
Tobacco rattle virus-induced PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH) gene silencing in Solanum pseudocapsicum L.
- Published
- Accepted
- Received
- Academic Editor
- Savithramma Dinesh-Kumar
- Subject Areas
- Agricultural Science, Biotechnology, Molecular Biology, Plant Science
- Keywords
- PHYTOENE DESATURASE (PDS), VIGS, Mg-chelatase H subunit (ChlH), Solanum pseudocapsicum L., TRV, Reference genes, Gene expression
- Copyright
- © 2018 Xu et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Tobacco rattle virus-induced PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH) gene silencing in Solanum pseudocapsicum L. PeerJ 6:e4424 https://doi.org/10.7717/peerj.4424
Abstract
Virus-induced gene silencing (VIGS) is an attractive tool for determining gene function in plants. The present study constitutes the first application of VIGS in S. pseudocapsicum, which has great ornamental and pharmaceutical value, using tobacco rattle virus (TRV) vectors. Two marker genes, PHYTOENE DESATURASE (PDS) and Mg-chelatase H subunit (ChlH), were used to test the VIGS system in S. pseudocapsicum. The photobleaching and yellow-leaf phenotypes of the silenced plants were shown to significantly correlate with the down-regulation of endogenous SpPDS and SpChlH, respectively (P ≤ 0.05). Moreover, the parameters potentially affecting the efficiency of VIGS in S. pseudocapsicum, including the Agrobacterium strain and the inoculation method (leaf syringe-infiltration, sprout vacuum-infiltration and seed vacuum-infiltration), were compared. The optimized VIGS parameters were the leaf syringe-infiltration method, the Agrobacterium strain GV3101 and the growth of agro-inoculated plants at 25°. With these parameters, the silencing efficiency of SpPDS and SpChlH could reach approximately 50% in S. pseudocapsicum. Additionally, the suitability of various reference genes was screened by RT-qPCR using three candidate genes, and the results demonstrated that glyceraldehyde 3-phosphate dehydrogenase (GAPDH) can serve as a suitable reference for assessing the gene expression levels of VIGS systems in S. pseudocapsicum. The proven application of VIGS in S. pseudocapsicum and the characterization of a suitable reference gene in the present work will expedite the functional characterization of novel genes in S. pseudocapsicum.
Introduction
Solanum pseudocapsicum L., which belongs to the genus Solanum and the family Solanaceae, possesses high ornamental value and is widely cultivated as an indoor ornament due to its bright red berries at maturity (Aliero, Grierson & Afolayan, 2006b). This plant also has important pharmaceutical value, as reflected by its use for the treatment of boils, gonorrhoea and acute abdominal pain (Aliero, Grierson & Afolayan, 2006a). The total alkaloid fraction from S. pseudocapsicum has hepatoprotective properties and antitumour, antifungal, and antimicrobial activities (Mitscher, Juvarkar & Beal, 1976; Badami et al., 2003; Vijayan et al., 2003; Aliero, Grierson & Afolayan, 2006b). An ethyl acetate extract from S. pseudocapsicum showed promising antifeedant and insecticidal activities (Jeyasankar, Premalatha & Elumalai, 2012). Despite the great ornamental and pharmaceutical value of S. pseudocapsicum, scarce molecular-level information is currently available due to the lack of gene function identification techniques and the unavailability of related genome sequences. Gene function studies would yield a better understanding of the molecular mechanisms that regulate trait development in S. pseudocapsicum and could provide a gene source for molecular breeding and drug development. Consequently, the development of techniques for identifying the functions of genes in S. pseudocapsicum is vital.
