
Differing responses of red abalone (Haliotis rufescens) and white abalone (H. sorenseni) to infection with phage-associated Candidatus Xenohaliotis californiensis
- Published
- Accepted
- Received
- Academic Editor
- Lesley Hoyles
- Subject Areas
- Aquaculture, Fisheries and Fish Science, Conservation Biology, Microbiology, Virology, Natural Resource Management
- Keywords
- Bacteriophage, Abalone, Candidatus xenohaliotis californiensis, Withering syndrome, Phage therapy, Ocean warming, White abalone, Haliotis rufescens, Haliotis sorenseni, Rickettsia
- Copyright
- © 2018 Vater et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Differing responses of red abalone (Haliotis rufescens) and white abalone (H. sorenseni) to infection with phage-associated Candidatus Xenohaliotis californiensis. PeerJ 6:e5104 https://doi.org/10.7717/peerj.5104
Abstract
The Rickettsiales-like prokaryote and causative agent of Withering Syndrome (WS)—Candidatus Xenohaliotis californiensis (Ca. Xc)—decimated black abalone populations along the Pacific coast of North America. White abalone—Haliotis sorenseni—are also susceptible to WS and have become nearly extinct in the wild due to overfishing in the 1970s. Candidatus Xenohaliotis californiensis proliferates within epithelial cells of the abalone gastrointestinal tract and causes clinical signs of starvation. In 2012, evidence of a putative bacteriophage associated with Ca. Xc in red abalone—Haliotis rufescens—was described. Recently, histologic examination of animals with Ca. Xc infection in California abalone populations universally appear to have the phage-containing inclusions. In this study, we investigated the current virulence of Ca. Xc in red abalone and white abalone at different environmental temperatures. Using a comparative experimental design, we observed differences over time between the two abalone species in mortality, body condition, and bacterial load by quantitative real time PCR (qPCR). By day 251, all white abalone exposed to the current variant of Ca. Xc held in the warm water (18.5 °C) treatment died, while red abalone exposed to the same conditions had a mortality rate of only 10%, despite a relatively heavy bacterial burden as determined by qPCR of posterior esophagus tissue and histological assessment at the termination of the experiment. These data support the current status of Ca. Xc as less virulent in red abalone, and may provide correlative evidence of a protective phage interaction. However, white abalone appear to remain highly susceptible to this disease. These findings have important implications for implementation of a white abalone recovery program, particularly with respect to the thermal regimes of locations where captively-reared individuals will be outplanted.
Introduction
Abalone are iconic benthic invertebrates that contribute to ecological health of northern Pacific coast kelp forests and serve as a food source for endangered sea otters Enhydra lutris. California’s wild abalone fishery flourished from the 1950s–1980s but was decommercialized in response to population declines from overexploitation followed by disease (California Department of Fish & Wildlife, 2005). Farmed abalone is of increasing economic significance; in 2008, it was estimated that over 129,000 metric tons of farmed abalone was supplied to the world market (Cook, 2016).
Withering syndrome (WS) was first reported in mid-1980’s at the Channel Islands, California and decimated black abalone—Haliotis cracherodii—populations (Haaker et al., 1992). The infection causes reduced feeding behavior and nutrient absorption; animals wither as they lose body mass through catabolism of the foot muscle (Gardner et al., 1995). The causative agent of the disease, Candidatus Xenohaliotis californiensis (Ca. Xc) is a member of the Order Rickettsiales of the Alphaproteobacteria (Friedman et al., 2000). Analysis of five genes (16S rRNA, 23S rRNA, ftsZ, vVirB11, and vVirD4) suggested that Ca. Xc is most closely related to the Neorickettsia genus and is the most ancestral form of the Anaplasmataceae family studied to data (Cicala et al., 2017b). Transmission of Ca. Xc appears to be fecal-oral (Moore et al., 2001). Ca. Xc infects the luminal epithelium of the posterior portion of the esophagus (PE) and digestive gland (Moore et al., 2001). The Ca. Xc bacterium forms large oblong inclusions in the digestive tract epithelium, which are easily identifiable in hematoxylin- and eosin-stained tissue sections (Friedman et al., 2000).
In red abalone, H. rufescens, exposure to warm water events in the presence of Ca. Xc exacerbates morbidity and mortality in two synergistic ways: it reduces the nutritional content of their primary food source, and is associated with elevated pathogen burdens (Vilchis et al., 2005). Trends of increasing frequency and intensity of Pacific ocean warming El-Nino events correspond to dramatic reductions in giant kelp densities (Tegner et al., 1996), and nitrogen nutrient concentration in seawater is inversely related to temperature (Tegner et al., 2001). Furthermore, ocean warming trends coincide with Ca. Xc disease outbreaks (Harvell et al., 1999). The results of laboratory studies using juvenile farm-raised red abalone showed that Ca. Xc has relatively little effect on the health of abalone held at temperatures of approximately 14 °C, while animals held in water approximately 18 °C suffer high mortality rates in association with higher Ca. Xc body burdens (Braid et al., 2005; Moore, Robbins & Friedman, 2000; Rosenblum et al., 2005; Vilchis et al., 2005). Braid et al. (2005) demonstrated that clinical signs of withering syndrome are not solely due to warm water stress..
White abalone–H. sorenseni–health and fitness is best supported by a consistent 14 °C seawater environment as determined by optimization of captive breeding methods (Leighton, 1972; Rogers-Bennett et al., 2016). Increased seawater temperatures are associated with adverse health effects in white abalone that translate to population instability. For example, white abalone spawning success rates are significantly decreased in warm water (20 °C). Ocean warming events have the potential to powerfully exacerbate disease, particularly in the sensitive white abalone species, while simultaneously reducing general animal fitness. White abalone were the first marine invertebrate species to be recognized as federally endangered. Outplanting animals bred and raised in a captive rearing program is considered the key restoration approach to increase densities quickly enough to reduce the probability of extinction (Rogers-Bennett et al., 2016; Stierhoff et al., 2014). White abalone are highly susceptible to WS (Friedman et al., 2007). This and globally increasing sea water temperatures may challenge future outplanting efforts.
After the catastrophic population declines reported in the early 90’s, wild black abalone survival rates began improving in 1996 which could not be fully attributed to ocean cooling trends that repress expression of WS (Chambers et al., 2005; Chambers et al., 2006). Further, anecdotal observations from California red abalone farmers beginning in 2006 indicated that the incidence and severity of WS had diminished; this change correlated with the appearance of a novel bacteriophage hyperparasite associated with Ca. Xc (Crosson et al., 2014; Friedman & Crosson, 2012). This bacteriophage was first described in farmed red abalone from Cayucos, California examined in 2009 (Friedman & Crosson, 2012). Its presence was visualized by histology as morphologically distinct inclusions and transmission electron microscopy (TEM) confirmed the presence of phage particles in several studies. These distinct pleomorphic inclusions, independent from confirmatory TEM, are currently recognized as representing phage-containing Ca. Xc (Brokordt et al., 2017; Crosson et al., 2014; González et al., 2014). Further characterization suggests this phage is a member of the Siphoviridae family and employs a lytic life cycle, which results in lysis of the host cell and replicates its genetic material separately from that of the host (Cruz-Flores & Cáceres-Martínez, 2016).
