
Diversity of cultivable protease-producing bacteria and their extracellular proteases associated to scleractinian corals
- Published
- Accepted
- Received
- Academic Editor
- Anastazia Banaszak
- Subject Areas
- Marine Biology, Microbiology
- Keywords
- Diversity, Cultivable, Protease-producing bacteria, Scleractinian corals, Extracellular protease diversity, Hydrolytic ability, Serine proteases, Metalloprotease, Firmicutes, Proteobacteria
- Copyright
- © 2020 Su et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Diversity of cultivable protease-producing bacteria and their extracellular proteases associated to scleractinian corals. PeerJ 8:e9055 https://doi.org/10.7717/peerj.9055
Abstract
Protease-producing bacteria play a vital role in degrading organic nitrogen in marine environments. However, the diversity of the bacteria and extracellular proteases has seldom been addressed, especially in communities of coral reefs. In this study, 136 extracellular protease-producing bacterial strains were isolated from seven genera of scleractinian corals from Luhuitou fringing reef, and their protease types were characterized. The massive coral had more cultivable protease-producing bacteria than branching or foliose corals. The abundance of cultivable protease-producing bacteria reached 106 CFU g−1 of coral. Phylogenetic analysis of 16S rRNA gene sequences revealed that the isolates were assigned to 24 genera, from which 20 corresponded to the phyla Firmicutes and Proteobacteria. Bacillus and Fictibacillus were retrieved from all coral samples. Moreover, Vibrio and Pseudovibrio were most prevalent in massive or foliose coral Platygyra and Montipora. In contrast, 11 genera were each identified in only one isolate. Nearly all the extracellular proteases from the bacteria were serine proteases or metalloproteases; 45.83% of isolates also released cysteine or aspartic proteases. These proteases had different hydrolytic ability against different substrates. This study represents a novel insight on the diversity of cultivable protease-producing bacteria and their extracellular proteases in scleractinian corals.
Introduction
In coral reef ecosystems, nitrogen is as essential as carbon and other nutrients, but it is present at very low concentrations in the water surrounding coral reefs (Suzuki & Casareto, 2011). Abundant particulate organic material (including proteins, amino acids and various biomolecules) and detritus that carry organic nitrogen (OrgN), are the main nitrogen sources in coral reefs (Atkinson, Falter & Hearn, 2001). The imbalance of consumption and production between inorganic and organic nitrogen suggests that OrgN is produced in dissolved forms (dissolved organic nitrogen, DON), which are important components of coral reef metabolism and biogeochemical cycles (Larkum, Kennedy & Muller, 1988; Rädecker et al., 2015). In this cycle, particulate OrgN must be first decomposed into dissolved OrgN and then undergo ammonification, nitrification, and denitrification resulting in formation of nitrogen gas, which will be released into the atmosphere. These processes are mainly performed by bacteria through degradation enzymes (Hunter, Mills & Kostka, 2006; Olson & Lesser, 2013).
A large amount of the dry tissue weight (about 50%) of corals is composed of proteins that exist in many forms and have dozens of functions, such as comprising muscle protein and scaffolds for cytoskeletons, acting as biological catalysts, and giving corals wondrous colorations (Fitzgerald & Szmant, 1997). Organisms absorb organic matter from the environment to assemble tissue. Protease-producing bacteria are considered the main degraders of OrgN in the marine environment because they secrete extracellular proteases that hydrolyze the OrgN into peptides and amino acids that are easily taken up by organisms for subsequent catabolism (Chen et al., 2003; Hui-Lin et al., 2012). Protease-producing bacteria not only enhance protein digestibility and improve the growth of the host but also reduce organic waste in aquaculture (Amin, 2018; Shi et al., 2016). Some bacteria are potential dietary probiotics that promote growth and improve immunity and disease resistance (Amin, 2018; Selim & Reda, 2015).
The diversity of protease-producing bacteria and extracellular proteases from coastal and ocean sediments has been studied (Li et al., 2017; Ming-Yang et al., 2013; Zhou et al., 2009). Protease-producing bacteria have been found into four major phyla, Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes, and are dominated by Pseudoalteromonas, Pseudomonas, and Bacillus genera. Most extracellular proteases secreted by these bacteria are serine proteases and/or metalloproteases isolated from the marine sediments. Many of these proteases are in a relatively low proportion of cysteine proteases (Ming-Yang et al., 2013; Zhang et al., 2015). It is well known that corals harbor abundant, highly biodiverse, and multifunctional prokaryotic communities that play significant roles in nutrient procurement and material transformation to maintain the health of coral reef ecosystems (Blackall, Bryan & Van Oppen, 2015), which population structure is distinct from that of the contiguous sediment (Melissa & Farooq, 2010) and seawater (Shinichi, Woodley & Mónica, 2010). However, there are few reports on bacterial metalloproteases from corals and on coral extracellular proteases that has significant physiological effect on coral symbionts (Anithajothi et al., 2014; Meir et al., 2009; Santos et al., 2011).
A previous study by Zhao et al. (2008) showed that the Luhuitou reef consists of 69 species of hermatypic corals that belong to 24 genera and 13 families (Zhao et al., 2008). It covers an area that has a remarkable amount of biological diversity, including typical coral reefs, many valuable fisheries, and countless microorganisms. However, the functional roles of the microbial symbionts of corals are still poorly understood, and the diversity and functional redundancy of protease-producing bacteria and their extracellular proteases has seldom been addressed. In this paper, seven genera of scleractinian corals from Luhuitou fringing reef were sampled, representing several different skeletal morphology of coral. After isolation and screening, 136 cultivable protease-producing bacterial strains were obtained from these corals, and their diversity was investigated by phylogenetic analysis of 16S rRNA gene sequences. The diversity of bacterial extracellular proteases secreted by these strains was studied using different substrates and inhibitors tests. This study aimed to explore the diversity of protease-producing bacterial communities in corals and characterize their extracellular proteases.