Virus-induced gene silencing (VIGS) is a recently developed gene transcript suppression technique for characterizing gene function in plants (Burch-Smith et al., 2004). This technique has many advantages for unravelling gene function compared with the loss-of-function approaches that are currently available for plants, and these include easy and rapid gene silencing, its transformation-independent nature, and the requirement of only some sequence information (Burch-Smith et al., 2004). Increasing numbers of viruses have recently been used to derive VIGS vectors for unravelling gene functions, such as tobacco rattle virus (TRV) (Ratcliff, Martin-Hernandez & Baulcombe, 2001), barley stripe mosaic virus (BSMV) (Holzberg et al., 2002), cabbage leaf curl virus (CaLCuV) (Muangsan et al., 2004), potato virus X (PVX) (Faivre-Rampant et al., 2004) and cucumber mosaic virus (CMV) (Tasaki et al., 2016). Among these viral vectors, TRV has been widely used to construct VIGS vectors for silencing target genes in various plant species, such as Arabidopsis thaliana (Burch-Smith et al., 2006), Nicotiana benthamiana, Solanum lycopersicum (Liu, Schiff & Dinesh-Kumar, 2002), and Capsicum annuum (Chung et al., 2004), because it has a wide host range and induces mild infection symptoms. However, it is unknown whether TRV-based VIGS can be applied to unravel gene functions in S. pseudocapsicum. Although the sequence of the S. pseudocapsicum genome is not currently available, the genome sequences of other species belonging to the Solanum genus (Solanaceae), such as S. lycopersicum and C. annuum (Lin et al., 2014; Qin et al., 2014), have been published. Thus, the target gene sequence of S. pseudocapsicum can be obtained through homology-based cloning, making the identification of gene functions in S. pseudocapsicum possible. PDS encodes an important enzyme in carotenoid biosynthesis, and the ChlH gene encodes the H subunit of magnesium-protoporphyrin chelatase, which is involved in chlorophyll biosynthesis. Both the PDS and ChlH genes are commonly used as markers of VIGS because the silencing of one of these genes yields a photobleached and a yellow-leaf phenotype, respectively (Cunningham & Gantt, 1998; Hiriart et al., 2002; Liu et al., 2012). To assess whether the TRV-based VIGS system could be used to identify gene functions in S. pseudocapsicum, we investigated a TRV-induced PDS and ChlH silencing protocol involving agro-infiltration in S. pseudocapsicum. Moreover, the effects of the Agrobacterium strain, the inoculation methods and the growth temperature after Agrobacterium infiltration on S. pseudocapsicum were compared.
Reverse transcription quantitative real-time PCR (RT-qPCR) is a standard tool for the quantification of gene expression levels. In the present study, RT-qPCR was used to detect the PDS and ChlH gene expression levels. It is well known that the accuracy of RT-qPCR techniques is challenged by many sources of variation, including template quality, sampling errors, and amplification efficiency (Bustin, 2002). The selection of a suitable reference gene is vital in RT-qPCR analysis because it ensures the reliability of the RT-qPCR results (Vandesompele et al., 2002). Housekeeping genes are commonly used as reference genes for normalizing RT-qPCR analyses in molecular biology (Hong et al., 2008). For example, genes encoding actin (ACTIN), polyubiquitin (UBQ), and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are widely used as reference genes in gene expression studies (Czechowski et al., 2005; Jain et al., 2006). To date, a reference gene in S. pseudocapsicum has not been identified. Statistical algorithms such as geNorm, BestKeeper and NormFinder have been developed to evaluate the best-suited reference genes for normalizing real-time PCR data from a given set of biological samples (Vandesompele et al., 2002; Andersen, Jensen & Orntoft, 2004; Pfaffl et al., 2004). Thus, three reference genes (ACTIN, GAPDH and UBQ) identified in previous studies were selected as candidates in the present study for stability screening using the geNorm, BestKeeper and NormFinder algorithms prior to VIGS.
This study established a TRV-based VIGS protocol and provided the first screening of appropriate reference genes for S. pseudocapsicum. The results will expedite the characterization of gene expression levels and the analysis of gene functions in S. pseudocapsicum, which would allow the exploitation of desirable genes in S. pseudocapsicum.
Materials and Methods
Plant materials
Seeds from S. pseudocapsicum were soaked in water at 25 °C for five days and then sown in pots containing an autoclaved medium consisting of peat and vermiculite (at a ratio of 3:1). The seedlings were grown in a growth chamber (25 °C, 60–70% relative humidity, 16-h light/8-h dark cycle).
For the VIGS experiments, leaves exhibiting the silencing phenotype (only the phenotypic areas) were collected from five of the silenced seedlings, and leaves were also harvested from five mock-treated and untreated seedlings at the second-true-leaf stage. The collected leaves were used for analysis of the expression of PDS and ChlH genes.
For the screening of reference genes, leaves at different development stages (cotyledons stage, first-true-leaf stage and second-true-leaf stage), stems and roots were sampled to analyse the expression of the candidate reference genes. Samples were obtained from five different plants, three independent biological replicates were included for each sample, and each experiment was repeated three times. All the samples were immediately frozen in liquid nitrogen and maintained at −80 °C until RNA extraction.
RNA extraction and RT-PCR
Total RNA was extracted using an RNAprep Pure Plant Kit (Polysaccharide-& Polyphenolic-rich) (TIANGEN, Beijing, China) according to the manufacturer’s instructions, and any DNA contamination was removed with RNase-free DNase I (TIANGEN, Beijing, China). The RNA integrity was confirmed by denaturing 1.2% agarose gel electrophoresis, and the quantity and purity were determined with a Q3000 UV spectrophotometer (Quawell Technology Inc., San Jose, CA, USA). Only high-quality samples with an A260/A230 > 2.0 and 1.8 < A260/A280 < 2.1 were used for cDNA synthesis. First-strand cDNA was synthesized using 1 µg of total RNA using the TransScript® One-Step gDNA Removal and cDNA Synthesis SuperMix Kit (TRAN, Beijing, China).