Bacteriophages are more abundant than any other marine biological entity; however, there are only a relatively small number of published genomes. These likely fail to capture the high diversity of marine phages (Perez Sepulveda et al., 2016). In April 2018, the annotated genome of the Ca. Xc phage was published and represents the first phage of a marine rickettsial-like organism to be sequenced; the identified open reading frames had low levels of similarities to other known biological entities (Cruz-Flores et al., 2018).
While Friedman et al. (2014a) and Friedman et al. (2014b) demonstrated significant improvement in black abalone survival when challenged with phage-containing Ca. Xc, the mechanism by which the phage reduces pathogenicity is unclear. Complex host-pathogen-phage interactions and resulting selective pressures, which increase bacterial resistance to the phage and simultaneously diminished the bacteria’s virulence, have been observed in other microbial systems (León & Bastías, 2015). Evidence supports prophage-associated changes in virulence in other Rickettsiales (Masui et al., 2000). Mechanisms for variation in Ca. Xc virulence unrelated to phage have been explored in recent years. A preliminary study investigating effects of breeding found that there is a genetic component to Ca. Xc infection susceptibility (Brokordt et al., 2017). Interbreeding between abalone species has been show to transfer susceptibility to WS (González et al., 2014). Heritable and environmental variables may also influence the abalone host microbiome, which is generating interest as a possible factor associated with abalone health and resistance to withering syndrome (Cicala et al., 2017a; Connelly, Horner-Devine & Friedman, 2012).
In this study, we aimed to elucidate changes in Ca. Xc virulence in association with phage presence on WS expression in red and white abalone species under thermal conditions that are known to either enhance or retard expression of the disease.
Materials and Methods
Abalone and life support
This experiment was conducted in the California Department of Fish and Wildlife’s Pathogen Containment Facility at the UC Davis Bodega Marine Laboratory in Bodega Bay, California. Red abalone, approximately 2.2 cm in shell length, were purchased from an abalone farm in Goleta, California. White abalone, approximately 1.8 cm in shell length, were donated from the UC Davis Bodega Marine Laboratory’s White Abalone Recovery Project. Prior to challenge, feces from all tanks were tested for 16S rRNA Ca. Xc genes by quantitative PCR (qPCR) following a validated protocol (Friedman et al., 2014b). Although fecal qPCR is treated only as a proxy for live pathogen, it has been shown to be the most sensitive assay for Ca. Xc detection (Friedman et al., 2014b). To support the absence of infection, five animals from both the red and white groups were sacrificed for histologic examination. Animals were supplied with a combination of wild-collected kelp (Macrocystis pyrifera) and cultured dulse (Palmaria palmata) two to three times per month throughout the experiment. Because Ca. Xc is known to be present in local abalone, feed was soaked in freshwater for at least 5 min prior to distributing to the tanks; our ongoing unpublished observations have demonstrated that this is sufficient to inactivate residual Ca. Xc that may be present on algal feed (CDFW unpublished observations). All tanks received constant 20-µm filtered, aerated, UV-irradiated, flow-through seawater.
Experimental design
This study was constructed as a fully nested design with tanks nested within temperatures and Ca. Xc exposure challenge and abalone nested within tanks. One hundred ninety-two red and 192 white abalone, ∼2 cm in length, were randomly and evenly distributed into either of the two treatment groups (exposed), or the control group (Fig. 1).

Figure 1: Experimental set up: Experimental variables are illustrated as follows: (A) Candidatus Xenohaliotis californiensis (Ca. Xc) exposed, 18.5 °C; (B) Ca. Xc exposed, 13.6 °C; (C) Control, 18.5 °C.
Experimental units–tanks–are represented as cylinders; red or white fill represents those stocked with red or white abalone respectively. Elevated (18.5 °C) and ambient (13.6 °C) seawater flowed from header tanks holding infected or uninfected animals for the first 161 days to transmit Ca. Xc.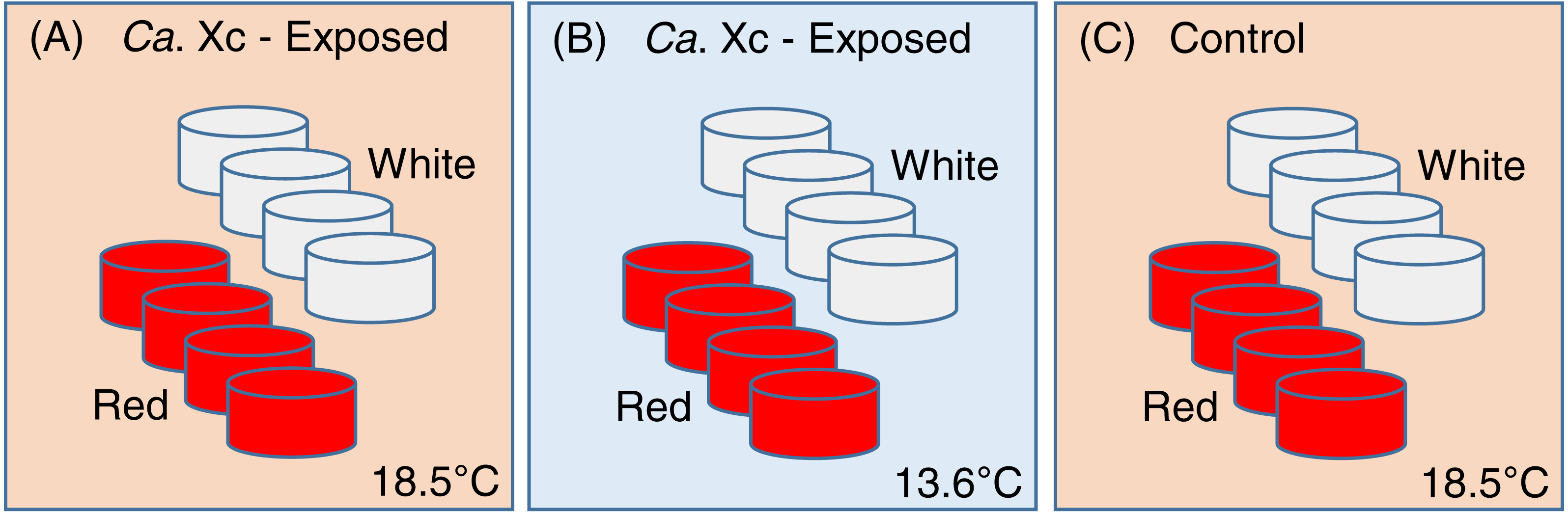{kind=link}
To avoid cross-contamination, we spatially organized the groups in lieu of random placing. Each treatment group was comprised of eight 3.8-L tanks, with eight animals housed in each tank. Two groups, one exposed to Ca. Xc and the other unexposed, received elevated temperature seawater (approximately 18.5 °C); a second Ca. Xc-exposed group received ambient water (approximately 13.6 °C). Temperature was measured hourly by automated temperature recorder placed in one tank per treatment group.
