
PHACTR1 is associated with disease progression in Chinese Moyamoya disease
- Published
- Accepted
- Received
- Academic Editor
- Joanna Elson
- Subject Areas
- Cell Biology, Genomics, Neurology
- Keywords
- Whole-exome sequencing, Moyamoya disease, Mutation, PHACTR1
- Copyright
- © 2020 Yang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. PHACTR1 is associated with disease progression in Chinese Moyamoya disease. PeerJ 8:e8841 https://doi.org/10.7717/peerj.8841
Abstract
Moyamoya disease (MMD) is a progressive stenosis at the terminal portion of internal carotid artery and frequently occurs in East Asian countries. The etiology of MMD is still largely unknown. We performed a case-control design with whole-exome sequencing analysis on 31 sporadic MMD patients and 10 normal controls with matched age and gender. Patients clinically diagnosed with MMD was determined by digital subtraction angiography (DSA). Twelve predisposing mutations on seven genes associated with the sporadic MMD patients of Chinese ancestry (CCER2, HLA-DRB1, NSD-1, PDGFRB, PHACTR1, POGLUT1, and RNF213) were identified, of which eight single nucleotide variants (SNVs) were deleterious with CADD PHRED scaled score > 15. Sanger sequencing of nine cases with disease progression and 22 stable MMD cases validated that SNV (c.13185159G>T, p.V265L) on PHACTR1 was highly associated with the disease progression of MMD. Finally, we knocked down the expression of PHACTR1 by transfection with siRNA and measured the cell survival of human coronary artery endothelial cell (HCAEC) cells. PHACTR1 silence reduced the cell survival of HCAEC cells under serum starvation cultural condition. Together, these data identify novel predisposing mutations associated with MMD and reveal a requirement for PHACTR1 in mediating cell survival of endothelial cells.
Introduction
Moyamoya disease (MMD) is a progressive stenosis at the terminal portion of internal carotid artery (ICA) with compensatory development of a hazy network of basal collaterals called Moyamoya vessels (Scott & Smith, 2009; Suzuki & Takaku, 1969). The prevalence of MMD is the highest in East Asian countries, including Japan, Korea and China (Kuroda & Houkin, 2008; Miao et al., 2010). The annual incidences of MMD in China and Japan are 0.43 and 0.54 per 100,000, which are significantly higher than that in the USA (0.086 per 100,000) and Europe (0.3 per 100,000) (Kraemer, Heienbrok & Berlit, 2008; Kuriyama et al., 2008; Miao et al., 2010; Uchino et al., 2005). Epidemiology studies had revealed several risk factors associated with MMD, such as Asian ethnicity, female gender and family history (Ganesan & Smith, 2015). Although the heritability of MMD is unknown, the genetic components may play an important role in the etiology of MMD (Inayama et al., 2018).
Previous studies have explored and revealed several genetic loci associated with MMD, such as 3q24-p26, 6q25, 8q23, 17q25 (Ikeda et al., 1999; Inoue et al., 2000; Sakurai et al., 2004; Yamauchi et al., 2000). A polymorphism of c.14576G>A in the RNF213 gene (RNF213) on the 17q25-ter region was identified as a novel susceptibility gene for MMD in Japanese and Chinese populations with a founder effect (Liu et al., 2011). RNF213 is correlated with early onset and severe forms of MMD. Recently, rare variants on the C-terminal of RNF213 were found correlated with MMD arteriopathy in patients of European ancestry (Guey et al., 2017), while a genome-wide association study involving a large case-control study among Chinese ancestry revealed 10 novel loci could be responsible for MMD, which extended our knowledge of MMD (Duan et al., 2018). However, the etiology of MMD is far more complicated than we expected, and further study on genome wide association is necessary.
Disease progression is among the most frequent reason for clinical symptomatic events. The clinical characteristics has been increasingly recognized as a prevalent cause of disease progression in those patients with natural course, such as early age onset, autoimmune factors and family heritability, but the evaluation is mostly focused on clinical characteristics (Dlamini, Muthusami & Amlie-Lefond, 2019; Grangeon et al., 2019; Jiang et al., 2018; Kim & Jeon, 2014). The relationship between genetic risk factor and disease progression in MMD patients remains unknown.