Cloning of the candidate reference genes and the PDS and ChlH genes
To acquire the S. pseudocapsicum orthologues of the reference genes as well as PDS and ChlH genes that were previously reported in other plant species, publicly available gene fragments, including ACTIN, GAPDH, UBQ, PDS and ChlH, from species in Solanaceae (tomato, pepper, tobacco and potato) were downloaded. The ACTIN, GAPDH, UBQ, PDS and ChlH gene fragments in S. pseudocapsicum were amplified by PCR using homology-based cloning as previously reported (Zhong et al., 2014). Primers for the cloning of SpACTIN were designed according to the conserved regions among S. lycopersicum (SlACTIN, NM_001330119.1), C. annuum (CaACTIN, XM_016683691.1), Solanum tuberosum (StACTIN, XM_015308091.1) and N. tabacum (NtACTIN, EU938079.1); primers for the cloning of SpGAPDH were designed according to the conserved regions among S. tuberosum (StGAPDH, XM_006352526.2), C. annuum (CaGAPDH, NM_001324619.1), N. tabacum (NtGAPDH, XM_016612593.1) and S. lycopersicum (SlGAPDH, XM_004248266.3); primers for the cloning of SpUBQ were designed according to the conserved regions among C. annuum (CaUBQ, AY486137.1), S. tuberosum (StUBQ, XM_015307154.1), S. lycopersicum (SlUBQ, XM_019214130.1) and N. tabacum (NtUBQ, XM_016589747.1); primers for the cloning of SpPDS were designed according to the conserved regions among S. lycopersicum (SlPDS, EF650011.1), C. annuum (CaPDS, NM_001324813.1) and N. tabacum (NtPDS, DQ469932.1); and primers for the cloning of SpChlH were designed according to the conserved regions among N. tabacum (NtChlH, NM_001325713.1), S. lycopersicum (SlChlH, XM_015217369.1) and C. annuum (CaChlH, XM_016716472.1). RT-PCR was performed using the cDNA of leaves of S. pseudocapsicum as the template. The RT-PCR temperature protocol was as follows: (1) 95 °C for 5 min, (2) 35 cycles of 95 °C for 30 s, 55 °C for 30 s and 72 °C for 1 min, and (3) maintenance at 72 °C for 2 min. To obtain the longer PDS gene fragment, the PDS gene sequence was further cloned by 3′-rapid amplification of cDNA ends (3′-RACE) using the GeneRacer™ kit (Invitrogen, Carlsbad, CA, USA). The gene-specific primers (GSPs) for 3′-RACE were designed based on the sequence of the PDS fragment amplified using homology-based cloning according to the manufacturer’s instructions provided with the GeneRacerTM kit. The reaction programmes were as followers: (1) 95 °C for 3 min, (2) 35 cycles of 98 °C for 20 s, 58 °C for 15 s and 72 °C for 2 min, and (3) maintenance at 72 °C for 2 min. The above-mentioned primer sequences are shown in Table S1.
Analysis of gene expression by RT-qPCR and detection of TRV by RT-PCR
Real-time PCR was performed using SYBR Premix Ex Taq (Takara, Japan). The primers used to amplify the ACTIN, GAPDH, UBQ, PDS and ChlH segments are shown in Table 1. The reaction programmes were as followed: (1) 95 °C for 1 min, (2) 40 cycles of 95 °C for 20 s, 60 °C for 10 s, and 72 °C for 25 s, and (3) a melt curve programme (65 °C to 95 °C with an increment in temperature of 0.5 °C every 0.05 s). The signals were monitored using a CFX96 Real-Time System (Bio-Rad, Hercules, CA, USA). For each primer pair, the amplification efficiency was derived from a standard curve generated from five-fold serial dilution points of a mixture of the cDNA from all the samples. The expression stabilities of the reference genes were analysed using geNorm, BestKeeper and NormFinder software (Vandesompele et al., 2002; Andersen, Jensen & Orntoft, 2004; Pfaffl et al., 2004). All the software tools were performed according to their manuals. The average Cq value was calculated from three biological and three technical replicates. To normalize the differences in the amounts of mRNA from other genes, the amount of SpGAPDH mRNA was determined for each sample. The relative expression level of SpPDS and SpChlH were analysed using the 2−ΔΔCT method (Livak & Schmittgen, 2001). The error bars represent the ±SEs from three independent experiments. The data were analysed by ANOVA using SAS software.
