
CRISPR screening identifies M1AP as a new MYC regulator with a promoter-reporter system
- Published
- Accepted
- Received
- Academic Editor
- Francois van der Westhuizen
- Subject Areas
- Cell Biology, Molecular Biology, Oncology
- Keywords
- CRISPR library screening, MYC
- Copyright
- © 2020 Yamamoto et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. CRISPR screening identifies M1AP as a new MYC regulator with a promoter-reporter system. PeerJ 8:e9046 https://doi.org/10.7717/peerj.9046
Abstract
Background
MYC is one of the proto-oncogenes contributing to tumorigenesis in many human cancers. Although the mechanism of MYC regulation is still not fully understood, learning about the comprehensive mechanism controlling the transcriptional activity of MYC will lead to therapeutic targets. The CRISPR/Cas9 library system is a simple and powerful screening technique. This study aims to identify new transcriptional upstream activators of MYC using the CRISPR activation library with new promoter-reporter systems.
Methods and Results
The MYC promoter-reporter system was developed with a photoconvertible fluorescent protein, Dendra2, and named “pMYC-promoter-Dendra2.” This MYC promoter-reporter system was designed to harbor a proximal MYC promoter at (3.1 kb). Both the CRISPR activation library and pMYC-promoter-Dendra2 were induced to HEK 293T cells, and Dendra2-positive cells, that are supposed that MYC should be upregulated, were collected individually by a cell sorter. Among the 169 cells collected, 12 clones were successfully established. Then, pMYC-promoter-Dendra2 was transfected again into these 12 clones, and two of 12 clones showed Dendra2 positivity. In this procedure, the cells with non-specific autofluorescence were correctly distinguished by utilizing the photoswitchable character of Dendra2. Using extracted genomic DNA of these two Dendra2 positive clones, polymerase chain reaction (PCR) was performed to amplify the guide RNA (gRNA) containing region, which was introduced by the CRISPR activation library. Eventually, PLEKHO2, MICU, MBTPS1, and M1AP were identified, and these gRNAs were transfected individually into HEK 293T cells again using the CRISPR activation system. Only M1AP gRNA transfected cells showed Dendra2-positive fluorescence. Then, the overexpression vector for M1AP with a doxycycline-inducible vector confirmed that M1AP induced high MYC expression by real-time quantitative PCR and western blot. Furthermore, the dual-luciferase assay showed a significant increase of promoter activity, and MYC mRNA was higher in M1AP- overexpressing cells. M1AP is highly expressed in several cancers, though, a positive correlation between M1AP and MYC was observed only in human acute myeloid leukemia.
Conclusion
The present study confirmed that the experimental method using the CRISPR library technology functions effectively for the identification of molecules that activate endogenous MYC. This method will help elucidate the regulatory mechanism of MYC expression, as well as supporting further drug research against malignant tumors.
Introduction
MYC is one of the most well-known proto-oncogenes, contributing to tumorigenesis in many human cancers. It can function as a transcriptional factor in both cancerous and non-cancerous cells, and it is involved in cell growth, proliferation, metabolism, and transformation (Dang, 2012). The Wnt- β-catenin signaling pathway is one of the most well-described pathways involved in the regulation of MYC. Due to mutations in the adenomatous polyposis coli (APC) gene or abnormalities in glycogen synthase kinase-3 β, Wnt-signal is stimulated and the excessive accumulation of β-catenin occurs. Then, active β-catenin moves into the nucleus and binds to T cell factor (TCF), followed by inducing MYC expression (Liu et al., 2002; Novak & Dedhar, 1999). Mitogen-activated protein kinase (MAPK), which is a serine/threonine kinase, and its subfamily extracellular signal-regulated kinase (ERK), regulate MYC transcriptional activity by inducing phosphorylation of MYC (MAPK/ERK signaling pathway). Furthermore, as non-coding RNA, microRNAs like miR-34 (Kong et al., 2008) and let-7 (Sampson et al., 2007) suppress MYC expression by binding to the 3′-untranslated region of MYC mRNA. As for long noncoding RNAs, PVT1 regulates the expression of MYC by inhibiting the degradation of MYC protein (Tseng et al., 2014). Thus, it has been reported that various regulatory mechanisms function in a complex fashion regarding MYC expression, although other molecules and elements like crosstalk between each pathway are not fully understood.
Concerning the high MYC expression rate of multiple malignant tumors, elucidating the mechanism controlling the transcriptional activity of MYC is a crucial issue because it will lead to therapeutic targets. Using the CRISPR/Cas9 library system, with its collection of more than 70,290 different guide RNAs (gRNAs), enables us to identify molecules that relate knockout or activation of endogenous genes. To date, the CRISPR library has been utilized for various studies. Shalem et al. (2014) identified genes whose loss is involved in resistance to vemurafenib by introducing the knockout library into a human melanoma cell line followed by treatment with BRAF inhibitor vemurafenib. In contrast, Konermann et al. (2015) identified molecules that exhibit resistance to PLX-4720 by introducing an activation library. In this way, the CRISPR library has been used to identify the certain genes involved in drug sensitivity or resistance, toxin resistance, and hypoxia response (Konermann et al., 2015; Gilbert et al., 2014; Kampmann, 2018; Kurata et al., 2018a; Kurata et al., 2018b; Shalem et al., 2014; Wang et al., 2014; Koike-Yusa et al., 2014; Doench et al., 2016; Jain et al., 2016); however, a screening of molecules that act on the promoter region of a specific gene and control transcriptional activity has not yet been tested. In addition, autofluorescence is highly problematic for collecting fluorescent cells in the screening. Therefore, a photoconversion protein, Dendra2, was used in this system. Dendra2 is derived from the octocoral Dendronephthya sp. T. and can change color with UV-violet light (405 nm laser). This study aims to utilize the CRISPR activation library for the detection of new transcriptional activators of MYC. For this purpose, we developed an original tool for evaluating MYC activity and establishing screening methods.
Materials & Methods
MYC promoter-reporter system and lentivirus plasmids
The Polymerase chain reaction (PCR) primers were designed across the proximal MYC promoter at −3.1 kb from the MYC transcriptional start site (TSS), and PCR was performed with KOD-FX (Toyobo). The primers were 5′-TTAGCTAGCGAGGGTT- TTCTTTGAGGGGC-3′ (forward) and 5′-AACGGATCCCGGAGATTAGCGAGAGAGGA-3′ (reverse).
A plasmid of Dendra2 (pDendra2) was purchased from Takara-Clontech. The cytomegalovirus immediate early promoter (PCMVIE) was removed with restriction enzymes (Afl III and Nhe I), and the multi-cloning site was replaced with Nhe I and BamH I. Amplified fragments of −3.1 kb of the MYC promoter region with Nhe I and BamH I restriction enzyme sites were integrated into pDendra2.The new reporter system with Dendra2 and the −3.1 kb MYC promoter region (pMYC-promoter-Dendra2) was used for further experiments. The double-stranded DNA fragments of M1AP with attB sites were purchased from gBlocks® (IDT) and generated into the pENTR221 vector using Gateway® BP clonase. Then they were transferred to the TripZ-TRE-DEST-IRES-GFP-EF1A-rtTA vector (Kurata et al., 2016) using Gateway® LR clonase.
