
Using Ipomoea aquatic as an environmental-friendly alternative to Elodea nuttallii for the aquaculture of Chinese mitten crab
- Published
- Accepted
- Received
- Academic Editor
- María Ángeles Esteban
- Subject Areas
- Agricultural Science, Aquaculture, Fisheries and Fish Science, Microbiology, Freshwater Biology, Environmental Contamination and Remediation
- Keywords
- Macrophyte system, Chinese mitten crab, Water nutrients, Bacterioplankton community
- Copyright
- © 2019 Shi et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Using Ipomoea aquatic as an environmental-friendly alternative to Elodea nuttallii for the aquaculture of Chinese mitten crab. PeerJ 7:e6785 https://doi.org/10.7717/peerj.6785
Abstract
Elodea nuttallii is widely used in Chinese mitten crab (CMC) rearing practice, but it is not a native aquatic plant and cannot endure high temperature. Thus, large E. nuttallii mortality and water deterioration events could occur during high-temperature seasons. The aim of this study was to identify the use of local macrophytes in CMC rearing practice, including Ipomoea aquatic and Oryza sativa. A completely randomized field experiment was conducted to investigate the crab yield, water quality, bacterioplankton community and functions in the three different systems (E. nuttallii, I. aquatic, and O. sativa). Average crab yields in the different macrophyte systems did not differ significantly. The I. aquatic and O. sativa systems significantly decreased the total nitrogen and nitrate-N quantities in the outflow waters during the rearing period compared to the E. nuttallii system, and the I. aquatic and O. sativa plants assimilated more nitrogen than the E. nuttallii plant. Moreover, the significant changes of bacterioplankton abundances and biodiversity in the three systems implied that cleanliness of rearing waters was concomitantly attributed to the differential microbial community and functions. In addition, principle component analysis successfully differentiated the bacterioplankton communities of the three macrophytes systems. Environmental factor fitting and the co-occurrence network analyses indicated that pH was the driver of bacterioplankton community structure. Functional predictions using PICRUSt (v.1.1.3) software based on evolutionary modeling indicated a higher potential for microbial denitrification in the I. aquatic and O. sativa systems. Notably, the O. sativa plants stopped growing in the middle of the rearing period. Thus, the I. aquatic system rather than the O. sativa system could be a feasible and environmental-friendly alternative to the E. nuttallii system in CMC rearing practice.
Introduction
The Chinese mitten crab (CMC), Eriocheir sinensis, is considered an invasive species in Europe and North America (Brodin & Drotz, 2014; Hanson & Sytsma, 2008), but it is an expensive delicacy in Asia (Chen & Zhang, 2007; Food and Agriculture Organization of the United Nations (FAO), 2019). In 2014, 796,621 tons of farmed CMCs were produced (Zeng et al., 2013); crabs were primarily bred in ponds and lakes (Zeng et al., 2013). The mitten crabs produced in Yangcheng Lake, Suzhou, China, are of high quality and have high economic value (Gu et al., 2013). Most of the crabs produced in Yangcheng Lake are exported to Shanghai, Hong Kong, and high-profit foreign markets.
Aquatic plants are required for mitten crab farming. The plants provide shelter for the crabs during exuviation, which is an important part of crab growth (Meng et al., 2013). In addition, aquatic plants assimilate excess nutrients, improve water cleanliness, and absorb solar radiation to maintain cool water temperatures. These properties increase crab growth, yield, and quality (Zhan & Yang, 2015).
Elodea nuttallii, a perennial aquatic plant native to North America, provides these benefits, and is thus widely used in CMC aquaculture (Wang et al., 2016). However, E. nuttallii cannot withstand high temperatures (Zhan & Yang, 2015). In Yangcheng Lake, summer air and water temperatures typically reach 35–40 and 26–34 °C, respectively, which frequently leads to massive E. nuttallii die off, resulting in serious water quality deterioration (Wu et al., 2016). Under such conditions, crab growth is negatively affected due to loss of shelter for exuviation hiding places and poor water quality (Zhan & Yang, 2015). Consequently, alternative aquatic plants are required to facilitate mitten crab aquaculture in areas such as Yangcheng Lake.
Local plants that have adapted to local conditions are the best candidates for E. nuttallii replacement. For example, Ipomoea aquatica is a semiaquatic, tropical/subtropical plant that might be applicable to mitten crab aquaculture (Zhang et al., 2014). I. aquatic grows well in shallow waters, withstands high temperatures, and efficiently removes nutrients (e.g., nitrogen and phosphorous) from water bodies (Liu et al., 2007; Wei et al., 2017). Furthermore, the tender shoots and leaves of I. aquatic are consumable, and providing additional economic value. Alternatively, Oryza sativa is a submerged grain crop that is common in Asia, and its assimilation ability of nitrogen and phosphorus is similar to I. aquatic (Nawaz & Farooq, 2017). O. sativa is thus another candidate for E. nuttallii replacement.
Therefore, in this study, we aimed to answer three questions. Which of the two locally adapted plants would most adequately replace E. nuttallii, similarly improving crab yield and quality? How do the candidate plant systems affect water quality and the associated environmental characteristics? Considering microorganisms play important roles in nutrient conversion in wastewater (Daims, Taylor & Wagner, 2006), are the two plant systems associated with different bacterioplankton communities that differentially affect water quality? Answers to these questions are of critical importance to crab farmers, and those who are concerned with water quality. Despite the importance of these questions, they remain unanswered. We thus aimed to address the above questions by evaluating candidate aquatic plant systems, and exploring the associated bacterioplankton communities.
Material and Methods
Pond construction, seedling preparation, and floating system construction
All of the experiments were performed in Lianzigang, Suzhou, China (31°27′40.18″N, 120°43.5′5.32″E). This region has a subtropical monsoon climate, with an annual average rainfall of 1,076.2 mm (http://www.pmsc.cn/). Annual average temperature in Lianzigang is 15–17 °C, with high temperatures of 35–40 °C in July and August (http://www.pmsc.cn/). All of the experiments were conducted between May 1 and November 30, 2017.
The crab-rearing system used here consisted of a water inlet, a water outlet, a micro-porous aeration system, a pond, floating macrophytes, and vertical posts to anchor the macrophytes (Fig. 1). The pond, which was mechanically excavated, had a total area of 1,000 m2 and was 1.5-m deep. The pond was separated into nine sub-ponds with cement walls that were 0.4 m wide and 1.5 m high. Each sub-pond had an area of ∼100 m2 and an independent staff gauge. Sub-pond bottoms were left in a natural state to allow crabs to burrow, and to facilitate E. nuttallii root establishment and growth. I. aquatic, O. sativa, and E. nuttallii were grown in separate sub-ponds with three replicates per plant using a completely randomized design.
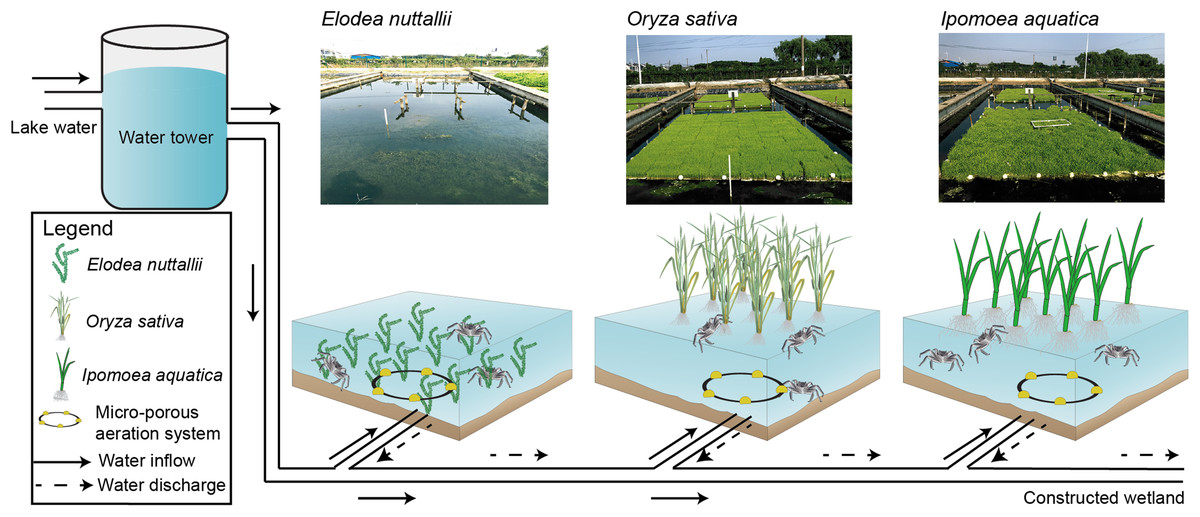
Figure 1: Experimental design and schematics for three macrophyte systems.