| Gene symbol | Primer name | Primer sequence (5′–3′) | Product length (bp) | R2 | E% |
|---|---|---|---|---|---|
| ACTIN | Sp-YG-ACT-F Sp-YG-ACT-R |
ATTGAGCATGGCATTGTGAGC GCGATTAGCCTTTGGATTGAGA |
137 | 0.982 | 97.7 |
| GAPDH | Sp-YG-GAPDH-F Sp-YG-GAPDH-R |
CCAACCCTTGTCTTCCCACC CTCAAACCTACCGCCTCCCT |
242 | 0.990 | 103.8 |
| UBQ | Sp-YG-UBQ-F Sp-YG-UBQ-R |
TTGGCAAGCAACAATCAT GCAGATGGACAGCAGGAC |
225 | 0.986 | 105.7 |
| PDS | Sp-YG-PDS-F Sp-YG-PDS-R |
TCATGTTGTCAAAACTCCAAGG TGTCAACTTCTTCTCGCTCC |
223 | 0.995 | 96.7 |
| ChlH | Sp-YG-ChlH-F Sp-YG-ChlH-R |
AAGCACCTGGTAATCTGAACTCTG CATCGGGTCACCTTCGTATC |
112 | 0.991 | 97.1 |
To determine whether leaf photobleaching was induced by TRV2-SpPDS and whether the yellow-leaf phenotype was induced by TRV2-SpChlH in S. pseudocapsicum, TRV RNA1 and RNA2 were detected using the following two primer sets: (1) pTRV1-F/pTRV1-R and (2) pTRV2-F/pTRV2-R. The primers were designed based on the TRV1 RNA1 sequence (GenBank ID AF406990) and the TRV2 RNA2 sequence (GenBank ID 406991). The PCR procedures were as follows: (1) 95 °C for 5 min, (2) 35 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min, and (3) maintenance at 72 °C for 2 min.
Construction of the pTRV2-SpPDS and pTRV2-SpChlH vectors
To generate the pTRV2-SpPDS vector, the inserted SpPDS fragment was amplified using the PDS_insert_EcoRI-F and PDS_insert_EcoRI-R primers, and the pTRV2- SpChlH vector was generated by amplifying the inserted SpChlH fragment using the ChlH_insert_SmaI-F and ChlH_insert_SmaI-R primers. These primers were designed according to the manufacturer’s instructions for a one-step seamless cloning kit (Ju Hua Tai Ke, China). The pTRV2 vector was then digested with EcoRI and SmaI (Thermo Fisher Scientific Inc., USA). After gel extraction, the inserted SpPDS fragment containing the EcoRI site adapter and the inserted SpChlH fragment containing the SmaI site adapter were ligated into the EcoRI-digested TRV2 vector and the SmaI-digested TRV2 vector, respectively, using a one-step seamless cloning kit (Ju Hua Tai Ke, China), and the resulting vectors were then transformed into E. coli DH5a cells (TransGen, China). The presence of the PDS-containing and ChlH-containing inserts was confirmed by PCR using the TRV2_insert_yz-F and TRV2_insert_yz-R primers through detection of the corresponding pTRV2 multiple cloning sites (MCS). The expected size of the PCR product from the empty TRV2 vector was 338 bp, whereas a 909-bp product indicated that the PDS-containing fragment of interest was inserted into the TRV2 vector, and a 809-bp product indicated that the ChlH-containing fragment of interest was inserted into the TRV2 vector. The plasmid was further sequenced to verify the correct insertion of the fragment. Finally, the successfully constructed vector was transformed into Agrobacterium strain GV3101 or LBA4404 using the freeze-thaw method, as previously reported (Jyothishwaran et al., 2007).
Agrobacterium inoculation and optimization of TRV-based VIGS conditions in S. pseudocapsicum
For the VIGS assay, the Agrobacterium strains GV3101 and LBA4404 containing the TRV-VIGS vector were cultured as described by Tian et al. (2013) with slight modifications. The Agrobacterium strains GV3101 and LBA4404 were harvested, resuspended in infiltration media (20 g/L sucrose, 5 g/L MS, 10 mM MES, and 200 mM acetosyringone) and adjusted to an OD 600 of 1.0. A mixture of Agrobacterium cultures containing pTRV1 and pTRV2 (or its derivatives) was placed in the dark at room temperature for 3 h before inoculation. Seedlings, seeds and sprouts were then infiltrated with Agrobacterium cultures containing pTRV1 and pTRV2 (or pTRV2 derivatives) (1:1, OD600 = 1.0) using three Agrobacterium inoculation methods. The detailed methods are described below:
Leaf syringe-infiltration method: When a seedling had developed two cotyledons, the underside of the cotyledons was rubbed gently with a 10-µL tip and then infiltrated with Agrobacterium inocula using a 1-mL needleless syringe.
