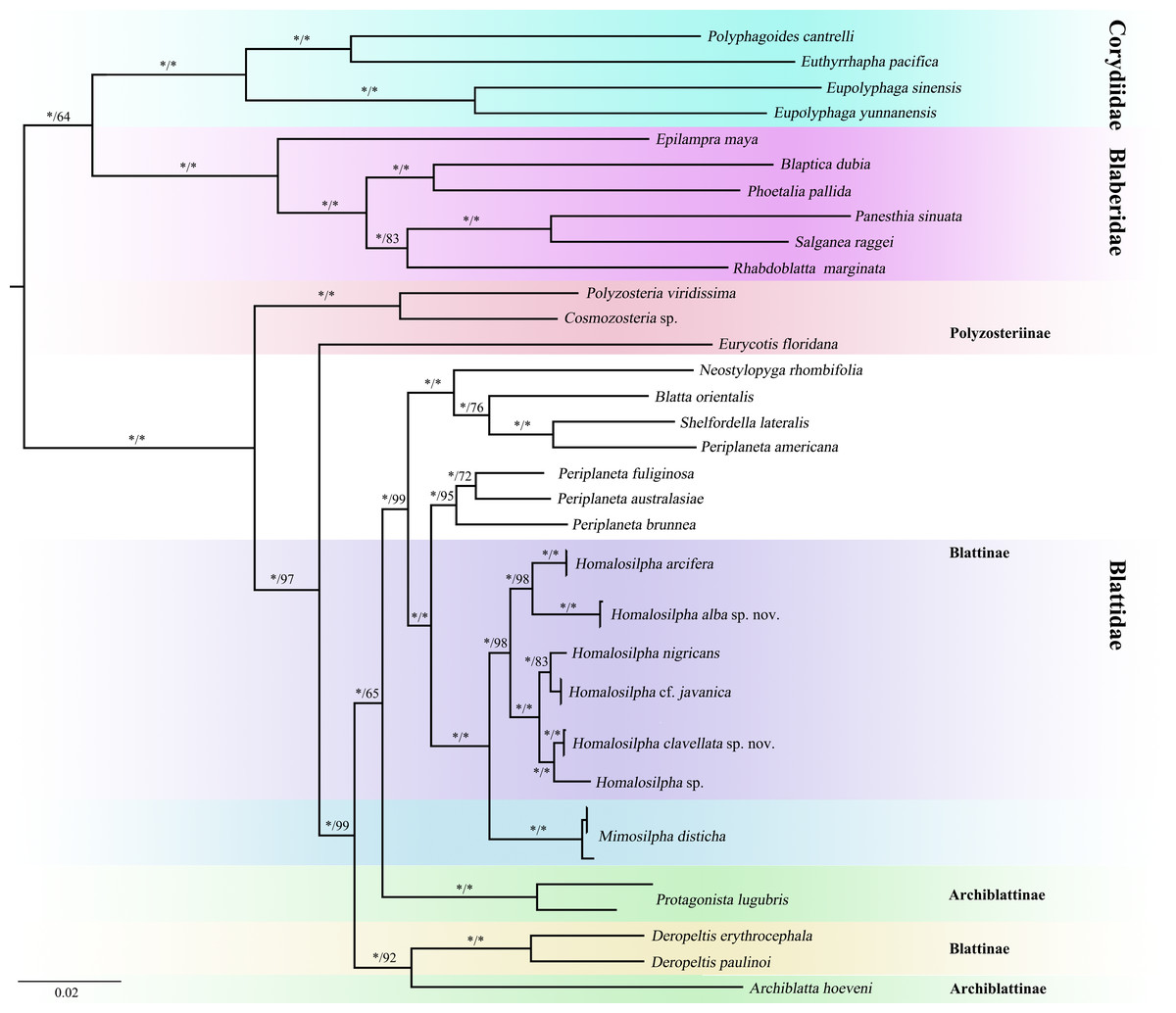
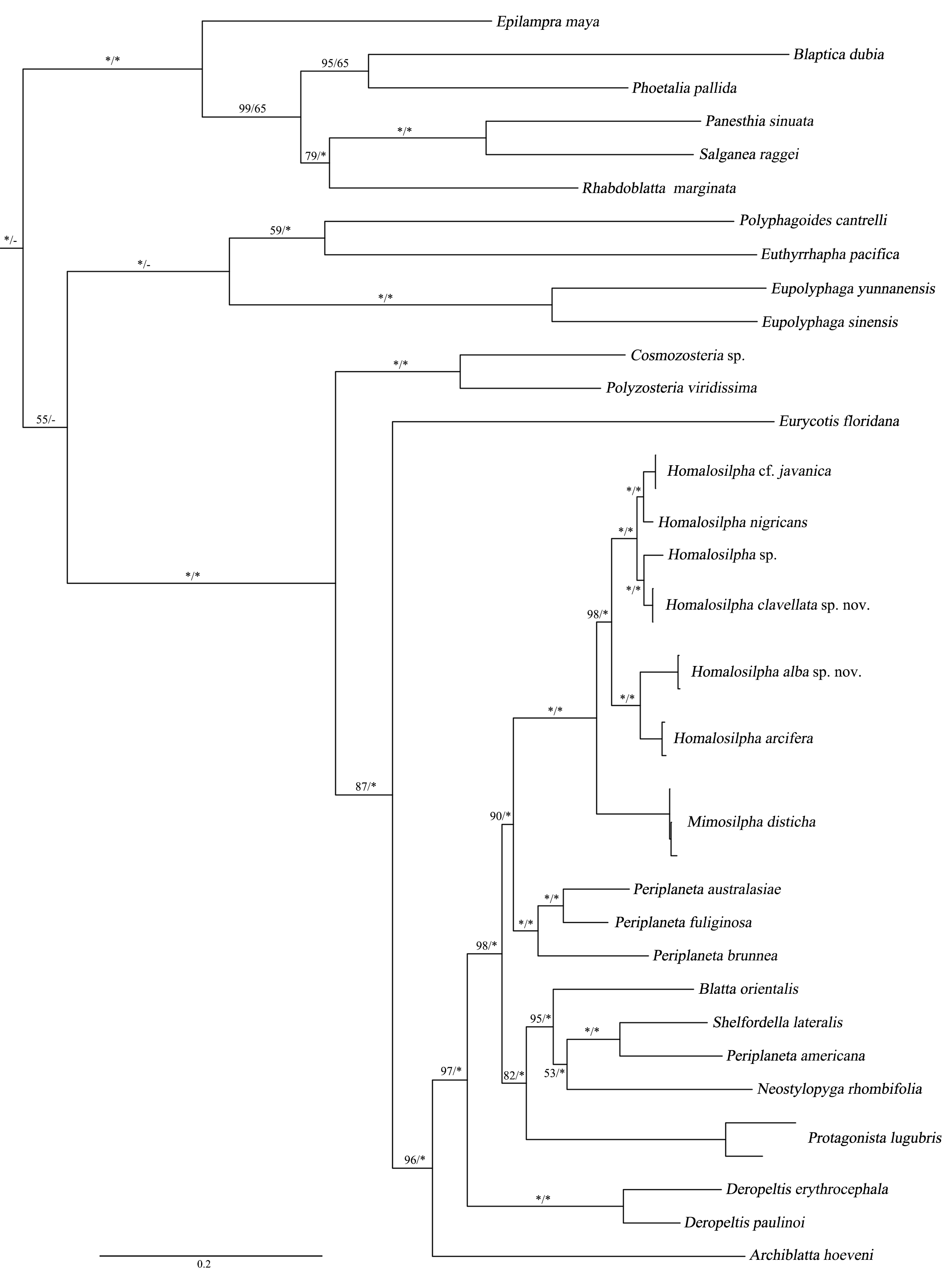
Figure 3: Maximum likelihood (ML) tree of the cockroaches based on six genes.