Exposure to Ca. Xc
To initiate Ca. Xc exposure, inflowing seawater was routed through 11-L conical header tanks with farmed red abalone prior to supplying the experimental tanks. The two exposed groups of each species received effluent water from a header tank containing eight farmed red abalone, each approximately 123 gm in weight, from a population shown by histology to be infected with Ca. Xc and its phage. The control groups of red abalone and white abalone were headed by a tank holding 50 farmed red abalone, approximately 2.4 gm in weight; these animals’ feces tested negative for Ca. Xc by PCR. The source water was directed through the headers through day 161 to ensure Ca. Xc exposure. Moore et al. (2001) showed that infection in red abalone was 100% after 111 days of Ca. Xc exposure to a header tank with infected red abalone at 18.5 °C (prior to appearance of the Ca. Xc phage).
Sampling schedule and processing
At selected time points (days 0, 62, 126, 161, 265, 343) all animals in the experiment were weighed and measured for shell length. A body mass Condition Index (CI) was calculated as: . At day 161, header tanks were removed and two randomly selected animals per tank were sacrificed and tested for Ca. Xc infection by qPCR from DNA extracted from post-esophagus (PE) tissue samples. Additionally, at day 161, six white abalone, three animals each from the heated and ambient exposed groups, were randomly selected and sacrificed for histological confirmation of transmission of Ca. Xc and its phage. At day 343, all surviving experimental animals were processed for analysis of infection in PE and digestive gland tissues by both qPCR and histology. For qPCR, PE tissue (∼30 mg) was excised from sacrificed animals and DNA extractions were performed using a DNeasy Blood and Tissue Kit (QIAGEN Germantown, MD) following the manufacturer’s protocol for pathogen detection. Tissue samples were processed for histology as previously described (Moore et al., 2001). Davidson’s-fixed (Shaw & Battle, 1957), hematoxylin- and eosin-stained 5 µm paraffin tissue sections containing PE and digestive gland were prepared from sacrificed animals. After termination of the experiment, slides were blindly assessed for presence/absence of Ca. Xc inclusions, and the inclusions present were categorized as having morphologies indicating phage infection (phage-containing) or lack of infection (classical).
Fecal bacterial sampling regimen and processing
Feces were collected from each tank bi-monthly for qPCR analysis. Feces from four tanks per group were pooled for Ca. Xc 16S rRNA gene detection. Fecal samples were weighed and frozen at −20 °C upon collection until analysis. DNA from fecal samples (∼250 mg) was extracted and purified with a QIAamp DNA Stool Mini Kit (QIAGEN) according to the manufacturer’s ‘Isolation of DNA from Stool for Pathogen Detection’ protocol. DNA obtained was eluted in 200 µl volumes and stored at −20 °C until analysis.
Quantitative PCR assays for Ca. Xc
We monitored Ca. Xc gene presence in post-esophagus and fecal samples using the methods developed and validated by Friedman et al. (2014a) and Friedman et al. (2014b). Standard curves were constructed using PCR product of the WSN1 primers: WSN1 F (5′AGTTTACTGAAGGCAAGTAGCAGA3′) and WSN1R (5′TCTAAC TTGGACTCATTCAAAAGC3′) and the P16RK3 plasmid (Friedman et al., 2014b). Plasmid concentration was quantified by Qubit fluorometer (ThermoFisher Scientific, Waltham, Massachusetts). Assayed tissue and fecal samples were considered positive if the mean copy number per ng of genomic DNA in triplicate samples was equal to or greater than one, and reactions prior to normalization calculations had at least three gene copies—as convention of Minimum Information for Publication of Quantitative Real-Time PCR Experiments guidelines describes (Bustin et al., 2009). For reporting purposes, and to meet assumption of residual normality for statistical analysis, reaction copy numbers were normalized by input DNA (ng) and log transformed.
Data analysis
A Cox proportional hazard model was used for survival analysis of treatment groups. A Chi-squared test was used to assess differences in survival of red abalone between this study and a historical study conducted prior to presence of the phage (Moore, Robbins & Friedman, 2000). Hazard ratio terminology refers to the likelihood of death associated with stratified variables: species, water temperature, Ca. Xc exposure. Variations in (1) condition index values and (2) qPCR data, were tested for significance by One-way ANOVA. Results of these models were assessed by post hoc Tukey comparisons; results are reported with estimates and standard deviations, describing the difference of the means and variance respectively from null hypothesis of the linear model. Residual errors from analysis were assessed for normality using the Wilk Shapiro test. Data from qPCR study was log-transformed. Both log-transformed qPCR data and condition index data was transformed by winsorization to meet the parametric test assumptions of normality. A test had a significant result if p ≤ 0.05 (level of significance α = 0.05). Statistical analysis was done with R version X R3.1.3 (R Core Team, 2014).
Results
Transmission of Ca. Xc
All Ca. Xc-exposed groups showed fecal shedding of Ca. Xc DNA after header tank removal from the system (day 161), indicating effective transmission to the exposed red abalone and white abalone under both temperature regimes. The unexposed tanks tested negative by feces PCR (Table 1).
| Group: species, treatment, temperature | Days | ||
|---|---|---|---|
| 0–113 | 126–236 | 251–342 | |
| Red, Ca. Xc-exposed, ambient | – | + | + |
| White, Ca. Xc-exposed, ambient | + | + | + |
| Red, Ca. Xc-exposed, elevated | + | + | + |
| White, Ca. Xc-exposed, elevated | + | + | NA |
| Red, control, elevated | – | – | – |
| White, control, elevated | – | – | – |
Notes:
- NA
-
no animals remain at this time period
+ greater or equal to 3 gene copies - less than 3 gene copies.
Pathogen transmission to all Ca. Xc-exposed tanks was confirmed by tissue qPCR from animals sacrificed at the end of the experiment with the exception of the Ca. Xc-exposed, elevated temperature white abalone which all died prior to day 343. Abalone that died of natural causes during the experiment were frozen and PE was excised and screened by qPCR for Ca. Xc. Of these animals, 16% from the control groups produced very low levels of Ca. Xc gene amplification (<three copies/ng input DNA). The source of the potential contamination is unknown. Other measurements from fecal qPCR, tissue qPCR from sacrificed animals, and histology assessments did not indicate transmission of Ca. Xc. to the control groups.
Analysis of survival
We examined survival in response to water temperature, abalone species, and Ca. Xc exposure (Fig. 2).
