
In silico analysis reveals a shared immune signature in CASP8-mutated carcinomas with varying correlations to prognosis
- Published
- Accepted
- Received
- Academic Editor
- Xiangqin Cui
- Subject Areas
- Bioinformatics, Genomics, Immunology, Oncology
- Keywords
- CASP8, HNSC, Necroptosis, UCEC, Inflammation, IL33, Neutrophils, TCGA
- Copyright
- © 2019 Ghanekar and Sadasivam
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. In silico analysis reveals a shared immune signature in CASP8-mutated carcinomas with varying correlations to prognosis. PeerJ 7:e6402 https://doi.org/10.7717/peerj.6402
Abstract
Background
Sequencing studies across multiple cancers continue to reveal mutations and genes involved in the pathobiology of these cancers. Exome sequencing of oral cancers, a subset of Head and Neck Squamous cell Carcinomas (HNSCs) common among tobacco-chewing populations, revealed that ∼34% of the affected patients harbor mutations in the CASP8 gene. Uterine Corpus Endometrial Carcinoma (UCEC) is another cancer where ∼10% cases harbor CASP8 mutations. Caspase-8, the protease encoded by CASP8 gene, plays a dual role in programmed cell death, which in turn has an important role in tumor cell death and drug resistance. CASP8 is a protease required for the extrinsic pathway of apoptosis and is also a negative regulator of necroptosis. Using multiple tools such as differential gene expression, gene set enrichment, gene ontology, in silico immune cell estimates, and survival analyses to mine data in The Cancer Genome Atlas, we compared the molecular features and survival of these carcinomas with and without CASP8 mutations.
Results
Differential gene expression followed by gene set enrichment analysis showed that HNSCs with CASP8 mutations displayed a prominent signature of genes involved in immune response and inflammation. Analysis of abundance estimates of immune cells in these tumors further revealed that mutant-CASP8 HNSCs were rich in immune cell infiltrates. However, in contrast to Human Papilloma Virus-positive HNSCs that also exhibit high immune cell infiltration, which in turn is correlated with better overall survival, HNSC patients with mutant-CASP8 tumors did not display any survival advantage. Similar analyses of UCECs revealed that while UCECs with CASP8 mutations also displayed an immune signature, they had better overall survival, in contrast to the HNSC scenario. There was also a significant up-regulation of neutrophils (p-value = 0.0001638) as well as high levels of IL33 mRNA (p-value = 7.63747E−08) in mutant-CASP8 HNSCs, which were not observed in mutant-CASP8 UCECs.
Conclusions
These results suggested that carcinomas with mutant CASP8 have broadly similar immune signatures albeit with different effects on survival. We hypothesize that subtle tissue-dependent differences could influence survival by modifying the micro-environment of mutant-CASP8 carcinomas. High neutrophil numbers, a well-known negative prognosticator in HNSCs, and/or high IL33 levels may be some of the factors affecting survival of mutant-CASP8 cases.
Introduction
Exome sequencing, RNA-sequencing, and copy number variation analysis of different cancers have revealed a cornucopia of disease-relevant mutations and altered pathways (Cancer Genome Atlas Research Network et al., 2013b). The identified genes included those with broad relevance across different cancers, as well as those relevant in one or few cancer types. The next phase will involve parsing this voluminous data to generate ideas and hypotheses with the potential for clinical impact, and then testing them experimentally.
We are particularly interested in the heterogeneous group of Head and Neck Squamous cell Carcinomas (HNSCs) as these account for a large number of mortalities each year in the Indian subcontinent (Ferlay et al., 2010; Gupta et al., 2011). Multiple exome sequencing studies have revealed the landscape of recurrent somatic mutations in HNSCs and its prevalent subtype of Oral Squamous Cell Carcinomas (OSCCs) (Agrawal et al., 2011; India Project Team of the International Cancer Genome Consortium, 2013; Pickering et al., 2013; Stransky et al., 2011; Hayes et al., 2016). While TP53 was the most significant recurrently mutated gene in this cancer type, several other genes such as CASP8, FAT1, and NOTCH1 were also unearthed as significantly recurrently mutated by these large-scale sequencing studies. Barring TP53, the roles of these genes in oral epithelium homeostasis, and how this is altered owing to their mutation in cancer remain to be fully elucidated (Rothenberg & Ellisen, 2012). In this study, we chose to focus on the CASP8 gene, which is mutated in ∼10% of all HNSC cases, and more specifically in 34% of cases with OSCC of the gingiva-buccal sulcus (OSCC-GB), the subtype that accounts for the majority of HNSC cases in the Indian subcontinent (Agrawal et al., 2011; Stransky et al., 2011; Hayes et al., 2016). The types of mutations in CASP8 reported in these HNSC cases included loss of function due to frameshift, nonsense mutation or splice mutation as well as missense and deletion mutations.
Apart from HNSC, Uterine Corpus Endometrial Carcinoma (UCECs) carried the most numbers of mutations in the CASP8 gene, as was observed upon searching the Genomic Data Commons (Grossman et al., 2016). We found that CASP8 was recurrently mutated in about 10% of UCEC cases. Here again, the role of CASP8 in endometrial tissue homeostasis, and how this is altered owing to its mutation in UCEC remains unclear. CASP8 was also mutated in other cancer types, however, the numbers of such tumors are too low for meaningful analyses. Thus, using the sequencing data on 528 head and neck, and 560 uterine corpus endometrial carcinoma tumors available in The Cancer Genome Atlas (TCGA) (Cancer Genome Atlas Network, 2015; Cancer Genome Atlas Research Network et al., 2013a), we sought to identify distinctive features of mutant-CASP8 tumors.
CASP8 regulates two pathways of programmed cell death; it is a key protease required for the initiation of the extrinsic apoptotic pathway that is targeted by some drug-resistant tumors, and it is an important negative regulator of necroptosis (Pasparakis & Vandenabeele, 2015; Feltham, Vince & Lawlor, 2017; Günther et al., 2011; Weinlich et al., 2013). Loss-of-function mutations in CASP8 could lead to reduced apoptosis and promote tumor survival (Salvesen & Walsh, 2014). It could also lead to enhanced necroptosis and promote tumor cell death (Günther et al., 2011; Weinlich et al., 2013). Interestingly, it has been proposed that the necroptotic pathway could be utilized to develop anti-cancer treatments for countering cancers with resistance to apoptosis (Su et al., 2016). At least four HNSC-associated CASP8 mutations have been reported to inhibit activation of the extrinsic apoptosis pathway suggesting loss-of-function, however necroptosis was not analyzed in this study (Li et al., 2014). On the background of these observations, tumors harboring CASP8 mutations offer a tractable, physiologically relevant opportunity to understand the changes brought about by CASP8 mutation, how it affects survival, and if CASP8 or the necroptotic pathway could be a potential drug target.
In this study, we describe the comparison of RNA-sequencing (RNA-seq) data from head and neck squamous cell carcinoma, and later from uterine corpus endometrial carcinoma, that are mutant or wild type for CASP8. We report distinctive molecular features of mutant-CASP8 HNSCs and UCECs that this comparison revealed. In addition, we describe results obtained by correlating these features to overall survival in the affected patients.
Materials and Methods
Differential gene expression analysis of wild-type-CASP8 and mutant-CASP8 cases
Data for 528 head and neck squamous cell carcinoma (HNSC) cases available at The Cancer Genome Atlas (TCGA) were downloaded in May–June 2017 from https://portal.gdc.cancer.gov/. Clinical data files, Mutation Annotation Format (MAF) files, and mRNA quantification files such as HT-Seq files (files with number of reads aligning to each protein-coding gene) and FPKM-UQ files (files with number of fragments aligning per kilobase of transcript per million mapped reads normalized to upper quartile) were downloaded. The HPV status of HNSC cases at TCGA has been reported earlier (Chakravarthy et al., 2016), and these data were used to assign HPV-positive and HPV-negative cases.