ML tree derived from analysis of combined data 12S, 16S, 18S, 28S, COII, and H3 genes. Branch labels are support for our analyses in the following order: bootstrap supports of the maximum-likelihood tree, Bayesian posterior probabilities of the Bayesian tree; asterisks (*) indicate 100% support for a given analysis. The topology shown was totally similar to that derived from BI analysis.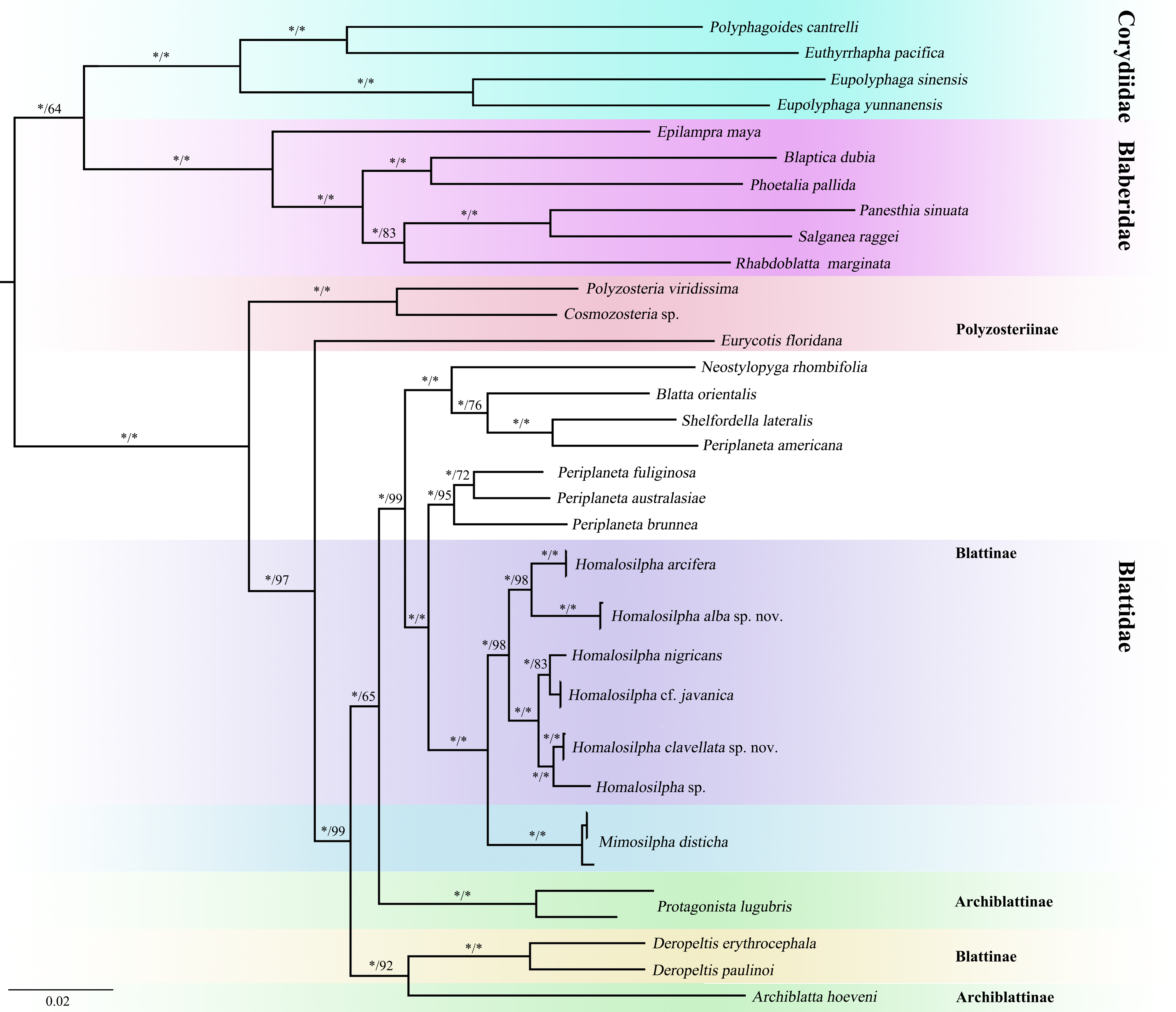{kind=link}

Figure 4: Photographs of middle and hind tibiae of Homalosilpha and Mimosilpha species.
(A–B) H. nigricans Princis, 1966a. (C–D) H. alba sp. nov. (E–F) M. disticha. Middle and hind tibiae are seen on dorsal view. Scale bars = 5 mm.{kind=link}
Relationship of HomalosilphaandMimosilphainferred from two phylogenetic analyses
We acquired 12 12S, 12 16S, 11 18S, 12 28S, 10 COII, 12 H3 sequences with accession numbers MW218556–MW218578, MW219581–MW218604 and MW201809–MW201830 (Table S2). For the concatenate dataset (12S, 16S, 18S, 28S, COII, H3), our likelihood and Bayesian phylogenetic analyses yielded totally identical topologies with generally high node support (Fig. 3 and Fig. S1). All genera of Blattidae clustered together and formed a highly supported monophyletic group (MLB = 100, BPP = 100). Five recognized major lineages of Blattidae from ML and BI inferences were recovered, represented by members of Polyzosteriinae, Archiblattinae and Blattinae. According to our inferred trees, Polyzosteriinae, Archiblattinae and Blattinae were paraphyletic with high support values.
The topology derived from ML and BI analyses shows that all Homalosilpha members cluster together and are recovered as the sister group of Mimosilpha (MLB = 100, BPP = 100). H. clavellata sp. nov. and Homalosilpha sp. were recovered as sister groups with strong support values, which is consistent with the inference based on COI data. And the result is same as the trees of seven genes (12S, 16S, 18S, 28S, COI, COII, H3) (Figs. S2 and S3).
The electronic version of this article in Portable Document Format (PDF) will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: [74F6BF88-9FFD-40DB-9C8B-D7650491F0EE]. The online version of this work is archived and available from the following digital repositories: PeerJ, PubMed Central and CLOCKSS.
Taxonomy
Homalosilpha Stål, 1874
Homalosilpha Stål, 1874: 13. Type species: Homalosilpha ustulata Burmeister, 1838. Kirby, 1904: 143; Shelford, 1910: 19; Hanitsch, 1915: 112; Princis, 1966a: 49; Princis, 1966b: 457; Roth, 1999: 172.
Diagnosis. Body medium to large, flat, smooth, generally brownish to blackish. Antennae long, slender. Pronotum flat, decorated with maculae, sides not deflexed, greatest width near the middle. Tegmina and wings extending considerably beyond the apex of the abdominal tip. Legs long, front femora Type A2, the outer edge of the middle and hind tibiae each with three rows of spines (Fig. 4); tarsal claws simple, symmetrical. Supra-anal plate broad. Cerci long. Subgenital asymmetrical. Male genitalia: left and right phallomere all consisting of three parts. L1 fold sclerite; margin of L2d with serration; posterior of L2v produced with a spiny projection to the end; L3 unciform, the terminal bifurcate. R1 foot-shaped, left margin with two spines; terminal of R2d projected, R2v folded sclerite; R3 broad and folded sclerite. In the nymph, the body shows zebra-like stripes.
Checklist of Homalosilphaworldwide
| Homalosilpha alba sp. nov. China |
| Homalosilpha arciferaBey-Bienko, 1969 China, Vietnam |
| Homalosilpha contraria (Walker, 1868) Philippines |
| Homalosilpha decorata (Serville, 1838) Indonesia (Sumatra), Indonesia (Java Island), Borneo Island |
| Homalosilpha gaudensShelford, 1910 China, Vietnam |
| Homalosilpha haaniPrincis, 1966a Indonesia |
| Homalosilpha clavellata sp. nov. China |
| Homalosilpha javanicaBey-Bienko, 1969 Indonesia |
| Homalosilpha kryzhanovskiiBey-Bienko, 1969 China |
| Homalosilpha nigricans Princis, 1966 Congo Democratic Republic |
| Homalosilpha obtusangula sp. nov. China |
| Homalosilpha recta sp. nov. China |
| Homalosilpha quadrimaculataRoth, 1999 Malaysia |
| Homalosilpha ustulata (Burmeister, 1838) China, Myanmar, Malaysia, Indonesia, Philippines |
| Homalosilpha validaBey-Bienko, 1969 China |
| Homalosilpha vicina (Brunner von Wattenwyl, 1865) Guinea, Cameroon, Uganda |
| Homalosilpha sp. China |
Key to Homalosilpha species worldwide
-
1. Pronotum mostly black …2
-
Pronotum mostly white…3
-
2. Pronotum with two vertical white stripes near the lateral margin …H. decorata
-
Pronotum with four white spots respectively at the anterior margin and middle…H. quadrimaculata
-
3. Pronotum with a large black spot at center …4
-
Pronotum with multiple black spots or stripes …6
-
4. The large spot connecting both the front and hind margins …H. haani
-
The large spot only connected with the hind margin …5
-
5. Pronotum broadest near the hind margin, trapeziform …H. nigricans
-
Pronotum broadest near the middle …H. gaudens
-
6. Face pale yellow …H. arcifera
-
Face black …7
-
7. Vertex yellow to yellowish brown …8
-
Vertex black …10
-
8. Abdomen with yellow spots …H. valida
-
Abdomen brownish black to dark …9
-
9. The hind margin of supra-anal plate slightly concave, lateral angles blunt …H. recta sp. nov.
-
The hind margin of supra-anal plate round, lateral angles sharp …H. alba sp. nov.