Sprout vacuum-infiltration method: Sprouts that were 0.5–1 cm in length were submerged into agro-inocula in a beaker within a desiccator, pulled by a vacuum pump until the pressure reached 0.07 kPa, and maintained for 1 min.
Seed vacuum-infiltration method: Seeds were treated using the same protocol described for the sprout vacuum-infiltration method.
In the temperature optimization experiment, seedlings were treated with Agrobacterium strain GV3101 containing TRV2-SpChlH using the leaf syringe-infiltration method and then grown at 18 °C, 25 °C, or 30 °C. The number of seedlings subjected to each treatment was 50, and each experiment was repeated three times.
Results
Cloning and sequence analysis of S. pseudocapsicum orthologs for the reference genes and the PDS and ChlH genes
The reference genes and PDS and ChlH gene fragments were amplified by homology-based cloning as previously reported (Zhong et al., 2014). The results showed that the following fragments were obtained: approximately 600-bp ACTIN and GAPDH fragments, 200-bp UBQ fragment, 600-bp PDS fragment and 500-bp ChlH fragment (Fig. 1). A BLASTX search (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastx&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome) of the sequence results was conducted, and the search results showed that the ACTIN, GAPDH, UBQ, PDS and ChlH amplification fragments were derived from the corresponding genes. To obtain the longer PDS sequences, 3′-rapid amplification of cDNA ends was performed, and an approximately 761-bp partial PDS fragment was obtained (Fig. 1). In addition, an approximately 1,151-bp partial PDS fragment was obtained by sequence assembly based on the two above-mentioned PDS fragments using DNAMAN software (Lynnon Biosoft). Alignment of the ACTIN, GAPDH, UBQ, PDS and ChlH fragments of C. annuum and S. pseudocapsicum showed that these gene fragments shared high similarity between the two species; the ACTIN fragment (GenBank accession number: MG825852) in S. pseudocapsicum showed 99% similarity with the ACTIN gene fragment in C. annuum, whereas the GAPDH (GenBank accession number: MG825856), UBQ (GenBank accession number: MG825855), PDS (GenBank accession number: MG825854) and ChlH (GenBank accession number: M825853) gene fragments exhibited 88%, 100%, 95% and 97% similarity, respectively (Table 2). These results showed that the orthologous ACTIN, GAPDH, UBQ, ChlH and PDS gene fragments were cloned successfully in S. pseudocapsicum.
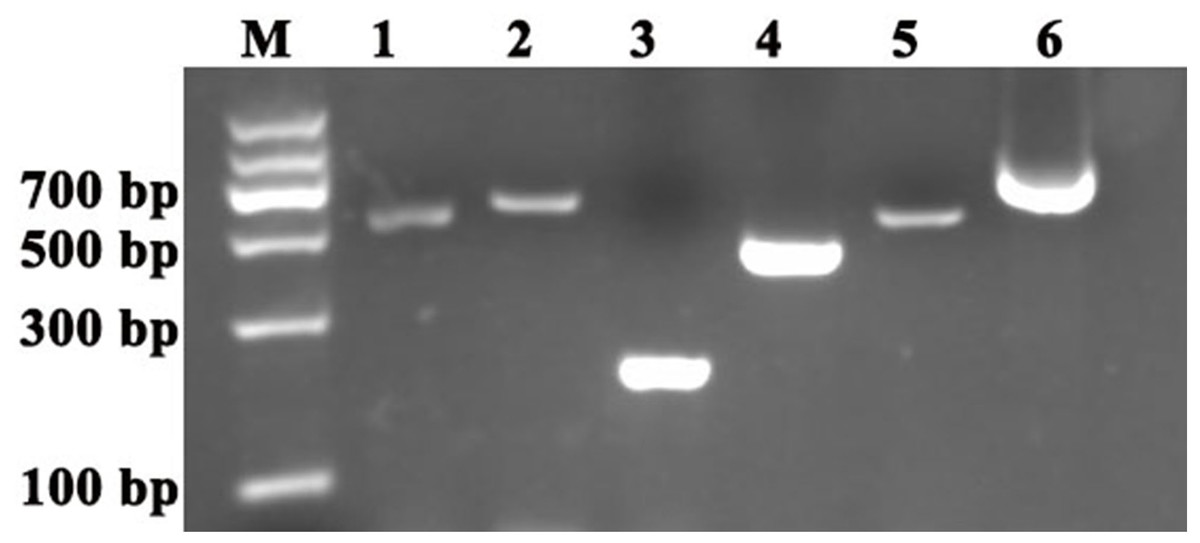
Figure 1: Cloning of the reference genes and PDS and ChlH gene fragments in S. pseudocapsicum.