Cases with and without CASP8 mutation were selected as shown in Fig. 1. CASP8 mutations in HNSC cases were identified using the Mutation Annotation Format (MAF) files available at TCGA. The workflow for somatic mutation calling at TCGA uses four different pipelines: SomaticSniper, MuSE, MuTect2, and VarScan2. The variants called by these four pipelines are further annotated to infer the biological context of each variant using Variant Effect Predictor (VEP). VEP predicts the effect of variants based on its location and information from databases such as GENCODE, sift, ESP, polyphen, dbSNP, Ensembl genebuild, Ensembl regbuild, HGMD and ClinVar. This annotation results in a list of variants with three predicted effects; high impact variants arising from frame-shift or nonsense mutations, variants with moderate impact which include missense mutations and low impact which include variants with synonymous mutations. The information regarding the impact of mutations was available in the MAF file from each somatic mutation calling pipeline employed by TCGA.

Figure 1: A flowchart indicating the sequence of processes used to select the HNSC cases used in this study.
HNSC cases with CASP8 mutation were identified using MAF files from TCGA. Out of 528 HNSC cases available at TCGA, 55 cases had mutations in CASP8. All cases with CASP8 mutation were HPV-negative. Hence, HPV-negative wild-type cases were considered for use as control. In addition, as CASP8-MT cases occurred in specific subsites in oral cavity (Table S1), CASP8-WT cases from these same subsites were selected as control. Thus, 369 HNSC cases with wild-type-CASP8 were selected as control. Gene expression data was available for 53 cases with CASP8 mutations and 354 cases with wild type CASP8. Data from HT-Seq files of selected cases with CASP8 mutation and corresponding wild-type control cases was analyzed using edgeR to identify genes that were differentially expressed in CASP8-MT HNSCs as compared to CASP8-WT. DEGs, Differentially Expressed Genes; FDR, False Discovery Rate.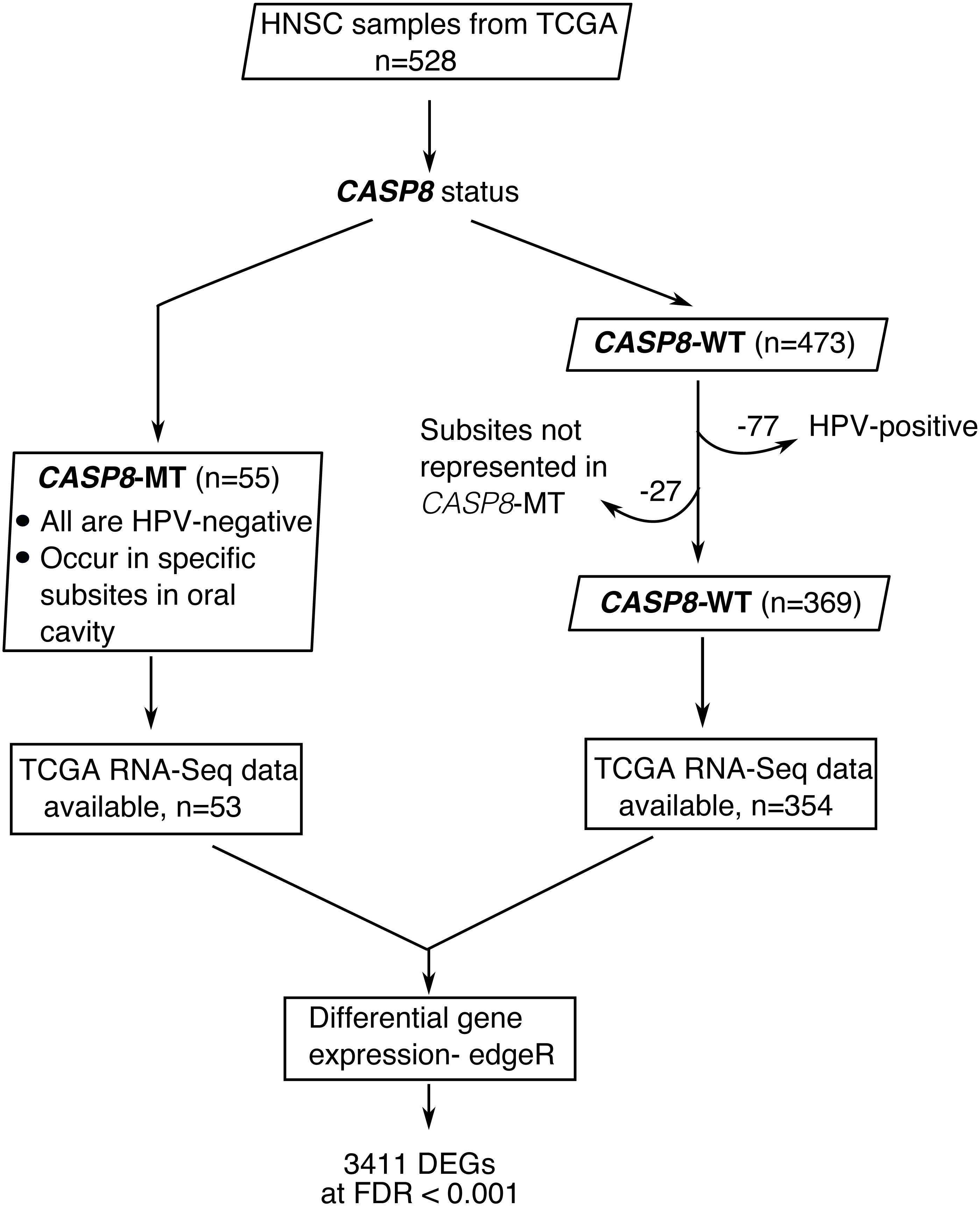{kind=link}
Fifty-five HNSC cases with non-synonymous CASP8 mutations were identified from MAF files. Notably, the majority (80%) of the identified CASP8 mutations were predicted by more than one somatic mutation calling pipeline, and all had either high or moderate impact on function. All cases with CASP8 mutation were HPV-negative and were found in tumors at specific sites in oral cavity. Therefore, HNSC cases that were from these same subsites and were HPV-negative were used as wild type control. A total of 424 HNSC cases of which 369 had wild-type-CASP8 (CASP8-WT) and 55 had mutant-CASP8 (CASP8-MT) were thus selected (Table S1). Among selected cases, RNA-seq data was available for 354 cases with wild-type-CASP8 and 53 cases with mutant-CASP8.
Transcripts that were differentially expressed in CASP8-MT as compared to CASP8-WT cases were identified using edgeR (Robinson, McCarthy & Smyth, 2010). edgeR uses raw read counts as input, which were obtained from HT-Seq files. The analysis was performed using quantile-adjusted conditional maximum likelihood (qCML) method without any filters. All transcripts with FDR < 0.001 and showing a fold-change of at least 2.5-fold (logFC of 1.3) were deemed to be significantly differentially expressed.
Similarly, clinical data and HT-Seq files for 560 uterine corpus endometrial carcinoma (UCEC) cases available at TCGA were downloaded in February 2018. CASP8 mutations of high or moderate impact were present in 56 UCEC cases. Cases without CASP8 mutations were used as wild type control. RNA-seq data was available for 476 CASP8-WT tumors and 56 CASP8-MT tumors. Transcripts that were differentially expressed in CASP8-MT as compared to CASP8-WT cases were identified using edgeR as described for differential gene expression analysis of HNSC.
Gene Ontology and Gene Set Enrichment Analysis (GSEA)
Enrichment analysis was performed at http://geneontology.org/ to identify biological processes overrepresented among transcripts that were differentially expressed between CASP8-WT and CASP8-MT HNSCs (The Gene Ontology Consortium, 2017). Genes that passed the following criteria: (a) FDR < 0.001 (b) FDR < 0.001 and log2FC < −1.3, (c) FDR < 0.001 and log2FC > 1.3, (d) b and c merged, were used to create input gene sets for gene ontology analysis performed using PANTHER version 13.1 (release 2018-02-03). The Binomial test was used to determine statistical significance and the Bonferroni correction for multiple testing was applied.
GSEA was performed using a pre-ranked gene list and hallmark gene sets available at the Molecular Signature Database (Subramanian et al., 2005). The hallmark gene sets use either HGNC or entrez gene ids as the gene identifier. Out of the 60,483 transcripts analyzed by edgeR, HGNC gene symbols could be assigned to 36,095. logFC values for these 36,095 transcripts from the edgeR output from HNSC and UCEC differential gene expression analyses were used to generate the pre-ranked gene list. Gene sets with FDR < 25% and with distinct enrichment at the beginning or end of the ranked list (as observed in enrichment plots) were taken to be significantly enriched gene sets. To perform GSEA with genes that were up-regulated in the skin of mice lacking functional Caspase-8, the top 100 up-regulated genes that were reported were selected (Kovalenko et al., 2009). Of these 100 genes, 80 genes had corresponding human orthologs (CASP8-KOSET) as identified using tools available at http://www.informatics.jax.org/. GSEA was then performed using the pre-ranked gene list and the CASP8-KOSET.