-
10. Interocular space with a yellow straight band …11
-
Interocular space with a yellow band …H. ustulata
-
11. The lateral pattern and vertical pattern of pronotum connected with posterior band, spots on both sides of the lateral pattern …H. vicina
-
The middle of pronotum with spots or spots and stripes, the both sides of lateral pattern without spots…12
-
12. Pronotum without vertical pattern …H. contraria
-
Pronotum with vertical pattern …13
-
13. The vertical pattern of pronotum with one stripe …14
-
The vertical pattern of pronotum with spots …16
-
14. The vertical pattern with one short and bold stripe …H. clavellata sp. nov.
-
The vertical pattern with one long stripe and posterior vertical pattern bifurcated …15
-
15. The vertical pattern with pinstripe and the anterior pattern connected with lateral pattern …H. obtusangula sp. nov.
-
The vertical pattern with black stripe and the central pattern connected with vertical pattern …Homalosilpha sp.
-
16. The hind margin of supra-anal plate with two sharp valves …H. javanica
-
The hind margin of supra-anal plate with two round valves …H. kryzhanovskii
Holotype. CHINA: Tibet: male (SWUB125-H) Beibeng Township, Mêdog County, 2013.VII.31, Xinglong Bai & Junsheng Shan leg.
Paratype. 1 female (SWUB125-P1), CHINA: Yunnan: Mt. Fenshuiling, Jinping County, 2016.V, Tianlong He leg.
Etymology. The species epithet comes from the Latin word obtusangulus in reference to the lateral angles of supra-anal plate being obtuse.
Description. Male. Body length 24.4 mm; overall length including tegmen 33.8 mm; pronotum length × width 6.4 × 8.4 mm. Female. Body length 26.5 mm; overall length including tegmen 36.3 mm; pronotum length × width 6.8 × 8.9 mm.
Coloration. Similar in both sexes. Body generally dark brown (Figs. 5A and 5B). Interocular space with a whitish transverse band. Pronotum white, margins black outlined, hind margin black; center of the pronotum with symmetrical blackish markings (Figs. 5C and 5D). Tegmen reddish brown, wing hyaline, brownish except the anal area (Figs. 7A and 7B). Tibiae and tarsi dark reddish brown (Fig. 5B).
Head with vertex unsheltered by pronotum. Interocular distance slightly narrower than the distance between antennae sockets. Pronotum nearly heptagonal, broadest near the middle; the middle of pronotum with black and symmetrical patterns, the anterior pattern connected with lateral pattern, the lateral pattern arc-shaped, central pattern dart-like, the posterior of vertical pattern bifurcated, the posterior band bold (Figs. 5C and 5D). Male supra-anal plate broad and symmetrical, nearly trapezoidal, but with a deep, obtuse-angled invagination at the middle of hind margin; male subgenital plate asymmetrical, left hind corner more protruded than on the right, styli similar, long (Figs. 8A and 8B).
Male genitalia. Left phallomere: L1 fold sclerite; margin of L2d with small serration, posterior of L2v produced as a spiny projection to the end; L3 unciform, the base broad, tapering down, the terminal bifurcate, one branch short, the other long (Fig. 8C). Right phallomere: R1 foot-shaped, left margin with two spines; terminal of R2d sharp, R2v fold sclerite; R3 broad and fold sclerite (Fig. 8C).
Remarks. H . obtusangula sp. nov. resemble H. kryzhanovskii, but can be distinguished from H. kryzhanovskii by the following characters: (1) the pronotum with stripes in the former, while the latter with spots; (2) the hind margin of supra-anal plate deeply concave, while the latter, hind margin slightly concave; (3) the lateral corner sharp in the former, while the latter, the lateral corner rounded
Holotype. CHINA: Yunnan: male (SWUB126-H) Banhong Township, Cangyuan County, 1130 m, 2008.VII.16-18, Jishan Xu & Zhenhua Gao leg.
Etymology. The species epithet ‘rectus’ refers to the hind margin of supra-anal plate and subgenital plate being straight.
Description. Male. Body length 28.3 mm; overall length including tegmen 39.9 mm; pronotum length × width 7.3 × 10.6 mm.
Coloration. Body generally dark brown (Figs. 5E and 5F). Vertex and the area between eyes and ocelli white. Pronotum white, subhyaline, with black outline weak, hind margin blackish weakly, middle of the pronotum with sparse, symmetrical blackish markings (Figs. 5G and 5H). Tegmina yellowish brown, wings hyaline, dark brownish except the anal area (Figs. 7C and 7D). Tibiae and tarsi dark reddish brown.
Head with vertex unsheltered by pronotum. Interocular distance narrower than distance between antennae sockets. Pronotum nearly hexagonal, anterior and hind margins slightly outward, broadest near the middle; the anterior pattern triangular, the lateral pattern and central pattern with two parts, the vertical pattern with only one spot, the posterior band indistinct (Figs. 5E and 5F). Male supra-anal plate broad and symmetrical, nearly trapezoidal, with a very shallow, obtuse-angled invagination at the middle of hind margin, hind lateral angles slightly protruded laterally; subgenital plate of male asymmetrical, quadrate, right lateral hind corner narrowed, styli similar, long (Figs. 8D and 8E).
Male genitalia. Left phallomere: L1 fold sclerite; posterior margin of L2d with small spines, posterior of L2v produced as a spiny projection to the end; L3 unciform, the base broad, tapering down, the terminus bifurcate (Fig. 8F). Right phallomere: R1 foot-shaped, left margin with two unequal spines, the spine on the top longer than the bottom; terminal of R2d unciform, R2v fold sclerite; R3 broad (Fig. 8F).
Remarks. H. recta sp. nov. is similar to H. kryzhanovskii (Figs. 6I–6L, 8M–8O) and H. alba sp. nov. (Figs. 5I–5L, 8G–8I), but can be distinguished from H. kryzhanovskii by the following characters: (1) the composition of black spots in pronotum; (2) vertex white, while in H. kryzhanovskii black; and (3) the hind margin of supra-anal plate slightly concave and with lateral sides curving laterally, however in H. kryzhanovskii the two hind angles nearly round. Furthermore, it can be distinguished from H. alba sp. nov. by the following characters: (1) the composition of black spots with slight difference; and (2) the hind margin of the supra-anal plate slightly concave and blunt, however in H. alba, the lateral angles of the supra-anal plate are sharp.

Figure 5: Photographs of bodies, pronotums and faces of four species of Homalosilpha.
(A–D) H. obtusangula sp. nov., male holotype. (E–H) H. recta sp. nov., male holotype. (I–L) H. alba sp. nov., male holotype. (M–P) Homalosilpha sp., female. Scale bars = 10 mm (A, B, E, F, I, J, M, N); 5 mm (C, D, G, H, K, L, O, P).{kind=link}

Figure 6: Photographs of bodies, pronotums and faces of two species of Homalosilpha and one species of Mimosilpha.
(A–D) H. clavellata sp. nov., male holotype. (E–H) H. clavellata sp. nov., female paratype. (I–L) H. kryzhanovskii. (M–P) M. disticha. Scale bars = 10 mm (A, B, E, F, I, J, M, N); 5 mm (C, D, G, H, K, L, O, P).{kind=link}
Holotype. CHINA: Hainan: male (SWUB127-H) Yajiang Scenery Spot, Mt Bawangling, Cangjiang County, 1130 m, 2015.IV.30, Lu Qiu & Qikun Bai leg.
Paratypes. China: Hainan: 1 female (SWUB127-P1). Maogan Township, Baoting County, 2015.IV.11-12, Lu Qiu & Qi-kun Bai leg; 1 female (SWUB127-P2). Mt Diaoluoshan, Lingshui County, 1964.III.28, Si-Kong Liu; 1 female (SWU) Ledong County, 1954.V.4, Keren Huang leg.