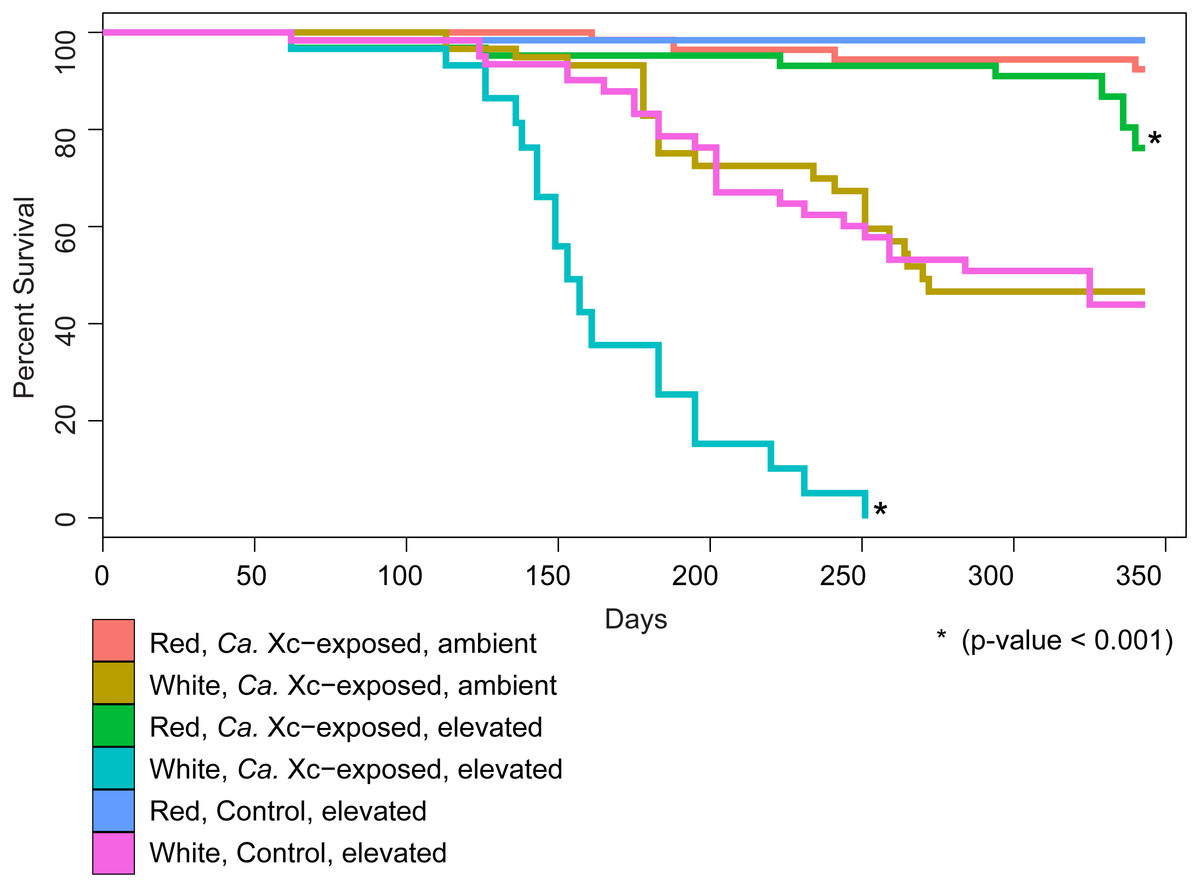
Figure 2: Percent survival of red and white abalone held in ambient water (13.6 °C) or at elevated temperature (18.5 °C) with and without Candidatus Xenohaliotis californiensis (Ca. Xc) exposure for 343 days; N = 64 for each group at day.
Each curve represents one of the six treatment groups, with variables: Ca. Xc exposure, seawater temperature, and species. We observe a significant difference between survival curves of red and white abalone held under elevated seawater temperature and Ca. Xc exposure conditions.{kind=link}
Our analysis indicated that white abalone exposed to Ca. Xc and elevated temperature were 10.9 times more likely to die than red abalone held in the same conditions (hazard ratio = 10.9; 95% CI [5.97–19.87]; P < 0.001). By day 251, all Ca. Xc-exposed white abalone in the elevated temperature treatment died, while red abalone held under the same conditions maintained a survival rate of 91%. Elevated temperature increased mortality risk for both red and white abalone 3.3 times that of the ambient treatment groups (hazard ratio = 3.3; 95% CI [1.1–10.2]; P = 0.039). Specifically, under elevated temperature, Ca. Xc-exposure increased the mortality risk of both species of abalone 12.5 times (hazard ratio = 12.5; 95% CI [1.6–96.0]; P = 0.015). Generally, under all conditions, white abalone had a risk of mortality that was 41 times that of red abalone (hazard ratio = 41.0; 95% CI [5.6–303.1]; P < 0.001).
| Average seawater temperature | N | Cumulative mortality events at day = 220 | Percent survival | |
|---|---|---|---|---|
| Current study | 18.5 | 46 | 4 | 91% |
| Moore, Robbins & Friedman (2000) | 18.5 | 30 | 10 | 67% |
The survival rate of red abalone in the current study, exposed to Ca. Xc with phage, was 36% higher than the previous experiment, in which animals were exposed to phage-free Ca. Xc (Table 2) (Moore, Robbins & Friedman, 2000). We evaluated survival at day 220, immediately prior to termination day in the 2000 study (Moore, Robbins & Friedman, 2000). Between Days 1 and 220, in both experiments, the average water temperature was 18.5 °C (Moore, Robbins & Friedman, 2000). Both experiments used farm-origin juvenile red abalone. The 60 animals in Moore, Robbins & Friedman’s (2000) study averaged 8 cm in length and were selected from a farmed population with a known low-intensity Ca. Xc infection; however, this population did not express clinical symptoms of WS (Moore, Robbins & Friedman, 2000). Comparison of survival in the historical (phage-free) and current (phage-containing) experimental data by Chi-squared test shows a significant difference (X-squared = 25.37, df = 1, P < 0.001) favoring survival in the presence of phage.
Body condition indices
Assessment of animal health—as a function of weight, normalized by length—over time showed that red abalone remained heathy under all experimental conditions, unlike their white abalone counterparts (Fig. 3).