Cell culture and SAM library screening
Human embryonic kidney cells (HEK 293T) were kindly provided by Dr. David Laraespada, Masonic Cancer Center, University of Minnesota, MN, USA. and maintained in Dulbecco’s modified Eagle’s medium (DMEM, Fuji Film) with 10% fetal bovine serum and penicillin/streptomycin with 5% CO2 at 37 °C . To transduce the lentivirus, HEK 293T cells (5 × 105 cells/well) were seeded in a six-well cell culture plate one day before the transfection. The next day, 3 µg of lentivirus plasmid was transfected with 1 µg of pMDG and 2 µg of pCMV using Lipofectamine™ 3000 Reagent (Invitrogen) according to previous work (Kurata et al., 2016). Twelve hours after transfection, the medium was changed to fresh DMEM. The virus supernatant was harvested at 48 h after transfection, then filtered with Millex™ -HP 0.45 µm (Millipore). The target HEK 293T cells (5 × 105 cells/well) were seeded in a six-well plate one day prior to transduction and were transduced with this lentivirus supernatant with 5 µg/ml polybrene (Sigma). The used lentiviral plasmids were lentiMPHv2 (Addgene, #89308), lentidCAS-VP64_blast (Addgene, #61425), and lentiSAMv2 (Addgene, #61597). At first, lentiMPHv2 was transduced into HEK 293T and treated with Hygromycin B for 2 weeks. Then, lentidCAS9-VP64 was induced in MPH-expressing HEK 293T cells and treated with Blasticidin for 2 weeks. The MPH-dCas9-VP64-expressing HEK 293T applied to further experiments. The control and MYC activation gRNA (Fig. S1C) were induced in MPH-dCas9-VP64-expressing HEK 293T cells. The CRISPR activation library, lentiSAMv2, was also transduced into MPH-dCas9-VP64-expressing HEK 293T cells and then treated with Zeocin (300 µg/ml, Invivogen) for 2 weeks.
Collection of Dendra2-positive cells and identification of gRNAs
After 2 weeks of selection with Zeocin, pMYC-promoter-Dendra2 was transfected into SAM library lentivirus-transduced cells with Lipofectamine™ 3000 Reagent (Invitrogen). HEK 293T cells transfected with pMYC-promoter-Dendra2 were evaluated for Dendra2-positive cells by fluorescence microscope (Keyence). Cells were collected at 72 h and washed with phosphate-buffered saline (PBS) and applied to the cell sorter. The Dendra2-positive cells were collected into a 96-well plate with MoFlo XDP (Beckman Coulter).
A total of 169 cells were collected, and 12 clones were successfully established. Collected clones were centrifuged and washed with PBS solution and pelletized. Lysis buffer solution with proteinase K (20 mg/ml) was added to cells and left for 30 min at 60 °C. After treatment at 94 °C for 10 min, the lysate was centrifuged for 15 min, and the supernatant was collected as genome DNA (gDNA) containing fractions. Using extracted gDNA of each collected clone, PCR was conducted with GoTaq® qPCR Master Mix (Promega) with the following primers: 5′-GAGGGCCTATTTCCCATGAT -3′ (forward) and 5′-GAGGGCCTATTTCCCATGAT-3′ (reverse). PCR product containing the gRNA region was employed for TA cloning with pGEM®-T Vector Systems (Promega) and gRNAs were sequenced by Sanger-sequencing.
Preparation of CRISPR backbone plasmid with target gRNAs and transcriptional activation for MYC using the CRISPR activation system
The oligonucleotides for each target gRNA sequence were designed using the list from the SAM library and integrated into the pENTR221/U6/stuffer/MS2 vector (Kurata et al., 2018a). The MPH-dCas9-VP64-expressing HEK 293T cells (1 × 105 cells/ml) were seeded in a 24-well cell culture plate. The next day, 2.5 µg of individual gRNA vector and pMYC- promoter-Dendra2 were transfected with Lipofectamine ™ 3000 Reagent. At 48 h after the transfection, the MYC transcriptional activation was evaluated by Dendra2-positivity by Keyence.
Evaluation of MYC expression
A pTRIPZ-M1AP-GFP was transduced into the HEK 293T cells. These transduced cells were selected with puromycin (1 µg/ml, Invitrogen) for 2 weeks. Seventy-two hours after the induction of Doxycycline (DOX) (1 µg/ml), the collected cells were employed for RNA isolation.
RNA was isolated using an RNeasyR Mini Kit (Qiagen) according to the manufacturer’s instructions. Complementary DNA (cDNA) was generated from RNA with ReverTra AceR qPCR RT Master Mix (Toyobo). Beta-ACTIN was used as an endogenous control. Using the ABI Prism 7900HT (Applied Biosystems), quantitative PCR (qPCR) analysis was performed to quantify the RNA level using SYBR Mix. The sequences for the PCR primers used in gene expression were as follows: MYC : 5′-CGACTCTGAGGAGGAACAAGAA-3′ (forward) and 5′-CAGCAGAAGGTGATCCAGACT-3′ (reverse), β-ACTIN: 5′-CACAGAGCCTCGCCTTTGCC-3′ (forward) and 5′-CACAGAGCCTCGCCTTTGCC-3′ (reverse). The mRNA level of the targeted gene was analyzed by comparison with the standard calibration curve. Western blot analysis was performed according to standard procedures (Kurata et al., 2016). The primary antibodies used were anti-c-Myc (Abcam ab32072) and anti-GAPDH (Santa Cruz sc47724), each at a dilution of 1:1000. The primary antibodies were detected with horseradish peroxidase (HRP)-conjugated secondary antibodies at 1:50000. Protein bands were visualized with ClarityTM Western ECL Substrate (Bio-Rad).
Dual-Luciferase reporter assay
The fragments of the MYC promoter was integrated into a pGL4 vector (Promega). A pT3.5-CAG-M1AP was generated with pENTR221-M1AP and pT3.5-CAG-DEST (Kurata et al., 2018a) using Gateway® LR clonase. The luciferase reporter vectors, the pT3.5-CAG-M1AP vector, and the pRL Renilla luciferase reporter vector were co-transfected in the HEK 293T cells. The samples were harvested 48 and 72 h after the transfection. Each assay was biologically triplicated and repeated. Luciferase activities were measured using the dual-luciferase reporter assay system (Promega) and Lumat LB9507 (Perkin Elmer). RNA was harvested at the same time points and the expression of MYC mRNA was analyzed by qPCR.