Materials and methods
Sampling and coral identification
Coral samples were collected from the Luhuitou coral reef (109°28′E, 18°13′N), located in the south of Hainan Island, east of Sanya Bay, and west of Luhuitou Peninsula in China. In September 2017, coral fragments (approximately 10 × 10 cm) were collected from seven healthy corals using a hammer and punch at a depth of 2–10 m (Table 1). The collected samples were gently washed with sterile seawater and placed in sterile plastic bags. All samples were stored at low temperatures (0–4 °C) to reduce mucus secreting and immediately transported to the laboratory for isolation of protease-producing bacteria. After removing tissue using an airbrush, the coral skeleton was ready for immediate species identification. All coral samples were identified through ecological and morphological characteristics, according to the book by Veron (2009).
| CORAL GENERA | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| A | L | B | C | F | D | Q |
Total and Rate (%) |
||
| GENERA DISTRIBUTION | |||||||||
| Fimicutes(5) | Bacillus | 5 | 2 | 1 | 5 | 3 | 6 | 22 (16.18%) | |
| Lysinibacillus | 2 | 2 (1.47%) | |||||||
| Paenisporosarcina | 1 | 1 (0.74%) | |||||||
| Fictibacillus | 3 | 1 | 3 | 1 | 5 | 2 | 2 | 17 (12.50%) | |
| Exiguobacterium | 1 | 1 | 2 | 4 (2.94%) | |||||
| Proteobacteria(15) | |||||||||
| Sphingomonas | 1 | 1 (0.74%) | |||||||
| Paracoccus | 1 | 1 (0.74%) | |||||||
| Ruegeria | 1 | 1 (0.74%) | |||||||
| Roseibacterium | 1 | 1 | 2 (1.47%) | ||||||
| Oceanicola | 1 | 1 (0.74%) | |||||||
| Pseudovibrio | 4 | 15 | 1 | 20 (14.71%) | |||||
| Stenotrophomonas | 1 | 1 (0.74%) | |||||||
| Pseudomonas | 1 | 1 | 1 | 3 (2.21%) | |||||
| Erwinia | 1 | 1 (0.74%) | |||||||
| Shewanella | 1 | 1 (0.74%) | |||||||
| Pseudoalteromonas | 1 | 1 (0.74%) | |||||||
| Idiomarina | 1 | 1 (0.74%) | |||||||
| Alteromonas | 1 | 1 | 2 (1.47%) | ||||||
| Microbulbifer | 3 | 3 | 1 | 2 | 9 (6.62%) | ||||
| Vibrio | 6 | 2 | 9 | 2 | 1 | 14 | 34 (25.00%) | ||
| Actinobacteria(3) | |||||||||
| Microbacterium | 2 | 2 | 1 | 1 | 6 (4.41%) | ||||
| Micrococcus | 1 | 1 (0.74%) | |||||||
| Brevibacterium | 1 | 1 (0.74%) | |||||||
| Bacteroidetes(1) | Aquimarina | 2 | 1 | 3 (2.21%) | |||||
|
Total strain number (136) |
11 | 21 | 12 | 33 | 21 | 14 | 24 | 136 | |
Notes:
Pocillopora, Porites, Platygyra, Turbinaria, Faviia, Acropora, Montipora were represented by A, B, C, D, F, L, Q.
Cultivation and screening of protease-producing bacteria
Protease-producing bacteria were cultivated and screened according to a previous study (Li et al., 2017). Briefly, 1 g fresh weight of corals, including tissue, mucus and skeleton (triplicate samples collected from one species were weighed equally before mixing), was diluted in 10 mL sterile sea water and homogenized by vortexing with sterile three mm glass beads for 10 min at a speed setting of 6.0. A portion of the resulting homogenates was pooled and filtered at 0.22 µm to remove bacteria and simulate coral environment for fostering more bacteria on plate. 100 mL eluate was then added to media cooled to 50 °C, which was composed of 1/10 2216E (tenfold dilution of 2216E) and 1.5% (w/v) agar powder in 1 L seawater at pH 8.0, and supplemented with 1% (w/v) casein. The preparation and addition of coral tissue homogenates was performed according to Rebecca et al. with minor modifications (Certner & Vollmer, 2015). Aliquots of 100 µL of the homogenates were serially diluted (10−1–10−6) and separately spread on screening plates with three replicates. All plates for bacterial screening were incubated at 25 °C until a clear hydrolytic zone formed around the colonies. Positive colonies were selected and purified by repeated streaking until uniform or pure colonies could be detected on the same plate. Morphological characteristics (e.g., colony color and zone, hydrolytic zone) were analyzed. Pure cultures were preserved in 25% (v/v) glycerol at −80 °C for further use.