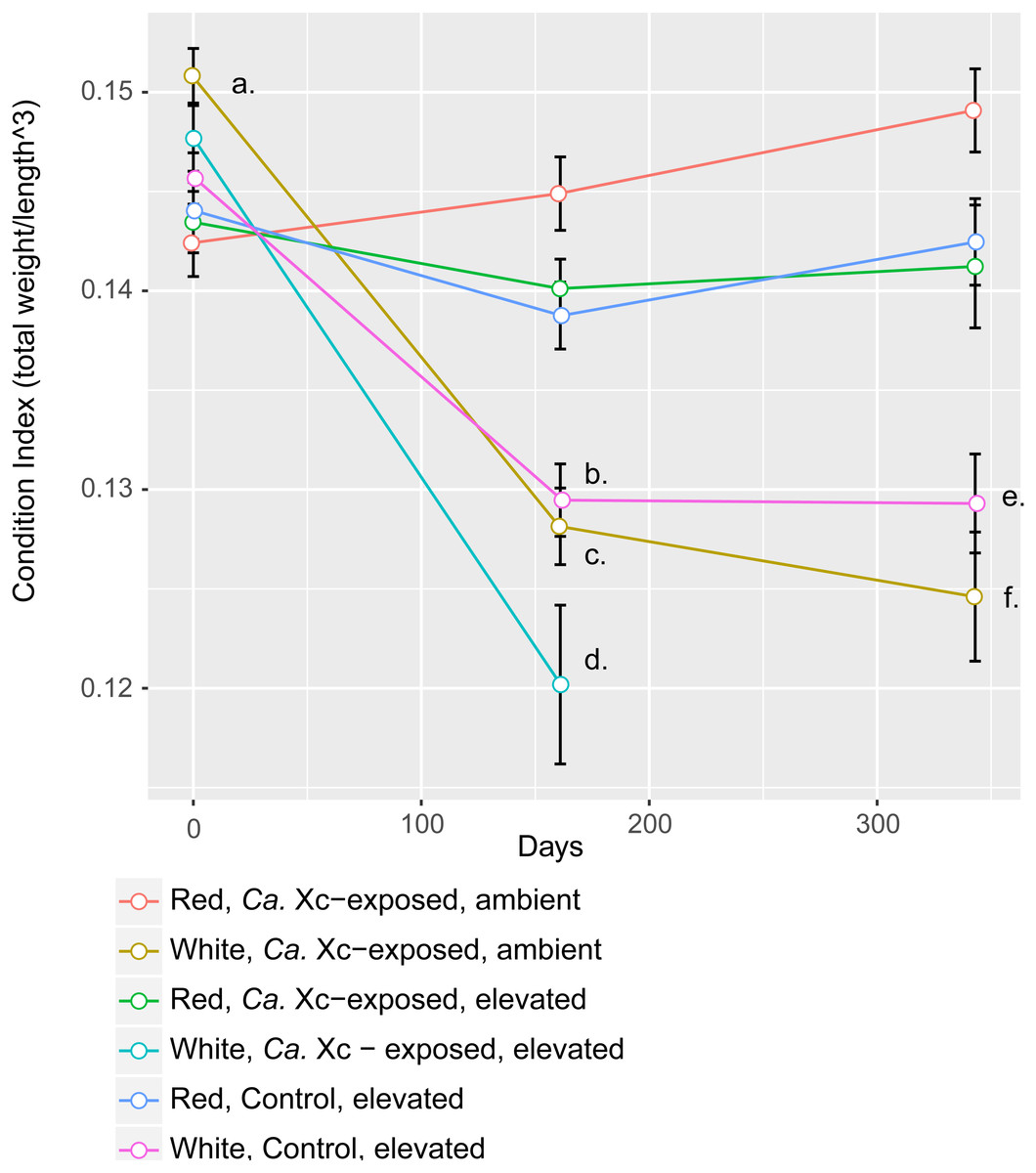
Figure 3: Longitudinal plot of mean values in Condition Index (CI), calculated by total abalone weight divided by shell length cubed, over time of experimental groups.
Error bars represent standard error of the mean. (A) Initial CI values of white abalone, destined for Ca. Xc exposure were significantly greater than the other groups. (p-values < 0.05) (B) CI values of white, control abalone were significantly lower than red, control abalone (p-value = 0.007) (C) CI values of white, Ca. Xc-exposed, ambient abalone were significantly lower than red, Ca. Xc-exposed, ambient abalone (p-value < 0.001) (D) CI values of white, Ca. Xc-exposed, elevated abalone were significantly lower than red, Ca. Xc-exposed, elevated abalone (p-value < 0.001) (E) CI values of white, control abalone were significantly lower than red, control abalone (p-value = 0.007) (F) CI values of white, Ca. Xc-exposed, ambient abalone were significantly lower than red, Ca. Xc-exposed, ambient abalone (p-value < 0.001).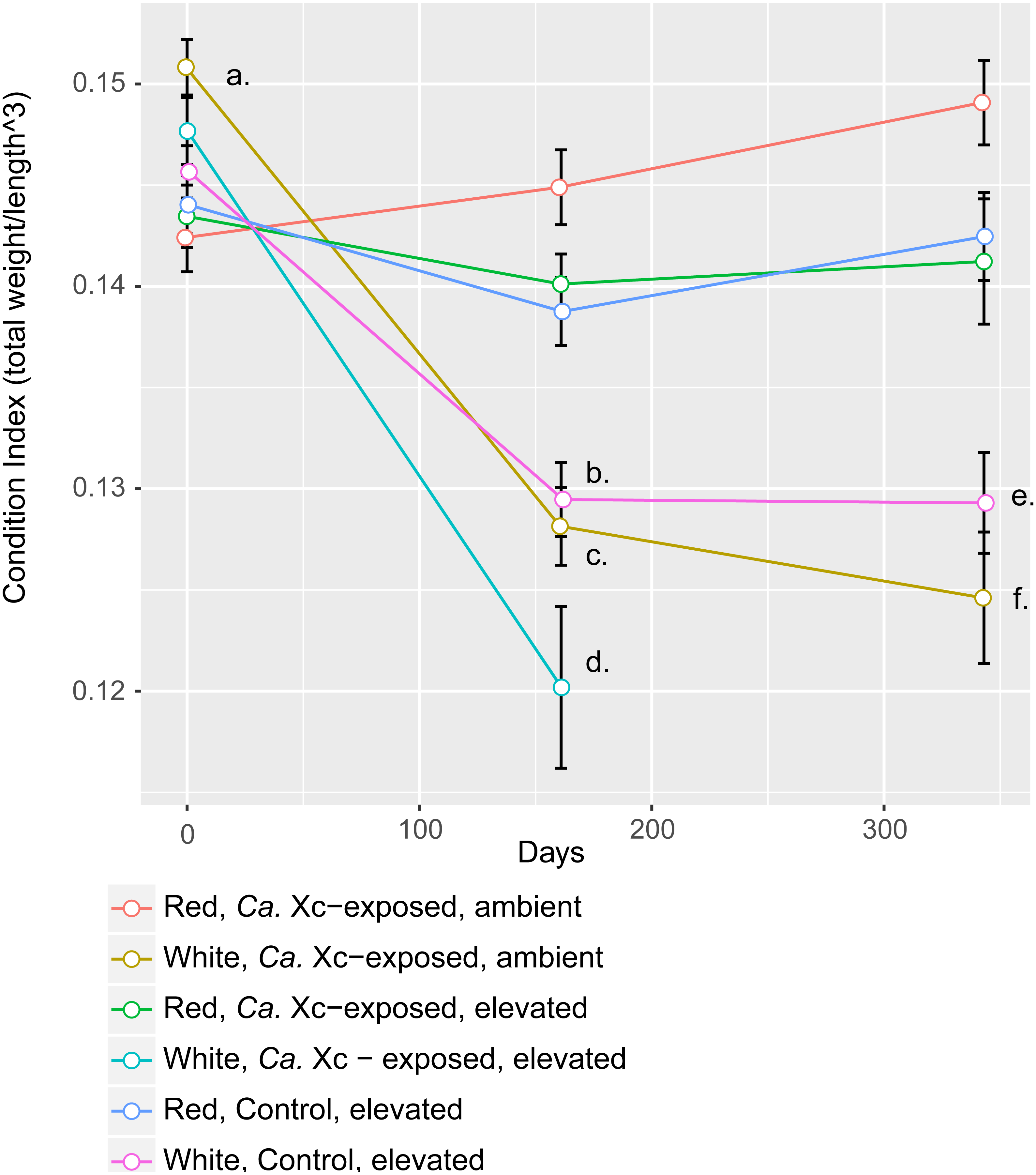{kind=link}
While data were collected at additional time points, we focused on three—beginning, mid, and end—for visual clarity. At the outset of the experiment (Day 0), the only group that showed a significant difference in CI values was the white, ambient group destined for Ca. Xc exposure, with a greater condition index values than the other five groups (Estimate 0.008, Std. Error 0.002, t-value = 3.950, P < 0.001). Mid-way through the experiment (Day 161) all three groups of white abalone overall had lower mean condition index values than their red counterpart groups. Statistical analysis showed significant differences between control, elevated temperature red and white groups (Estimate −0.009, Std. Error 0.003, t-value = −3.497, P < 0.007) with somewhat greater differences between the exposed red and white groups in both elevated temperature (Estimate −0.020, Std. Error 0.003, t-value = −6.003, P < 0.001) and ambient (Estimate −0.017, Std. Error 0.003, t-value = −6.459, P < 0.001) seawater treatments. However, no differences were observed between elevated temperature and ambient white, Ca. Xc-exposed groups (Estimate −0.008, Std. Error 0.003, t-value = −2.386, P = 0.162) more than half of the exposed elevated temperature white abalone had died prior to this time point. Based on visualization of longitudinal changes, white abalone fared worse under all treatments; Ca. Xc exposure appeared to be associated with increased withering in white abalone, while red abalone did not appear to decrease in body condition in response to Ca. Xc exposure. At the end of the experiment (day 343), white and red abalone under the same treatment of Ca. Xc-exposure in ambient seawater showed significant differences in body condition (Estimate −0.024, Std. Error 0.004, t-value = −5.823, P < 