In this study, we performed a case-control design with whole-exome sequencing analysis on 31 sporadic MMD patients (nine cases with disease progression and 22 stable MMD cases) and 10 normal controls with matched age and gender. Predisposing mutations were discovered and then validated by Sanger sequencing. We identified 12 predisposing mutations on seven genes associated with the sporadic MMD patients of Chinese ancestry and further validated that SNV (c.13185159G>T, p.V265L) on PHACTR1 was highly associated with the disease progression of MMD. Finally, we evaluated the effect of PHACTR1 on cell survival of endothelial cells.
Material and Methods
Subjects
We enrolled 31 MMD subjects and 10 non-related healthy controls with matched age and gender during 2018 to 2019 at Department of Neurosurgery, The Affiliated Drum Tower Hospital of Nanjing University Medical School, Nanjing, China. Subjects recruited into this study were all Chinese ancestry. The study was reviewed and approved by the research ethics committee, Nanjing Drum Tower Hospital of Nanjing University Medical School (2018-173-01). Written informed consent was taken from each participant.
Patients with MMD determined by digital subtraction angiography (DSA) were based on the guidelines established by the Japanese Research Committee on Moyamoya disease of the Ministry of Health, Welfare and Labor, Japan (RCMJ) (Fukui, 1997). Information on family histories, gender, age, onset symptoms were obtained by interview. Cases with additional evidence of arthrosclerosis, meningitis, autoimmune diseases, brain neoplasm, Down syndrome, Recklinghausen disease, irradiation or other obvious specific etiologies were excluded. All included MMD subjects all first time came to the hospital and diagnosed with the MMD. All these MMD subjects did not have any drug treatment for MMD or other conditions.
Treatment and clinical follow-up
Through the use of single-photon emission computed tomography (SPECT), cerebral blood flow (CBF) was semi-quantitatively measured for all MMD patients 1 month after the initial onset. After preoperative imaging evaluation, surgical revascularization, including superficial temporal artery to middle cerebral artery anastomosis combined with encephalo-duroarterio-synangiosis (EDAS), was conducted in the hemisphere with intracranial hemorrhage or with more severe ischemia. If a patient elected not to undergo surgery, conservative management was used instead. After baseline investigation and surgical intervention, all patients underwent clinical follow-up. Brain MRI and MRA were performed every 6 to 12 months. During the follow-up period, if major intracranial artery stenosis progression was suspected on MRA or cerebrovascular events occurred, cerebrovascular DSA was performed within 1 week. The presence or absence of disease progression was evaluated at the final follow-up visit which defined as the progression of any major intracranial artery stenosis >50% in the ICA and/or the posterior cerebral artery (PCA). Representative cases with disease progression are shown in Fig. 1.