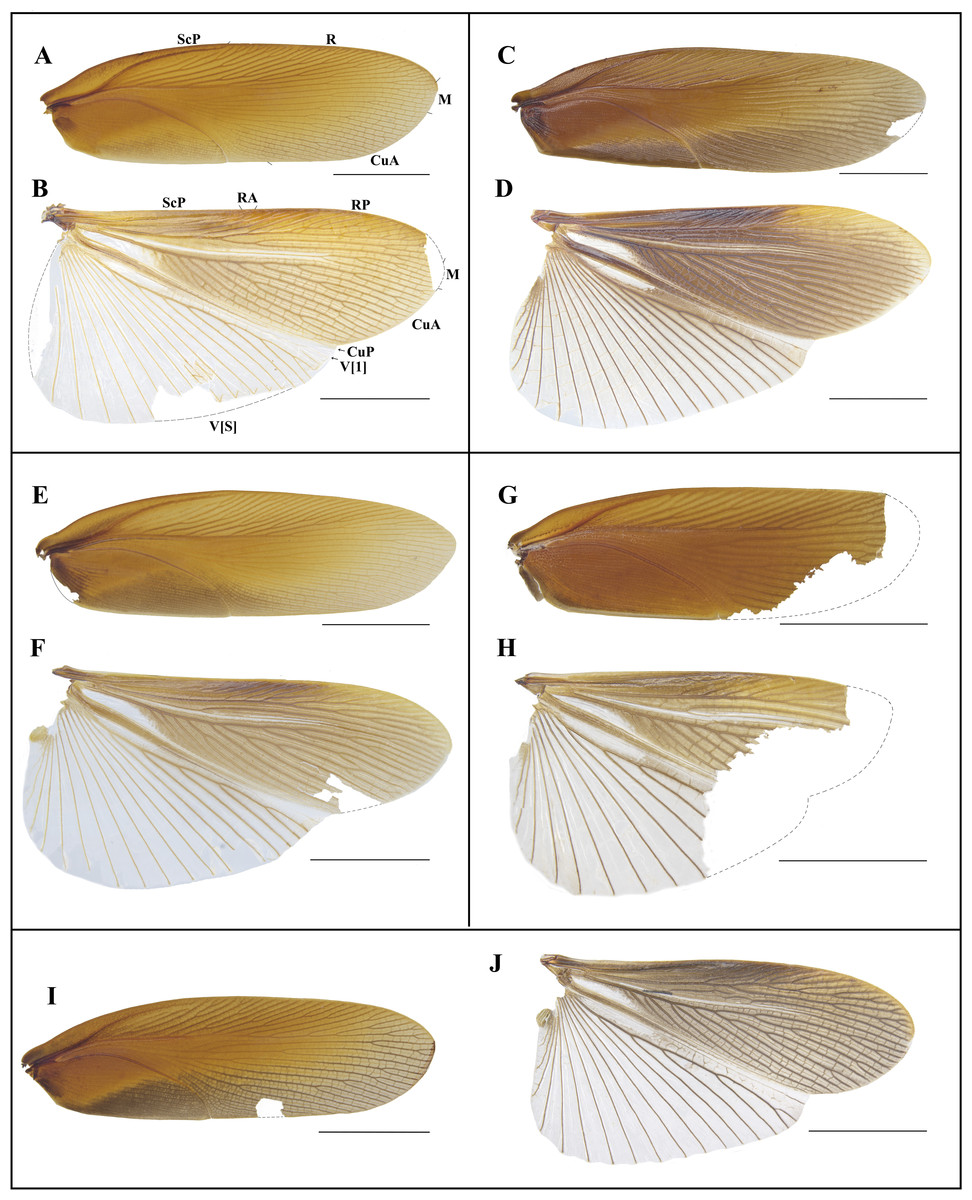
Figure 7: Photographs of tegmen and hind wings of Homalosilpha.
(A–B) H. obtusangula sp. nov. (C–D) H. recta sp. nov.; (E–F) H. alba sp. nov. (G–H) Homalosilpha sp. (I–J) H. clavellata sp. nov. Scale bars = 10 mm.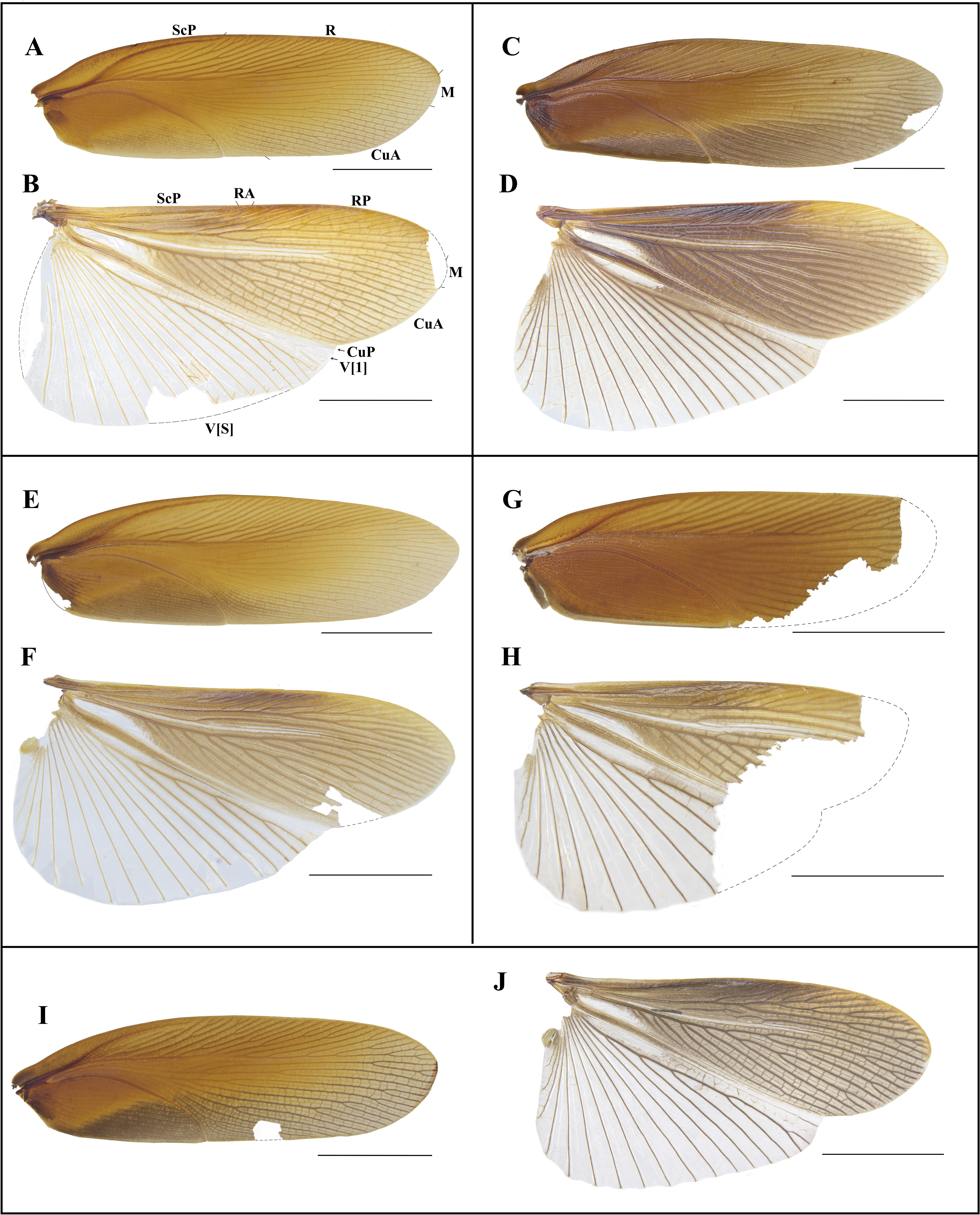{kind=link}
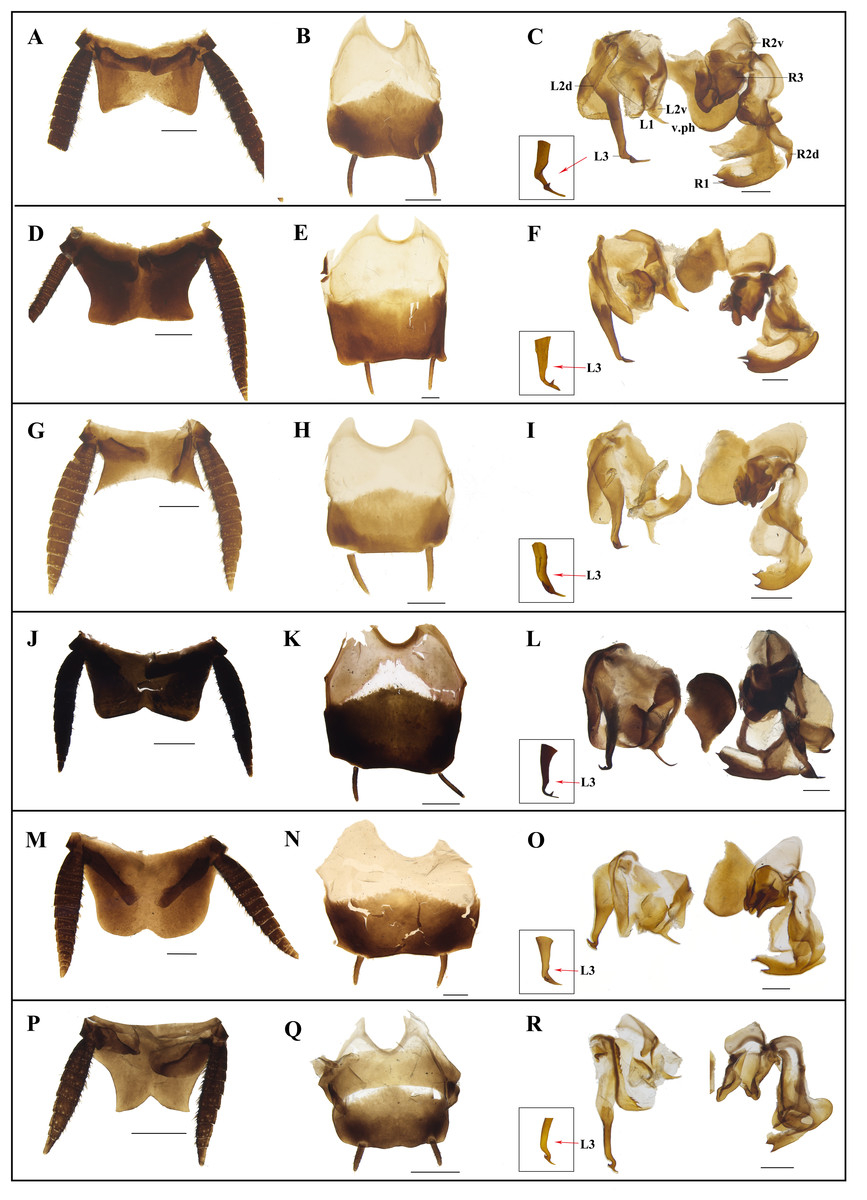
Figure 8: Photographs of male genitalia of Homalosilpha and Mimosilpha species.
(A–C) H. obtusangula sp. nov., holotype. (D–F) H. recta sp. nov., holotype. (G–I) H. alba sp. nov., holotype. (J–L) H. clavellata sp. nov., holotype. (M–O) H. kryzhanovskii. (P–R) M. disticha. A, D, G, J, M, P, supra-anal plate, dorsal view; B, E, H, K, N, Q, subgenital plate, ventral view; C, F, I, L, O, R, phallomere, dorsal view. Scale bars = 1 mm.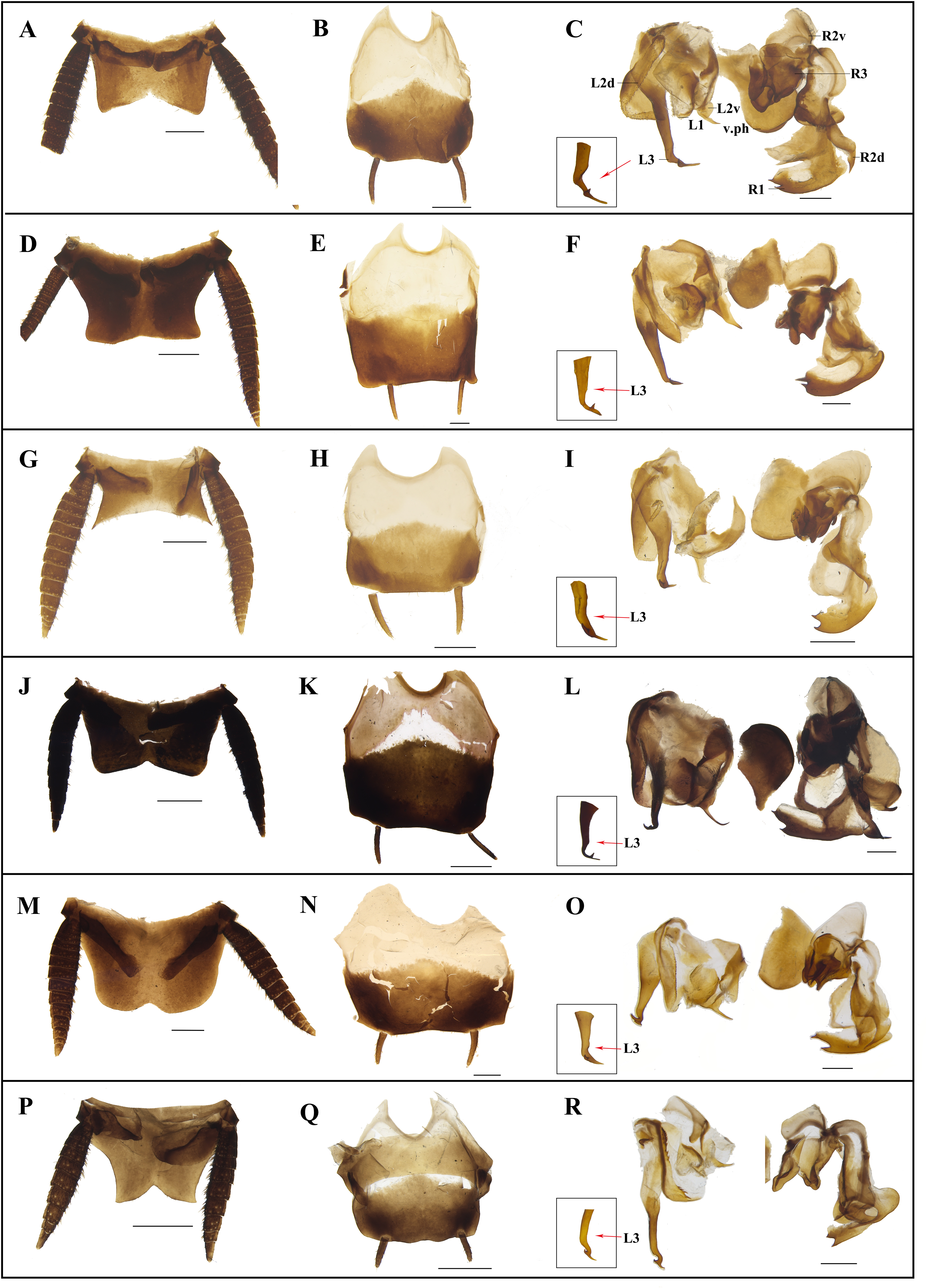{kind=link}

Figure 9: Habitats of Homalosilpha. and Mimosilpha species from China.
(A) M. disticha found on lichenous and mossy tree trunk (Dadugang, Jinghong, Yunnan). (B) H. kryzhanovskii found under the bark (Dadugang, Jinghong, Yunnan). (C) Nymphs of M. distincta, one was eating a dipteran insect (Dadugang, Jinghong, Yunnan). (D) A female Homalosilpha sp. (Mt. Ailaoshan, Yunnan). (E) A nymph of Homalosilpha species from Mengla, Xishuangbanna, Yunan (collected by Jianyue Qiu and Hao Xu). (A–C) photographed by Xinran Li. (D–F) photographed by Lu Qiu.{kind=link}
Etymology. The species epithet is derived from the Latin word ‘albus’, referring to the vertex being white.