Molecular identification of the bacterial strains
After incubation in liquid screening medium without agar, genomic DNA was extracted from the bacterial strains using the Hipure bacterial DNA kit (Magen, China). The 16S rRNA gene was amplified using the universal primer pair 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′). PCR amplifications were performed in a Mastercycler pro (Eppendorf, Germany) in a final volume of 30 µL, containing 1.2 µL (10 µM) of each primer, 0.1 µL of template DNA (approximately 60 ng), and 25 µL Easy Taq super Mix (Transgen Biotech, Beijing). The PCR conditions were as follows: 95 °C for 4 min; 30 cycles at 94 °C for 45 s, 54 °C for 45 s, 72 °C for 120 s; followed by 72 °C for 10 min. PCR products were sequenced by Sangon Biotech (Shanghai, China). Identification of bacterial strains was carried out by comparison with available 16S rRNA gene sequences in GenBank using BLASTn approach to determine their closest relatives and approximate phylogenetic affiliation (Tomova et al., 2013). Samples with the same 16S rRNA gene sequence (or with only one base difference) were categorized as the same strain. A phylogenetic gene tree was generated from 16S rRNA using the neighbor-joining method with MEGA package version 7.0 (Kumar, Stecher & Tamura, 2016). The 16S rRNA gene sequences of this study were deposited in GenBank database under accession numbers MK617631 –MK617766.
Hydrolysis of casein, elastin, and gelatin by bacterial extracellular proteases
Three solid basic media (2216E) were prepared, and supplemented with 1% (w/v) casein, 0.5% (w/v) gelatin, or 0.5% (w/v) elastin powder, respectively (Li et al., 2017). Protease-producing isolates were inoculated with sterilized toothpicks on plates and then incubated at 25 °C for 3 days. The ratio of the hydrolytic zone diameter to the colony diameter (hydrolytic zone/colony, H/C) was calculated to indicate the enzymatic activity for each substrate.
Effect of protease inhibitors on protease activity
The inhibitor assay was performed as described in a previous paper (Li et al., 2017). Each protease-producing strain was cultivated in the screening medium without agar at 25 °C with shaking (170 rpm) for 3 d. The culture supernatant was obtained by centrifugation at 14,000×g at 4 °C for 5 min. The supernatant was diluted to maintain OD660 between 0.4 and 0.8 in 20 mM Tris–HCl (pH 8.0), and incubated at 15 °C for 20 min in the presence of different inhibitors including phenylmethylsulfonyl fluoride (1.0 mM, PMSF, Aladdin), 1,10-Phenanthroline (1.0 mM, OP, Aladdin), E-64 (0.1 mM, Sigma), and pepstatin A (0.1 mM, Aladdin). The residual activity of each sample was measured in triplicates using samples without inhibitors as controls (100%), and the relative activity (%) of samples was calculated.
Results
Sample collection and coral identification
The 17 scleractinian coral samples were classified into seven genera, corresponding to branching (Pocillopora and Acropora), massive (Porites, Platygyra, and Favia), and foliose (Turbinaria and Montipora) corals according to skeletal morphology, and used for incubation and screening of protease-producing bacteria.
Isolation and quantification of protease-producing bacteria from corals
After incubation at 25 °C for 12–15 days, approximately 20–100 colonies of various color, size, and morphology, appeared in samples diluted to 10−3–10−4. Manual counts indicated that the most abundant bacteria reached 107CFU g−1 in Platygyra coral samples. Nearly 20% of the colonies had hydrolytic zones. The massive coral had more cultivable protease-producing bacteria than branching or foliose corals. Approximately 200 colonies were obtained for further identification and characterization.
Phylogenetic Diversity of Protease-Producing Bacteria Isolated from Corals
The 16S rRNA gene of the 136 isolates, which contained nearly 1,400 bp, were amplified and partially sequenced. Samples with the same 16S rRNA gene sequence (or with only one base difference) were categorized as the same strain, resulting in a total of 76 different strains. Phylogenetic affiliation was assigned based on the sequences of the 16S rRNA genes. As shown in Table 1, the 136 strains were assigned to 24 genera. Most of the bacterial isolates were assigned to 20 genera within the phyla Firmicutes and Proteobacteria, except for isolates (sC2, sC26, sJ58) from Platygyra and Faviidae belonging to Aquimarina in the phylum Bacteroidetes, and eight isolates belonging to Microbacterium (sA9, sA61, sD33, sF16b, yB24b, yB18), Micrococcus (sF19) and Brevibacterium (yF15) in the phylum Actinobacteria. The Firmicutes included Bacillus, Lysinibacillus, Paenisporosarcina, Fictibacillus, and Exiguobacterium. The Proteobacteria were mainly affiliated with the phyla Alphaproteobacteria and Gammaproteobacteria, including Sphingomonas, Paracoccus, Ruegeria, Roseibacterium, Oceanicola, Enterobacteriaceae, Pseudovibrio, and Stenotrophomonas, Pseudomonas, Erwinia, Shewanella, Pseudoalteromonas, Idiomarina, Alteromonas, Microbulbifer, Vibrio, and Lucibacterium. Four of the most predominant genera were Bacillus (16.18%) and Fictibacillus (12.50%) in the phylum of Firmicutes that were present in all coral samples, Pseudovibrio (14.71%) and Vibrio (25.00%) in the phylum of Proteobacteria that hat were present in almost all samples. The largest number of protease-producing bacteria was in Platygyra and Montipora which corresponded massive or foliose corals. Sphingomonas, Paracoccus, Ruegeria, Stenotrophomonas, Oceanicola, Shewanella, Pseudoalteromonas, Idiomarina, Alteromonas, Micrococcus and Brevibacterium were represented only by one isolate each, 11 isolates in total, corresponding to 8.1% of the total. The rest of genera constituted 1.4% to 7.3% of all strains. These results revealed that the predominant bacteria in corals of various genera were different. In massive coral (Platygyra), Pseudovibrio (42.80%) and Vibrio (25.00%) were the predominant protease-producing bacteria. In foliose coral (Montipora), Bacillus (24.00%) and Vibrio (56.00%) were the predominant protease-producing bacteria.