Elodea nuttallii, Oryza sativa, and Ipomoea aquatica. Each macrophyte was planted in a separate sub-pond (three replicate ponds per species). The sub-ponds were connected by a series of PVC pipes. Pond water was replenished from the Yangcheng Lake water, and discharged water flowed into a constructed wetland. The growth of all three macrophytes was restricted to 60% of the pond areas. A micro-porous aeration system was used nightly to ensure sufficient aeration. Photo credit: Linlin Shi.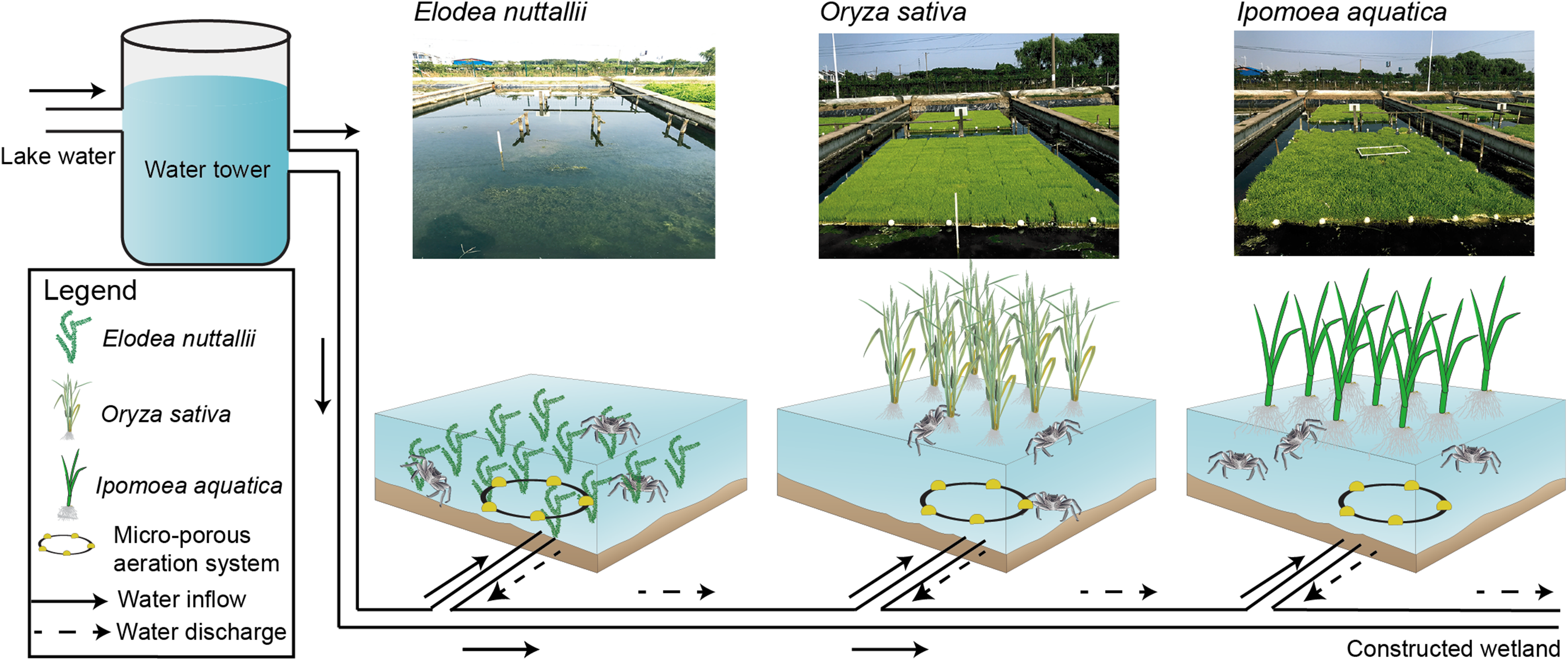{kind=link}
Sub-ponds were disinfected using sodium hypochlorite. After disinfection, water was pumped into each sub-pond to a depth of ∼10 cm. Basal fertilizer (45 kg ha−1 of compound-fertilizer; N:P2O5:K2O = 15:15:15) was applied 2–3 days before seedling transplantation.
Oryza sativa and I. aquatic seedlings were prepared by sowing seeds on a patented nutritional matrix (0.16 m2) containing sufficient nutrients for seedling growth. The matrix had sufficient buoyancy to allow seedlings to float on the surface of the water. Seeds were germinated on the matrix, grown for 30 days in a greenhouse at 25 °C, and then transferred to the bottom of the appropriate sub-ponds. Seedling matrices were arranged side by side until seedling coverage reached 60% of the total water surface area. Additional water was then pumped into each sub-pond to increase the water depth to 20 cm, and seedlings were allowed to re-establish growth for 5–7 days. Upon growth re-establishment, water was again gradually and gently pumped into the sub-ponds to increase water depth to 1.0 m. At this depth, all of the seedlings were floating. Seedlings were fixed in place using ropes attached to buoys, which were fastened to posts in the sides of the cement dividers. E. nuttallii seedlings were directly transplanted from other ponds. E. nuttallii cluster spacing was 0.5 × 0.5 m, with 40 seedlings per cluster. The transplanted E. nuttallii plants covered ∼60% of the total water surface area. Upon growth re-establishment, water was pumped into the E. nuttallii sub-ponds to a depth of 1.0 m.
Crab pond management, nutrient measurements, and crab yield determination
Crab ponds were managed using standard CMC rearing methods (Zhan & Yang, 2015). Juvenile crabs, with an average weight of 15 g, were purchased from a local company (Su’an Fishery Co., Ltd., Nantong, China) and added to the experimental sub-ponds at a density of about 12,000 individuals per hectare. Crab feed (bait) was purchased from the Tongwei Group (http://www.tongwei.com/). The nutrient composition of the feed was varied to meet the different needs at each of the growth stages. Crabs were fed twice per day dependent on growth stage, as recommended by the bait manufacturer. Crabs were not dosed with antibiotics or chemical fishery drugs. A micro-porous aeration system was used nightly to ensure sufficient aeration. Approximately five cm of water was pumped out of all ponds each week, and replaced with an equal volume of fresh water from Yangcheng Lake (depending on local precipitation).
Outflow water samples were taken every 7–10 days during the rearing period, and immediately frozen at −20 °C. At the end of the experiment, nutrients in the outflow samples were measured using an auto analyzer (SKALAR SAN++, Breda, the Netherlands); the nutrients measured included total nitrogen (TN), total phosphorous (TP), ammonium-N (NH4+-N), nitrate-N (NO3−-N), and nitrite-N (NO2−-N). Absolute cumulative nutrient quantity in discharged water was measured by nutrient concentration in discharged water and outflow water quantity estimated by staff gauge. Outflow pH was measured using a WTW portable pH meter (ProfLine 3310; WTW, Weilheim, Germany). Inlet water samples were taken and measured every month during the rearing period, and average nutrient concentrations were finally calculated. Dissolved oxygen (DO) concentrations were not measured, because preliminary studies identified large spatial and temporal variations in DO concentrations (Fig. S1) due to uncontrollable factors (i.e., air temperature, air pressure, water disturbances, aeration, activities of aquatic organisms, and photosynthesis of plants and algae) (Dai et al., 2013).
The biomass yields (TBM) of E. nuttallii, I. aquatic, and O. sativa were estimated by dry matter productivity in unit area (DM) and aquatic plant areas (APA) in each pond. The fresh plant tissues in one m2 area were collected and oven-dried with three replicates in each pond at the end of the experiment, and average dry weight in one m2 area was considered as DM. APA was equal to 60% of total area of each pond. Thus, the TBM could be calculated by the equation: TBM = DM × APA. Trimmed plant tissues were included in the biomass production estimates. As I. aquatic and E. nuttallii has a sprawling growth pattern, it was periodically cut back outside of the rope-restricted area to maintain ∼60% coverage to keep constant aquatic plant coverage (Trimming details see Figs. S1 and S2). Nutrients assimilated by the plants were calculated based on plant biomass and nutrient concentrations. The tissue-mixed plant samples that were collected in different growth stages were used for nutrient concentration measurement. Table S1 showed the average nutrient concentrations of plants.
To assess crab production, mature crabs were harvested using crab traps, and remaining crabs were captured by hand at night, after the pond water was completely drained. Males and females were manually separated and weighed.
Characterization of bacterioplankton communities
As crab rearing is sensitive to high air temperatures (Yuan et al., 2017), the bacterioplankton communities during periods of high temperatures were more interesting. Thus, the bacterioplankton community was assessed on July 2, 2017, when the maximum air temperature reached 37 °C. To obtain sufficient bacterioplankton biomass for community profiling via DNA sequencing, we collected 10 L of water at 20 cm depth from each replicate pond. Water samples were filtered through a 0.22-μm polycarbonate membrane (Millipore, Billerica, MA, USA). DNA was extracted from the filtered biomass using a FastDNA Spin Kit for soil (MP bio, Solon, OH, USA), following the manufacturer’s protocols. Extracted DNA concentration was determined with a NanoDrop 2000 UV–vis spectrophotometer (Thermo Scientific, Wilmington, NC, USA); DNA quality was assessed with gel electrophoresis on a 1% agarose gel. The V4 hypervariable region of the bacterial 16S rRNA gene was amplified using PCR, with the primers 563F (5′-AYTGGGYDTAAAGVG-3′) and 802R (5′-TACNVGGGTATCTAATCC-3′) (Cardenas et al., 2010), following previously described protocols (Wang et al., 2017). The amplified PCR products were purified using an AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA), and quantified using a QuantiFluor-ST kit (Promega, Madison, WI, USA), following the manufacturer’s instructions. Purified amplicons were pooled in equimolar concentrations and sequenced on an Illumina MiSeq platform (Illumina, San Diego, CA, USA) at Majorbio Bio-Pharm Technology Co. Ltd. (Shanghai, China), following standard protocols.
Raw sequence reads were demultiplexed using QIIME (v.1.9.1) (Caporaso et al., 2010a). Barcoding adapters and PCR primers were cleaved using cutadapt (v.1.16) (Martin, 2011). Low-quality reads were removed from the dataset with USEARCH10 (Edgar & Flyvbjerg, 2015), using the “fastq_filter” command with the parameters maxee = 1 and truncqual = 15. The remaining paired-end reads were merged using the “fastq_mergepairs” command in USEARCH10 (Edgar & Flyvbjerg, 2015). Merged read abundances were normalized across samples by randomly subsampling 28,000 sequences from each sample. A zero-radius OTU (zOTU) table was produced from the sequence reads using the Unoise3 algorithm (Edgar, 2018). The taxonomic classification of each zOTU was assigned using the UCLUST algorithm against the Silva (SSU123) 16S rRNA database with default parameters (Caporaso et al., 2010b; Edgar, 2010).