Immune cell infiltration in HNSC and UCEC cases
The abundance estimates of six immune cell types; B cells, CD4+ T cells, CD8+ T cells, Neutrophils, Macrophages, and Dendritic cells in TCGA cases was downloaded from Tumor IMmune Estimation Resource (TIMER) at https://cistrome.shinyapps.io/timer/ (Li et al., 2017). This data was available for 353 CASP8-WT and 51 CASP8-MT cases from HNSC. The comparison of immune cell infiltration levels across CASP8-WT, CASP8-MT, and HPV-positive cases was performed using a two-sided Wilcoxon rank test and the graphs were plotted using R. Similar analysis was performed for all CASP8-MT and CASP8-WT cases from UCEC.
Survival analysis
Survival analysis was performed to investigate the difference in the survival of CASP8-WT and CASP8-MT patients from HNSC and UCEC. Survival analysis was also performed to investigate the effect of factors such as expression levels of certain genes and immune cell infiltration on survival. The expression levels of genes of interest were obtained from FPKM-UQ files from TCGA and the data for distribution of immune cell infiltration was obtained from TIMER. Kaplan–Meier curves for CASP8-WT and CASP8-MT cases were plotted using the Survival and Survminer packages in R and the plots were compared using the log-rank test (Therneau, 2015).
To investigate the effect of genes of interest (such as those from gene sets enriched in GSEA or genes involved in necroptosis) and immune cell infiltration levels on survival, multivariate Cox proportional hazards test was performed for HNSC cases. In addition, Cutoff Finder tool available at http://molpath.charite.de/cutoff/ was used to investigate the influence of a single continuous variable on survival (Budczies et al., 2012). In the Cutoff Finder tool, the cutoff for dichotomization of a continuous variable was determined as the point with the most significant split by log-rank test, using coxph and survfit functions from the R package survival. Survival analysis of either CASP8-WT or CASP8-MT cases was performed using this method by dichotomizing gene expression or immune cell infiltration levels.
Results
Genes involved in immune response are up-regulated in mutant-CASP8 HNSCs
To investigate the significance of CASP8 mutations in head and neck squamous cell carcinoma (HNSC), we performed differential gene expression analysis using RNA-seq data from HNSC cases with and without CASP8 mutations. As reported previously, HNSCs carrying CASP8 mutations occurred predominantly in sites within the oral cavity such as the cheek mucosa, floor of mouth, tongue, larynx, and overlapping sites of the lip, oral cavity, and pharynx (Table S1). In addition, since HPV-positive (Human Papillomavirus-positive) HNSCs constitute a molecularly distinct subtype; we examined the HPV status of the 55 mutant-CASP8 HNSCs using data from Chakravarthy et al. (2016). Based on this reported data, all 55 mutant-CASP8 HNSCs were found to be HPV-negative. Since all the HNSC cases carrying CASP8 mutations were HPV-negative, and were from specific sites within the oral cavity, HNSCs carrying wild-type-CASP8 that were HPV-negative and also from these same sites were selected as controls for all subsequent analyses. A total of 424 HNSC cases of which 369 had wild-type-CASP8 (CASP8-WT) and 55 had mutant-CASP8 (CASP8-MT) were thus selected (Fig. 1, see also Table S1). Of these, RNA-seq data was available for 354 CASP8-WT and 53 CASP8-MT cases.
Raw sequencing reads from CASP8-WT and CASP8-MT cases, obtained from HT-Seq files, were subjected to edgeR analysis for differential gene expression (Table S2). At FDR < 0.001, 186 genes were up-regulated in CASP8-MT with log2FC > 1.3 while 1,139 genes were down-regulated in CASP8-MT with log2FC < −1.3 (Fig. 2A). There was also a statistically significant 1.3-fold increase in the expression level of the CASP8 gene perhaps to overcome the loss of function (Table S2, gene ESNG00000064012 in HNSC edgeR output).
To identify biological processes specifically enriched in the CASP8-WT or CASP8-MT cases, enrichment analysis was performed with the differentially expressed genes using tools available at the Gene Ontology (GO) Consortium. As seen in Fig. 2A, distinct processes were enriched in the CASP8-WT and CASP8-MT cases. For example, genes involved in the regulation of immune response (p-value = 3.30E−02) were enriched in CASP8-MT HNSCs while genes with roles in synaptic transmission (p-value = 2.85E−07), synaptic vesicle exocytosis (p-value = 3.90E−03), and muscle contraction (p-value = 2.29E−03) were the top three biological processes enriched in CASP8-WT HNSCs. Please refer to Table S3 for the full list.
We further analyzed the differential gene expression data using the Gene Set Enrichment Analysis (GSEA) tool. After generating a pre-ranked gene list based on logFC values from the edgeR analysis, we queried this list in the GSEA software using hallmark gene sets available at the Molecular Signatures Database. Several gene sets were enriched in upregulated or downregulated genes in CASP8-MT cases at FDR < 25%. Particularly, gene sets involved in immune regulation such as allograft rejection, complement, inflammatory response, interferon-α response, and interferon-γ response, were specifically enriched in the CASP8-MT HNSCs, in sync with the GO results (Figs. 2B–2F and Table S4). The hallmark gene sets enriched in CASP8-WT HNSCs were epithelial-mesenchymal transition (EMT), myogenesis, and the KRAS pathway (Figs. 2G–2I and Table S4).
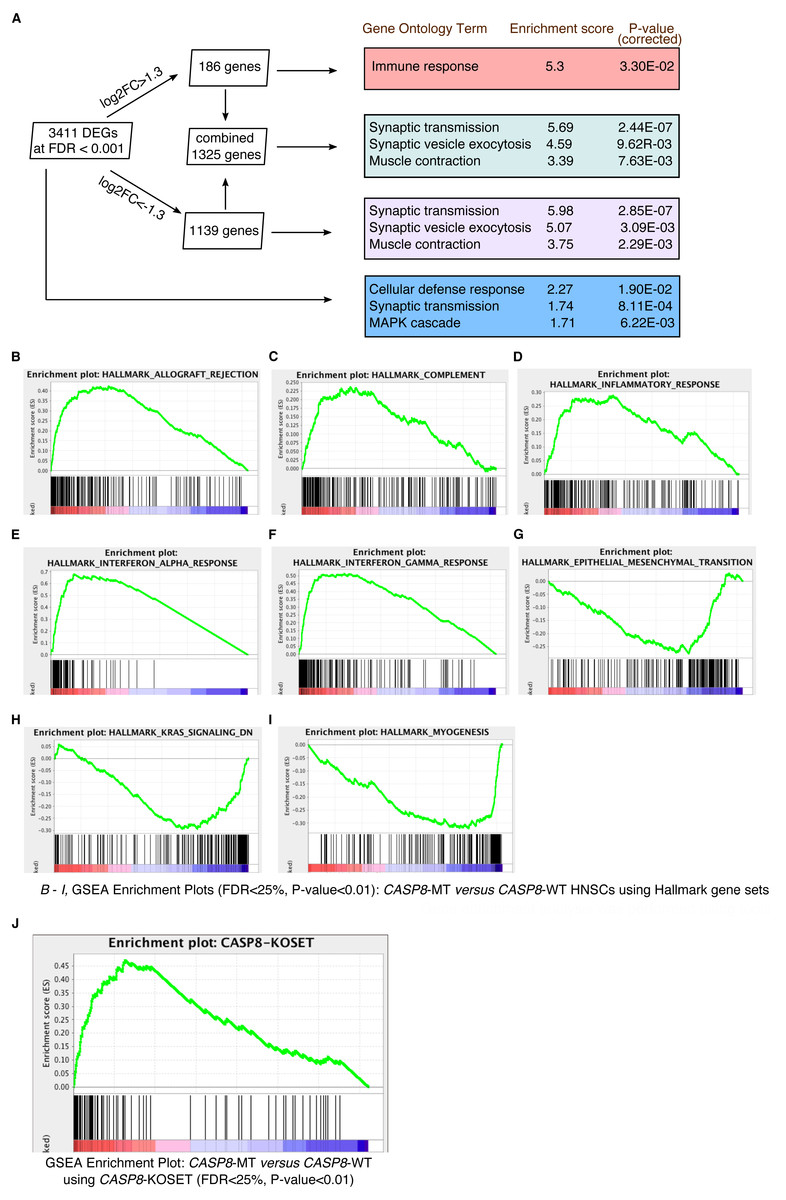
Figure 2: Gene enrichment analyses reveal a prominent immune signature in CASP8-MT HNSCs.