All related sequences from this study and reference sequences from the GenBank database were used to construct a neighbor-joining tree (Fig. 1 and 2). Approximately thirty one strains from Poritidae, Platygyra, and Montipora formed Branch 1 in Fig. 1, with 99% sequence similarity with the type strain Vibrio parahaemolyticus NBRC 12711. Nineteen Pseudovibrio strains (Branch 3 from three corals) (Fig. 1) clustered with Pseudovibrio denitrificans DSM 17465 with 99–100% similarity. Thirteen Bacillus strains (Branch 6 from four corals) were closest to Bacillus oceanisediminis H2T (99.2–100%). The phylogenetic relationships of the other strains are shown in Fig. 1 and 2. The results suggest that the diversity and composition of protease-producing bacterial communities was various in different genera of coral, and the highest diversity and evenness in community were present in Acroporidae, Faviidae and Platygyra (Table 1).
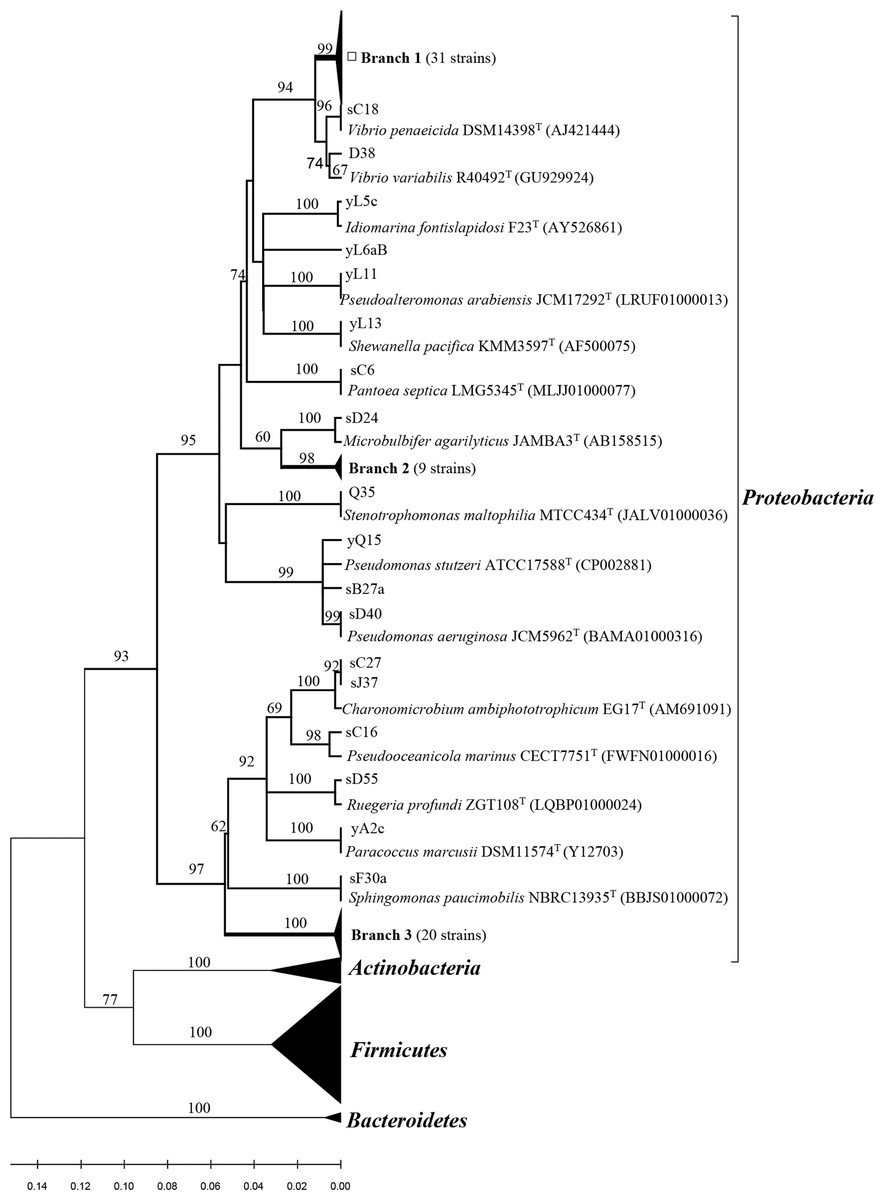
Figure 1: Phylogenetic tree of the protease-producing Proteobacteria isolated from Scleractinian corals based on 16S rRNA gene sequences.