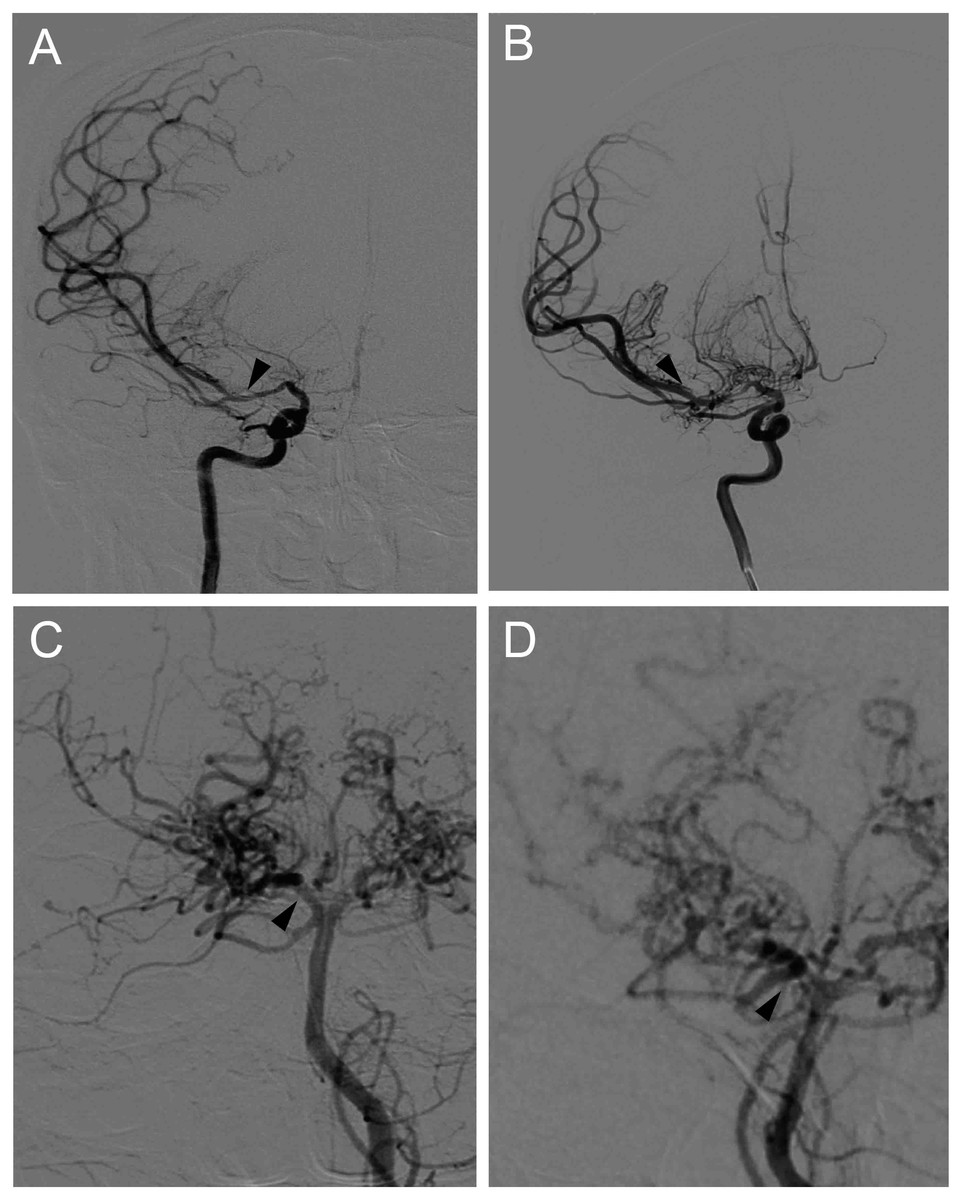
Figure 1: Representative cases of disease progression in MMD.
(A) Right internal carotid angiograms of a 48 y/r man, showing artery stenosis progression in the proximal portion of right MCA (arrows). (B) Right posterior cerebral angiograms of a 51 y/r woman, showing artery stenosis progression in the proximal portion of right PCA (arrows).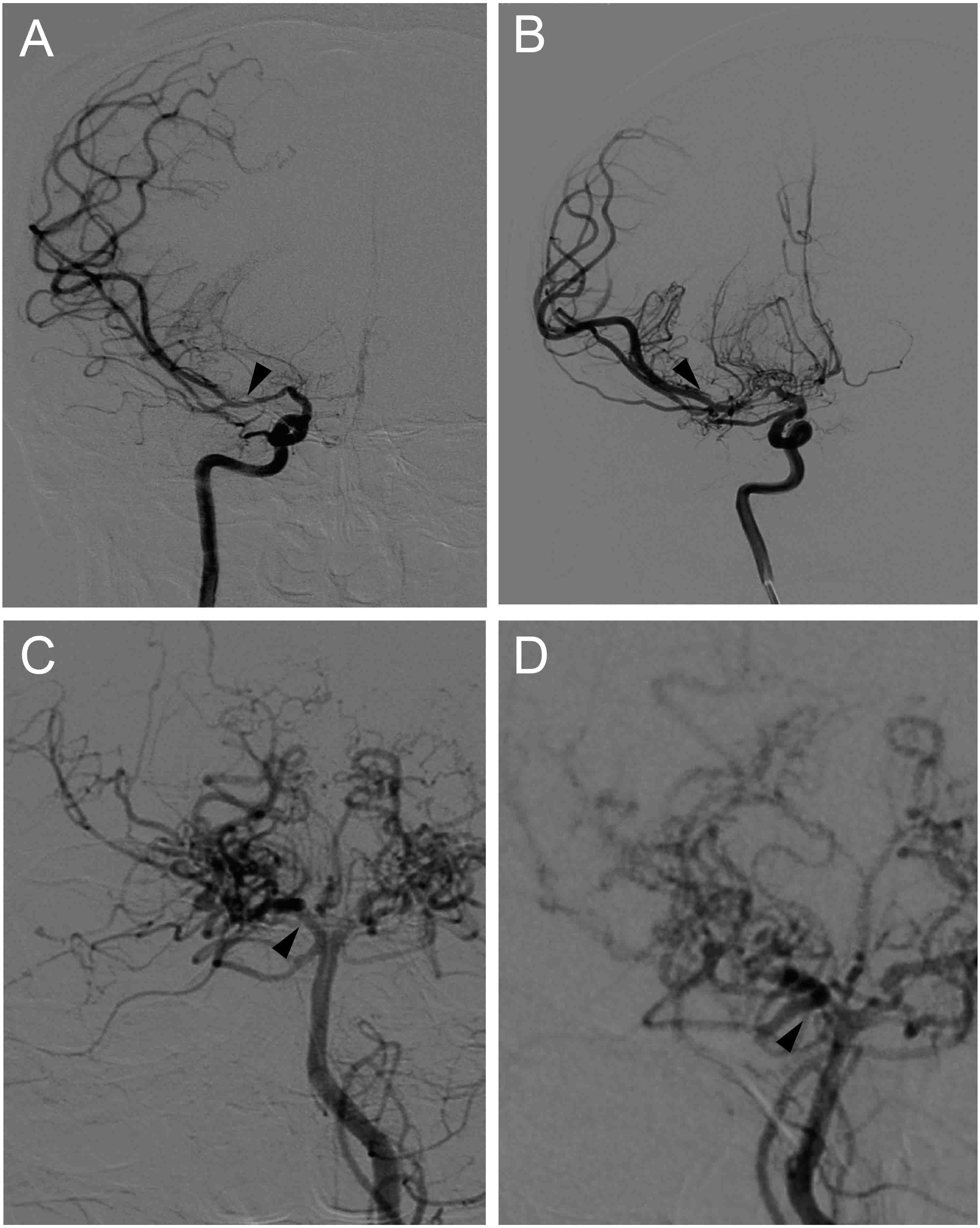{kind=link}
Blood sample extraction and storage
Three tubes of 15 mL peripheral blood samples were collected and subjected to DNA extraction (Qiagen). DNA extraction method is seen in the exome sequencing analysis section. DNA samples were used immediately to next experiments, or store at −80 °C for later usage.
Whole-exome sequencing and Sanger sequencing
We conducted exome sequencing analysis on 31 sporadic MMD subjects and 10 non-related healthy control subjects. Peripheral blood samples were collected from the subjects, and DNA was extracted using the QIAamp™ DNA and Blood Mini kit (Qiagen™, Munich, Germany), and sheared using acoustic fragmentation (Covaris) and purified using a QIAquick PCR Purification Kit (Qiagen). The whole-exome DNA library was sequenced on an Illumina HiSeq X Ten platform and performed as previously described (Stachler et al., 2015).
The Sanger sequencing was performed in Genscript Ltd. (Nanjing, China).