An additional OTU table was generated to predict bacterioplankton functions. The abundance-normalized sequences were clustered at the 97% nucleotide similarity cutoff level into OTUs using the “pick_closed_reference_otus.py” function in QIIME (v.1.9.1) (Caporaso et al., 2010b). OTUs were taxonomically classified using the UCLUST algorithm (Edgar, 2010) against the Greengenes (gg_13_5) reference database (DeSantis et al., 2006) with default parameters. The resulting OTU table was analyzed using PICRUSt (v.1.1.3) (scripts “normalize_by_funtion.py,” “predict_metagenomes.py,” “categorize_by_function.py,” and “metagenome_contributions.py”) (Langille et al., 2013). The PICRUSt algorithm produced a table of functional Kyoto Encyclopedia of Genes and Genomes orthologs (KOs). To obtain OTU-specific counts of genes (Da Fonseca et al., 2019; Fan et al., 2018) associated with nitrification and denitrification, the PICRUSt script “metagenome_contributions.py” with −l option was applied to selected KOs (K00370, K00371, K00374, K02567, K02568, K00368, K15864, K04561, K02305, K00376, K10535, K10944, K10945, and K10946) (Script S1).
Statistical analysis
Statistical analyses were primarily performed in R (v.3.3.2) (R Core Team, 2013). To identify significant differences among treatments, the Levene and Kolmogorov–Smirnov tests were used to check the homogeneity of variances and data normality, respectively. One-way ANOVA was used to determine the significance among the treatments, and Tukey’s HSD test was then applied for multiple comparisons. If the measurement variable did not meet the normality assumption, a Kruskal–Wallis test was performed instead of one-way ANOVA. We considered P < 0.05 to be statistically significant, unless otherwise noted.
The bacterioplankton communities were analyzed using the vegan package (Dixon, 2009) in R. The matrix of zOTU abundances was transformed prior to distance-based analyses using the “decostand” function with the Hellinger method (Legendre & Gallagher, 2001). Principal component analyses were performed using the "rda" function in vegan to visualize differences among the bacterioplankton communities from the different macrophyte systems. Environmental variables were then fitted and projected onto an ordination using “envfit” function in vegan based on 1,000 permutations.
To investigate the correlation of microbial taxa and environmental variables, we constructed a co-occurrence network using CoNet (Faust & Raes, 2016), based on the zOTU table and the environmental variables. In the co-occurrence analysis, the read count matrix was first filtered, and only those zOTUs with at least seven minimum occurrence values across the nine samples were retained. Pair-wise associations among zOTUs and environmental factors were calculated using the Pearson, Spearman, Kendall, Bray–Curtis, and Kullback–Leibler correlation methods. The initial top and bottom edge numbers were set at 1,000. For each edge and each measure of association, 1,000 permutation scores and 1,000 bootstrap scores were computed. The resultant networks were visualized using Cytoscape (v.3.2.1) (Shannon et al., 2003).
Results
System performance, crab yield, and water quality
The macrophyte systems floated steadily throughout the whole experiment despite two moderate windstorms. Thus, the floating systems described here were suitable for plant growth on water surfaces. All of the plants grew well initially (between May and August). However, O. sativa began to go to seed (indicating the start of the reproductive stage) in early August, about 25 days earlier than the normal (September, if O. sativa is grown in a field). After early August, O. sativa plants yellowed and the roots darkened. In contrast, I. aquatic and E. nuttallii grew well until the end of the experiment.
Crab yields among the three macrophyte systems did not differ significantly. The average yields of male and female crabs were ∼740 and ∼267 kg ha−1, respectively (Figs. 2A and 2B). Individual weight is one of the most important factors determining the value of commercial crabs. Here, the individual weight distributions did not differ significantly among plant systems (P > 0.05). The median weights of male and female crabs were 168 and 109 g, respectively (Figs. 2C and 2D).
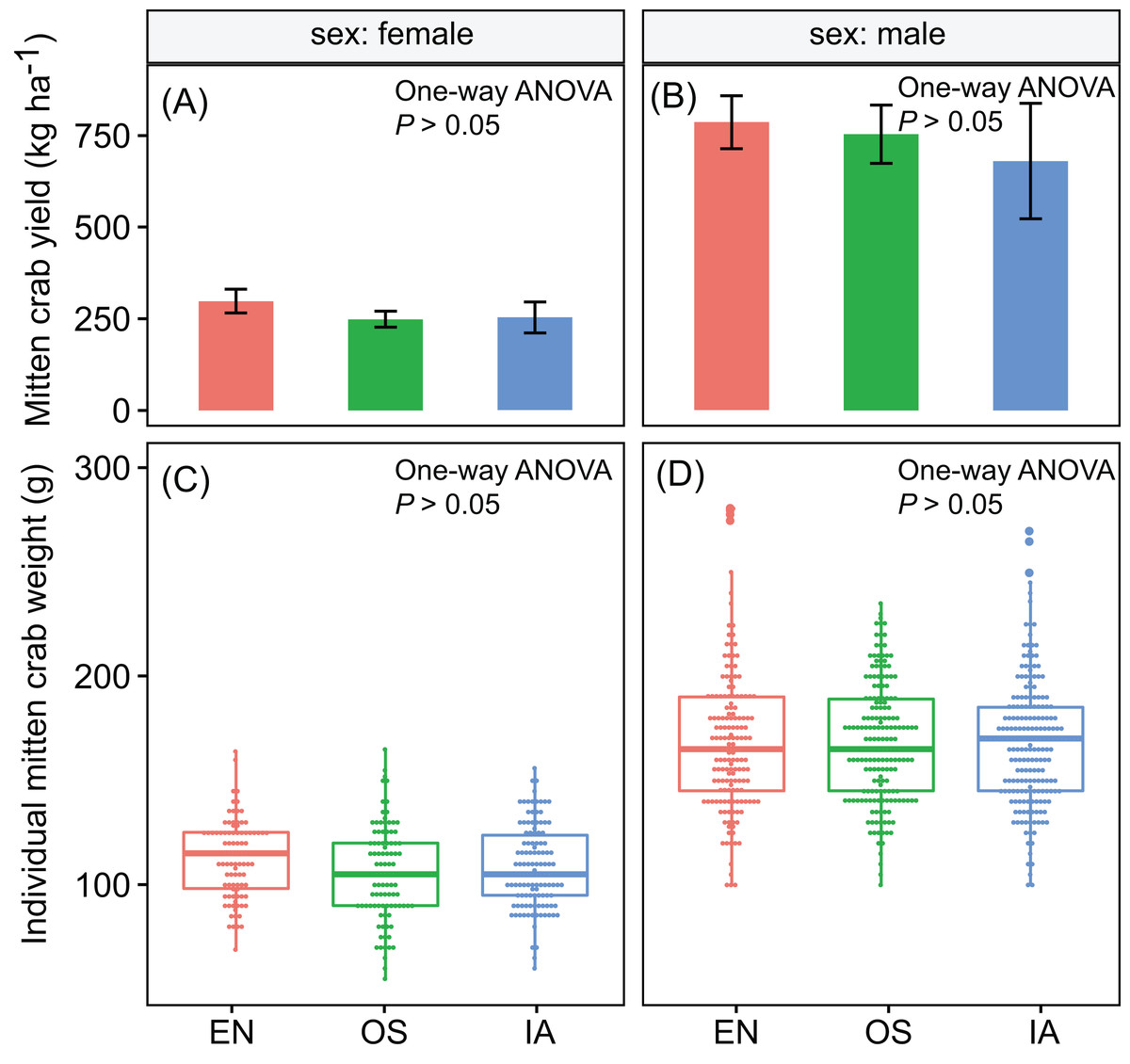
Figure 2: (A–D) the yield and individual weight distributions of the Chinese mitten crabs did not differ significantly among macrophyte systems.