M1AP and MYC expressions in databases
For the purpose of comparing gene expression between cancer and non-cancerous tissue, the GEPIA web server was employed (Tang et al., 2017) (http://gepia2.cancer-pku.cn/).
Datasets for AML samples were used in BloodSpot database, that database was used to retrieve gene expression data from curated human and murine microarray and RNA-sequencing (RNA-Seq) datasets (Bagger et al., 2016). GSE42519, GSE13159, GSE15434, GSE61804, GSE14468, and The Cancer Genome Atlas (TCGA) datasets were used for analysis. In addition, following database were used; The Human Protein Atlas (https://www.proteinatlas.org/) and cBioPortal (https://www.cbioportal.org) using TCGA data (Cancer Genome Atlas Research Network et al., 2013; Gao et al., 2013; Cerami et al., 2012).
Statistical analysis
The data were statistically analyzed using the EZR version 1.36 software. A value of p < 0.05 was considered statistically significant for all analyses.
Results
Development of the MYC promoter-reporter system for library screening
Using PCR primers designed across the proximal MYC promoter at −3.1 kb (−3134 to −40) from the MYC TSS, PCR was performed. The MYC promoter region was integrated upstream of Dendra2. The pDendra2 harboring the 3.1 kb MYC promoter region was called “pMYC-promoter-Dendra2” for further experiments (Fig. S1A). Plasmids of pMYC-promoter-Dendra2 and gRNA of MYC or HPRT (as a negative control) were transfected in HEK 293T cells using the CRISPR activation system. We confirmed that Dendra2 showed positivity only in the cells with gRNA for MYC (Figs. S1B, S1C). Therefore, pMYC-promoter-Dendra2 was utilized as a MYC promoter-reporter analysis tool in this study.
MYC transcriptional activator screening Using the CRISPR activation library
To screen transcriptional activators working on the MYC promoter region, the CRISPR activation library was employed. The SAM library lentivirus was infected into MPH-dCas9-VP64-expressing HEK 293T cells (1 × 107 cells). After selection, pMYC- promoter-Dendra2 was transfected into these HEK 293T cells. The multiplicity of infection (MOI) ranged from 1.4 to 5.2.
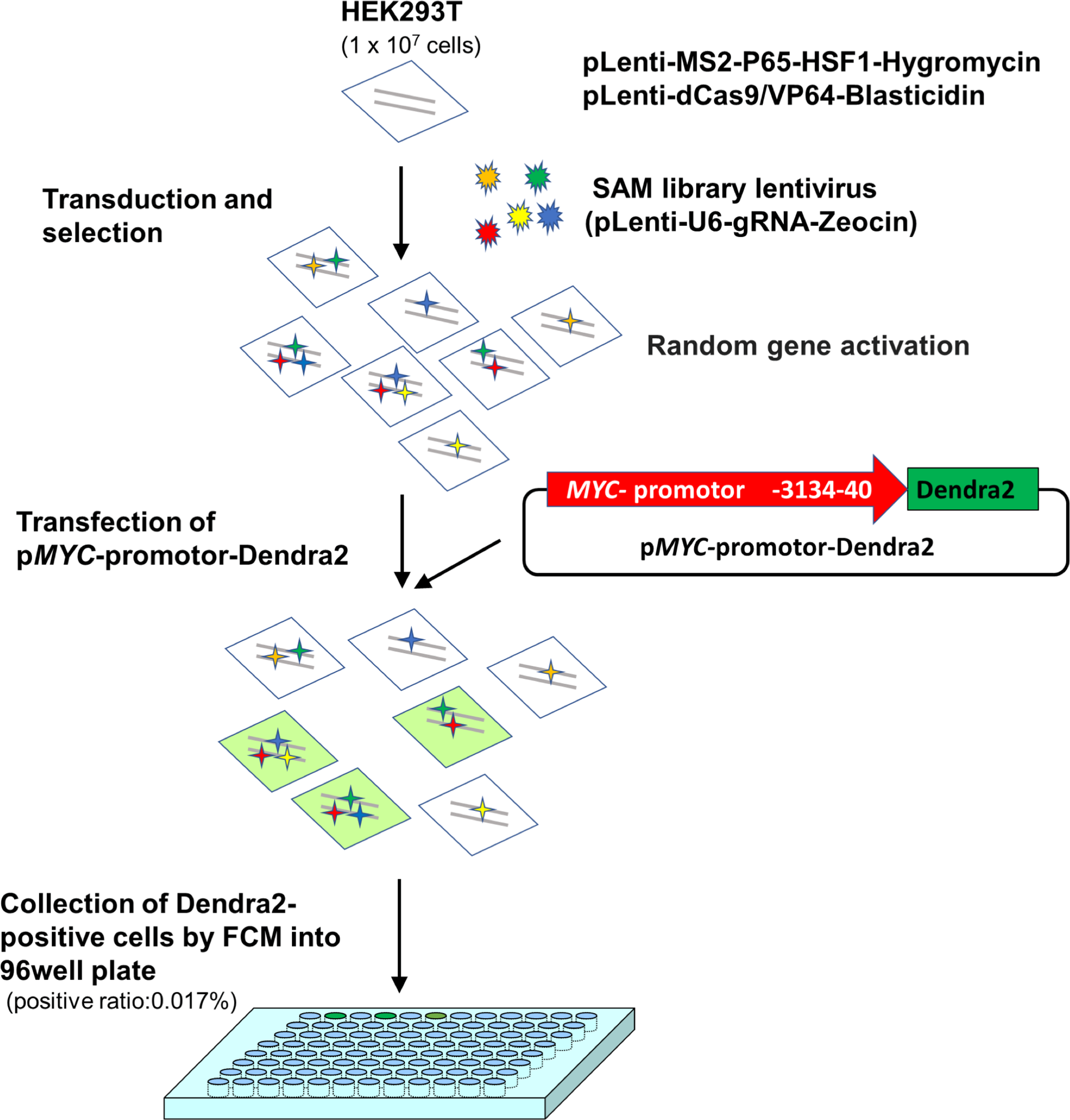
At 72 h after transfection, cells were proceeded to Dendra2-positive cell collection using flow cytometry (positive ratio: 0.017%). Sorted single cells were collected into a 96-well plate and incubated for 2 weeks. Among the collected 169 cells, 12 clones were successfully established (Fig. 1).