Description. Male. Body length 27.2 mm; overall length including tegmen 33.1 mm; pronotum length × width 6.1 × 8.5 mm. Female. Body length 28.3 mm; overall length including tegmen 36.4 mm; pronotum length × width 7 × 9.9 mm.
Coloration: Similar in both sexes. Body dark brown (Figs. 5I and 5J). Vertex and the area between eyes and ocelli white. Pronotum white, margins black outlined, hind margin black (Figs. 5K and 5L). Tegmina reddish brown, wings hyaline, brown except the anal area (Figs. 7E and 7F). Middle of the abdomen yellowish brown. Tibiae and tarsi reddish brown (Fig. 5L).
| Genes | Forward/ Reverse | Sequences (5′-3′) | References |
|---|---|---|---|
| 12S | F | ATCTATGTTACGACTTAT | Inward, Beccaloni & Eggleton (2007) |
| R | AAACTAGGATTAGATACCC | Kambhampati (1995) | |
| 16S | F | CGCCTGTTTAACAAAAACAT | Simon et al. (1994) |
| R | CGCCTGTTTAACAAAAACAT | Cognato & Vogler (2001) | |
| 18S | F | CTGGTTGATCCTGCCAGT | Hillis & Dixon (1991) |
| R | TAATGATCCTTCCGCAGGTTCACCT | Halanych, Lutz & Vrijenhoek (1998) | |
| 28S | F | ACACGGACCAAGGAGTCTAAC | Inward, Beccaloni & Eggleton (2007) |
| R | GTCCTGCTGTCTTAAGCAACC | ||
| COII | F | AGAGCWTCACCTATTATAGAAC | Park et al. (2004) |
| R | GTARWACRTCTGCTGCTGTTAC | ||
| H3 | F | ATGGCTCGTACCAAGCAGACVGC | Inward, Beccaloni & Eggleton (2007) |
| R | ATATCCTTRGGCATRATRGTGAC | ||
| COI | F | GGTCAACAAATCATAAGATATTGG | Folmer et al. (1994) |
| R | TAAACTTCAGGGTGACCAAAAAATCA |
Head with vertex unsheltered by pronotum. Interocular distance narrower than the distance between antennae sockets. Pronotum nearly a hexagon, anterior and hind margins slightly outward, broadest near the middle; the anterior pattern and central pattern triangular, the lateral pattern with three parts, the vertical pattern with only one spot, the posterior band bold and with two spots on the band (Fig. 5K). Male supra-anal plate symmetrical, nearly trapezoidal, hind margin widely concave, lateral hind angles sharp and protruded outward; subgenital plate of male asymmetrical, left hind corner angled while right hind corner round, styli similar (Figs. 8G and 8H).
| Species | Accession number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Homalosilpha cf. javanica | MW201581 | ||||||||||||
| Homalosilpha cf. javanica | MW201582 | 0.000 | |||||||||||
| Homalosilpha nigricans | MW201583 | 0.036 | 0.036 | ||||||||||
| Homalosilpha clavellata sp. nov. | MW201584 | 0.044 | 0.044 | 0.039 | |||||||||
| Homalosilpha clavellata sp. nov. | MW201585 | 0.044 | 0.044 | 0.039 | 0.000 | ||||||||
| Homalosilpha sp. | MW201586 | 0.054 | 0.054 | 0.047 | 0.031 | 0.031 | |||||||
| Homalosilpha kryzhanovskii | MW201587 | 0.051 | 0.051 | 0.046 | 0.035 | 0.035 | 0.056 | ||||||
| Homalosilpha arcifera | MW201588 | 0.079 | 0.079 | 0.084 | 0.072 | 0.072 | 0.083 | 0.084 | |||||
| Homalosilpha alba sp. nov. | MW201590 | 0.070 | 0.070 | 0.081 | 0.064 | 0.064 | 0.079 | 0.077 | 0.047 | ||||
| Homalosilpha alba sp. nov. | MW201591 | 0.072 | 0.072 | 0.082 | 0.065 | 0.065 | 0.081 | 0.079 | 0.049 | 0.002 | |||
| Mimosilpha disticha | MW201592 | 0.081 | 0.081 | 0.077 | 0.074 | 0.074 | 0.079 | 0.084 | 0.075 | 0.075 | 0.077 | ||
| Mimosilpha disticha | MW201593 | 0.081 | 0.081 | 0.077 | 0.074 | 0.074 | 0.079 | 0.084 | 0.075 | 0.075 | 0.077 | 0.000 | |
| Mimosilpha disticha | MW201594 | 0.077 | 0.077 | 0.077 | 0.069 | 0.069 | 0.077 | 0.081 | 0.070 | 0.074 | 0.075 | 0.006 | 0.006 |
Male genitalia: Left phallomere: L1 lamellar; margin of L2d with small spines, posterior of L2v produced with a spiny projection to the end; L3 with a spiny projection, the basal broad, tapering down, the terminus bifurcate, the lengths equal (Fig. 8I). Right phallomere: R1 foot-shaped, left margin with two equal spines; terminal of R2d unciform, R2v fold sclerite; R3 with complicated and broad sclerite (Fig. 8I).
Remarks. H. alba is similar to H. kryzhanovskii (Figs. 6I–6L, 8M–8O) but can be distinguished from H. kryzhanovskii by the following characters: (1) the black spots smaller and thinner than H. kryzhanovskii in pronotum; and (2) the lateral angles of supra-anal plate sharp, however in H. kryzhanovskii nearly round.
Holotype. CHINA: Yunnan: male (SWUB128-H), Tongbiguan Township, Yingjiang County, 1130 m, 2018.IV.30, Lu Qiu & Wenbo Deng leg.
Paratype. 1 female (SWUB128-P1), CHINA: Yunnan: Tongbiguan Township, Yingjiang County, 1130 m, 2018.IV.30, Lu Qiu & Wenbo Deng leg.
Etymology. This species epithet is derived from the Latin word ‘clavellatus’ referring to the terminal of R2d being produced in a stick shape.
Description. Male. Body length 22.8 mm; overall length including tegmina 29.0 mm; pronotum length × width 4.4 × 7.1 mm. Female. Body length 23.2 mm; overall length including tegmina 29.3 mm; pronotum length × width 5.5 × 8.1 mm.
Coloration: body generally black. Interocular space with a white transverse band, distal half of clypeus white. Pronotum white, margins black outlined; center of the pronotum with symmetrical blackish markings (Figs. 6A and 6B). Tegmen reddish brown, wings hyaline, blackish brown except the anal area (Figs. 7G and 7H). Tibiae and tarsi reddish brown (Fig. 6B).
Head with vertex unsheltered by pronotum. Interocular distance slightly narrower than the distance between antennae sockets. Pronotum nearly hexagonal, broadest on the basal half but near the middle, the anterior pattern connected with lateral pattern, the lateral pattern bold, the vertical pattern connected with posterior pattern, the posterior band bold and with two spots on the band (since the pronotum of the holotype male is deformed, thus this description is only based on the female paratype) (Figs. 6A–6H). Male supra-anal plate broad and symmetrical, nearly trapezoidal, but with a shallow, obtuse-angled invagination at the center of hind margin; male subgenital plate asymmetrical, left hind corner more protruded than that of the right, styli similar, thin (Figs. 8J and 8K).
Male genitalia: Left phallomere: L1 broad and lamellar; margin of L2d with small and dense spines, posterior of L2v produced as an acerose spiny projection to the end; L3 with a spiny projection, the basal broad, tapering down, the terminal bifurcate (Fig. 8L). Right phallomere: R1 foot-shaped, posterior with two spines; terminal of R2d clubbed, R2v fold sclerite and the end with a spine; R3 with broad sclerite (Fig. 8L).
Remarks. H . clavellata sp. nov. is similar to H. arcifera, but can be distinguished by the following characters: (1) different irregular maculae on the pronotum, large in H. clavellata sp. nov., while H. arcifera, small; (2) face and body black in H. clavellata sp. nov., while in H. arcifera, yellowish white. Besides, the pronotum of the male holotype is deformed with posterior and anterior margins deeply concave. We utilized DNA barcoding to pair this species, and the genetic distance between them was 0.00%.
Material examined. 2 females (SWUB129-1, SWUB129-2), CHINA: Yunnan: Yaonan Village, Mt Ailao, Xinping County, 2018.V.24, Lu Qiu & Zhiwei Dong leg.
Description. Female. Body length 22.8–23.5 mm; overall length including tegmen 29.0–30.2 mm; pronotum length × width 4.4–5.0 × 6.9–7.1 mm.