Bioinformatics analysis
Raw sequencing reads and all qualified reads were processed with an in-house bioinformatics pipeline, which followed as previously described (Stachler et al., 2015). Duplicated fragments were marked by Picard v1.141. After converting the data into bam format, GATK BaseRecalibration module was used to improve the base quality and then HaplotypeCaller module was used to discover genetic variants (SNV/INDEL). SnpEff 4.3i and Gemini v0.18.0 were used for functional annotation with Online Mendelian Inheritance in Man (OMIM), the Exome Aggregation Consortium (ExAC) Browser, MutationTaster2 and the Combined Annotation Dependent Depletion (CADD).
Variants filtering and selection
Variants were excluded if they had a call rate <98%, major allele frequency <1%, abnormal heterozygosity >(mean ± 3SD), or significant deviation from Hardy–Weinberg equilibrium among controls (Phwe < 1 ×10−4). The non-coding, synonymous, impact_severity=low, MAF (minor allele frequency) >0.01 variants were excluded from the raw data. SO_IMPACT=HIGH was referenced using MAF database including ESP, 1KG, ExAC database. The definition of “SO_IMPACT=HIGH” is based on the Gemini (https://gemini.readthedocs.io/en/latest/content/database_schema.html#details-of-the-impact-and-impact-severity-columns). The filtered variants was compared with known MMD related genes, such as RNF213, CCER2, HLA-DRB1. SNVs with CADD PHRED scaled score >15 were regarded as deleterious.
siRNA transfection and Western blotting
Human coronary artery endothelial cell (HCAEC) was kindly gifted from Dr. Shengnan Li (Nanjing Medical University) and cultured in DMEM plus 10% fetal bovine serum (FBS, Hyclone, Waltham, MA). SiRNA targeting PHACTR1 (sc-95456, Santa Cruz Biotechnology, Santa Cruz, CA) and scrambled siRNA were transiently transfected into HCAECs using Lipofectamine 2000 Reagent (Invitrogen, Carlsbad, CA). HCAEC cells 48 h after transfection were lysed with ice-cold RIPA lysis buffer. The SDS-PAGE was performed as the standard procedure. Anti-PHACTR1 (sc-514800, Santa Cruz Biotechnology) and anti- β-actin (Sigma, St. Louis, MO) were used. Protein bands were visualized with ECL reagent (Thermo Scientific, Rockford, IL) and recorded by Tanon 5200 Multi Imaging Workstation (Tanon, Shanghai, China).
Cell survival assay
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) was used for the assessment of HCAEC cell survival and performed as previously described (Zhu et al., 2012).
Statistical analysis
Continuous variables were described as mean ± SD, and categorical variables were presented as number and percentage. An independent t-test, and a chi-square or Fisher exact test were used to compare patients with and without disease progression. A P value <0.05 was considered significant. All statistical analyses were performed using SPSS 23.0 (IBM, Chicago, Illinois).
Results
Characteristic of subjects and variants quantity
The mean age (years ± SD) of all enrolled cases was 33 ± 8.46, and sex ratio of male: female was 0.34. The mean age and sex ratio of these healthy controls were 35.1 ± 6.58 and 0.3, which matched the disease group (Table 1). Of the 31 patients, 14 (45.2%) presented as intracranial hemorrhage and 17 (54.8%) presented as ischemic stroke at baseline. Among these 31 subjects, 9 of them suffered from disease progression in 10 hemispheres during the follow-up periods including 8 in the anterior circulation and 2 in the posterior circulation, and the other 22 cases were diagnosed as stable MMD disease.
| Characteristics | MMD group | Healthy group |
|---|---|---|
| Age (Mean ± SD) | 33 ± 8.46 | 35.10 ± 6.58 |
| Gender (Male/Female) | 0.34 | 0.30 |
| Ages < 18y (%) | 7 (22.58%) | 2 (20%) |
| Ages ≥ 18y (%) | 24 (77.42%) | 8 (80%) |
| Family history | No | No |
| Disease progression (%) | 9 (29.03%) | — |
| Stable disease (%) | 22 (70.97%) | — |
| Other combined severe conditions | No | No |
By comparing with whole-exome sequencing results of the 31 sporadic MMD subjects with the human genome GRCh37.75, total 15,206 genes were successfully picked out. The main sequencing quality and depth were 85.42 M ±11.32 M reads, 99.69 ± 0.16 map (%), 111.87 ± 12.42 depth, suggesting a high quality of whole-exome sequencing. Total 196,365 variants were found including 176,582 site nucleotide polymorphisms (SNP), 8,906 insertions (INS) and 10,877 deletions (DEL) (Table 2). Total 117,748 (51.359%) variants were mis-senses, 1,525 (0.665%) variants were non-senses, and 109,992 (47.976%) variants were silent types. By comparing with the healthy subjects, 10,241 variants in exons were discovered.