Error bars denote the standard deviations of the mean (n = 3). EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.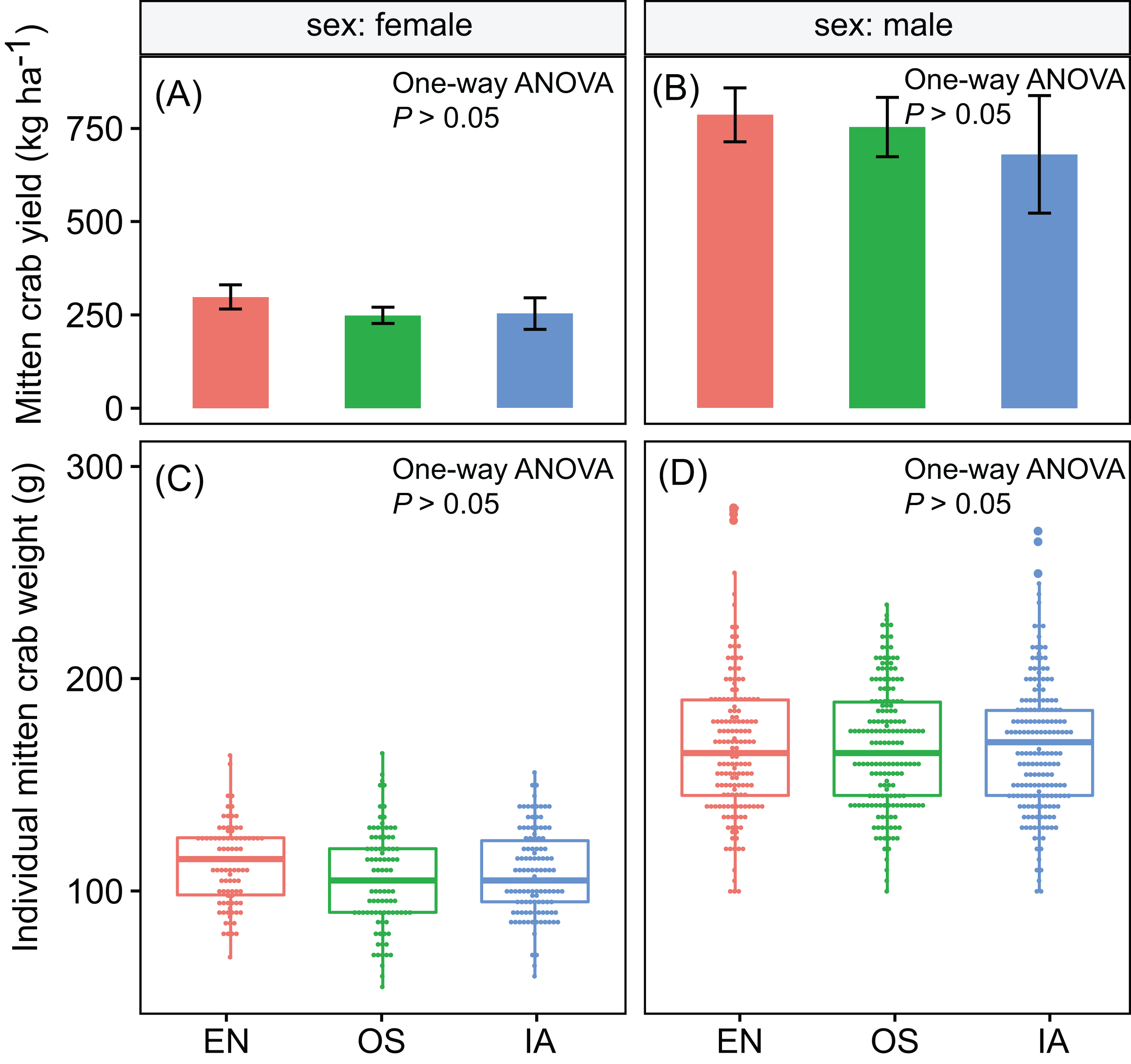{kind=link}
The nitrogen assimilated by I. aquatic was estimated to be 118 kg ha−1, based on macrophyte biomass weight and nutrient concentrations. This level of assimilation was 3.5 times that of E. nuttallii, and 1.5 times that of O. sativa (Fig. 3). In contrast, the phosphorous assimilated by I. aquatic, E. nuttallii, and O. sativa did not differ significantly.
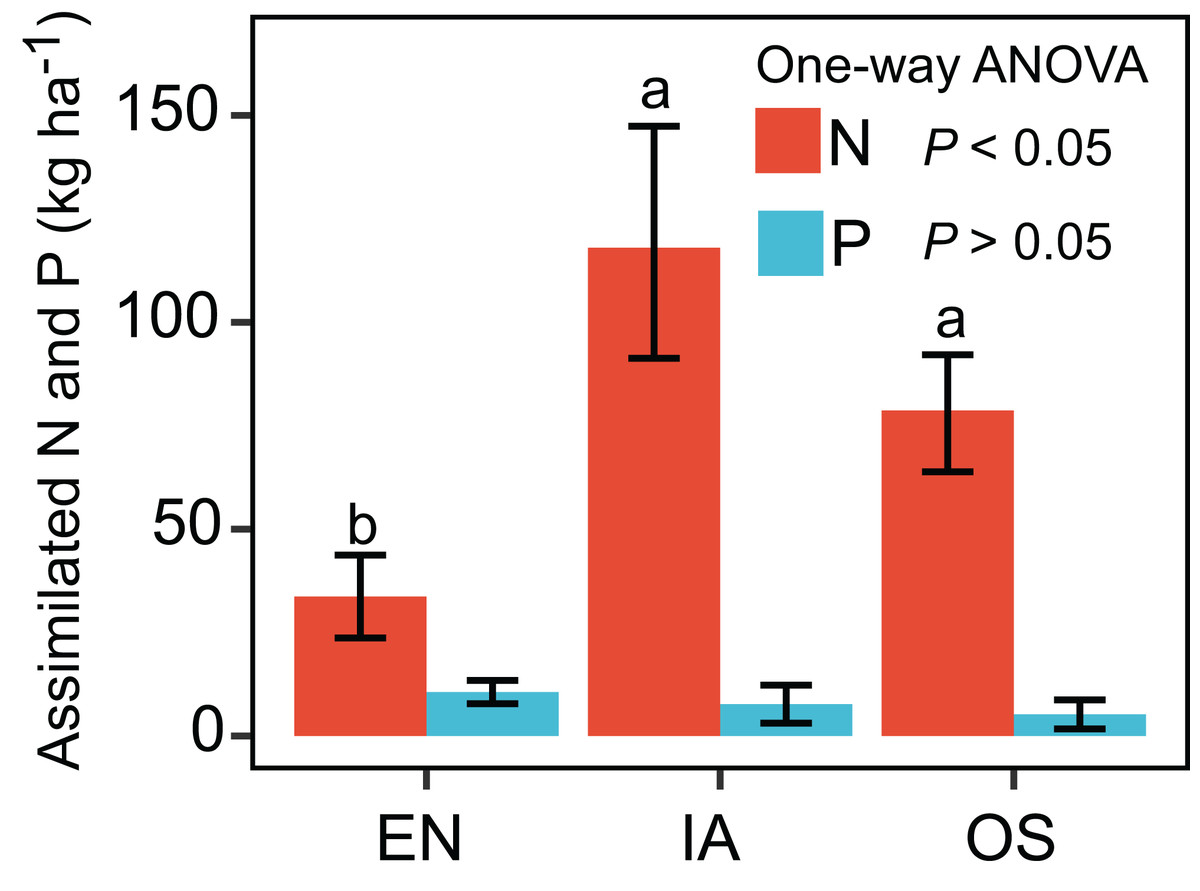
Figure 3: Assimilation of nitrogen and phosphorous by EN, Elodea nuttallii; IA, Ipomoea aquatica; and OS, Oryza sativa.
Different letters above bars represent significant differences among plant systems (P < 0.05; Tukey’s HSD test). Error bars denote the standard deviations of the means (n = 3).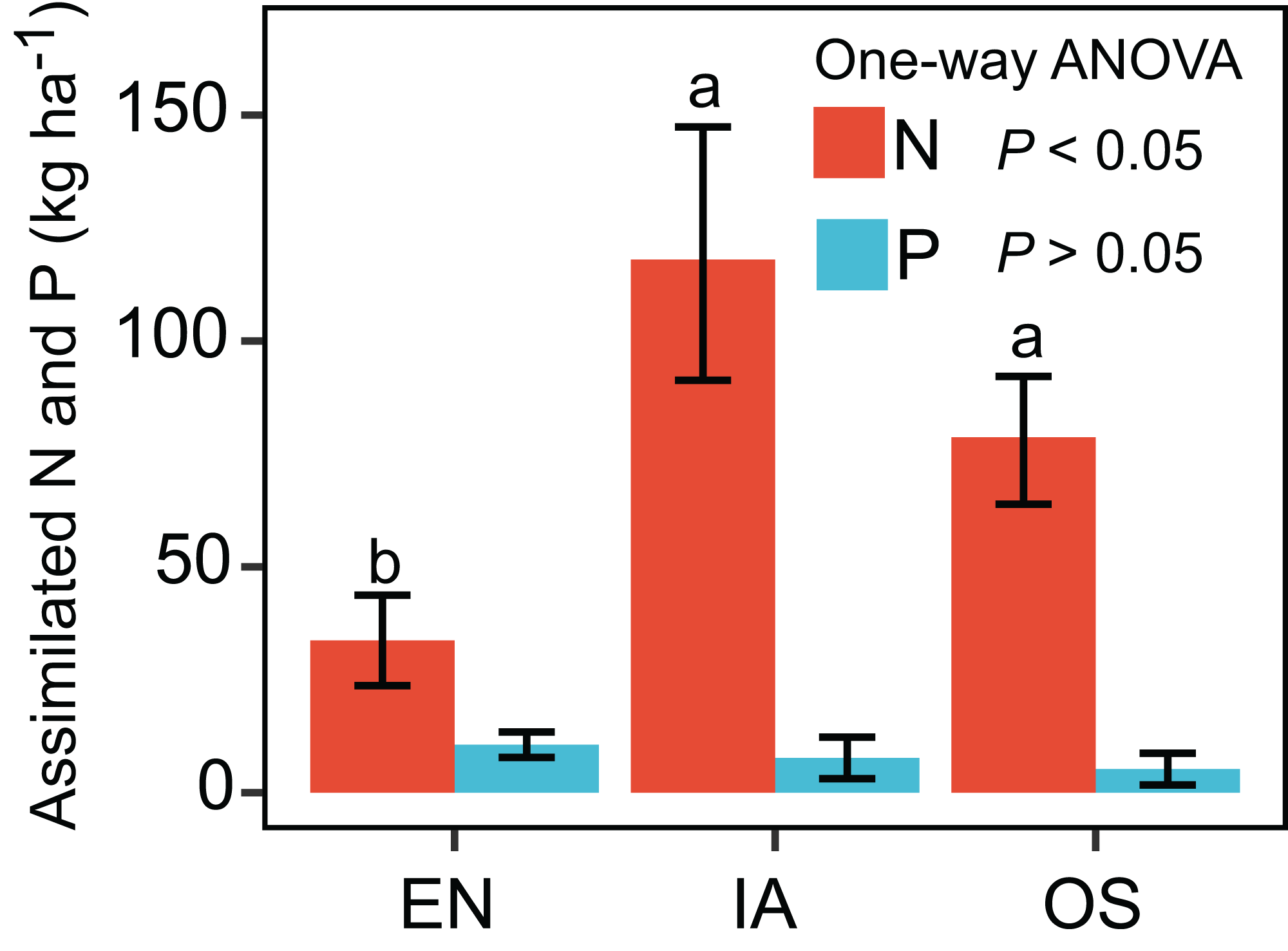{kind=link}
The average TN, TP, NH4+-N, and NO3−-N concentrations of inlet waters in the rearing period were 1.25 ± 0.44 (mean ± SD), 0.03 ± 0.02, 0.25 ± 0.43, and 0.57 ± 0.47 mg L–1, respectively (Fig. 4). The TN content in 31.3% of the samples from the E. nuttallii system exceeded the Environmental Quality Standards for Surface Water (GB3838-2002) limit for type III water. In contrast, the TN content in 9.4% and 15.6% samples from the O. sativa and I. aquatic systems, respectively, exceeded the type III water quality limit (GB3838-2002) (Fig. 4). The number of samples that exceeded the TP and NH4+-N type III water quality limits did not differ substantially among treatments. No samples exceeded the water quality limit for NO3−-N (10 mg L−1). Cumulative curve analysis indicated that amount of TP and NH4+-N accumulated over the course of the experiment (up to November 27) did not differ significantly among different plant systems. However, the levels of accumulated TN and NO3−-N were significantly higher in the E. nuttallii system than in the I. aquatic or O. sativa systems (Fig. 5). Average pH was 7.48 in the I. aquatic system, 8.16 in the E. nuttallii system, and 7.9 in the O. sativa system (Fig. 6).