Gene enrichment analysis was performed using tools available at the Gene Ontology Consortium (A), as well as using the Gene Set Enrichment Analysis tool (B–J). (A) Enrichment analysis was performed using genes with FDR < 0.001 and/or showing log2FC greater than 1.3 or less than −1.3. The top three gene ontology terms, based on enrichment scores, among the PANTHER GO-Slim Biological Processes significantly enriched in these gene lists are indicated along with Bonferroni-corrected P-values. (B–I) GSEA was performed using a pre-ranked list generated using log2FC values from the edgeR analysis. GSEA Hallmark gene sets enriched in CASP8-MT HNSCs (B–F) or CASP8-WT HNSCs (G–I) with FDR < 25%, P-value < 0.01, and showing enrichment at the top or bottom of the list are shown. (J) Enrichment plot of a GSEA performed with the same pre-ranked list that was analysed in (B–I) and a gene set of human orthologs of the genes up regulated in the skin epidermis of Casp-8F/−K5-Cre mice (CASP8-KOSET) is shown (FDR < 25%, P-value < 0.01).{kind=link}
Gene expression in the skins of epidermal Caspase-8 knockout mice mirrors the expression pattern of mutant-CASP8 HNSCs
Expression of an enzymatically inactive Caspase-8 mutant or the deletion of wild-type Caspase-8 in the mouse epidermis leads to chronic skin inflammation (Kovalenko et al., 2009; Lee et al., 2009). A microarray analysis performed by Kovalenko et al. to identify genes specifically up-regulated in the skin epidermis of Casp-8F/−K5-Cre (relative to Casp-8F/+K5-Cre epidermis) mice revealed increased expression of several immune-regulatory and inflammatory genes including several cytokines. Using the human orthologs of these up-regulated genes (Table S5), we again queried the pre-ranked gene list with the GSEA tool. As seen in Fig. 2J, genes highly expressed in the Casp-8F/−K5-Cre mouse skins were also significantly enriched in CASP8-MT HNSCs (as opposed to their wild-type counterparts), indicating that the inactivation of CASP8 leads to the up-regulation of a similar set of genes in both mouse and human epidermal tissues.
Enrichment of immune response gene sets correlates with increased infiltration of specific immune cell types in mutant-CASP8 HNSCs
Gene sets involved in immune response were specifically enriched in CASP8-MT HNSCs. HPV-positive HNSCs, a subset of HNSCs, also display high immune cell infiltration, as compared to HPV-negative HNSCs (Chakravarthy et al., 2016; Mandal et al., 2016; Nguyen et al., 2016; Russell et al., 2013). We investigated if the enrichment of immune response genes in CASP8-MT HNSCs was correlated with increased infiltration of immune cells, and if it was comparable to the immune cell infiltration levels in HPV-positive HNSCs. Immune cell infiltration levels in three subsets of HNSCs; CASP8-WT, CASP8-MT (both HPV-negative), and HPV-positive (which is CASP8-WT), were compared using the Wilcoxon test; the comparisons were: (1) CASP8-WT and CASP8-MT (2) CASP8-WT and HPV-positive CASP8-WT, and (3) CASP8-MT and HPV-positive CASP8-WT. We checked if there was a difference in the numbers/types of immune cell infiltrates between these three subsets of HNSCs using the data available at Tumor IMmune Estimation Resource (TIMER) (Li et al., 2017), a comprehensive resource for immune cell infiltration of TCGA tumors.
Consistent with the GSEA results, CASP8-MT cases showed significantly higher infiltration of CD8+ T cells, neutrophils, and dendritic cells as compared to CASP8-WT cases (p-values < 0.0005), suggesting that the immune response to the tumor in WT and MT cases was different (Figs. 3A–3C). Also, in agreement with previous reports, HPV-positive HNSCs had significantly higher infiltration of all immune cell types as compared to the CASP8-WT HNSCs. A comparison of immune cell infiltration levels in HPV-positive and CASP8-MT HNSCs showed that the extent of infiltration of CD8+ T cells, neutrophils, and dendritic cells (Figs. 3A–3C) in these two subsets was also similar. However, HPV-positive HNSCs had higher infiltration of CD4+ T cells and B cells, compared to the other two subsets (Figs. 3D, 3E).
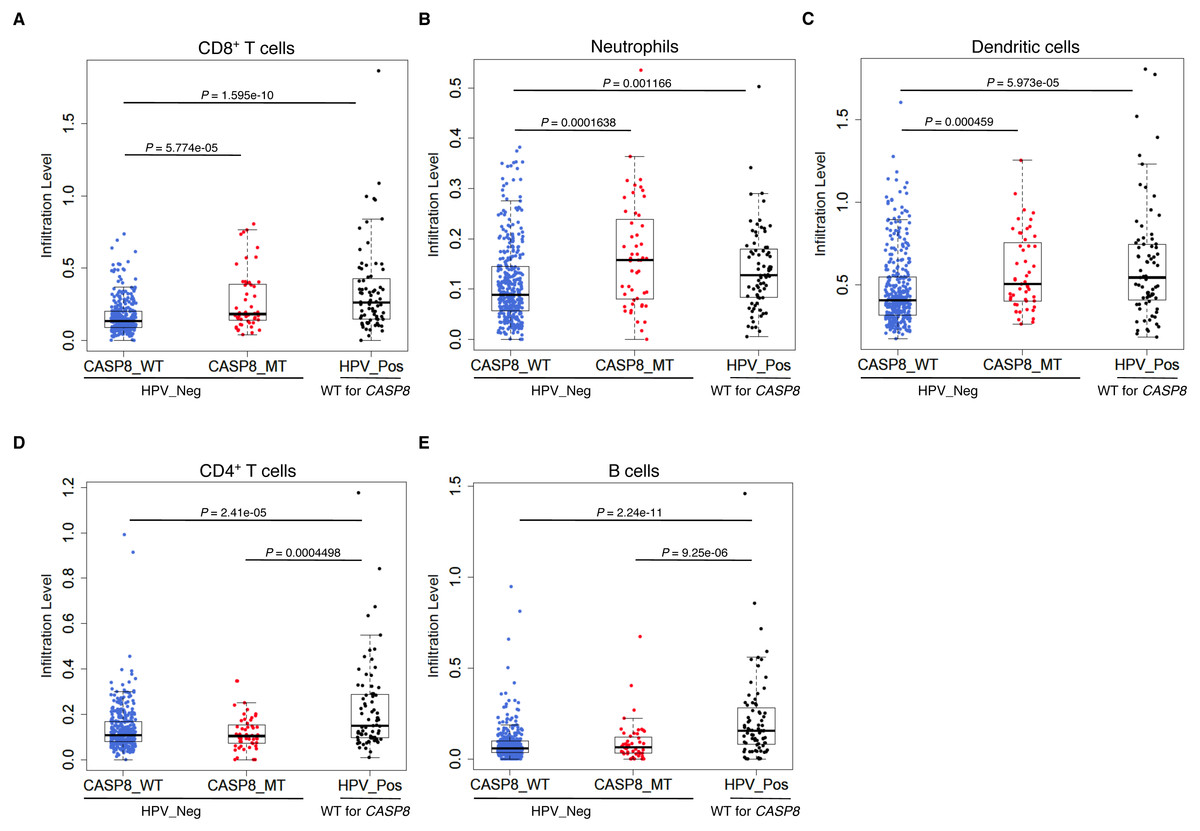
Figure 3: CASP8-MT HNSCs have higher numbers of certain types of infiltrating immune cells compared to CASP8-WT HNSCs.