| Chromosome | Length | Variants |
|---|---|---|
| 1 | 249,250,621 | 19,250 |
| 2 | 243,199,373 | 13,079 |
| 3 | 198,022,430 | 10,188 |
| 4 | 191,154,276 | 7,014 |
| 5 | 180,915,260 | 8,074 |
| 6 | 171,115,067 | 12,066 |
| 7 | 159,138,663 | 10,358 |
| 8 | 146,364,022 | 6,694 |
| 9 | 141,213,431 | 8,301 |
| 10 | 135,534,747 | 8,454 |
| 11 | 135,006,516 | 11,547 |
| 12 | 133,851,895 | 9,960 |
| 13 | 115,169,878 | 3,211 |
| 14 | 107,349,540 | 6,748 |
| 15 | 102,531,392 | 7,578 |
| 16 | 90,354,753 | 9,221 |
| 17 | 81,195,210 | 10,799 |
| 18 | 78,077,248 | 3,263 |
| 19 | 59,128,983 | 13,925 |
| 20 | 63,025,520 | 4,873 |
| 21 | 48,129,895 | 2,394 |
| 22 | 51,304,566 | 5,289 |
| X | 155,270,560 | 3,838 |
| Y | 59,373,566 | 241 |
| Total | 3,095,677,412 | 196,365 |
Significant predisposing mutations associated with disease progression
By comparing with the known MMD-related genes, 12 predisposing mutations on seven gene exons had from medium to high impact on gene modification (Table 3). These genes includes RNF213, CCER2, HLA-DRB1, NSD-1, PDGFRB, PHACTR1 and POGLUT1, 8 of these SNVs were deleterious with CADD PHRED scaled score >15. Next, we performed Sanger sequencing to validate the association between predisposing mutations and MMD disease progression. SNV (c.13185159G>T, p.V265L) on PHACTR1 was found in 3 cases with disease progression and shown significantly associated with its progression.
| Locus | Gene | Position | Ref | Alta | Codon change | Amino acid change | Qual | Depth | Impact | CADD raw score | CADD PHRED scaled scored | Progression/ Stable | wild typeb | Mutantb | P valuec |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 19q13.2 | CCER2 | 39401715 | C | T | c. 39401715C>T | p.G67R | 1763.4 | 3451 | Medium | 1.187 | 14.37 | Progression | 9 | 0 | 0.71 |
| Stable | 21 | 1 | |||||||||||||
| 6p21.32 | HLA-DRB1 | 32549352 | G | C | c. 32549352G>C | p.P212A | 372.77 | 6450 | Medium | 1.940 | 18.63 | Progression | 8 | 1 | 0.71 |
| Stable | 22 | 0 | |||||||||||||
| 32551942 | T | C | c. 32551942T>C | p.D105G | 2688.4 | 16207 | Medium | 2.289 | 21.90 | Progression | 8 | 1 | 0.71 | ||
| Stable | 22 | 0 | |||||||||||||