Phylogenetic tree of the protease-producing Proteobacteria isolated from Scleractinian corals based on 16S rRNA gene sequences. The tree was constructed by neighbor-joining method using MEGA package version 7.0. Only bootstrap values greater than 50% are presented in the nodes. The scale bar represents 2% nucleotide substitution. Branch 1 indicates 31 Vibrio strains similar to Vibrio owensii LMG 25443T (JPRD01000038). Branch 2 indicates nine Microbulbifer strains similar to Microbulbifer variabilis Ni2088T (AB167354). Branch 3 indicates 20 Pseudovibrio strains similar to Pseudovibrio denitrificans DSM17465T (jgi.1107980).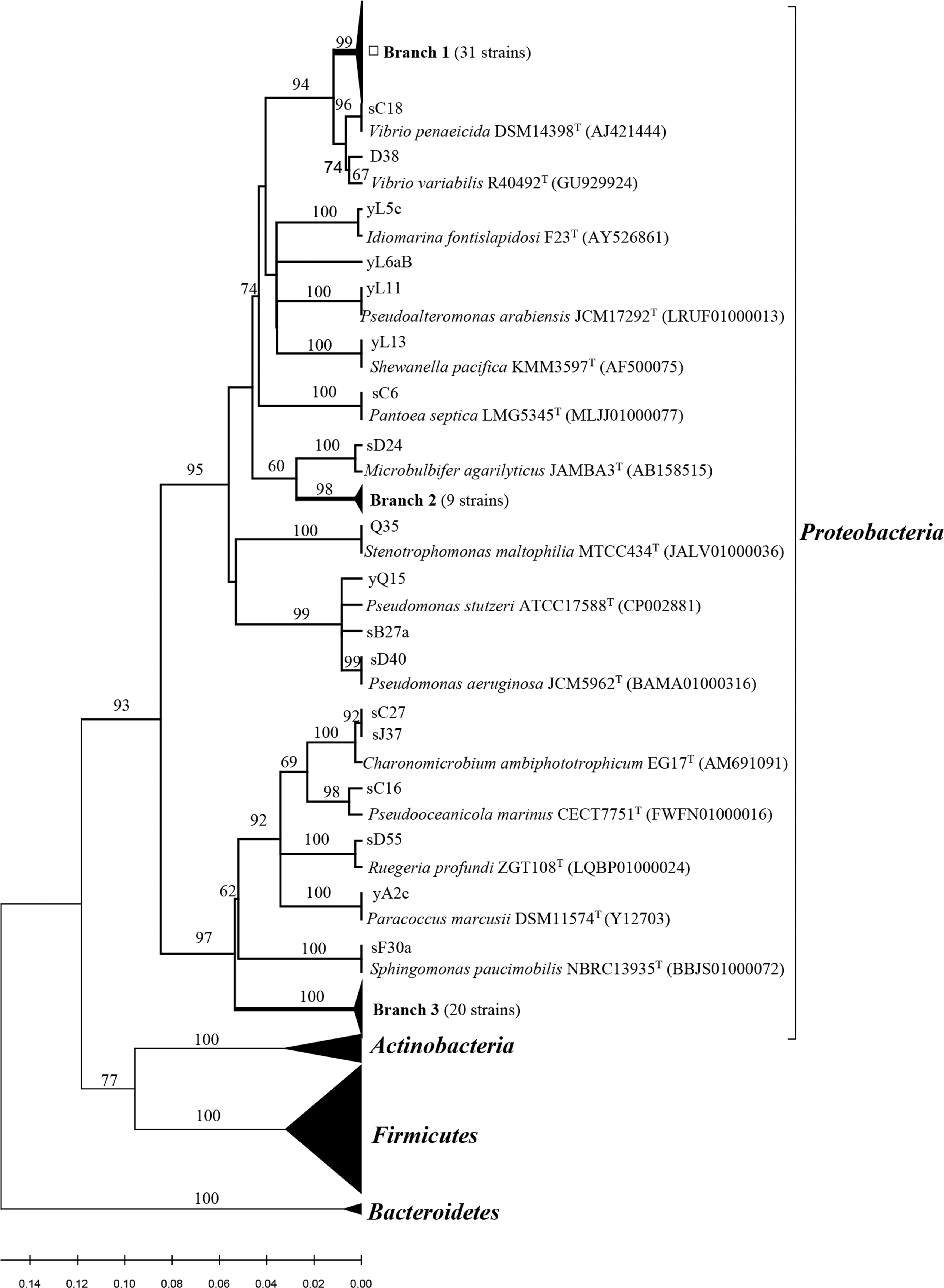{kind=link}

Figure 2: Phylogenetic tree of the protease-producing Actinobacteria, Firmicutes and Bacteroidetes isolated from Scleractinian corals based on 16S rRNA gene sequences.
The tree was constructed by neighbor-joining method using MEGA package version 7.0. Only bootstrap values greater than 50% are presented in the nodes. The scale bar represents 2% nucleotide substitution. Branch 4 indicates four Micrococcus strains similar to Micrococcus aloeverae AE-6 T (KF524364). Branch 5 indicates 13 Fictibacillus strains similar to Fictibacillus nanhaiensis JSM082006 T (GU477780). Branch 6 indicates 13 Bacillus strains similar to Bacillus megaterium NBRC 15308 T (JJMH01000057).{kind=link}
Diversity of the extracellular proteases produced by the bacteria
The diversity of the bacterial extracellular proteases in corals was investigated using substrate specificity assays that measured the H/C ratio on plates containing casein, gelatin, or elastin (Table 2). Of the 76 isolates, 48 produced enough protease for enzymatic assays. The extracellular proteases generated by these strains produced obvious hydrolytic zones on casein and gelatin media. These isolates were screened from medium containing casein and gelatin; however, they showed very different hydrolytic abilities against casein and gelatin. The extracellular proteases from isolates sC18 and sL3 belonging to Vibrio, isolates sC1, sD13, and sD19 belonging to Microbulbifer, isolate yL5c belonging to Idiomarina, and isolates sC2 and yF15 belonging to Brevibacterium had high caseinolytic activity with H/C ratios over 4.0. The extracellular proteases from Vibrio (sL3), Paracoccus (yA2c), Brevibacterium (sC2), Bacillus (yF12B, sA10, sJ2 and yQ16) showed high gelatinolytic activity with H/C ratios over 6.0. However, only 24 isolates (50.0% of the total isolates) were capable of hydrolyzing elastin. All elastinolytic isolates produced proteases with various levels of activity against all three proteins, for example, bacillus (yQ16). Although the isolates belong to the same genus, they showed different levels of degradation of elastin. Different substrate specificities reflect that these extracellular proteases may belong to different kinds of metalloproteases or serine proteases.