0.001). However, we observed significant, yet smaller differences between the control groups (Estimate −0.013, Std. Error 0.004, t-value = −3.242, P < 0.0121). There was not a significant difference in white abalone body condition between the control and Ca. Xc-exposed, ambient, groups (Estimate 0.004, Std. Error 0.005, t-value = 0.955, P = 0.872); additionally, there was no difference between red abalone in the control and the exposed, elevated temperature groups (Estimate 0.001, Std. Error 0.003, t-value = 0.369, P = .996).
Candidatus Xc prevalence and infection intensity
Candidatus Xc 16S rRNA gene copy numbers obtained by qPCR from PE tissue can serve as a proxy for bacterial burden, and were assessed at days 161 and 343 (Fig. 4).

Figure 4: Log transformed qPCR-derived Candidatus Xenohaliotis californiensis gene copy numbers from PE tissue at days 161 and 343.
(A) At day 161, no Ca. Xc genes amplified in the red, ambient group (B) No Ca. Xc genes amplified in the control groups (C) Prior to day 343, all abalone in the white, Ca. Xc- exposed, elevated group died.{kind=link}
The qPCR data from tissue samples taken on day 161 were too skewed to transform such that residuals met normality assumptions for appropriate parametric statistical analysis. However, visualization of data trends suggests that at day 161, elevated temperature resulted in higher pathogen tissue burdens in white abalone (Fig. 4); the mean Ca. Xc gene copy number per ng DNA from tissue samples of exposed white abalone in the elevated temperature regimen was 4,248; 326 times greater that of their red counterparts. Notably, white animals sacrificed at this time point were survivors—60% of their cohort already died. There was no Ca. Xc amplification in the red, exposed ambient group at day 161. At day 343, red abalone in the exposed, ambient group had significantly lower pathogen gene numbers than (a) the exposed elevated temperature red group (Estimate 2.485, Std. Error 0.3167, t-value =7.845, P < 0.001) and (b) their exposed, ambient white counterparts (Estimate 2.045, Std. Error 0.3167, t-value =6.475, P < 0.001). No Ca. Xc DNA was amplified from PE tissue samples of sacrificed animals in the unexposed groups at either time point.
At day 161, three white abalone from each of the exposed ambient and exposed elevated temperature groups were sacrificed and used for histopathology to corroborate transmission data from fecal and tissue qPCR samples and determine whether any Ca. Xc inclusions present included those with morphology indicating phage infection. For the first time, we morphologically identified the phage-containing inclusions in white abalone (Fig. 5).
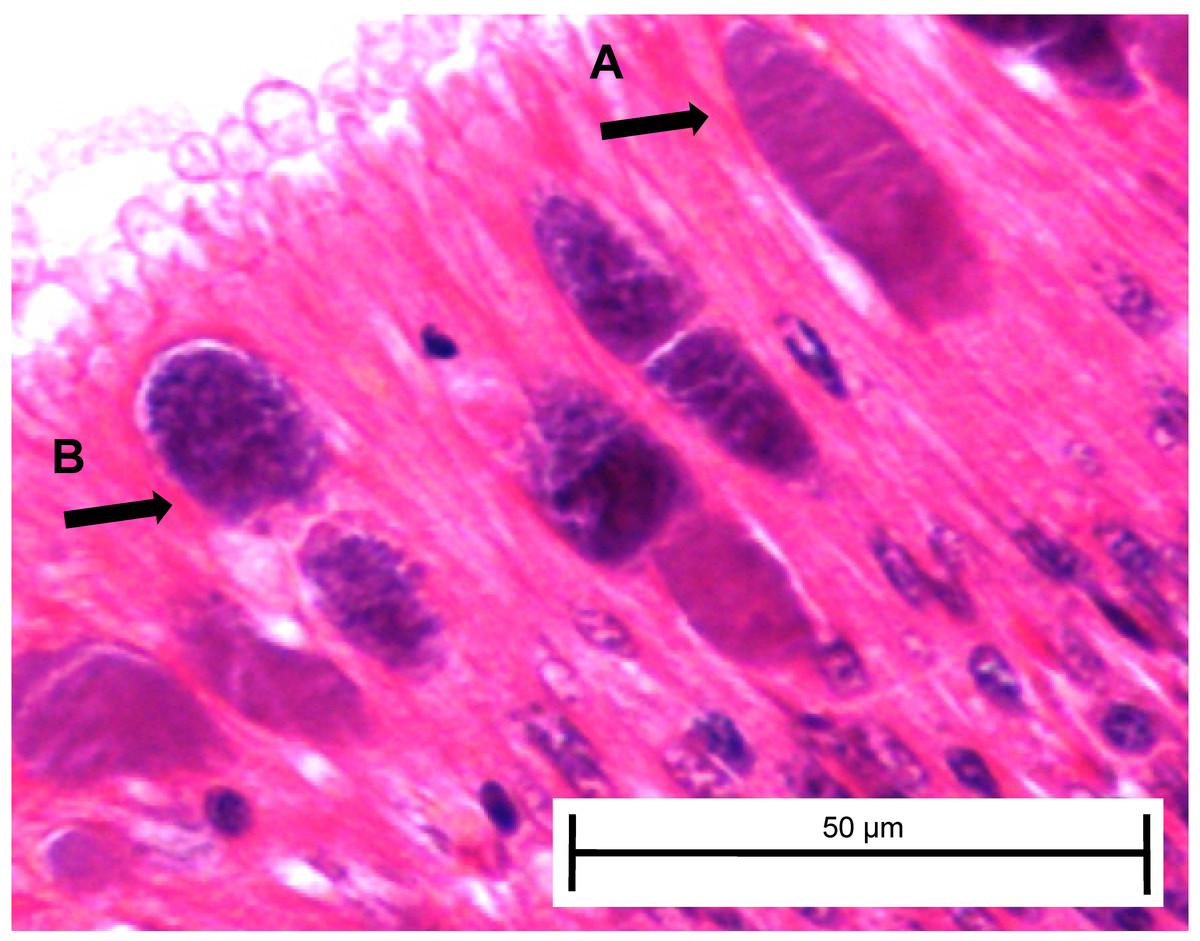
Figure 5: Candidatus Xenohaliotis californiensis inclusions within posterior esophagus epithelia from white abalone held at 18.5 °C and Ca. Cx exposed at 161 days.