| 5q35.3 | NSD1 | 176562643 | T | C | c. 176562643T>C | p.I180T | 1044.4 | 2736 | Medium | 2.862 | 23.30 | Progression | 9 | 0 | 0.71 |
| Stable | 21 | 1 | |||||||||||||
| 176638711 | A | G | c. 176638711A>G | p.H1104R | 1108.4 | 2199 | Medium | 0.003 | 2.68 | Progression | 9 | 0 | 0.71 | ||
| Stable | 21 | 1 | |||||||||||||
| 5q32 | PDGFRB | 149513304 | G | A | c. 149513304G>A | p.P260L | 2883.4 | 4330 | Medium | 2.866 | 23.30 | Progression | 9 | 0 | 0.71 |
| Stable | 21 | 1 | |||||||||||||
| 6p24.1 | PHACTR1 | 13185159 | G | T | c. 13185159G>T | p.V265L | 4277.88 | 4183 | Medium | 2.622 | 22.80 | Progression | 6 | 3 | 0.019 |
| Stable | 22 | 0 | |||||||||||||
| 3q13.33 | POGLUT1 | 119211265 | A | T | c. 119211265A>T | p.M387L | 1367.4 | 2013 | Medium | 0.846 | 12.20 | Progression | 9 | 0 | 0.71 |
| Stable | 21 | 1 | |||||||||||||
| 17q25.3 | RNF213 | 78291014 | A | T | c. 78291014A>T | p.Q995H | 736.4 | 3008 | Medium | 0.677 | 10.91 | Progression | 9 | 0 | 0.71 |
| Stable | 21 | 1 | |||||||||||||
| 78319385 | T | G | c. 78319385T>G | p.I2466S | 3066.4 | 3554 | Medium | 3.499 | 25.10 | Progression | 9 | 0 | 0.71 | ||
| Stable | 21 | 1 | |||||||||||||
| 78320960 | T | C | c. 78320960T>C | p.V2991A | 2086.4 | 3562 | Medium | 1.924 | 18.48 | Progression | 9 | 0 | 0.71 | ||
| Stable | 21 | 1 | |||||||||||||
| 78321631 | A | G | c. 78321631A>G | p.I3215V | 1814.4 | 3802 | Medium | 2.739 | 23.10 | Progression | 9 | 0 | 0.71 | ||
| Stable | 21 | 1 |
The effect of PHACTR1 on cell survival
We hypothesized that PHACTR1, one of seven genes mentioned above, may mediate cell survival of human endothelial cells. We knocked down the expression of PHACTR1 by transfection with siRNA targeting PHACTR1 (Fig. 2A), and then measured the cell survival of HCAEC cells (Fig. 2B). Western blotting revealed that siRNA targeting PHACTR1 largely downregulated the expression of PHACTR1 in HCAEC cells, insulting in a ∼80% decrease (Fig. 2A). After the successive 96 h measurement of cell survival rate, we found that PHACTR1 silence reduced the cell survival of HCAEC cells under serum starvation cultural condition for 96 h, but not for 24∼72 h (Fig. 2B).
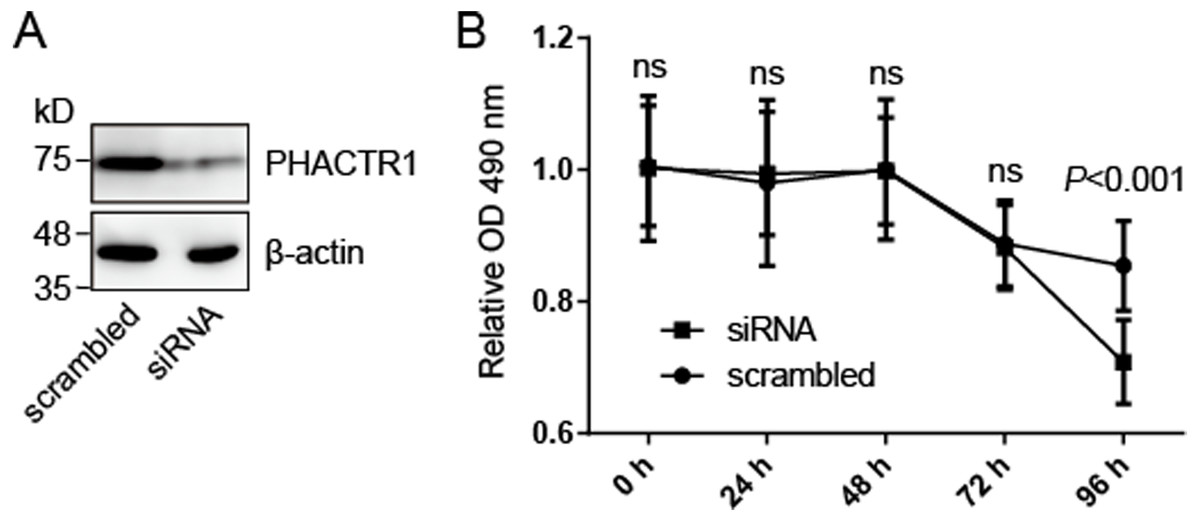
Figure 2: PHACTR1 silence reduces the cell survival of HCAEC cells.