Amplification of SpACTIN, SpGAPDH, SpUBQ, SpPDS and SPChlH from S. pseudocapsicum leaves. The primers for SpACTIN, SpGAPDH and SpUBQ were designed from conserved regions of ACTIN, GAPDH and UBQ based on the alignment of tomato, pepper, tobacco and potato CDS sequences. The primers for SpPDS and SPChlH were designed from conserved regions of PDS and ChlH based on the alignment of tomato, pepper, and tobacco CDS sequences. The cDNA from S. pseudocapsicum leaves was used as the template for the amplification of the corresponding PCR products. Lane 1, RT-PCR products of the ACTIN gene; Lane 2, RT-PCR product of the GAPDH gene; Lane 3, RT-PCR product of the UBQ gene; Lane 4, RT-PCR product of the ChlH gene; Lane 5, RT-PCR product of the PDS gene; and Lane 6, 3′RACE product of the PDS gene. The gene-specific primers (GSPs) for 3′RACE were designed based on the sequence of Lane 5 according to the manufacturer’s instructions provided with the GeneRacer™ kit (Invitrogen, Carlsbad, CA, USA). All the primers are shown in Table S1.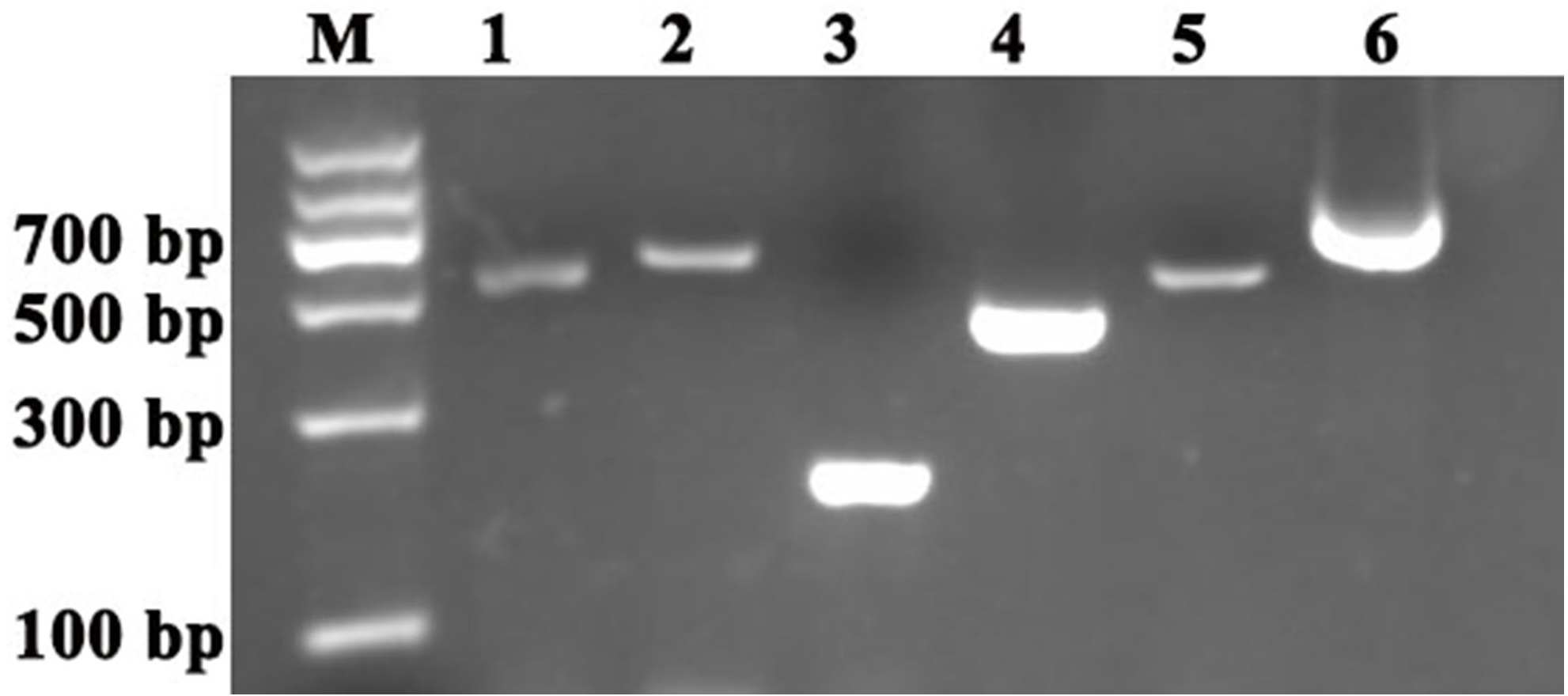{kind=link}
Assessment of primer specificity and expression levels of candidate reference genes
For identification of RT-qPCR primer specificity, the PCR-amplified products were analysed by agarose gel electrophoresis and melting curve analyses. The three sets of RT-qPCR primer pairs generated single bands with the expected size (Fig. 2A). Additionally, melting curve analyses also yielded a single peak with no visible primer-dimer formation (Figs. 2B–2D). Thus, both analyses confirmed that the three reference gene primer pairs yielded specific amplification of the reference genes in S. pseudocapsicum. The correlation coefficients (R2) of the standard curve were higher than 0.98 (Table 1), indicating good linear relationships among all the samples. The amplification efficiencies ranged from 97.7% for ACTIN to 105.7% for UBQ, suggesting that the reference gene primer pairs are suitable for further gene expression analyses (Table 1).