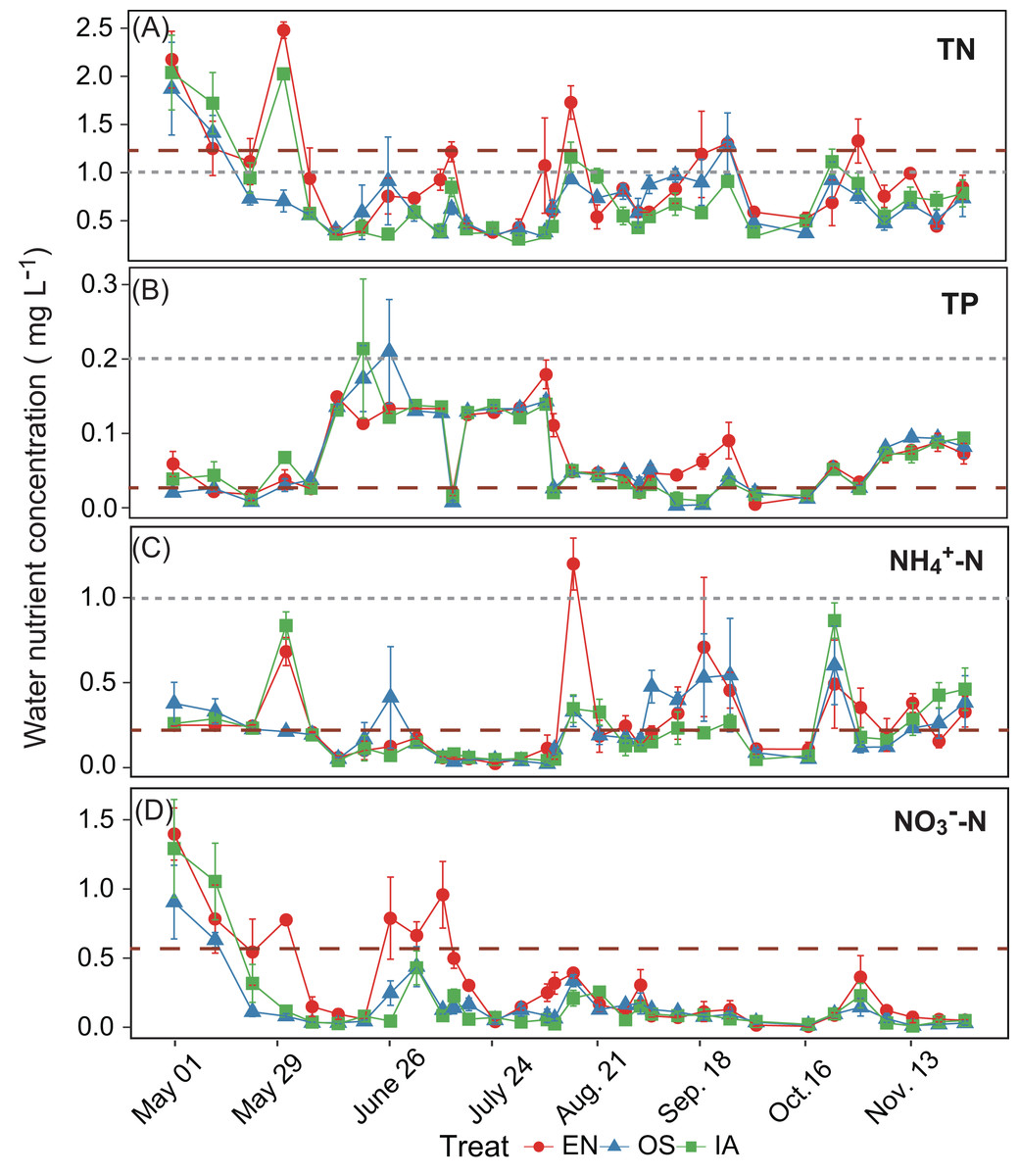
Figure 4: (A–D) separately shows the TN, TP, NH4+-N, and NO3−-N concentrations in the rearing pond.
Error bars denote the standard deviations of the means (n = 3). The brown dashed lines represent the average nutrient concentrations in the inlet waters during the rearing period. The gray dashed lines represent the type III water quality limits from the Chinese Environmental Quality Standard for Surface Water (GB3838-2002). NO3−-N concentrations were all below the type III water quality limit (10 mg L−1). EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.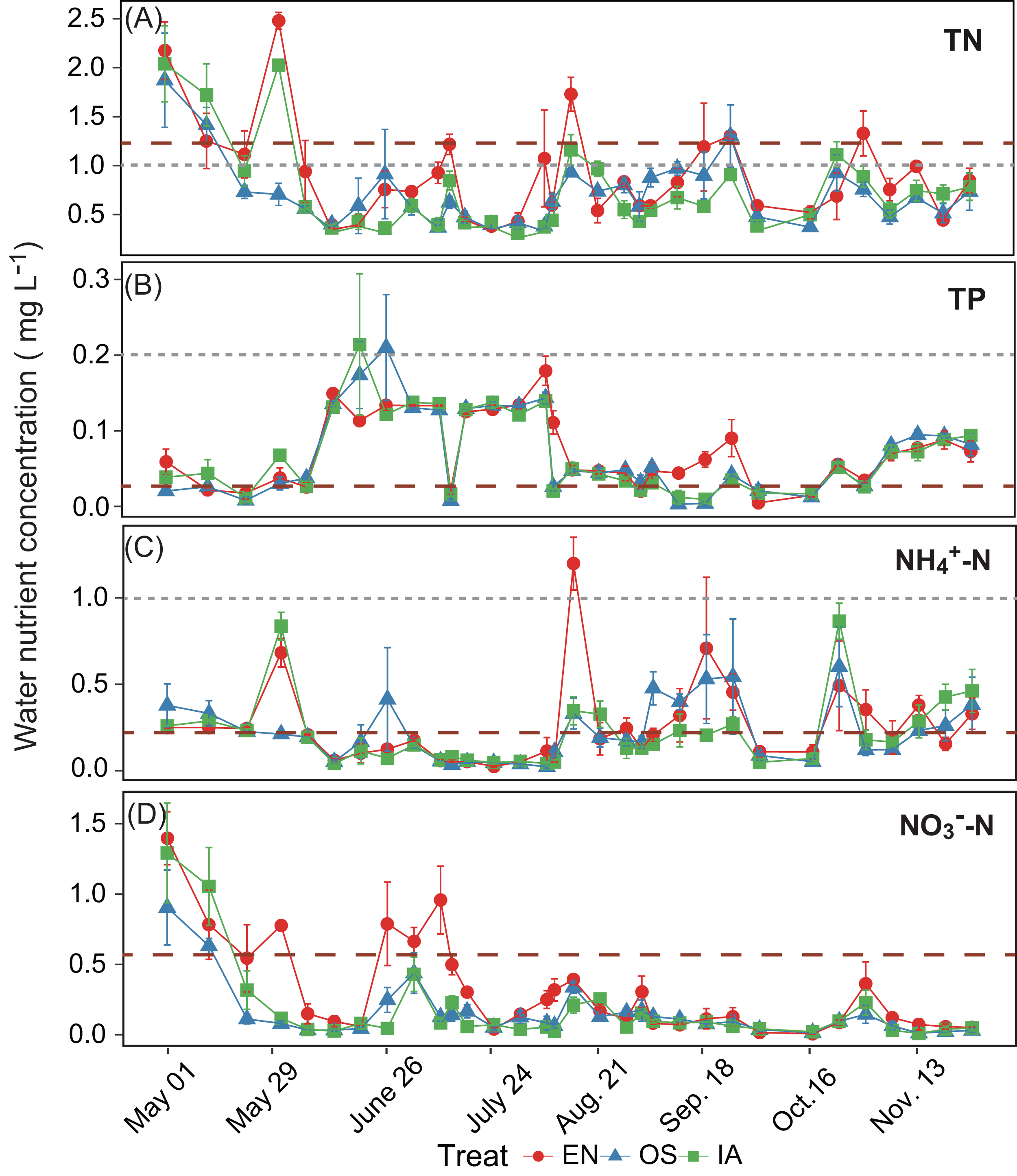{kind=link}

Figure 5: (A–D) separately shows the absolute cumulative TN, TP, NH4+-N, and NO3−-N quantities in rearing period.
The statistical significance of differences in cumulative nutrient quantity among macrophyte systems was determined with a Kruskal–Wallis test (n = 3) at the end of the rearing periods (November 27). Different lowercase letters represent significant differences (P < 0.05; Tukey’s HSD test). EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.{kind=link}

Figure 6: pH of the outflow water samples during the rearing period.
EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.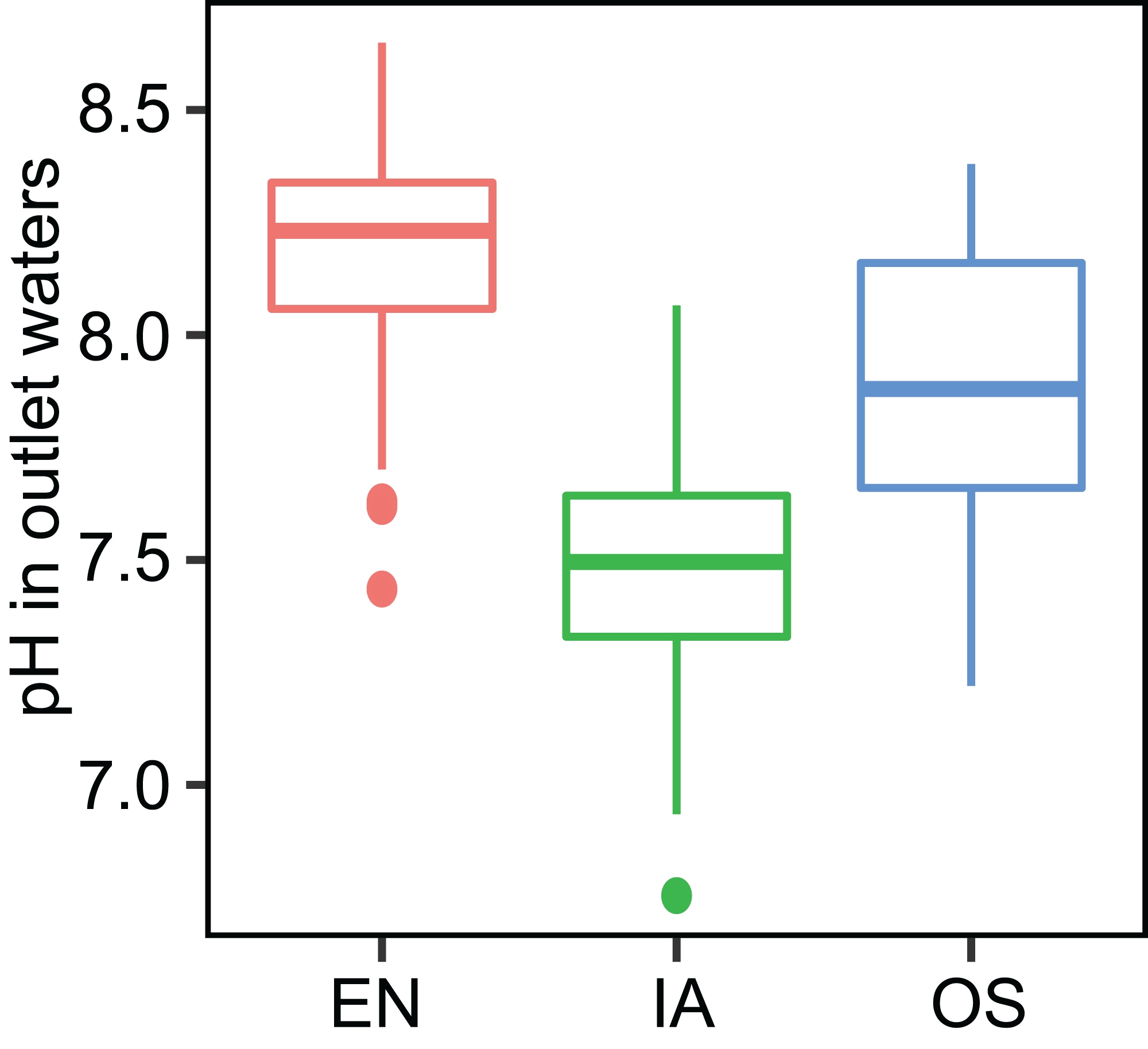{kind=link}
Bacterioplankton communities and predicted functions associated with nitrification/denitrification
The abundances of nearly all of the bacterioplankton phyla differed significantly among macrophyte systems (Fig. 7A). The abundances of Acidobacteria, Chloroflexi, Firmicutes, β-Proteobacteria, and δ-Proteobacteria were significantly higher in the O. sativa and I. aquatic systems as compared to the E. nuttallii system, while the abundances of Actinobacteria, Armatimonadetes, and γ-Proteobacteria were significantly lower. OTU-based diversity (α-diversity) was higher in the O. sativa and I. aquatic systems than in the E. nuttallii system (Fig. 7B), suggesting that bacterioplankton communities were more complex in the O. sativa and I. aquatic systems than in the E. nuttallii system.
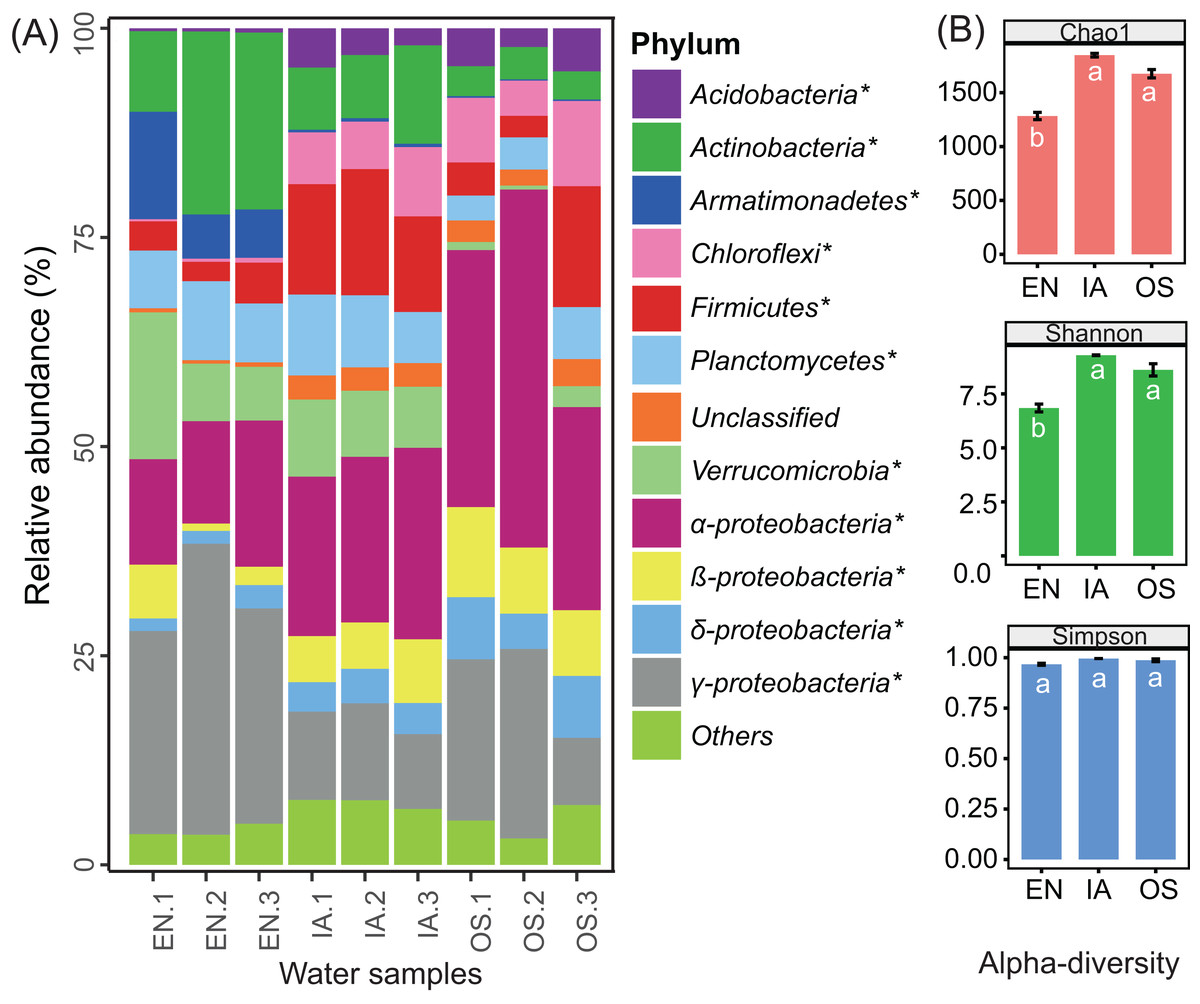
Figure 7: Bacterioplankton abundances and diversity indices of the three macrophytes systems.
(A) The relative abundances of the dominant bacterial phyla in the bacterioplankton communities and (B) the associated alpha diversity. Asterisks indicate significant differences in abundances among the macrophyte systems (P < 0.05 level; one-way ANOVA; n = 3). Error bars denote the standard errors of the means. Different letters above bars represent significant differences among treatments (P < 0.05; Tukey’s HSD test; one-way ANOVA; n = 3). EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.{kind=link}
Principal component analyse indicated that the bacterioplankton communities were discrete among the different plants, with the first two axes explaining 83.67% of the total variation (Fig. 8A). After fitting environmental variables (Table S2) and bacterioplankton community ordination, we found that pH was significantly associated with community differences (P < 0.05) (Fig. 8A). The co-occurrence network analysis also identified pH as the only environmental factor that co-varied with certain taxa. These pH-correlated taxa included zOTUs belonging to the phyla Acidobacteria, Chloroflexi, Firmicutes, Proteobacteria, and Planctomycetes (Fig. 8B). The total average abundance of co-occurring zOTUs was 7.7%, suggesting that pH influenced the relative abundance of the most abundant members of each community.