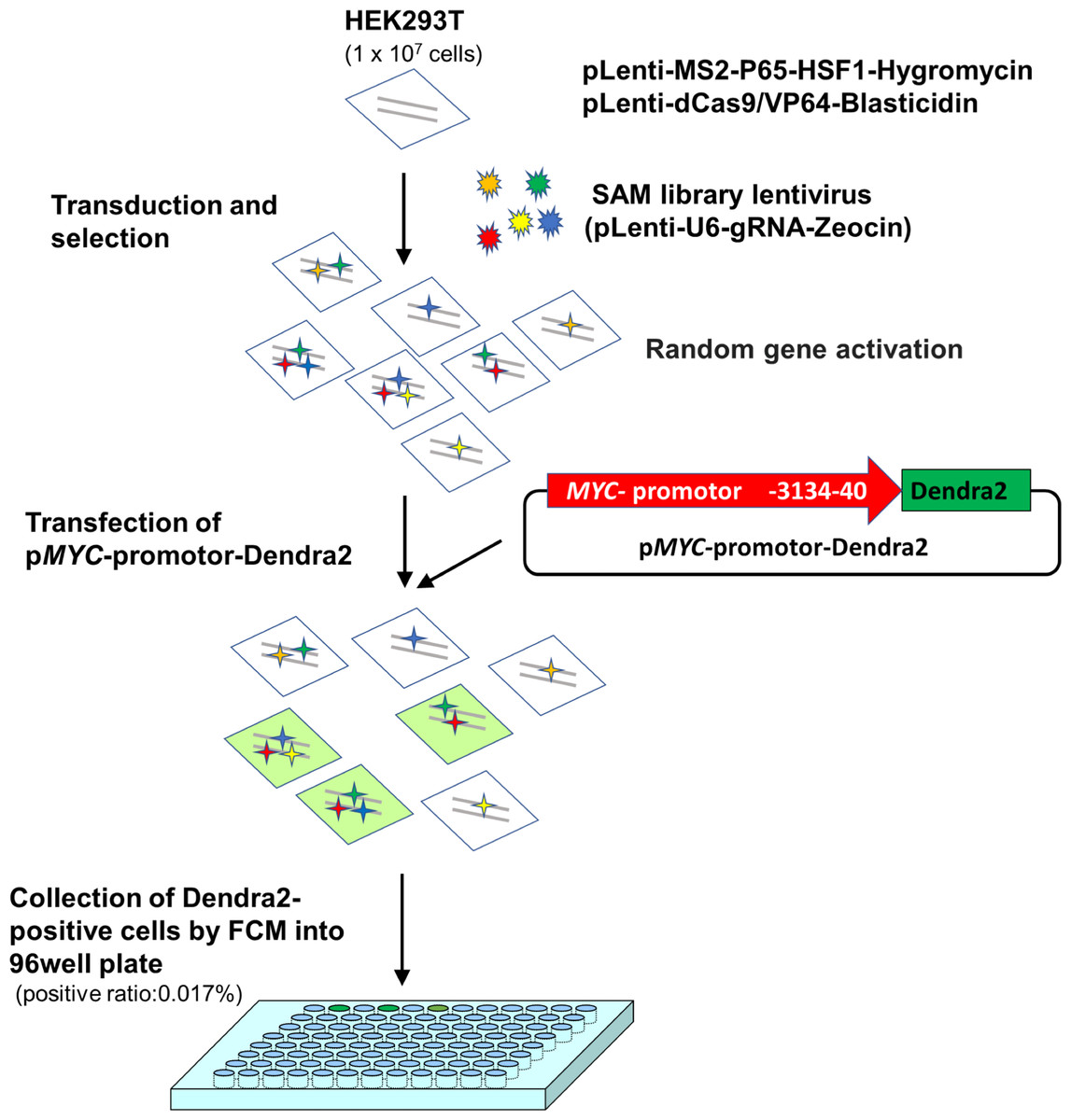
Figure 1: A scheme for MYC transcriptional activator screening with the CRISPR activation library.
{kind=link}
Second screening to identify candidate genes as MYC activators
For the second screening, pMYC-promoter-Dendra2 was again transfected into the 12 clones. Using fluorescence microscope, clones 3 and 5 were shown to be truly Dendra2 positive (Figs. 2A–2L), and the photoswitchable attribute of Dendra2 was useful for distinguishing it from cells with non-specific autofluorescence (Fig. S2). Using extracted gDNA of clones 3 and 5, PCR was performed to amplify the gRNA containing region, which was introduced by the CRISPR activation library. Then, the PCR product was employed for TA cloning, followed by colony PCR. By this method, four candidate genes—PLEKHO2 , MICU, MBTPS1, and M1AP were identified (Table 1).
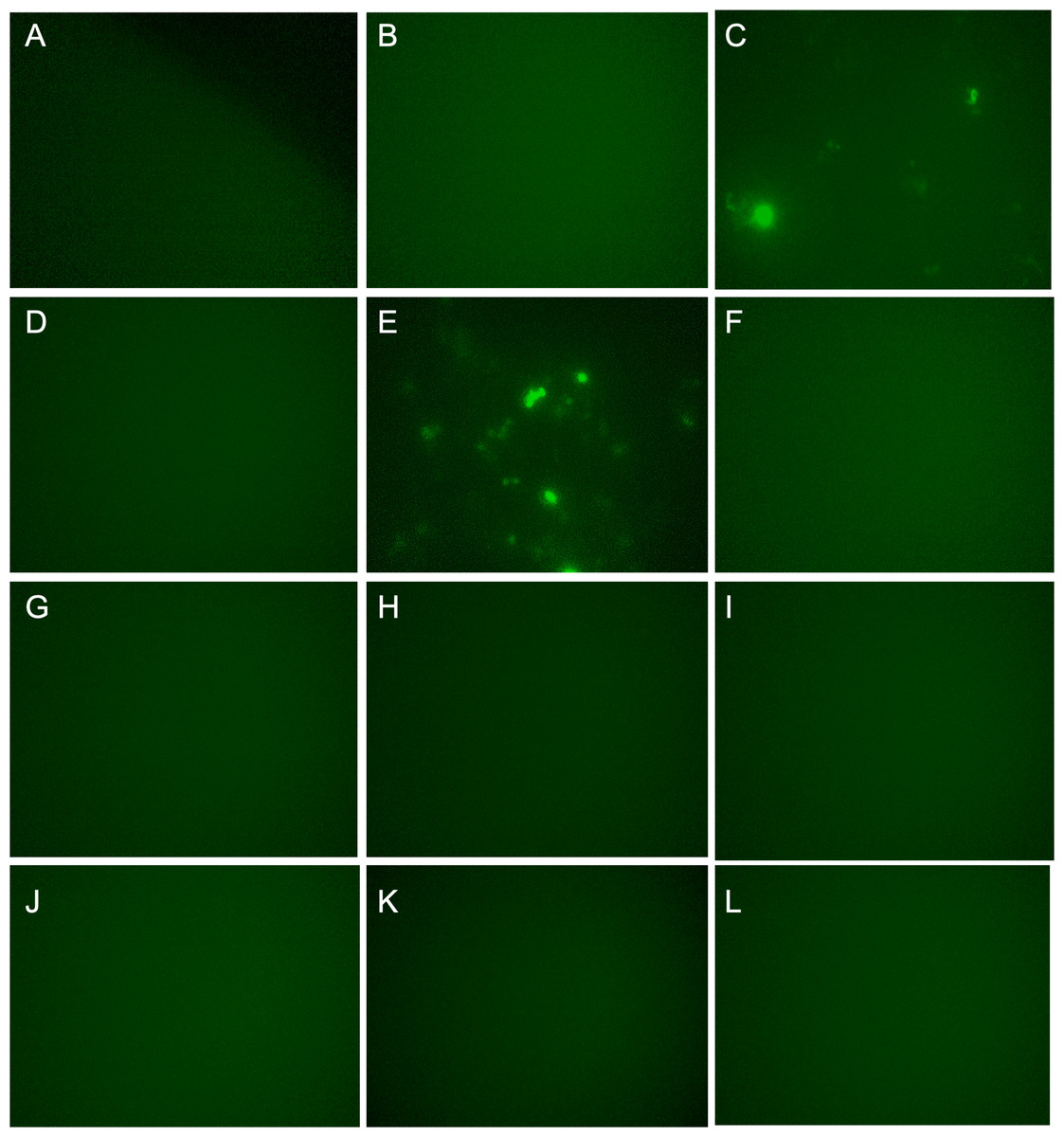
Figure 2: Dendra2-positive cells using fluorescence microscope.