Coloration: Body generally black. Interocular space with a white transverse band. Pronotum white, margins with black outline, hind margin black; center of the pronotum with symmetrical blackish markings. Tegmina brown, wings hyaline. Abdomen brown. Tibiae and tarsi yellow brown.
Head with vertex unsheltered by pronotum. Interocular distance slightly narrower than the distance between antennae sockets. Pronotum nearly hexagonal, broadest near the middle; the anterior pattern polygonal, the lateral pattern with two patterns, the central pattern connected with vertical pattern, the vertical pattern bold and the posterior bifurcated, the posterior band bold. Supra-anal plate broad, symmetrical and with ridges, cerci with obvious segmentation. Subgenital plate symmetrical, divided in two valves.
Natural History. Individuals were captured during night on the village roads (Lu Qiu, personal observation).
| MimosilphaBey-Bienko, 1957 |
| MimosilphaBey-Bienko, 1957: 905. Type species: Mimosilpha distichaBey-Bienko, 1957 |
| Princis, 1966b: 459. |
Diagnosis. Body small. The diagnoses are similar to Homalosilpha except the outer edge of middle and hind tibiae with two rows of spines (Fig. 4) and the margin of L2d with bigger serration (Fig. 8).
| Mimosilpha distichaBey-Bienko, 1957 |
| (Figs. 6M–6P, 8P–8R) |
| Mimosilpha distichaBey-Bienko, 1957: 905; Princis, 1966b: 459 |
Material examined. 11 males, 7 females (SWUB130-1, SWUB130-2…SWUB130-17, SWU130-18), CHINA: Yunnan: Dadugang, Jinghong City, Xishuangbanna, 2014.IV.28-29, Xinran Li & Hongguang Liu leg; 1 male (SWU130B-19), CHINA: Yunnan: Jinghong City, Xishuangbanna, 545 m, 1974.V.14-15, Io Chou & Feng Yuan leg.
Redescription. Male. Body length 18.2–20.5 mm; overall length including tegmen 22.6–24.9 mm; pronotum length × width 3.7–4.6× 5.4–6.3 mm. Female. Body length 19.2–22.4 mm; overall length including tegmen 21.3–25.7 mm; pronotum length × width 4.3–4.7 × 5.7–6.5 mm.
Coloration. Similar in both sexes. Body dark brown (Figs. 6M and 6N). Pronotum white, with a large central black spot on middle disk, margins with black outline, hind margin black (Figs. 6O and 6P). Tegmen dark brown, wing hyaline, brown except the anal area. Abdomen brown to dark brown. Tibiae and tarsi brown (Fig. 6N).
Head with vertex unsheltered by pronotum. Interocular distance narrower than the distance between antennae sockets. Pronotum nearly hexagonal, broadest near the middle, large black spot connected to the hind margin. Legs with sparse spines. Supra-anal plate of male broad and symmetrical, nearly trapezoidal, posterior margin widely concave, the lateral margin concave, both lateral hind angles sharp; male subgenital plate asymmetrical, left hind corner round, hind margin wavy (Figs. 8P and 8Q).
Male genitalia: Left phallomere: L1, L2 and L3. L1 lamellar; margin of L2d with small spines, posterior of L2v produced a spiny projection to the end; L3 with a spiny projection, the basal broad, tapering down, the terminal bifurcated (Fig. 8R). Right phallomere: R1, R2 and R3. R1 foot-shaped, posterior with two spines; terminal of R2d unciform, R2v fold sclerite; R3 simple and broad (Fig. 8R).
Discussion
Due to the similar pattern bearing in the pronotum by some Homalosilpha species (Bey-Bienko, 1969), i.e., H. arcifera, H. javanica, H. kryzhanovskii, H. ustulata, H. valida, H. alba sp. nov., H. recta sp. nov., it is very challenging to distinguish them based only on morphological data. Fortunately, our results show that DNA-based species delimitation methods perform well for Homalosilpha species, and also did well in matching male and female. Therefore, COI can be recommended as a single DNA barcode for Homalosilpha.
Homalosilpha is closely related to Mimosilpha not only in appearance but also in phylogenetic analysis. Apart from the slight difference in body size (Homalosilpha: medium to large, Mimosilpha: small), the diagnostic character given by Bey-Bienko (1957) to separate Mimosilpha and Homalosilpha was the number of rows of spines existing in the edge of hind tibiae (Homalosilpha: 3, Mimosilpha: 2). ML and BI analyses showed that Homalosilpha and Mimosilpha were monophyletic groups. Homalosilpha was recovered as the sister group of Mimosilpha.
Periplaneta americana, whose tibiae are also ornamented with strong spines, usually depend on their spines to brace themselves against the sides of a burrow and provide a stable platform for the transmission of force (Bell, Nalepa & Roth, 2007). Sometimes spines at the tip of tibiae will assist the tarsal claws to climb rough surfaces (Bell, Nalepa & Roth, 2007). It can be inferred that the spines on the tibiae of Mimosilpha and Homalosilpha also play a certain role in the process of movement. According to information observed so far (25 specimens of Homalosilpha and 25 specimens of Mimosilpha, including 12 Homalosilpha species from original description), we inferred the number of rows of spines to be stable in different individuals and even in nymphs. In field collection, one H. kryzhanovskii was found on wooden poles and two Homalosilpha sp. were found on the road, but more Mimosilpha disticha were found on tree trunks covered with lichen and moss (Li XR, pers. obs., 2014) (Fig. 9). We speculate that different rows of spines exhibited by Mimosilpha and Homalosilpha on the edge of hind tibiae might reflect adaptation to different habitats. So we do not treat Mimosilpha as the junior synonym of Homalosilpha although only minor morphological differences exist between them. We will explore more evidence concerning this relationship later.
Although our data are insufficient, the ML and BI phylogenetic analyses also reflected that Polyzosteriinae, Archiblattinae and Blattinae are paraphyletic with high support values. At the beginning, some scholars proposed that Archiblatta should be placed in Blattinae because of the valvular structure of the female subgenital plate (Shelford, 1910; Bruijning, 1948); but Roth (2003) still recognized Archiblattinae as a subfamily because of the absence or greatly reduced femoral armament. Murienne (2009) concatenated 12S and H3 to construct ML and BI, and the results revealed Archiblattinae (Archiblatta) and Blattinae form a monophyletic group. However, Djernæs, Klass & Eggleton (2015) proposed that Polyzosteriinae (Drymaplaneta, Eurycotis), Archiblattinae (Archiblatta) and Blattinae (Periplaneta, Deropeltis) might be artificial and not natural groups based on a phylogenetic tree combining 8 genes (12S, 16S, 18S, tRNA-Leu, COII, tRNA-Lys, H3). Bourguignon et al. (2018) also discovered Archiblattinae (Protagonisa) and Blattinae are paraphyletic via mitochondrial phylogeny. As these issues are reflected by our results, the monophyly of Polyzosteriinae, Archiblattinae and Blattinae should be questioned and more attention paid to solving this dilemma. Therefore, additional sampling of Archiblattinae and Blattinae will help improve our understanding of Blattidae in the future.
Conclusion
Our study shows that molecular methods based on COI data delimit MOTUs for Homalosilpha that are highly consistent with those based on morphological characters. Therefore, COI is recommended as an effective DNA Barcode for Homalosilpha. Our phylogeny based on mitochondrial and nuclear genes reveals the close relationship of Homalosilpha and Mimosilpha, but the introduction of more samples is therefore recommended to better improve our understanding of those highly similar genera. Although limited samples of Blattidae are included in our study, the paraphyly of Polyzosteriinae, Archiblattinae and Blattinae is found with high support values and our study will pave the way for a better understanding of the relationship among those subfamilies.
Supplemental Information
Species included in this study, with details of families, references, collecting localities, and GenBank accession numbers
Bayesian inference (BI) tree of the cockroaches based on six genes
BI tree derived from analysis of combined data 12S, 16S, 18S, 28S, COII, and H3 genes. Branch labels are support for our analyses in the following order: bootstrap supports of the maximum-likelihood tree, Bayesian posterior probabilities of the Bayesian tree; asterisks (*) indicate 100% support for a given analysis.