In addition, the diversity of the bacterial extracellular proteases in corals was investigated using protease inhibitors such as PMSF (serine protease inhibitor), OP (metalloprotease inhibitor), E-64 (cysteine protease inhibitor), and pepstatin A (aspartic protease inhibitor). Among the 78 strains, only 48 secreted enough extracellular proteases for inhibition assays. The activities of most proteases were inhibited by PMSF ranging from 23.10% to 98.56%, indicating that these extracellular proteases may be serine proteases in different proportions. The protease activities of 14 strains were inhibited more than 90% by PMSF, which indicated that they were serine proteases. The protease activities of 41 isolates were inhibited by OP with an efficiency ranging from 24.58% to 92.47%. There was slight inhibition (2.48%–12.15%) in eight isolates, while no inhibitory effect was observed in seven isolates. This indicates that most isolates tested in corals produced metalloproteases. Forty-eight (83.33%) isolates were inhibited by both PMSF and OP at different levels, suggesting that the majority of isolates produced serine proteases or metalloproteases. E-64 showed less than 30% inhibitory effect on the proteolytic activities of all proteases, and pepstatin A also showed light inhibition (1.36%–27.77%) on 35 isolates, except on sA10 and sC26. These results indicate that 45.83% of isolates produced cysteine and/or aspartic proteases. Different inhibition levels indicate that nearly all the extracellular proteases from the bacteria associated to corals are serine proteases and/or metalloproteases, and half of the isolates produced cysteine proteases or/and aspartic proteases.
| Genera | Strains | H/C ratioa | Inhaibition ratiob(%) | |||||
|---|---|---|---|---|---|---|---|---|
| Casein | Gelatin | Elastin |
PMSF (1 mM) |
OP (1 mM) |
E64 (0.1 mM) |
P-A (0.1 mM) |
||
| Vibrio | sC18 | 4.20 | 5.16 | 0 | 54.57 | 49.26 | 26.43 | 19.43 |
| sC57 | 2.30 | 2.41 | 4.00 | 81.86 | 6.60 | 27.74 | 18.21 | |
| D38 | 3.81 | 4.10 | 0 | ND | 64.20 | ND | ND | |
| sL1 | 3.90 | 1.23 | 0 | 97.68 | 86.24 | 29.02 | 0 | |
| sL3 | 5.10 | 6.09 | 0 | 59.55 | 60.42 | 4.82 | 3.88 | |
| sQ3 | 1.88 | 3.56 | 1.41 | 91.09 | 87.88 | 2.67 | −3.66 | |
| sQ48 | 2.16 | 3.18 | 1.53 | 98.56 | 81.12 | 10.25 | 16.02 | |
| Idiomarina | yL5c | 5.13 | 6.17 | 0 | 84.56 | 46.45 | 0 | 0 |
| Pseudoalteromonas | yL11 | 4.73 | 4.02 | 1.67 | 61.22 | 16.42 | 0 | 0 |
| Shwanella | yL13 | 2.86 | 2.55 | 1.54 | 88.03 | −1.92 | 5.76 | 8.23 |
| Pantoea | sC6 | 2.16 | 2.02 | 0 | 65.2 | 92.47 | 6.12 | −4.00 |
| Microbulbifer | sC1 | 4.1 | 4.22 | 0 | 93.01 | 67.83 | 10.53 | 23.32 |
| sC9 | 3.78 | 2.16 | 0 | 91.78 | 75.26 | 9.25 | −2.973 | |
| sC10 | 3.66 | 2.45 | 0 | 86.5 | 0 | 9.32 | 2.70 | |
| sD13 | 4.52 | 3.21 | 0 | 92.87 | 2.78 | 14.38 | 15.88 | |
| sD19 | 4.62 | 3.20 | 0 | 80.04 | 0 | 0 | 1.72 | |
| sL30 | 3.71 | 4.11 | 0 | 74.35 | −3.52 | 6.33 | 8.76 | |
| Strenotrophomonas | Q35 | 3.62 | 2.75 | 1.63 | 92.52 | 83.72 | 0.90 | 20.68 |
| Pseudomonas | sB27a | 2.47 | 2.04 | 3.75 | 66.11 | 30.12 | −6.12 | 0 |
| sD40 | 3.20 | 2.81 | 2.36 | 91.30 | 82.74 | 0 | −15.86 | |
| yQ15 | 4.00 | 3.42 | 1.26 | 92.50 | 78.14 | 1.20 | 6.46 | |
| Oceanicola | sC16 | 3.60 | 3.25 | 1.58 | 63.14 | 51.51 | 17.27 | 27.77 |
| Ruegeria | sD55 | 3.02 | 3.56 | 2.36 | 53.03 | 57.04 | 12.23 | 24.30 |
| Paracoccus | yA2c | 3.65 | 7.50 | 3.24 | 40.32 | 25.52 | 1.21 | 1.36 |