Arrows indicate (A) classical inclusions, (B) Phage-containing variant inclusions. Haematoxylin and eosin. Bars, 50 µm.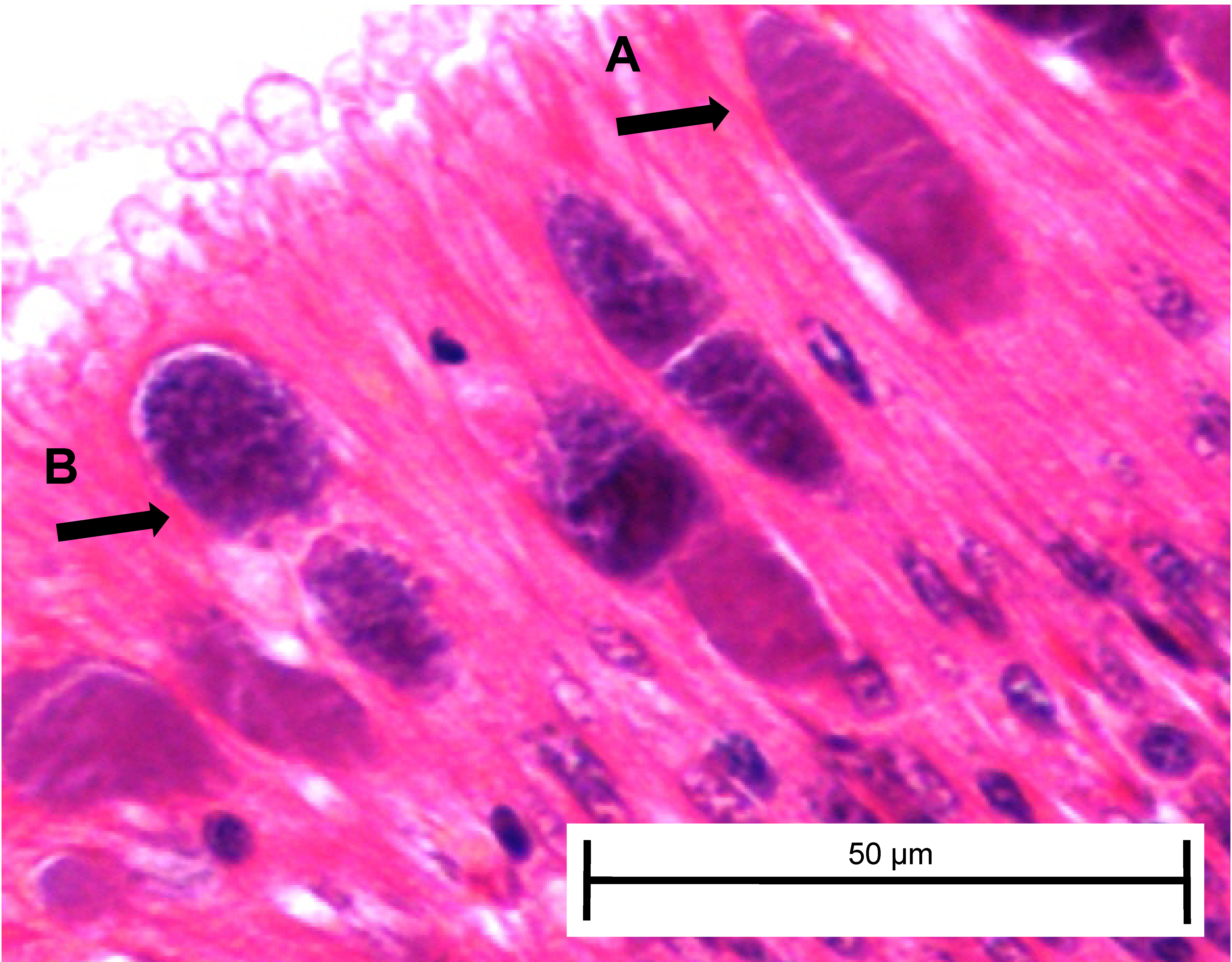{kind=link}
Histological examination of day 343 samples showed the presence of Ca. Xc classical inclusions and phage-containing inclusions in the red, elevated temperature and white, ambient temperature groups (Table 3). In concordance with the qPCR data, we observed nearly three times as many classical inclusions and more than five times as many phage-containing inclusions in post-esophagus samples from the exposed red, elevated temperature group compared to the exposed white, ambient group. Candidatus Xc inclusions (both classical and phage-containing) in digestive gland tissue were only observed in the Ca. Xc-exposed, elevated temperature, red abalone group.
| Species, treatment, temperature condition | Post-esophagus | Digestive gland | ||||
|---|---|---|---|---|---|---|
| Inclusion type* | Inclusion type* | |||||
| N | Classical | Phage-variant | N | Classical | Phage-variant | |
| Red, Ca. Xc-exposed, ambient | 34 | 0% | 0% | 54 | 0% | 0% |
| White, Ca. Xc-exposed, ambient | 14 | 29% | 14% | 19 | 0% | 0% |
| Red, Ca. Xc-exposed, elevated | 35 | 89% | 77% | 46 | 20% | 15% |
| White, Ca. Xc-exposed, elevated | 0 | NA | NA | 0 | NA | NA |
| Red, control, elevated | 27 | 0% | 0% | 62 | 0% | 0% |
| Red, control, elevated | 8 | 0% | 0% | 22 | 0% | 0% |
Notes:
Discussion
The initial goal of this study was to investigate potential diminished pathogenicity of Ca. Xc in association with phage presence. However, the clear differences in expression of WS between white and red species is a key and unexpected finding. The white abalone control group was adversely affected by the heated seawater—as observed by body condition and survival of the control group—perhaps contributing evidence for the frailty of the species. White abalone, exposed to Ca. Xc and held in 18.5 °C seawater, exhibited the highest mortality rate and most pronounced clinical signs of withering syndrome among the experimental groups, thus appearing to be highly susceptible to WS infection and disease, even with current phage-presence. However, under conditions that previously had been shown to exacerbate disease, the effects of WS from the current variant of Ca. Xc in association with the phage are reduced in red abalone relative to a similar study conducted more than a decade beforehand when no phage was evident.