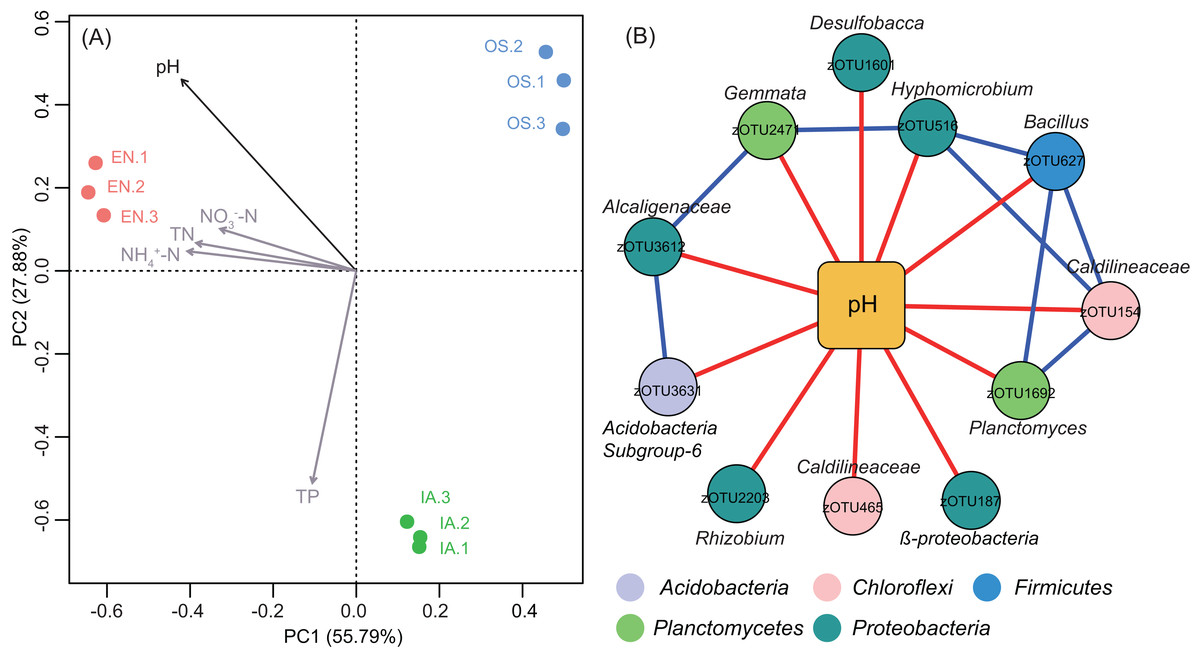
Figure 8: Bacterioplankton community structure and environmental factors.
(A) Principal component analysis (PCA) of bacterioplankton community composition and environmental variables. The significant environmental variable (P < 0.05) is shown with a black arrow; other factors are shown in gray. EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica. (B) Subnetwork showing the correlation between bacterial taxa and pH. Squared nodes correspond to environmental parameters and circle nodes correspond to zOTUs. Circle nodes not assignable to genus are labeled with the names of higher taxonomic ranks, and node colors represent phyla. Red and blue colors of edge represent negative and positive correlations, respectively.{kind=link}
Potential functions of the bacterioplankton communities were predicted using PICRUSt. We focused specifically on functional genes associated with nitrification and denitrification, as these processes are highly related to nitrogen cycling and are likely to affect nitrogen concentration in pond water. Genes associated with all of the steps of denitrification (napA, napB, nirK, norB, norC, and nosZ; Kanehisa et al., 2017) were generally more abundant in the I. aquatic and/or O. sativa systems than in the E. nuttallii system. However, the abundances of narI, narG, and narH, which are only involved in the reduction of nitrate to nitrite (Kanehisa et al., 2017), were lower in the I. aquatic and/or O. sativa systems (Fig. 9). The abundances of the nitrifying genes pmoA-amoA, pmoB-amoB, and pmoC-amoC, which are involved in the oxidation of ammonia to hydroxylamine (Kanehisa et al., 2017), were significantly higher in the O. sativa system than in the I. aquatic and E. nuttallii systems. The abundance of hao was not significantly different among the three macrophyte systems, although the abundances of narG and narH were lower in the I. aquatic and O. sativa systems than in the E. nuttallii system (Fig. 9).
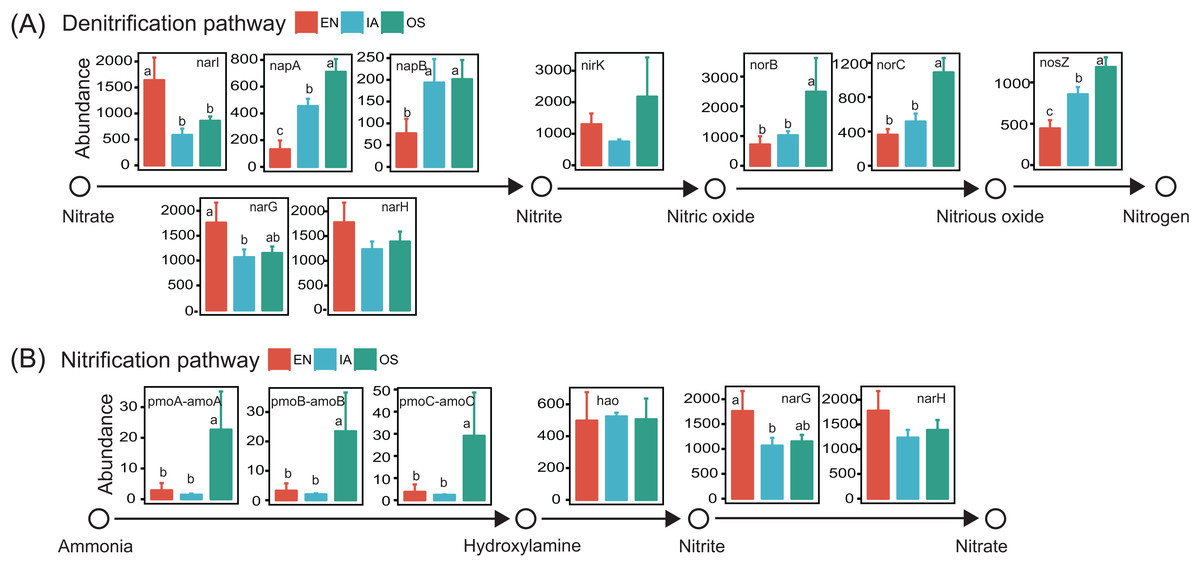
Figure 9: The abundances of the bacterioplankton genes associated with nitrification and denitrification among macrophyte systems, as predicted by PICRUSt.
(A) and (B) shows the denitrification and nitrification predicted gene abundances, respectively. Error bars denote the standard deviations of the means (n = 3). Different lowercase letters above bars represent statistically significant differences (P < 0.05; one-way ANOVA followed by Tukey’s HSD tests). EN, Elodea nuttallii; OS, Oryza sativa; IA, Ipomoea aquatica.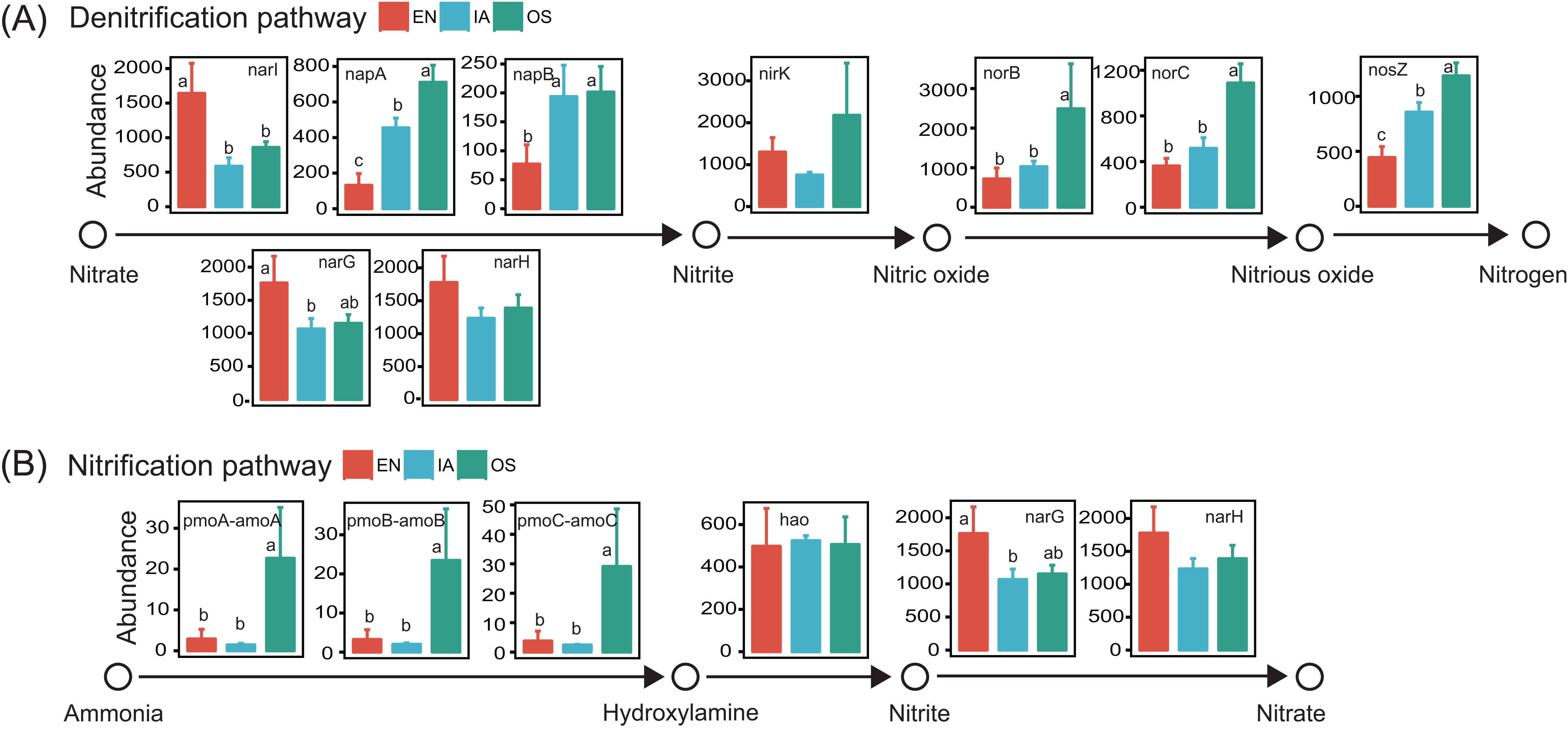{kind=link}
Discussion
The macrophyte system described here floated steadily throughout the whole experiment. In addition, mitten crab yield and quality did not differ significantly among plant systems (Fig. 2). Thus, our results demonstrated that local plants were a feasible alternative to E. nuttallii. Indeed, our data indicated that I. aquatic was be the best local replacement for E. nuttallii, as this plant grew well in ponds and provided sufficient shade. In contrast, O. sativa is not a suitable alternative to E. nuttallii, as it stopped growing in the middle of the experimental period. The mechanisms underlying the different performances of O. sativa and I. aquatic were related to the different growth behaviors and nutrient requirements of the two species. O. sativa has several distinctive growth stages, including tillering, heading, ripening and et al. (Zhang et al., 2018), which typically require different levels of nutrients (Fairhurst et al., 2007). For example, during the tillering stage, O. sativa biomass increases much faster than during other stages of O. sativa growth, and consequently requires a more intensive nutrient supply (Nawaz & Farooq, 2017). If the nutrient supply needs are not met (i.e., due to low nutrient concentrations in the water), many tillers become non-productive, and the few remaining productive tillers enter the reproductive stage earlier than normal (Nawaz & Farooq, 2017). In submerged paddy fields, the intensity of nutrient supply is manipulated by top-dressing fertilizers (Fairhurst et al., 2007). However, this technique cannot be used in crab-rearing ponds because it would pollute the water.