(A)–(L) correspond Clone 1-12. p MYC- promoter-Dendra2 was transfected into 12 clones that were successfully cloned after CRISPR activation screening. Clones 3 and 5 showed Dendra2 positivity.{kind=link}
Each candidate gRNA was transfected to the HEK 293T cells using the CRISPR activation system accompanied by pMYC-promoter-Dendra2. Because Dendra2 was positive only in the cells transfected with M1AP gRNA (Figs. 3A–3D), M1AP was considered as the most promising gene that would increase MYC transcriptional activity. Then, double-stranded DNA fragments coding the full length of M1AP induced into Doxycycline-inducible lentiviral vector (pTRIPZ-M1AP-GFP, Fig. 4A). The pTRIPZ-M1AP-GFP was introduced into HEK 293T cells. Seventy-two hours after the induction M1AP with DOX (1 µg/ml), high MYC expression was observed in DOX-induced cells by RT qPCR (t-test, p < 0.05, Fig. 4B). Furthermore, in the western blots, an increase of MYC expression was also observed in DOX-induced cells (Fig. 4C).
Additionally, a dual-luciferase assay conducted with a reporter containing the MYC promoter region showed a significant increase of promoter activity in M1AP-overexpressing cells at both 48 and 72 h (Figs. 4D and S3). On the other hand, the MYC mRNA, which was isolated at the same time points, showed mildly higher in M1AP-overexpressing cells at 48 h but not at 72 h (Figs. 4E and Fig. S3 ). Taken together, although M1AP was confirmed to activate the MYC promoter strongly, the MYC mRNA itself might be regulated also by other factors.
Correlation between MYC and M1AP in the database
According to the GEPIA web server (Tang et al., 2017) (http://gepia2.cancer-pku.cn/), high M1AP expression is observed in cervical carcinoma, acute myeloid leukemia (AML), ovarian carcinoma, thyroid carcinoma, uterine endometrial carcinoma, and uterine carcinosarcoma when compared to normal tissue (Fig. 4SA). Among these carcinomas, a correlation analysis using cBioPortal revealed a mild positive correlation between MYC and M1AP only in AML (Fig. 5A, Figs. S4B- S4F). Therefore, we focused on hematopoietic cells, and M1AP expression was analyzed using the BloodSpot database. As a result, M1AP was significantly higher in AML tissue compared to normal bone marrow tissue (Fig. 5B). Interestingly, M1AP was highly expressed in AML, especially with t (8;21) (Fig. 5C).
| Sequence of gRNA | Gene name |
|---|---|
| GGGCGGGAAATGGGTGGGGA | PLEKHO2 |
| TTGTGACACTACTCCAGCCT | MICU1 |
| AGGATCCCCGAAAAGGAGCA | MBTPS1 |
| TCTCAGGGTATCTAGGGACT | M1AP |
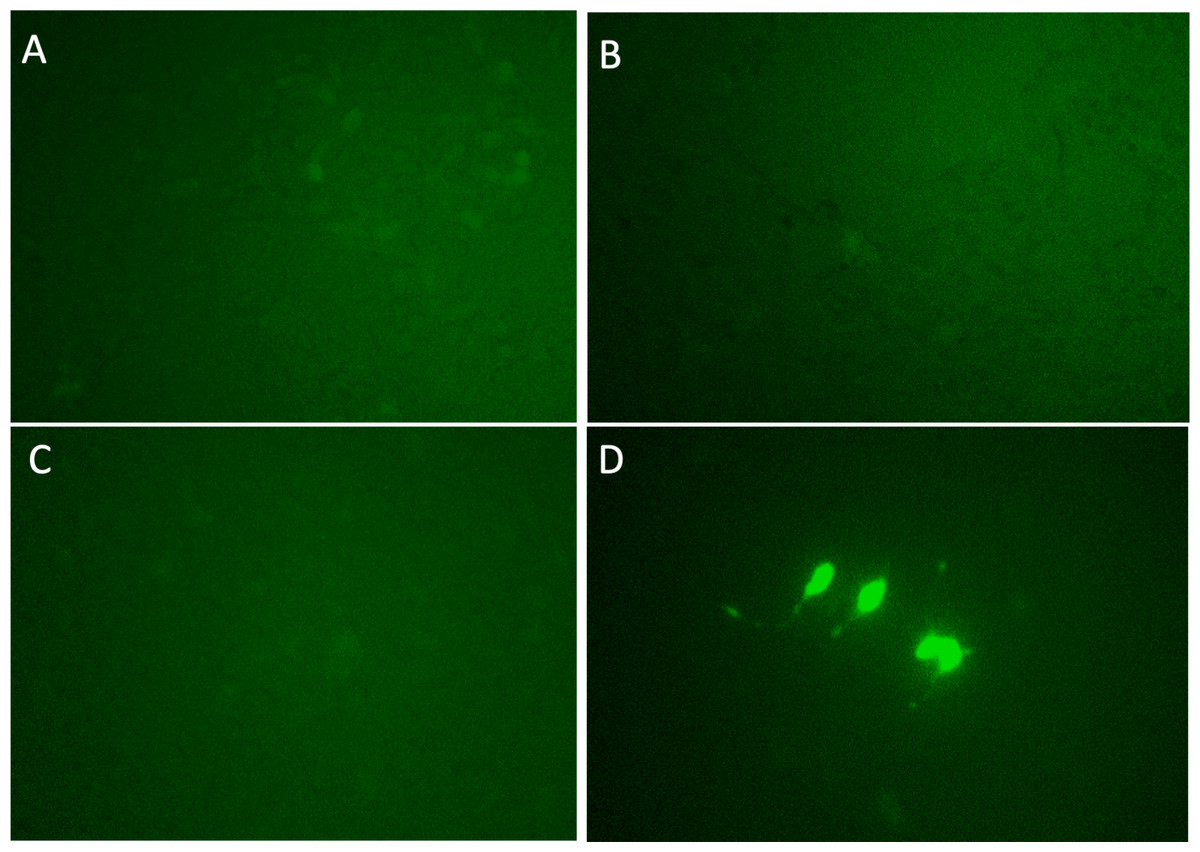
Figure 3: Validation experiments for candidate genes as MYC activators.