Immune cell infiltration levels in CASP8-WT (blue-filled circles), CASP8-MT (red-filled circles) (both HPV-negative), and HPV-positive (black-filled circles) HNSCs were compared using the immune cell infiltration data available at TIMER. Boxplots showing the levels of CD8+ T cells, neutrophils, and dendritic cells (A–C), as well as CD4+ T cells and B cells (D, E) in the three HNSC subsets are displayed. Significance testing was performed using the unpaired two-sided Wilcoxon test. All comparisons with P-value < 0.005 were considered significant and are indicated in the plots.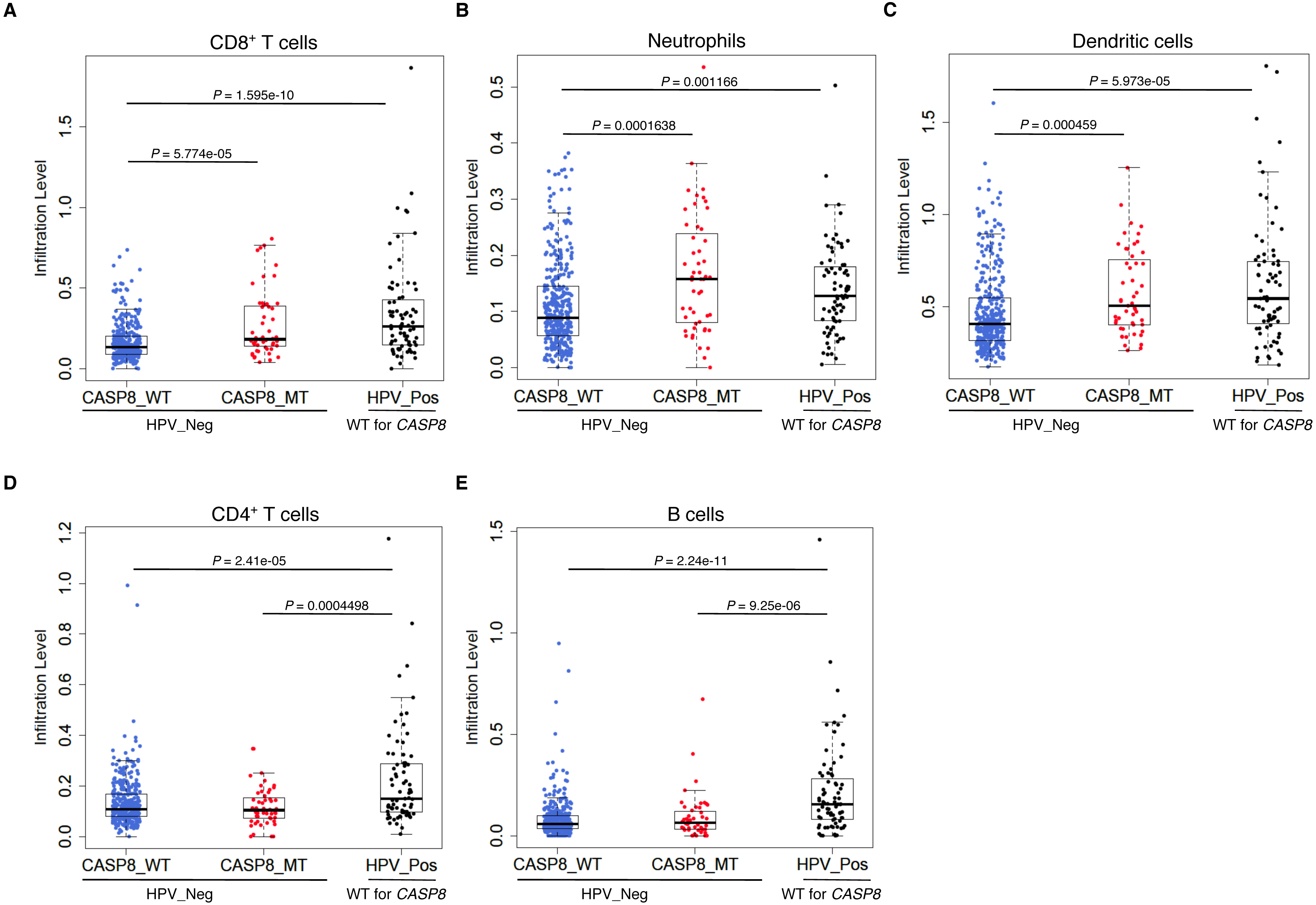{kind=link}
The “immune signature” of mutant-CASP8 HNSCs does not correlate to improved overall survival
High levels of immune cell infiltration in HPV-positive cases correlates with better survival in HPV-positive HNSC cases (Nguyen et al., 2016; Russell et al., 2013). To investigate if a similar effect could be observed in the survival of HNSC patients with and without CASP8 mutation, Kaplan–Meier analysis was performed on the CASP8-WT and CASP8-MT cases (filtered as per the schema in Fig. 1). There was no significant difference in the survival of patients with and without CASP8 mutations (p-value = 0.16, Fig. 4A), indicating that high levels of immune cell infiltration may not necessarily corelate with better survival in HNSCs.
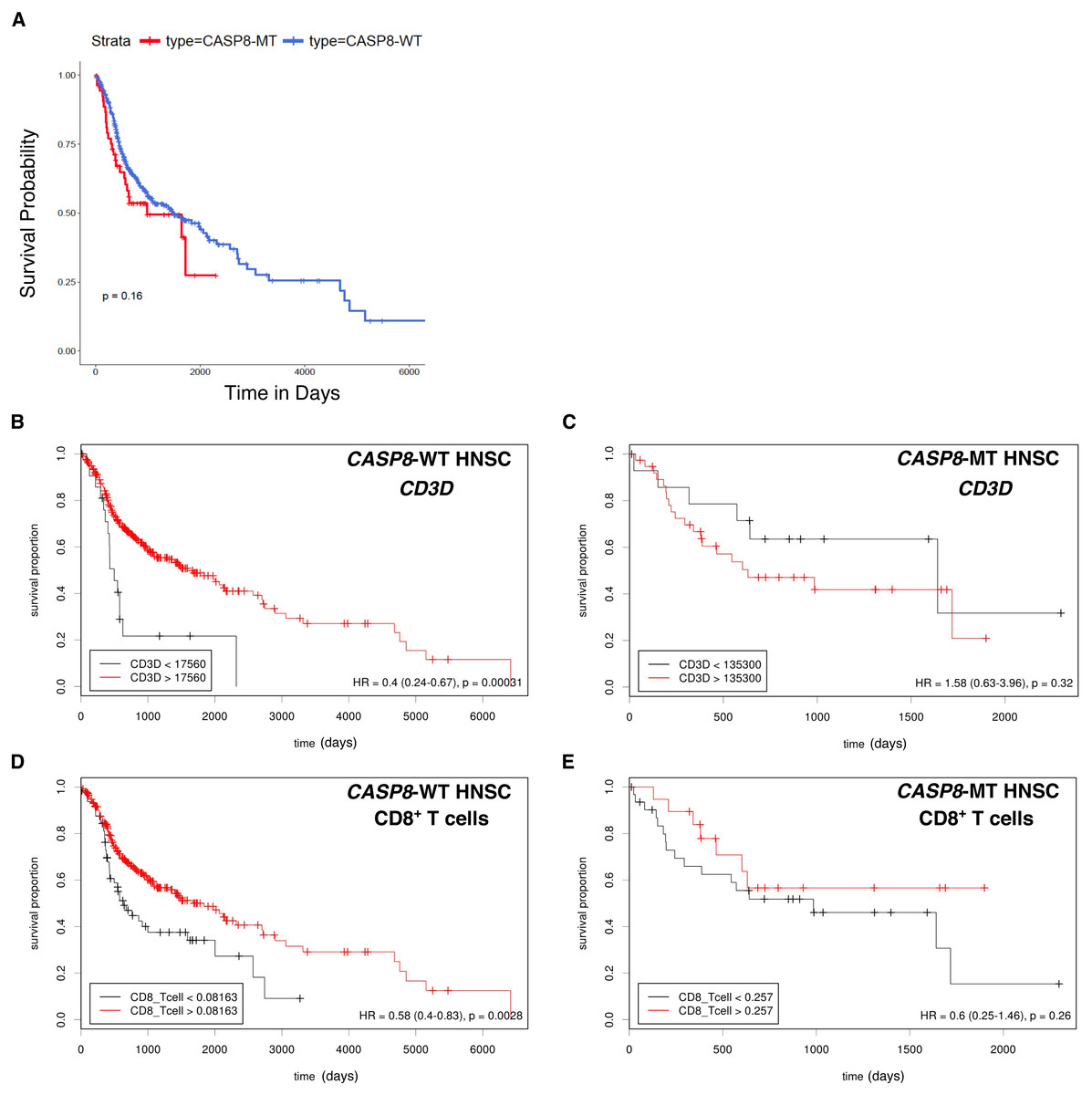
Figure 4: Survival analysis indicates lack of a survival advantage in CASP8-MT HNSCs in spite of their immune signature.