| Sphingomonas | sF30a | 3.26 | 3.12 | 2.77 | 62.79 | 56.68 | 14.31 | 14.73 |
| Pseudovibrio | sC3 | 2.30 | 2.35 | 0 | 84.73 | 73.26 | 13.03 | 9.81 |
| sC13 | 2.36 | 3.20 | 0 | 52.80 | 62.76 | 14.56 | 5.89 | |
| sC15 | 2.56 | 2.31 | 0 | 65.01 | 47.92 | 15.76 | 3.58 | |
| sL47 | 2.56 | 1.28 | 0 | 54.42 | 37.34 | 19.95 | 2.48 | |
| yF15 | 4.00 | 3.65 | 0 | 82.75 | 37.91 | 2.23 | 4.30 | |
| Micrococcus | sF19 | 1.76 | 2.15 | 0 | 88.26 | 8.24 | 16.75 | 17.67 |
| Fictibacillus | sD46 | 1.62 | 2.81 | 2.45 | 90.23 | 56.42 | 28.56 | 8.48 |
| sJ27 | 2.34 | 1.74 | 0 | 87.12 | 50.14 | 12.42 | 16.42 | |
| yQ18 | 1.88 | 2.01 | 2.01 | 84.12 | 40.13 | 8.45 | 3.88 | |
| Lysinibacillus | B21 | 3.21 | 4.21 | 2.54 | 41.60 | 45.25 | 2.47 | 8.29 |
| B24 | 3.02 | 4.05 | 3.21 | 65.21 | 42.00 | −12.10 | 8.14 | |
| Bacillus | sA10 | 1.64 | 5.21 | 0 | 92.85 | 2.48 | 15.88 | 33.29 |
| sA50 | 2.32 | 4.00 | 2.65 | 85.30 | 76.40 | 24.59 | 25.39 | |
| yF12B | 2.32 | 5.52 | 2.45 | 23.10 | 65.01 | −1.25 | 6.24 | |
| sJ2 | 2.15 | 5.22 | 0 | 85.09 | 44.68 | 15.51 | 12.44 | |
| sJ36 | 1.33 | 1.42 | 0 | 90.20 | −5.84 | 0 | 5.05 | |
| yL8 | 2.17 | 4.32 | 3.65 | 89.32 | 5.03 | 13.02 | 0 | |
| yL12 | 2.36 | 2.30 | 1.96 | 86.20 | 44.52 | 12.34 | 0 | |
| yQ16 | 3.02 | 5.25 | 5.63 | 56.56 | 43.22 | 0 | 0 | |
| Aquimarina | sC2 | 3.60 | 4.32 | 0 | 90.00 | 12.15 | 12.34 | 35.02 |
| sC26 | 2.46 | 3.30 | 2.26 | 82.07 | 0 | 5.76 | 8.23 | |
| sJ58 | 3.01 | 3.65 | 0 | 77.62 | 10.31 | 23.72 | 11.44 | |
Notes:
- PMSF
-
phenylmethylsulfonyl fluoride
- OP 1
-
10-phenanthroline
- P-A
-
pepstatin A
Pocillopora, Porites, Platygyra, Turbinaria, Faviia, Acropora, Montipora were represented by A, B, C, D, F, L, Q.
Discussion
In a previous study, the numbers of bacteria in mucus and tissue samples were detected by SYBR gold staining, and the cultivable population on Marine Agar was between 105 and 106/cm2 of coral surface (Koren & Rosenberg, 2006). Our report focused on protease-producing bacteria in corals. The highest abundance of cultured bacteria reached 107 CFU g−1 in some coral samples, which is higher than that in previous reports. Only 20% of the colonies had detectable protease activity, which corresponds to the community of protease-producing bacteria isolated from the South China Sea (106 CFU g−1) (Zhou et al., 2009), this is more than in sub-Antarctic sediments (105 CFU g−1) in Jiaozhou Bay (Zhang et al., 2015), and in Laizhou Bay (104 CFU g-1) (Li et al., 2017). And protease-producing bacteria can be found in all coral samples, this result indicates that a sizable population of protease-producing bacteria exist in corals.
Most reports estimate that important sources of DON come from bacteria and other organisms associated with corals (Ferrier, 1991; Grover et al., 2003; Renaud et al., 2008). Coral reefs are considered “oligotrophic areas” based on their low nitrate concentrations, which means that the degradation rate of organic matter in dissolved and particulate forms must be fast enough (Christian et al., 2004; Muscatine & Porter, 1977). It is not known how much DON is produced by bacteria, and few reports refer to the degradation of DON, which results in a failure to characterize microbial and to address the adaptive physiology and functional attributes of bacteria. In this study, we investigated the protease-producing bacterial communities and the diversity of extracellular proteases to reveal physiology and functional attributes of bacteria in scleractinian corals.