(A) PLEKHO2 (B) MICU3 (C) MBTPS1 (D) M1AP. With gRNAs of the four nominated genes, p MYC- promoter-Dendra2 was transfected into HEK 293T cells by applying the CRISPR activation system. Only M1AP-introduced cells were Dendra2 positive.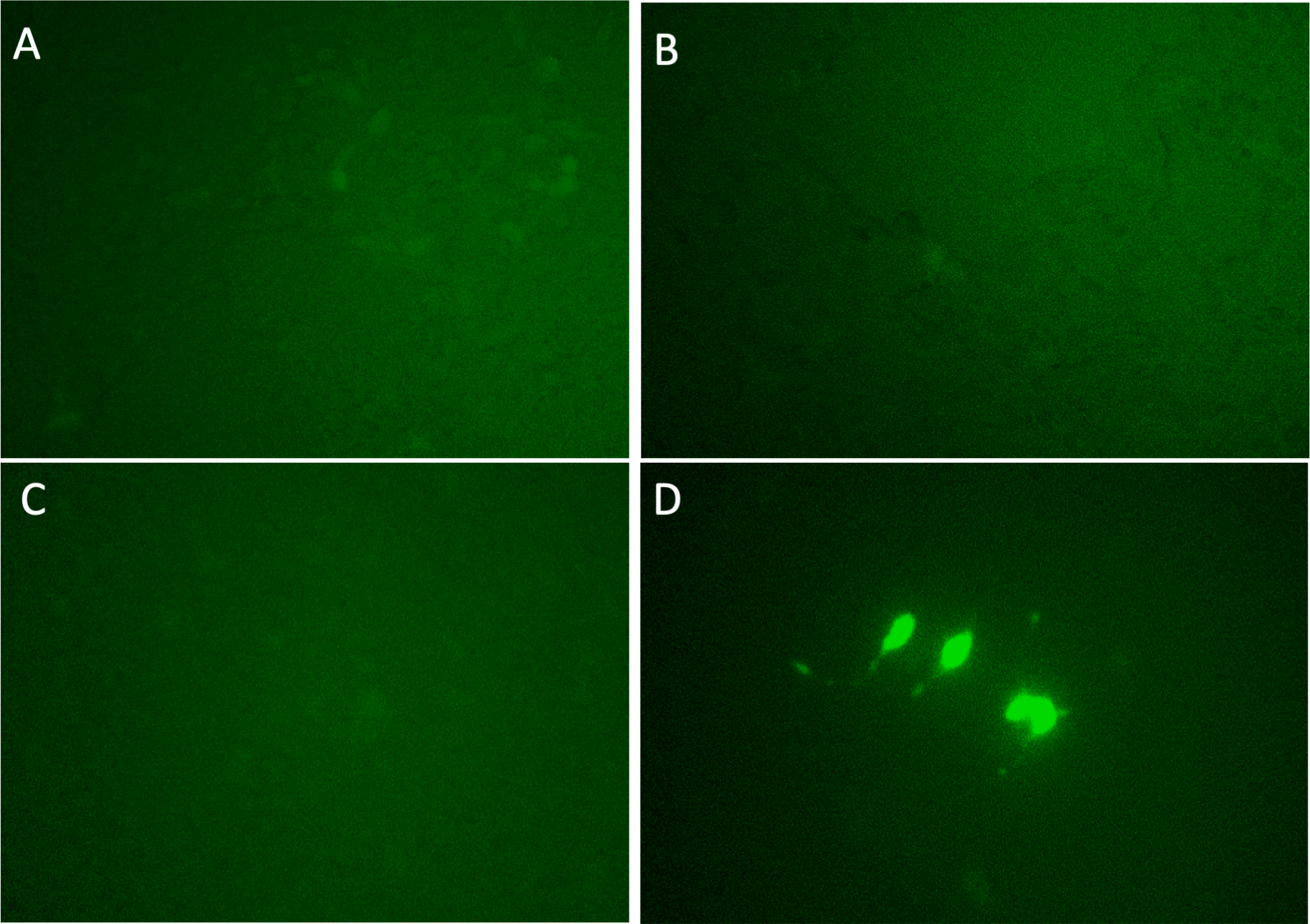{kind=link}

Figure 4: MYC mRNA expression analyzed by real-time quantitative polymerase chain reaction.
(A) Vector map of Dox-inducible M1AP expression vector. (B) High MYC expression was observed in DOX-induced cells (t-test, p < 0.05). (C) Western blotting analysis showed increasing of MYC protein in DOX- induced cells. (D) Luciferase activity at 48 hours was significantly higher in M1AP-overexpressing cells (t-test, p < 0.05). (E) MYC mRNA expression at 48 hours was mildly higher in M1AP-overexpressing cells (t-test, p < 0.05).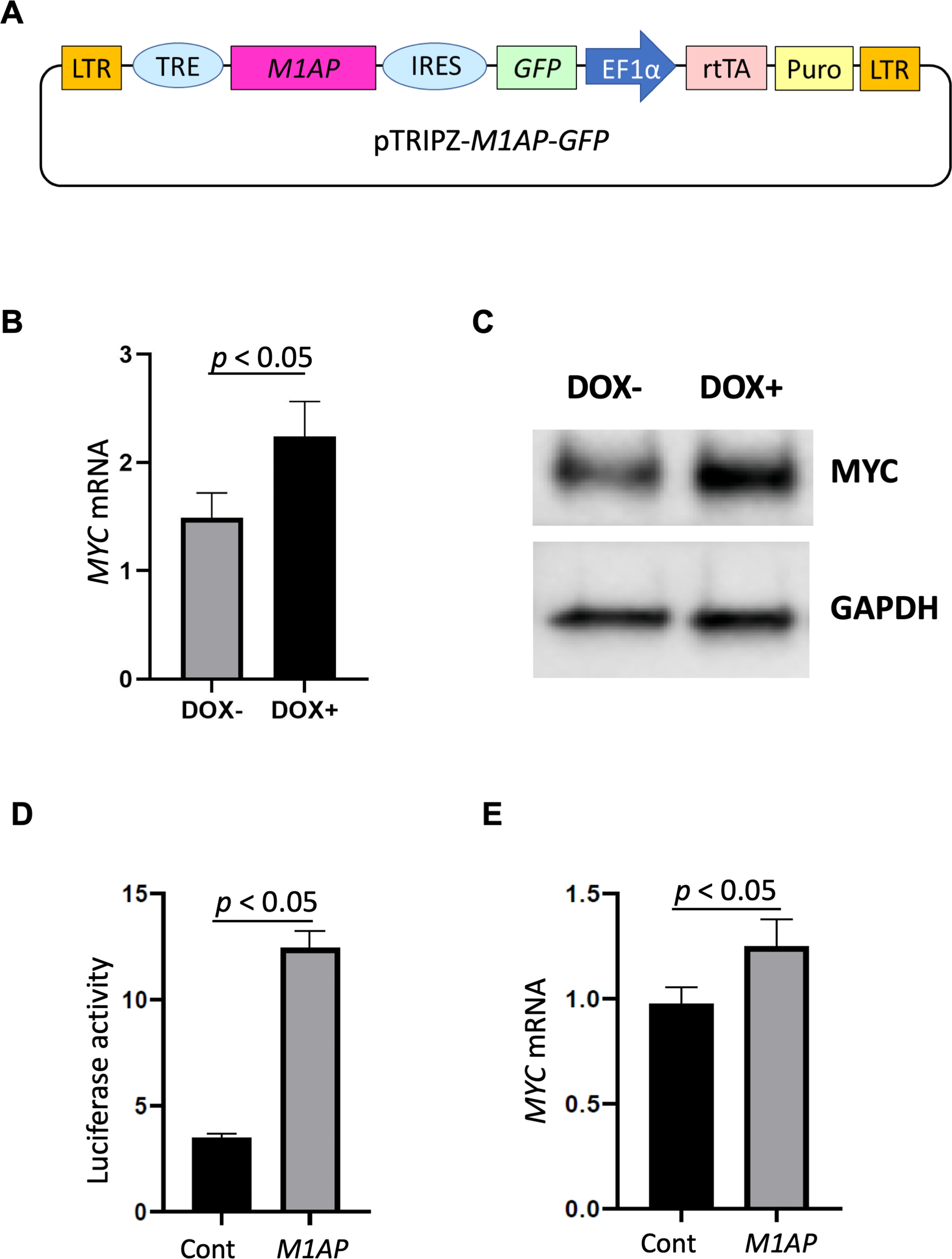{kind=link}
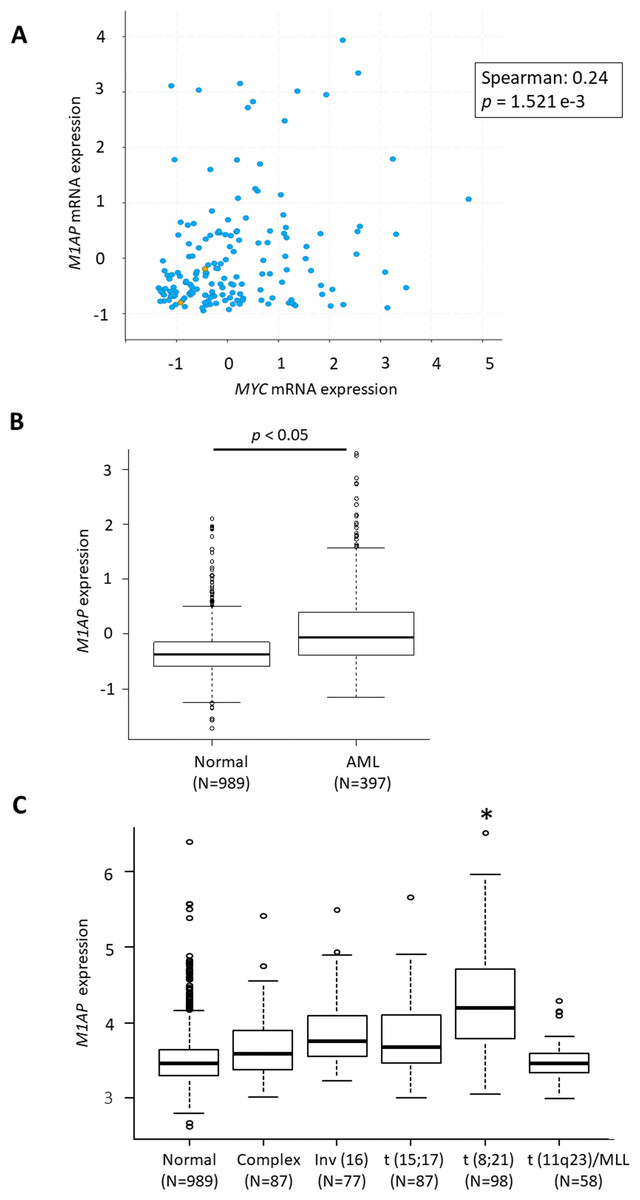
Figure 5: Correlation between MYC and M1AP in the database.
(A) In cBioPortal using TCGA data, a correlation analysis between MYC and M1AP in AML revealed a mild positive correlation. (B) Using the Bloodspot database M1AP mRNA expression in acute myeloid leukemia (AML, N = 989) was significantly higher than in normal tissue (N = 397). (Mann–Whitney U test, p < 0.05) (C) The BloodSpot database was used to retrieve gene expression data. M1AP expression was higher in each subtype of AML, especially with t (8;21). (Kruskal–Wallis test, *: p < 0.01).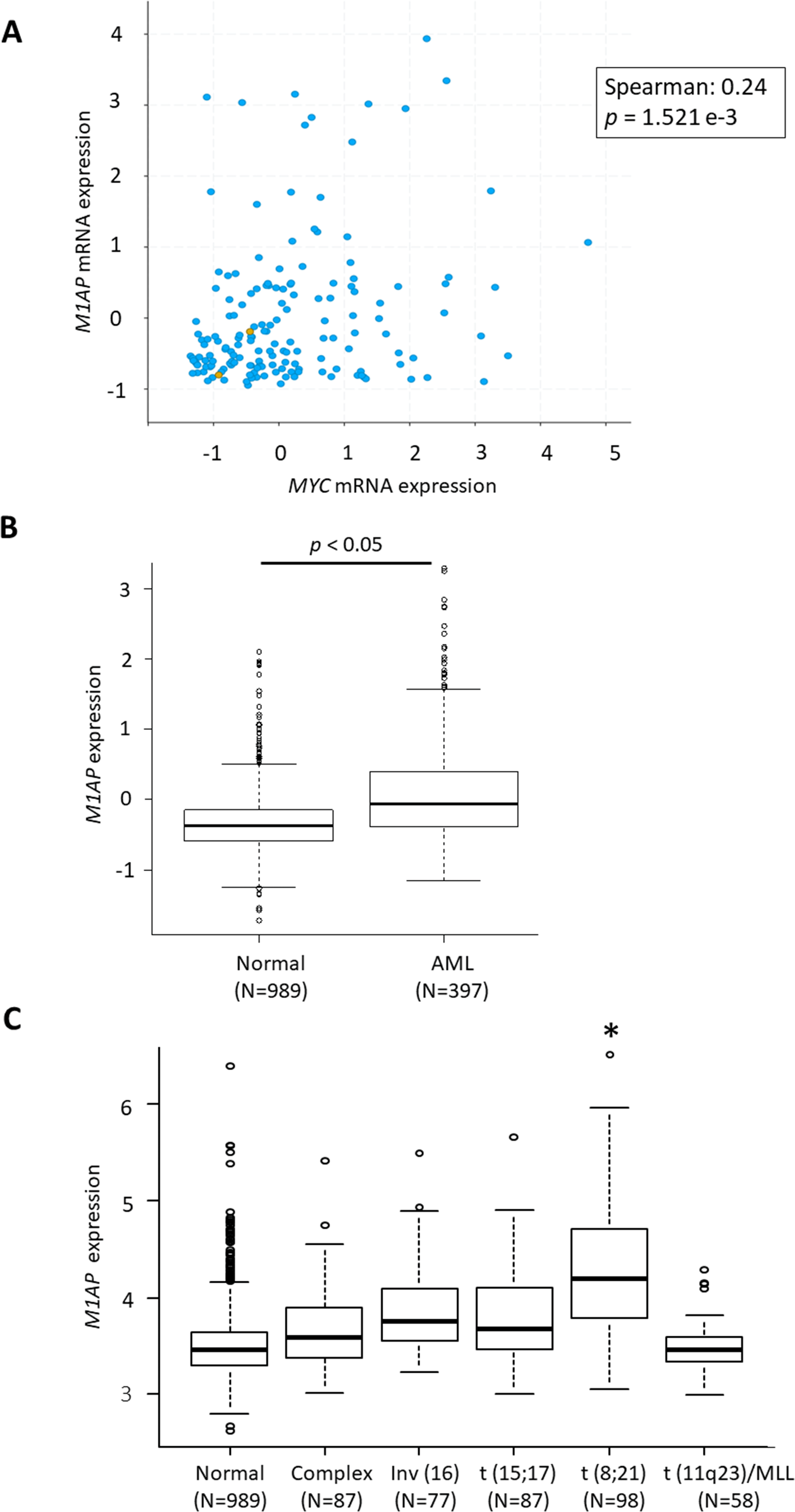{kind=link}
Discussion
In this study, we created a new reporter vector that incorporates the MYC promoter region across the proximal −3.1 kb from the TSS and photoconversion fluorescent protein to evaluate the transcriptional activity of endogenous MYC. Ideally, a large region of the MYC promoter on Chr.8q24 would be applied to better understand the full MYC promoter mechanism. However, even short region on the MYC promoter region are still not fully understood. In designing this new vector, 3.1 kb of the promoter region was applied because of the limited length of effective DNA insertion into the vector. Future improved experiments on larger region will help to clarify the whole mechanism of MYC regulation via the promoter region.