(A) Kaplan-Meier plots showing the survival probability of patients with CASP8-WT or CASP8- MT HNSC tumors (filtered as per the schema in Fig. 1). Log-rank test was used to compare the two curves and the log-rank P-value is indicated. (B–E) Survival plots generated using the Cutoff Finder tool showing the influence of the expression levels of CD3D and the levels of CD8+ T cells on overall survival in CASP8-WT (B, D) and CASP8-MT (C, E) cases. Gene expression data was obtained from FPKM-UQ files at TCGA and immune cell infiltration data was obtained from TIMER.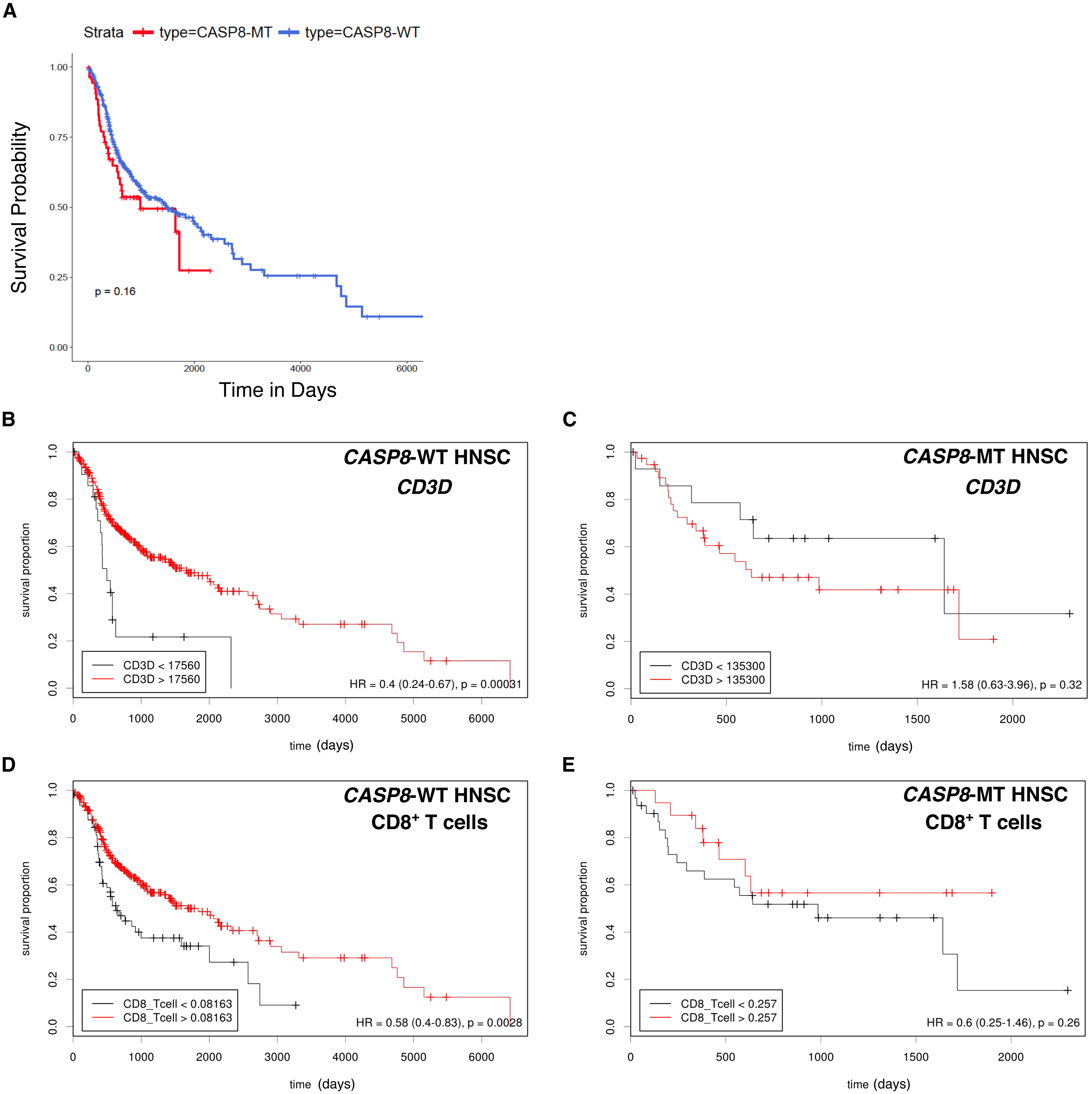{kind=link}
The effect of genes from pathways enriched either in CASP8-WT or CASP8-MT tumors (listed in Table S4) on survival was then investigated using the Cox proportional hazards model. Four genes from pathways enriched in CASP8-MT HNSCs; PRF1, CXCR6, CD3D, and GZMB, reduced the hazard ratio significantly in CASP8-WT cases, at p < 0.05 (Table S6). We also performed the survival analysis using Cutoff Finder to investigate the effect of the expression of individual genes on the survival of CASP8-WT and CASP8-MT cases. Increased expression of all these four genes was associated with higher overall survival in CASP8-WT (at p < 0.05). In contrast, in CASP8-MT cases, such association was seen only with GZMB expression levels (Figs. 4B, 4C and Fig. S1). Similarly, higher CD8+ T cell estimates (from TIMER) was also significantly associated with better survival in CASP8-WT but not in CASP8-MT HNSCs (Figs. 4D, 4E).
Since CASP8 is a negative regulator of the necroptotic pathway (Günther et al., 2011; Weinlich et al., 2013), we also investigated the effect of expression levels of genes involved in necroptosis on survival. Higher expression of RIPK1, RIPK3, and MLKL was associated with higher overall survival in CASP8-WT but not in CASP8-MT cases (Fig. S2). Additional factors that influenced survival are shown in Table S6.

Figure 5: CASP8-MT UCECs display an immune gene signature, have higher numbers of certain types of infiltrating immune cells, and survive better than CASP8-WT UCECs.