Unlike O. sativa, I. aquatic does not exhibit obvious physiological differences among growth stages. I. aquatic has sprawling growth, with biomass increasing over rearing time (from May to November) (Shaltout, Al-Sodany & Eid, 2010). Thus, low concentrations of nutrients in the water are sufficient. We consequently concluded that I. aquatic is an attractive macrophytic alternative to E. nuttallii in crab-rearing ponds.
Based on the Chinese national water quality standards (GB3838-2002) (State Environmental Protection Administration (SEPA), 2002), 31.3% of the outflow samples from the E. nuttallii system contained TN levels in excess of limits for type III waters (Fig. 4). Thus, the outflow from ponds using the E. nuttallii system is likely to pollute the downstream environment. Fewer outflow samples from the I. aquatic and O. sativa systems, as compared to the E. nuttallii system, had TN levels above the type III water quality limits. These data suggest I. aquatic or O. sativa systems would generate less environmental pollution than E. nuttallii systems. This suggestion was reinforced by the absolute cumulative nutrient quantities in outflow samples (Fig. 5), which showed that the TN and NO3−-N concentrations were significantly higher in the E. nuttallii system, as compared to the I. aquatic and O. sativa systems. Indeed, I. aquatic and O. sativa assimilated more nitrogen from the water than did E. nuttallii (Fig. 3). Thus, it was possible that I. aquatic and O. sativa more effectively improved water quality than E. nuttallii, which is a desirable property for macrophytes grown in crab-rearing ponds. An alternative explanation was that the differences in water quality among plant systems were a result of the activities of macrophyte-specific bacterioplankton communities.
Bacterioplankton communities differed significantly among the three macrophyte systems (Fig. 6), possibly because of the different root deposits produced by the three plants. Terrestrial and macrophytic plants release unique root exudates that drastically alter bacterioplankton community structure (Baudoin, Benizri & Guckert, 2003; Casamatta & Wickstrom, 2000; Nelson et al., 2013; Tanaka et al., 2012; Zhao et al., 2013). Bacterioplankton community composition might also be regulated by pH, as pH was the only environmental factor that was significantly associated with the relative abundances of specific bacterioplankton taxa (Fig. 8). These results were consistent with previous reports, which indicated that pH affects the community structures of both terrestrial bacteria (Fierer & Jackson, 2006; Lauber et al., 2009; Rousk et al., 2010) and bacterioplankton (Ren et al., 2015). Moreover, the significant increases in bacterioplankton diversity with lower water nutrients in the I. aquatic and O. sativa treatments (Figs. 5 and 7B) indicated that biodiversity helped improve water quality, as observed in other aquatic systems (Cardinale, 2011; Gregoracci et al., 2012).
The predicted functions of the bacterioplankton community also differed among the macrophyte systems, particularly those related to nitrification and denitrification. Most denitrifying genes, especially those associated with the reduction of nitrite, nitric oxide, and nitrous oxide, were more abundant in the I. aquatic and O. sativa bacterioplankton communities, as compared to the E. nuttallii bacterioplankton community (Fig. 9). Moreover, abiotic environmental factors usually control the denitrification process, that is, pH, temperature, and organic carbons (OC). The I. aquatic and O. sativa systems that could provide labile OC derived from root exudates (Wang et al., 2018; Xu et al., 2008; Zhu & Cheng, 2011) favored denitrification. Overall, the biotic and abiotic influences on denitrification in the I. aquatic and O. sativa systems were thought to be co-occurring, consistent with the conclusion in riparian wetlands (Xiong et al., 2017). In addition, concentrations of NO3−-N (and TN) were lower in the I. aquatic and O. sativa systems than in the E. nuttallii system. Thus, the I. aquatic and O. sativa systems might have a higher denitrification potential than the E. nuttallii system. Meanwhile, there were no obvious differences in the abundance patterns of nitrification-associated genes among macrophyte systems. The genes responsible for the oxidation of ammonium to hydroxylamine (pmoA-amoA, pmoB-amoB, and pmoC-amoC) were significantly more abundant in the O. sativa system (but not the I. aquatic system) as compared to the E. nuttallii system, but the abundances of these genes were relatively low across all of the plant systems, with only ∼2–20 copies per sample. In addition, the gene abundances in nitrification process usually could not accurately predict nitrification potential, which can be affected by other abiotic factors (Francis et al., 2005; Rocca et al., 2015; Yao et al., 2018). Thus, nitrification potential may not vary significantly among these three macrophytes, and further studies are required to validate these coupled nitrification–denitrification processes in future.
Conclusions
Elodea nuttallii is routinely cultivated in ponds used for mitten crab aquaculture. However, E. nuttallii temperature sensitivity often leads to plant deterioration and decreased water quality at high ambient temperatures, negatively affecting crab production. Here, we successfully designed a floating system to support the growth of I. aquatic and O. sativa on the surfaces of crab-rearing ponds. We then compared the crab yield, outflow water quality, and bacterioplankton communities among ponds with E. nuttallii, I. aquatic, and O. sativa macrophyte systems. Our results indicated that I. aquatic growth behavior was preferable to that of O. sativa. Crab yields did not differ significantly among systems. Moreover, outflow water quality, as indicated by TN and NO3−-N concentrations, was better in the I. aquatic and O. sativa systems than in the E. nuttallii system, due to the greater nitrogen assimilation of I. aquatic and O. sativa as compared to E. nuttallii. In addition, the microbial communities associated with I. aquatic and O. sativa had a greater denitrification potential than the microbial community associated with E. nuttallii. Thus, our results indicated mitten crabs could be successfully reared using native aquatic plants. Specifically, I. aquatic was a suitable and environmental-friendly replacement for E. nuttallii, but O. sativa was not.
Supplemental Information
DO concentrations measured in preliminary research in 2016.
DO concentrations measured in preliminary research in 2016. E. nuttallii, Elodea nuttallii; O. sativa, Oryza sativa; I. aquatic, Ipomoea aquatica. Different small letters on the box represent significant differences between treatments at 0.05 levels (Tukey HSD).
Macrophytes trimming methods in Chinese mitten crab ponds.
Figure S2. Planform of rearing ponds in this study. The frame is consisted of rope and plastic balls. The plants that grow out of frames would be cut.
Figure S3. The demo graph of E. nuttallii (Elodea nuttallii) trimming. The frame is consisted of rope and plastic balls. The stems and leaves of E. nuttallii that grow out of frames were trimmed manually using sickle, and just short stems (less than 1–2 cm) and roots of E. nuttallii were left.
Average nutrient concentrations of plants.
The tissue-mixed plant samples that collected in different growth stages were used for nutrient concentration measurement. Nitrogen and phosphorus contents are determined by the Kjeldahl Nitrogen Determination method and the molybdate-ascorbic acid method, respectively.
Water nutrient concentrations in July 2.
Different small letters in the same column represent significant differences among treatments.
The denitrification and nitrification contribution prediction.
The denitrification and nitrification contribution prediction using PICRUSt1.1.3.
Raw records of mitten crab yield and individual weight applied for preparation for Fig. 2.
Raw data records of mitten crab yield and individual weight applied for preparation for Fig. 2.
Raw data of assimilated nutrient contents of different macrophyte plants applied for preparation for Fig. 3.
Raw data of nutrient contents of different macrophyte plants applied for preparation for Fig. 3.
Raw data exported from the auto analyzer (SKALAR SAN++) and a WTW portable pH meter applied for reparation for Figs. 4–6 in the rearing period.
Raw data exported from the auto analyzer (SKALAR SAN++) and a WTW portable pH meter applied for reparation for Figs. 4–6 in the rearing period.
Raw data exported from USEARCH10 applied for data analyses and preparation for Figs. 7 and 8.
Raw data exported from USEARCH10 applied for data analyses and preparation for Figs. 7 and 8.