(A) HCAEC cells were transiently transfected with siRNA targeting PHACTR1 or scrambled siRNA. Western blotting verified the knockdown efficiency of siRNA targeting PHACTR1. β-actin as the loading control. (B) HCAEC cells transfected with siRNA targeting PHACTR1 or scrambled siRNA were cultured under serum starvation cultural condition for successive 96 h. Cell survival rate was assessed by cell survival assays. PHACTR1 silence reduced the cell survival of HCAEC cells under serum starvation cultural condition for 96 h.{kind=link}
Discussion
All of the recruited sporadic MMD patients are all first-time being diagnosed with this disease, and did not receive any clinical treatment or drugs administration for any other medical conditions. RNF213 was identified as a novel susceptible gene for MMD in East Asian population. This study identified 4 novel susceptibility loci and confirmed the previous reported susceptibility gene RNF213. Furthermore, we identified HLA-DRB1 variants are found in the sporadic MMD patients. HLA-DRB1 play a central role in the immune system by presenting peptides derived from extracellular proteins (Ji et al., 2018; Lauterbach et al., 2014). Although MMD patients with auto-immune diseases were excluded from this study, immune system dysfunction is still associated with the MMD pathophysiological process. CCER2 was reported as a biomarker for MMD by Japanese colleagues (Mukawa et al., 2017). NSD-1 is a transcriptional factor (Jo et al., 2016). In this study, we found a variant on CCER2 locus and two mutations on NSD-1.
Our results suggest that cytoskeleton system and cardiovascular development are involved with the MMD pathological process. PDGFRB and PHACTR1 work on the actin cytoskeleton system, and PDGFRB regulates cardiovascular development (Onel et al., 2018; Perez-Hernandez et al., 2016). POGLUT1 works on the NOTCH signaling pathway to regulate developments (Wu et al., 2017). According to Sanger sequencing, we found that SNV (c.13185159G>T, p.V265L) on PHACTR1 was significantly associated with MMD disease progression. Owing to the lack of appropriate animal model for MMD, we provisionally checked the effect of genes associated with MMD susceptibility on immortal endothelial cells. We found PHACTR1 mediates the cell survival of endothelial cells under serum starvation cultural condition. Nevertheless, the precise mechanisms of MMD needs to be further studied on molecular and cellular levels.
The main limitation of the study is the small size of samples. The subjects are difficult to recruit, we have attempted to perform a meta-analysis to strengthen or our conclusions. We have gone through three databases and obtained 76 articles about MMD (Pubmed, n = 23; Embase, n = 33; Web of science, n = 20). The duplicated articles ( n = 42) were removed. Then, 29 articles were excluded through sceening title or/and abstract, full text, due to review, animal studies, case report and/or unrelated outcomes, moyamoya syndrome or other diseases. After reading the full text of 13 sceened articles, we have not found these 8 SNVs mentioned in Table 3. So, we are unfortunately unable to perform a meta-analysis in the present research status.
Conclusions
We perform whole-exome sequencing on 31 sporadic MMD subjects and 10 healthy volunteers, and identify 12 predisposing mutations of seven genes (RNF213, CCER2, HLA-DRB1, NSD-1, PDGFRB, PHACTR1 and POGLUT1) associated with MMD and SNV (c.13185159G>T, p.V265L) on PHACTR1 associated with its progression. We preliminarily provide the rational evidence of the effect of PHACTR1 on endothelial cell survival and indicate its involvement in the pathophysiological process of MMD.
{kind=link}