Transmission of Ca. Xc was slowest in the red, ambient group based on fecal and tissue qPCR values. Variables that influenced fecal production and degradation, primarily feeding schedule, impaired normalization and thus interpretation accuracy of qPCR quantitative data, and therefore Table 1 summarizes only the presence/absence of Ca. Xc genes. Although, previous work has demonstrated PCR to be the most sensitive Ca. Xc detection method, it should be mentioned that due to assay sensitivity limitations, contamination in the control groups may have gone unidentified. In accordance with our results, slow infection progression in cold water has been show in previous studies (Braid et al., 2005; Moore, Robbins & Friedman, 2000). While the Moore, Robbins & Friedman (2000) study identified Ca. Xc inclusions in 90% of experimental animals held at ambient temperature (14.7 °C) at day 220, in the current study no inclusions were found by histology in the exposed red abalone held at 13.6 °C sacrificed on day 343. This finding from Moore, Robbins & Friedman’s (2000) study may be the result of ‘natural’ farm-associated transmission at a time point prior to the start of the study but it may also be indicative of a more virulent Ca. Xc strain. Additionally, the small histology subsample taken at day 161 from white abalone, held under ambient conditions did not have inclusions. We speculate that phage presence may also have extended the disease incubation period in the ambient temperature groups.
From weight and length data, we can infer that both the white and red abalone destined for Ca. Xc-exposure and elevated temperature treatment started with robust body condition, and thus were not at a disadvantage that would have predisposed them to withering and death. The similarity between the elevated temperature, control and ambient exposed white groups’ survival curves and condition index values supports the general sensitivity of these animals to adverse conditions and may also be a result of low genetic diversity (Gruenthal & Burton, 2005). The pronounced effects of temperature may be explained by white abalone’s natural habitat; they are a deeper-dwelling species and experience less fluctuation in water temperature than other species that inhabit intertidal and shallower subtidal zones.
In order to examine the impact of the phage on Ca. Xc pathogenicity, it would of course have been ideal to directly compare the phage-containing and uninfected pathogens. However, currently this is not believed possible because classical inclusions are consistently accompanied by phage-containing ones. This has been recorded in recent histology assessments from abalone populations in Baja Mexico and Cayucos, San Nicolas Island, and Carmel California (Cruz-Flores et al., 2016; Friedman et al., 2014a) and has been observed in abalone from Bodega Head (J Moore, pers. obs., 2015). Consequently, we attempted to replicate a thermal induction study undertaken with red abalone prior to appearance of the phage (Moore, Robbins & Friedman, 2000). Comparing our results with those from that study strongly suggest that red abalone are better able to withstand withering syndrome now than in years previous to phage presence. Anecdotal observations from California red abalone farms support this conclusion (J Moore, 2002, unpublished data).
However, these differences may be the result of other changes in the system, particularly as red abalone have been challenged by the disease for nearly 30 years. For example, red abalone may have developed heritable traits that confer immunity, as suggested by Brokordt et al. (2017). Alternatively, it is possible that Ca. Xc may have evolved to become less pathogenic through any number of mechanisms, including prophage; such a change cannot be elucidated by the experimental design of this study and thus we cannot decipher those mechanisms or causations. Recently, analysis of a subset of Ca. Xc coding genes showed an absence of genetic variation among samples taken from different geographic locations and from different abalone species; suggesting that genetic variability in the bacterium may have a limited contribution to the differing pathogenic effects we observe in this system (Cicala et al., 2018). A genome wide analysis might identify potentially attenuating virulence factors, which could be further investigated for population variation as explored in the (Cicala et al., 2018) study; however, whole genome sequence of Ca. Xc has yet to be published.
In this study red abalone exposed to the Ca. Xc showed no difference in body condition from their control counterparts, suggesting the limited pathogenic effects of the current variant Ca. Xc, or at a minimum the WS expression was significantly delayed. This is in contrast to the white abalone, for which we observed trends in body condition and survival that appear to be directly related to Ca. Xc gene copy numbers detected by qPCR. At the mid-point of the experiment, coinciding with the highest mortality rate and lowest body condition index values, Ca. Xc-exposed white abalone in the elevated temperature group also showed the greatest bacterial burden by qPCR. Despite any measured evidence in body shrinkage in the red groups, at the end of the experiment, Ca. Xc-exposed red abalone in the elevated temperature group had the highest pathogen gene copy and after day 300, the Ca. Xc-exposed, elevated temperature red group’s mortality rate appeared to increase; this might be indicative of disease expression after a nearly yearlong incubation period. The high level of detection of Ca. Xc DNA by tissue qPCR and histological changes at the end of the experiment could be associated with this apparent increase in mortality.
Our molecular data was supported by histological analysis. At day 343, Ca. Xc inclusions were only found in tissue samples from the two groups with the highest qPCR Ca. Xc gene amplification. Red abalone had a greater ratio of phage-containing inclusions to classical inclusions. However, it is difficult to assess and isolate the impact of temperature and species on our observations because we were only able to compare the red, elevated temperature group with the white, ambient group. While this study did not confirm phage particles by TEM, the highly-recognizable granular, pleomorphic inclusions have consistently revealed observable phage (Cruz-Flores & Cáceres-Martínez, 2016; Cruz-Flores et al., 2016; Friedman et al., 2014a). The mottled, complex morphology of the phage-containing inclusions has been interpreted as evidence of active phage-induced lysis (Friedman & Crosson, 2012). Inclusions with this morphology were observed in our experimental white abalone; this study is the first to document that white abalone are able to harbor the phage-containing variant of Ca. Xc.
Conclusion
The results of this study have implications for restoration strategies to ultimately remove white abalone from the US Endangered Species list. The federal white abalone recovery plan concluded that outplanting of hatchery-produced animals must be the key restoration action for successful recovery of the species (Team TWAR, 2008). White abalone in warm water appear highly susceptible to the current variant of Ca. Xc, and outplanting efforts should take place in cooler water to minimize thermal enhancement of disease expression. The white abalone captive breeding program has recently introduced new wild-origin to its broodstock pool, which may increase genetic variation and render the progeny to be less sensitive to natural environmental stressors and possibly more resistant to the effects of Ca. Xc. Studying heritable variation in susceptibility of white abalone families would be highly informative to the captive breeding efforts. In the Southern California Bight, cooler water typically translates to deeper water but also certain geographic locations with strong upwelling such as San Miguel Island (Erlandson et al., 2008). Our findings with red abalone align with the anecdotal reports from California abalone farmers that disease caused by Ca. Xc has been much less frequent and severe since the phage has been observed. We conclude that the current form of Ca. Xc, with its phage present, is associated with improved health and survival in red abalone under conditions that have previously exacerbated the disease. However, the stability of this development is unknown. Further investigation of the genome and the Ca. Xc phage’s phylogenetic relationships may shed light on the some of the host-pathogen-phage interactions and provide an explanation for the observed effects associated with the current pathogen variant. Future efforts may be directed towards whole genome sequencing of Ca. Xc and annotation of all mobile genetic elements of both the bacteria and the phage such that strains can be characterized and better understood to elucidate the mechanisms associated with pathogenicity in this important marine system.