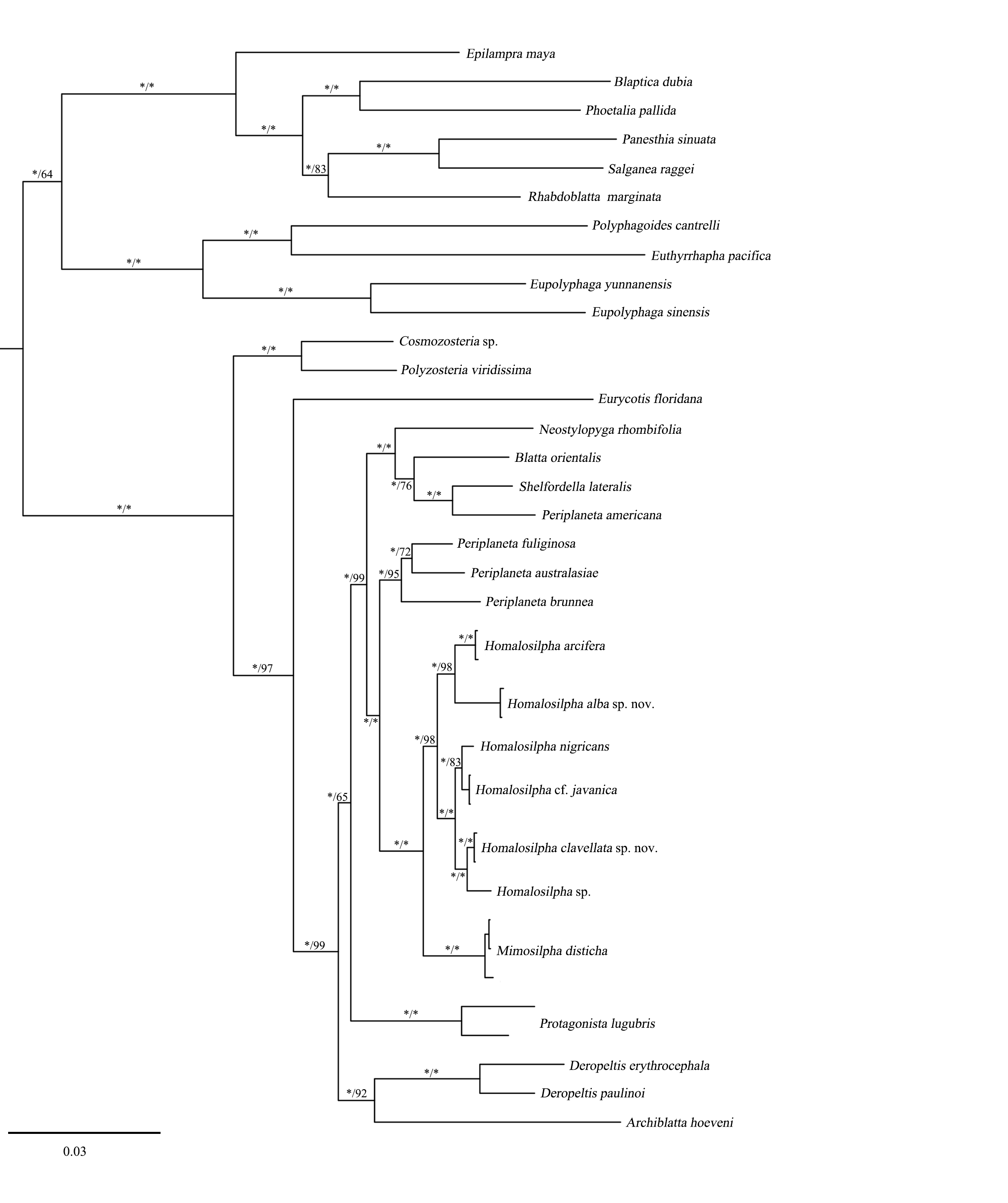{kind=link}
Maximum Likelihood (ML) tree of the cockroaches based on seven genes
ML tree derived from analysis of combined data 12S, 16S, 18S, 28S, COI, COII, and H3 genes. Branch labels are support for our analyses in the following order: bootstrap supports of the maximum-likelihood tree, Bayesian posterior probabilities of the Bayesian tree; asterisks (*) indicate 100% support for a given analysis
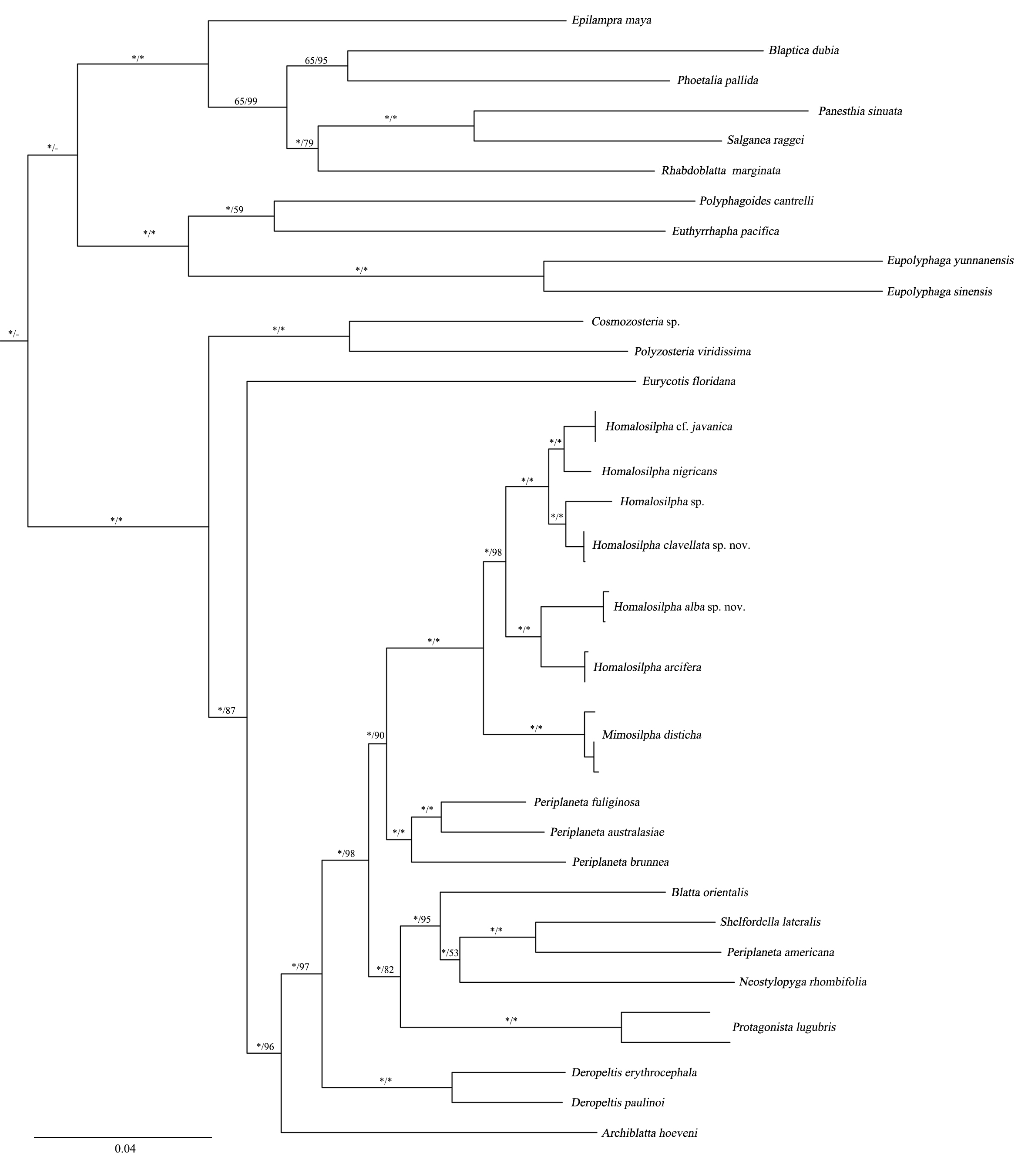{kind=link}
Bayesian inference (BI) tree of the cockroaches based on seven genes
BI tree derived from analysis of combined data 12S, 16S, 18S, 28S, COI, COII, and H3 genes. Branch labels are support for our analyses in the following order: bootstrap supports of the Bayesian tree, maximum-likelihood posterior probabilities of the maximum-likelihood tree; asterisks (*) indicate 100% support for a given analysis
{kind=link}