By applying this tool to the CRISPR library, we screened molecules that increase the expression of endogenous MYC. According to previous reports, by introducing the CRISPR library into a malignant tumor cell line and carrying out selection with a drug, molecules that cause susceptibility or resistance to that drug can be identified (Konermann et al., 2015; Gilbert et al., 2014; Kampmann, 2018; Kurata et al., 2018a; Kurata et al., 2018b; Shalem et al., 2014; Wang et al., 2014; Koike-Yusa et al., 2014; Doench et al., 2016; Jain et al., 2016). However, this is the first specific report of a molecule that activates transcription upstream of MYC.
MYC is an indispensable factor for cell growth and proliferation in normal tissues, but at the same time, it is one of the most well-known proto-oncogenes, with high amplification in various carcinomas. Moreover, MYC is widely involved in tumor development and growth (Dang, 2012). Elucidation of factors that regulate the transcriptional activity of MYC is crucial for establishing new therapeutic strategies for tumors. Although MYC expression is strictly regulated by mRNA and MYC protein, in 1998, He et al. reported that inactivation of APC located downstream of Wnt signaling pathway results in excessive accumulation of β-catenin and promotes endogenous MYC expression by acting on TCF present in the promoter region of MYC.
Regarding the research of the regulatory mechanism of MYC, Vo et al. (2018) transplanted CRISPR/dCas9-VP160 and gRNAs that activated MYC transcription in the mouse brain, resulting in the development of medulloblastoma. They concluded that this model is useful for future drug research by suppressing MYC expression.
Because this screening was adapted to about 70,290 kinds of gRNAs, initially, multiple molecules were expected to be identified; however, the screening eventually led to the identification of only one molecule, M1AP. The reason for this is that, on performing single-cell collection using flow cytometry, to avoid collecting false-positive cells, the cells with the highest fluorescence intensity were collected; as a result, the number of collected cells was extremely small. Furthermore, since the proliferation of the cells after the collection was poor, it is expected that collected cells included cells that were highly brightened