| Gene symbol | BlastX search | |||
|---|---|---|---|---|
| Capsicum orthologue locus | Capsicum locus description | Similarity (e-value) | Identity (%) | |
| ACTIN | XP_016543674.1 | Actin | 2e−139 | 99% |
| GAPDH | PHT69511.1 | Glyceraldehyde-3-phosphate dehydrogenase | 1e−123 | 88% |
| UBQ | AAR83898.1 | Ubiquitin-conjugating protein | 2e−47 | 100% |
| PDS | XP_016562403.1 | 15-cis-phytoene desaturase | 0.0 | 95% |
| ChlH | PHT80618.1 | Magnesium-chelatase subunit (ChlH) | 2e−101 | 97% |
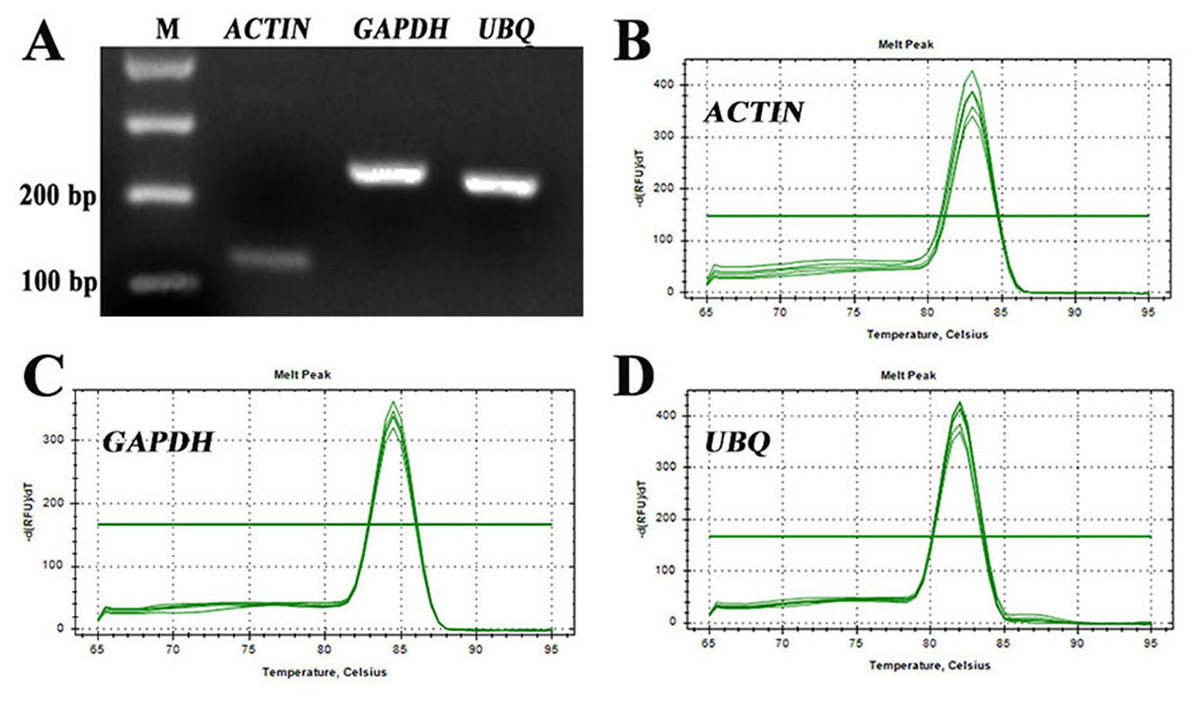
Figure 2: Assessment of the specificity of the primers used for RT-qPCR amplification.