The bacterial phyla associated with corals were identified with the development of culture-independent high-throughput DNA sequencing methods. Overall, Proteobacteria, Bacteroidetes, Firmicutes, Cyanobacteria, and Chloroflexi were the most abundant population with an abundance of over 90%, which is similar to other coral species (Jie et al., 2013; Liang et al., 2017). It have been found that the core microbiomes of different corals were reliably dominated by the phylum Proteobacteria, specifically Gammaproteobacteria and Alphaproteobacteria, and secondarily by Bacteroidetes and Actinobacteria (Chu & Vollmer, 2016). Similarly, in our study, Proteobacteria was the predominant cultivable protease-producing bacterial phyla (58.1%), represented by the genera Stenotrophomonas, Pseudomonas, Erwinia, Shewanella, Pseudoalteromonas, Idiomarina, Alteromonas, Microbulbifer, Vibrio, and Lucibacterium, followed by Gammaproteobacteria with an abundance of 39.0% (Table 1). Vibrio was the predominant genus, accounting for 23.5% of the total strains. This is different from the predominant protease-producing bacteria isolated from the sediments of the South China Sea, which belong to Alteromonas and Pseudoalteromonas (Zhou et al., 2009). Firmicutes was the second most abundant phylum (33.8%), with Bacillus as the dominant genus that was found in all coral samples (Table 1). This was consistent with a paper that described protease-producing bacteria and their extracellular proteases in the sediments of Laizhou Bay by Li et al. (2017) which illustrates that Firmicutes or Bacillus might play important roles in biodegradation in corals. Unlike these studies, ours is the first study reporting that numerous isolates belonging to Fictibacillus and Pseudovibrio possess protease-producing abilities.
There were differences in the diversity of protease-producing bacteria between species, although only a small portion of bacteria was screened. For example, the most abundant and diverse protease-producing bacteria was found in Platygyra sp. and Faviidae sp., both belonging to massive corals. This result is consistent with previous findings suggesting that abundant bacterial species may help corals to respond to abnormal changes of temperature (Liang et al., 2017). Although only 20 protease-producing isolates were obtained from Acroporidae sp. (a fast growing branching species) (Edwards & Clark, 1999), they have the second highest diversity.
Because of their broad biochemical diversity, microbes are an excellent source of proteases. Some proteolytic bacteria have been reported to be associated not only with fresh water but marine fish processing waste as well. The protease produced by Bacillus proteolyticus CFR3001 lyses the cells of pathogenic bacteria such as Escherichia coli, Listeria monocytogenes, Bacillus cereus, and Yersinia enterocolitica (Bhaskar et al., 2007). The protease-producing Bacillus subtilis E20 also shows great potential to increase the growth performance of Litopenaeus vannamei and increases disease resistance against the pathogen Vibrio alginolyticus (Liu et al., 2009; Tseng et al., 2009). To date, no study had demonstrated protease production by probiotic bacteria in corals; however, in the present study, we found that many Bacillus isolates from corals that displayed high proteolytic activity against casein, gelatin and/or elastin were widespread in all coral samples, and may play an important role in coral health. The analysis of protease-producing bacterial diversity in this study suggests that bacterial species may benefit coral growth, transplant, and regeneration.
Most isolates produced serine and/or metalloproteases, which was similar to protease-producing bacteria isolated from coastal sediments (Li et al., 2017; Ming-Yang et al., 2013; Zhou et al., 2009). Numerous studies have demonstrated that the zinc-metalloproteases are present in Vibrio pathogenic strains, for example, the zinc-metalloprotease of Vibrio coralliilyticus causes rapid photosystem II inactivation of Symbiodinium endosymbionts, leading to Pocillopora damicornis bleach (Yael, Maya & Eugene, 2003). V. coralliilyticus strain P1 has a diverse zinc-metalloproteases that possibly enables it to be an efficient coral pathogen (De et al., 2011). In this study, 32 isolates belonging to Vibrionaceae, but none belonging to V. coralliilyticus were found, suggesting that metalloproteases are widespread in protease-producing bacteria associated with corals. However, nearly 45.83% of isolates were also inhibited by E-64, which was different from the protease-producing bacteria isolated from coastal sediments (Ming-Yang et al., 2013; Zhou et al., 2009). Twenty-five isolates showed different levels of degradation activity against elastin, which is the most insoluble and difficult to degrade of the three protein substrates tested due to its high molecular weight protein polymer. Furthermore, the Bacillus isolates from corals displayed higher proteolytic activities against casein, gelatin and/or elastin than those in South China Sea (Zhou et al., 2009) or than those in the sediments of Laizhou Bay (Li et al., 2017). Strains with multiple enzyme types and different substrates specificities were observed in corals, which may allow the bacterial community to more effectively and rapidly hydrolyze complex OrgN sources for coral reefs.
Conclusion
This paper is the first systematic study to investigate the phylogenetic diversity of cultivable protease-producing bacteria associated with scleractinian corals and their extracellular protease types. Fictibacillus, Bacillus, Vibrio, and Pseudovibrio were the most retrieved genera, and Fictibacillus and Pseudovibrio that possess proteolytic activity were isolated for a first time from Luhuitou fringing coral reefs. In addition, some bacterial genera (Bacillus, Fictibacillus and Vibrio) were found in all samples but were most dominant in Platygyra and Montipora, which are massive or foliose coral. Forty-eight isolates showed different levels of degradation activities against casein and gelatin. Twenty-five of them simultaneously displayed hydrolytic activity against casein, gelatin, and elastin. Most protease-producing bacteria associated to scleractinian corals produced metalloproteases and/or serine proteases, and 22 isolates also released cysteine or aspartic proteases.
Supplemental Information
Bacteral culture on CFCF plate
A large number of coral symbiotic bacteria with different colors grown on CFCF plates
{kind=link}
Screening of protase-producing bacteria on plate
Cultivable protease-producing bacteria displayed a clear hydrolytic zone around the colonies.
{kind=link}