(A–D) GSEA was performed using a pre-ranked list generated using log2FC values from the edgeR analysis. Some GSEA Hallmark gene sets enriched in CASP8-MT UCECs (A–D) are shown. (E, F) Immune cell infiltration levels in CASP8-WT (blue-filled circles) and CASP8-MT (red-filled circles) UCECs were compared using the immune cell infiltration data available at TIMER. Boxplots showing the levels of CD8+ T cells and dendritic cells in the two UCEC groups are displayed. Significance testing was performed using the unpaired two-sided Wilcoxon test. All comparisons with P-value < 0.005 were considered significant and are indicated in the plots. (G) Kaplan-Meier plots showing the survival probability of patients with CASP8-WT or CASP8- MT UCEC tumors. Log-rank test was used to compare the two curves and the log-rank P-value is indicated.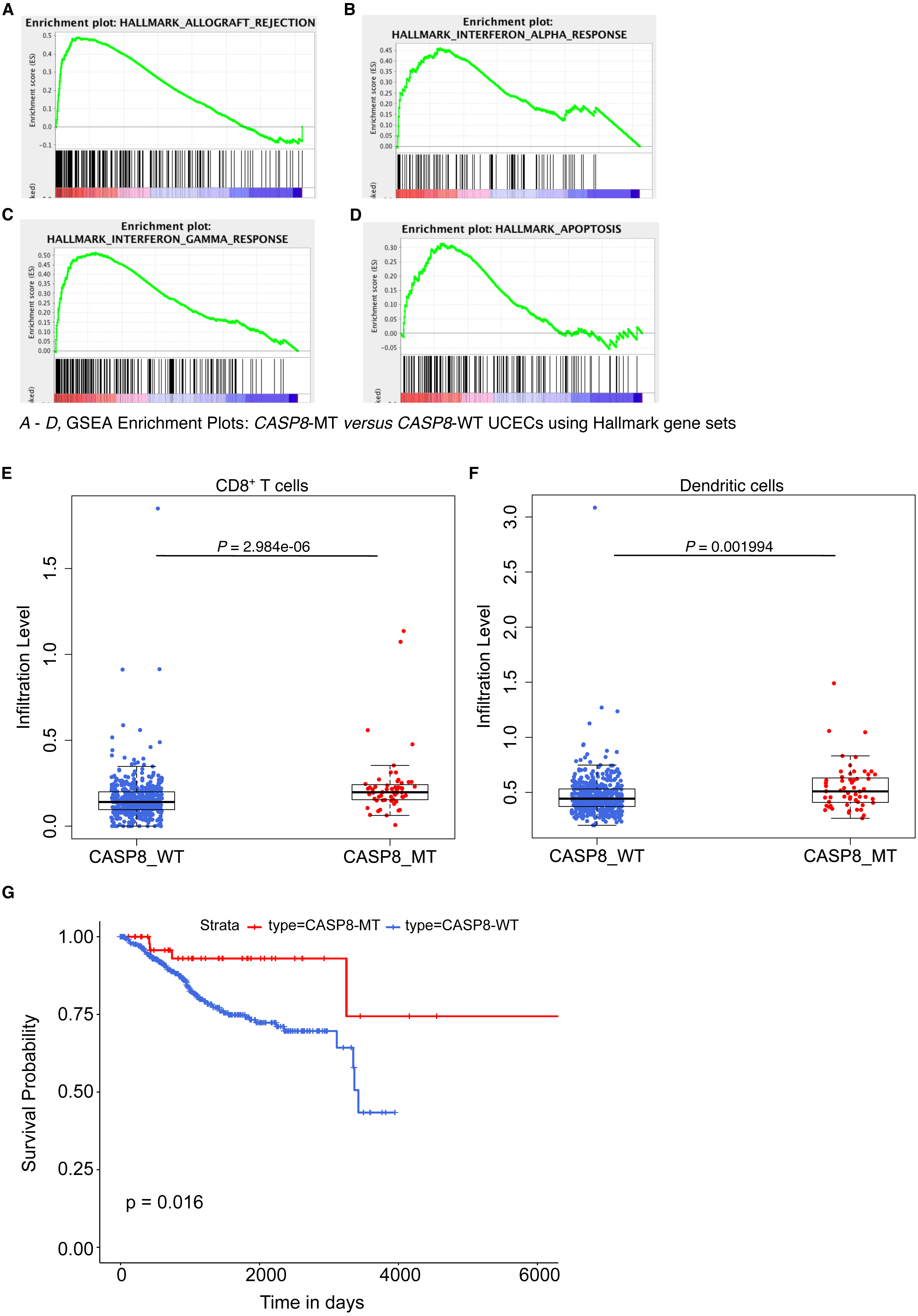{kind=link}
Mutant-CASP8 UCECs exhibit an immune signature similar to mutant-CASP8 HNSCs
We then investigated if this effect seen in CASP8-MT HNSCs was broadly applicable across other cancers carrying CASP8 mutations. On searching the Genomic Data Commons, we found that CASP8 was recurrently mutated in about 10% of (UCEC) cases. From a total of 560 UCEC cases, RNA-seq data was available for 476 CASP8-WT and 56 CASP8-MT cases. HTSeq files containing raw sequencing reads from these two groups were subjected to edgeR analysis for differential gene expression and further analyzed using the GSEA tool. Several gene sets involved in immune regulation were specifically enriched in the CASP8-MT UCECs. Notably, categories such as allograft rejection, interferon-α response, and interferon-γ response, were enriched in the CASP8-MT UCECs similar to the HNSC results (Figs. 5A–5C and 2B–2F). The genes that contributed to core enrichment in these gene sets in CASP8-MT UCECs also contributed to core enrichment of the same gene sets in CASP8-MT HNSCs, indicating that similar immune response genes were upregulated in the two carcinomas (Table S7). However, unlike HNSCs, the gene set for inflammatory response did not show any enrichment in CASP8-MT UCECs. CASP8-MT UCECs were additionally enriched for genes involved in apoptosis. Notably, this was not observed in the CASP8-MT HNSCs (Figs. 2B–2I and 5D).
High levels of IL33 and neutrophil infiltration are observed in mutant-CASP8 HNSCs but not in mutant-CASP8 UCECs
Using TIMER, we then checked the levels of infiltrating immune cells in the CASP8-WT and CASP8-MT UCECs. Consistent with the GSEA results, CASP8-MT UCEC cases showed significantly higher infiltration of CD8+ T cells and dendritic cells as compared to CASP8-WT cases (p-values < 0.005). However, in contrast to the HNSC data, the levels of neutrophils were not significantly higher in the CASP8-MT UCEC group (Figs. 5E, 5F, see also Fig. 3B). We then investigated if differences in the levels of neutrophil-active chemokines could potentially explain this observation (Sadik, Kim & Luster, 2011). From the edgeR differential expression data comparing the CASP8-MT and CASP8-WT groups in HNSC and UCEC, we obtained the fold change values and statistical significance of different chemokines known to attract neutrophils (Table S8). Interestingly, the cytokine IL33 was significantly up regulated (1.8 fold, FDR < 0.001) in CASP8-MT HNSCs but not in CASP8-MT UCECs.
Next, we performed Kaplan–Meier analysis on CASP8-WT and CASP8-MT UCEC cases. In contrast to the HNSC survival data, there was a difference in the survival of UCEC cases with and without CASP8 mutations, with cases harboring CASP8 mutations reporting better overall survival (p-value = 0.019, Fig. 5G).
Discussion
Here, we report a distinct class of carcinomas that have mutated CASP8. Using bioinformatics approaches to mine the TCGA data, we identified high expression of immune response-related genes (listed in Table S7) combined with high infiltration of CD8+ T cells and dendritic cells as a prominent shared immune signature in CASP8-MT carcinomas. In the first part of our analyses, we investigated the implications of the enrichment of this immune signature across different HNSC subtypes. Subsequently, in the second part, we investigated the correlation between immune signature and survival in two carcinomas having a significant number of cases with CASP8 mutations, HNSC and UCEC. Our analyses showed that despite similarities in the enrichment of gene sets, these carcinomas exhibited varying correlations of immune signature with survival. Our studies indicated that tissue-specific differences, such as the levels of infiltrating neutrophils and the cytokine IL33, could be responsible for the varying correlation of immune signature with survival.
Multiple studies have reported that HPV-positive HNSCs display a strong immune signature and high infiltration of immune cells that correlates with better survival (Nguyen et al., 2016; Russell et al., 2013). In contrast, our studies show that the enrichment of immune response genes and infiltration of immune cells seen in CASP8-MT HNSCs does not appear to correlate with improved prognosis. In fact, CASP8 mutation leads to the loss of a survival advantage that is observed in HNSC patients with wild-type CASP8 tumors under certain conditions. For example, higher expression levels of genes such as PRF1, CD3D, and CXCR6 are associated with better survival in CASP8-WT but not in CASP8-MT. It is possible that the higher expression of these genes results in higher extent of apoptosis leading to survival advantage. This perhaps does not take place in CASP8-MT, leading to the loss of survival advantage from higher expression of these genes. These results argue that a tumor microenvironment with high infiltration of immune cells does not necessarily provide a survival benefit in HNSCs. However, it is important to note that while the sample numbers of CASP8-MT cases are significant (n = 55), it is lesser than the number of CASP8-WT cases (n = 369). This may influence p-values, and it will be necessary to confirm these findings with greater numbers of CASP8-MT cases once more data becomes available.
We can think of at least two potential scenarios to explain the increased immune cell infiltration observed in CASP8-MT tumors. (a) Unregulated inflammatory and wound healing response: As mentioned earlier, loss of Caspase-8 in the mouse epidermis leads to chronic inflammation (Kovalenko et al., 2009). The infiltration of immune cells in mucosa lacking CASP8 accompanied by the enrichment of immune-associated gene sets is highly reminiscent of this phenotype. It has also been proposed that the loss of Caspase-8 in the mouse skin epidermis simulates a wound healing response (Lee et al., 2009). Both scenarios involve a gamut of immune cell types and secreted cytokine factors, leading to immune cell infiltration. It should however be noted that although similar gene sets are enriched in mouse skins lacking Caspase-8 and in CASP8-MT tumors, the types of immune cell infiltrates in the two are different. (b) Necroptosis: More recently, several studies have revealed a role for Caspase-8 as an inhibitor of necroptosis, a highly pro-inflammatory mode of cell death (Pasparakis & Vandenabeele, 2015; Feltham, Vince & Lawlor, 2017). In intestinal epithelia, the loss of Caspase-8 promoted necroptosis through the activation of RIP kinases and MLKL (Günther et al., 2011; Weinlich et al., 2013). A similar scenario could be occurring in CASP8-MT tumors leading to the expression of pro-inflammatory genes and the infiltration of immune cells.