due to cell senescence. Moreover, oncogene-induced cell senescence can be one explanation for why the survival rate of collected single cells was very low. Oncogene-induced senescence, which is now a well-known phenomenon, causes cell growth arrest where the oncogene is excessive. This was first reported in RAS (Serrano et al., 1997), and the same mechanism has been reported for MYC, where MYC oncogene overexpression results in inactivation of MYC, followed by the arrest of cell growth and proliferation (Wu et al., 2007). Excessive MYC expression due to the CRISPR activation system may have induced cell senescence and caused a low survival rate of sorted cells. We would like to use a next-generation sequencing system and adjust the multiplicity of infection in future screenings.
By newly established experimental line in this study, M1AP, which was found as a transcription factor for MYC, was proved to function strongly as a MYC activator by promoter assay. On the other hand, MYC expression was mildly elevated in the M1AP- overexpressing cells collected at the same time as the luciferase assay. These results suggest that the MYC mRNA is complexly controlled by multiple factors and multiple pathways.
The present study confirmed that this experimental method applying the CRISPR library technology functioned effectively for the identification of the molecule that activates endogenous MYC. This method will help elucidate the regulatory mechanism of MYC expression, as well as facilitating further drug research against malignant tumors. However, more investigation is required to establish a more accurate and precise screening technique.
Supplemental Information
{kind=link}
{kind=link}
{kind=link}
Luciferase activity in M1AP-overexpressing cells and MYC mRNA expression at 72 h
MYC promoters are strongly activated at 72 h after induction of M1AP expression, but the mRNA of MYC was not significantly upregulated at 72 h.
{kind=link}
M1AP expression and correlation with MYC in databases
(A) Comparison of M1AP expression in normal and tumor tissue in GEPIA. (B)–(F) Correlations between MYC and M1AP in cBioPortal. (B) Cervical carcinoma. (C) Ovarian carcinoma. (D) Thyroid carcinoma. (E) Uterine endometrial carcinoma. (F) Uterine carcinosarcoma.
{kind=link}