Why doesn’t the increased number of immune cells translate into improved prognosis in CASP8-MT HNSC tumors? Since CASP8 is an important mediator of the extrinsic apoptotic pathway, CASP8-MT tumors may have greater resistance to Fas- or DR5- mediated cell death pathways, which are typically employed by CD8+ T cells and Natural Killer cells to target infected/tumor cells (Li et al., 2014; Rooney et al., 2015). The survival analysis carried out in this study showed that CASP8-WT HNSC patients with higher expression of genes involved in T-cell mediated cytotoxicity had better survival. Importantly, this advantage was not seen in CASP8-MT patients.
Several studies have reported that high neutrophil numbers and an elevated neutrophil/lymphocyte ratio portended poorer prognosis in OSCC (Mahalakshmi et al., 2018; Glogauer et al., 2015). Thus, it is possible that elevated levels of neutrophil infiltration seen in CASP8-MT HNSC cases could be one of several events contributing to the poorer prognosis of CASP8-MT HNSCs. IL33, a cytokine and an alarmin linked to necroptosis may represent a possible mechanism for neutrophil recruitment in these cases (Alves-Filho et al., 2010; Hueber et al., 2011). High IL33 levels are also associated with poor prognosis in HNSCs (Chen et al., 2013). In addition, the pro-inflammatory environment generated during necroptosis may hold other advantages for the survival of CASP8-MT HNSCs. Necroptosis, IL33 levels, and neutrophil infiltration together or through independent mechanisms could be leading to a pro-tumor environment. Thus, promoting necroptosis may not necessarily translate into better survival for HNSC patients with apoptosis-resistant tumors.
Another reason for the lack of survival advantage in CASP8-MT HNSCs could be the composition of tumor-infiltrating immune cells in these tumors. For instance, HPV-positive tumors had higher levels of B cells and CD4+ T cells as compared to CASP8-MT tumors. It is likely that in addition to cytotoxic T cells, B cells and CD4+ T cells are required to mediate an immune response essential for tumor cell death, possibly for tumor antigen presentation or cytokine secretion.
A comparison of CASP8-MT HNSCs and CASP8-MT UCECs highlighted similarities and differences between the two carcinomas. Both CASP8-MT HNSCs and CASP8-MT UCECs showed an enrichment of gene sets involved in immune response such as interferon α response, interferon γ response, and allograft rejection. Most of the genes contributing to core enrichment in these gene sets in UCECs also contributed to core enrichment of these gene sets in HNSCs. Moreover, both these carcinomas showed high infiltration of CD8+ T cells and dendritic cells but not B cells or CD4+ T cells. CASP8 mutation thus led to a similar immune response in both HNSCs and UCECs. This shared immune signature, however, did not correlate with a specific survival outcome. Notably, CASP8-MT UCECs showed a significant survival advantage over CASP8-WT UCECs, unlike its HNSC counterpart. While we do not yet know the causal reason(s), the differences per se may be worth noting and could be responsible for this advantage. For instance, in contrast to CASP8-MT HNSCs, the gene set for inflammatory response was not enriched but the gene set for apoptosis was enriched in CASP8-MT UCECs. There was also no increased infiltration of neutrophils or transcriptional upregulation of IL33 in CASP8-MT UCECs. The up-regulation of apoptotic pathways together with the lack of enrichment of an inflammation-associated gene set that is typical of necroptosis perhaps indicates that apoptosis, rather than necroptosis, is the predominant mode of programmed cell death in CASP8-MT UCECs. This lack of inflammation may also be responsible for the lack of neutrophil infiltration in CASP8-MT UCECs since neutrophil chemoattractants, such as IL33, may not be released during apoptosis but is perhaps released during the highly inflammatory process of necroptosis, in turn leading to neutrophil infiltration.
It is also possible that necroptosis is initiated in CASP8-MT UCECs but the accompanying IL33 up-regulation and/or neutrophil infiltration seen in HNSCs does not take place due to tissue-specific differences. Under such conditions, apoptosis and necroptosis together could provide the survival advantage that is observed in CASP8-MT UCECs. Thus, in contrast to HNSCs, Caspase-8 pathway can be explored to identify potential drug targets in UCECs.
Conclusions
In this in silico study, we explore the implications of CASP8 mutations that have been identified across carcinomas through large-scale genomic studies. Our studies show that CASP8-mutated carcinomas display a shared immune signature. However, the consequences of this immune signature vary with CASP8- MT UCECs showing better survival while CASP8-MT HNSC cases do not have any survival advantage. Our analyses further suggest that neutrophil numbers and IL33 levels could be potential factors affecting the survival of mutant-CASP8 carcinomas. Broadly, our study highlights the need to further investigate the interaction between pathways of programmed cell death, immune response, and survival in carcinomas. Such studies could open a new window for therapeutic intervention in CASP8-mutated carcinomas.
Supplemental Information
Survival plots generated using the Cutoff Finder tool showing the influence of expression levels of GZMB, PRF1, and CXCR6 on overall survival in CASP8-WT (left) and CASP8-MT (right) cases
Gene expression data was obtained from FPKM-UQ files at TCGA and immune cell infiltration data was obtained from TIMER.
Survival plots generated using the Cutoff Finder tool showing the influence of expression levels of genes involved in necroptosis; RIPK1, RIPK3, and MLKL on overall survival in CASP8-WT (left) and CASP8-MT (right) cases
Gene expression data was obtained from FPKM-UQ files at TCGA and immune cell infiltration data was obtained from TIMER.
List of HNSC and UCEC tumors used in this study
The table lists HNSC and UCEC cases from TCGA analyzed in the study, along with the status of CASP8 mutation, availability of gene expression data, ICD-10-CM code and site of tumor.
Results of edgeR analysis
The table shows results of edgeR analysis performed to investigate differential gene expression using HTSeq files from CASP8-WT and CASP8-MT HNSC & UCEC cases. Differential expression and statistical significance of 60,483 transcripts are shown. Genes passing the cutoffs used (FDR < 0.001, and log2FC > 1.3 or < − 1.3) are also shown in separate sheets.
Results of Gene Ontology analysis
The tables list the enrichment of specific biological processes among the genes significantly differentially expressed between CASP8-WT and CASP8-MT HNSCs.
Top differentially expressed genes from GSEA
The tables list the gene sets and the top 10 or 12 genes in the GSEA Hallmark categories that were significantly enriched in CASP8-MT or CASP8-WT HNSCs.
List of mouse genes and their human orthologs used for GSEA
The table shows the list of mouse genes up regulated in Caspase-8 epidermal knockout mouse skin, and their human orthologs, which was used for GSEA (CASP8-KOSET). Homology information was obtained from http://www.informatics.jax.org/.
Results of Cox proportional-hazards regression analysis
The table shows the results of survival analysis performed using the Cox test to investigate the influence of expression levels of top genes from gene sets that were enriched in either CASP8-WT or CASP8-MT as well as to investigate the effect of immune cell infiltration. The covariates with p-value < 0.05 and with significant Cox model (p < 0.05 for likelihood test, Wald test, and logrank test) are highlighted in red. The covariates with p-values < 0.05 but where Cox model was not significant are highlighted in blue. The cells with results of genes that regulate necroptosis are colored blue while the cells with results of genes related to immune cell infiltration are colored green. *ATP4A is not considered to have a significant effect since 95% confidence interval for hazard ratio includes 1.
Shared Immune Signature Genes
The table lists genes that contribute to core enrichment of gene sets Allograft rejection, Interferon alpha response, and Interferon gamma response in both HNSC and UCSC in the GSEA.
edgeR analysis results for neutrophil-active chemokines
The table shows results of edgeR analysis reported in Table S2 for nine neutrophil-active chemokines. Fold change and statistical significance of the CASP8-MT versus CASP8-WT comparison in both HNSC and UCEC are shown.