(A) Agarose gel electrophoresis showing the specific RT-PCR products of the expected size obtained for the three candidate genes. (B–D) Melting curves of the ACTIN, GAPDH, and UBQ reference genes showing single peaks (each was obtained from five-fold serial dilutions of pooled cDNA samples from leaves of S. pseudocapsicum).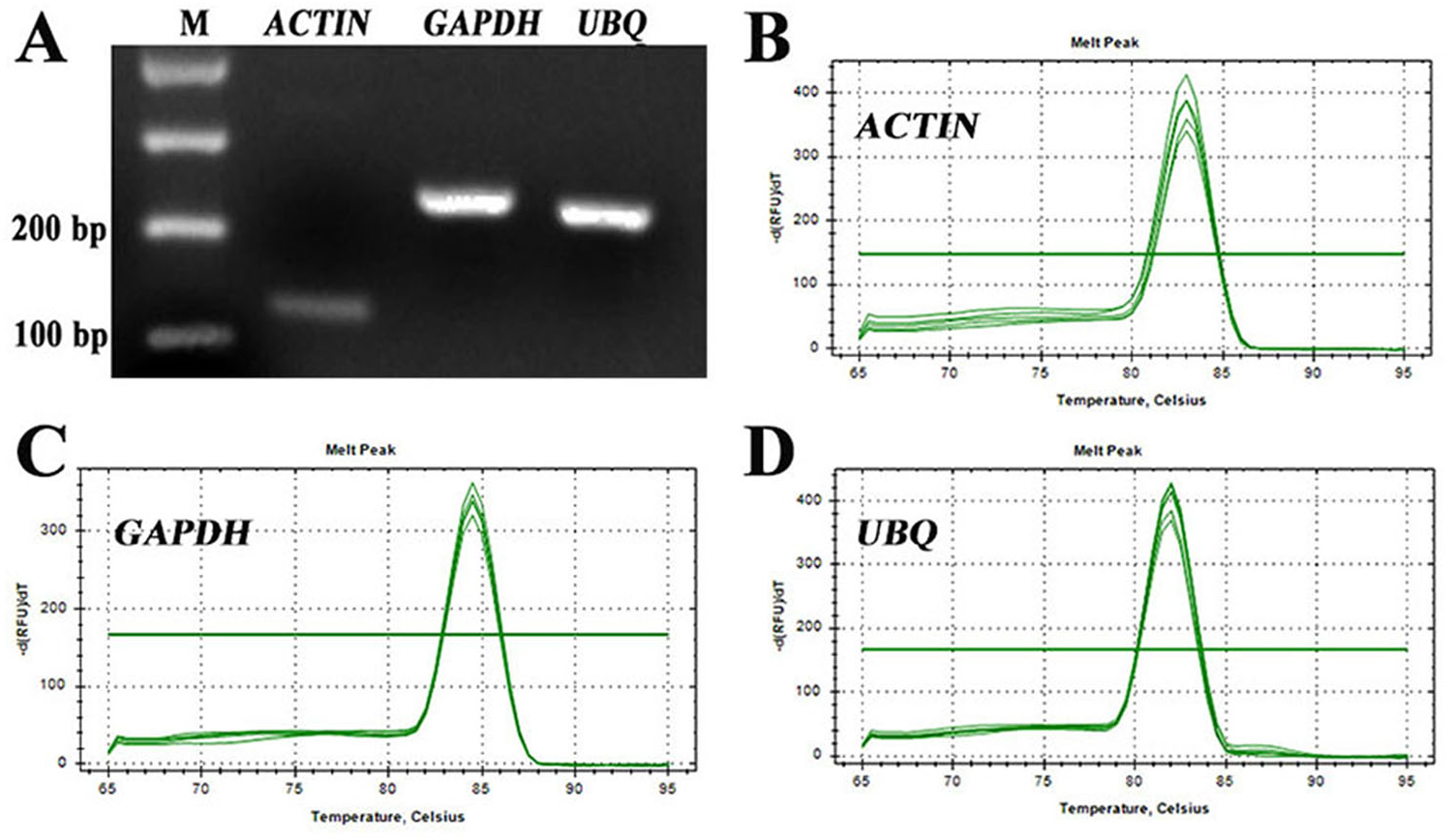{kind=link}
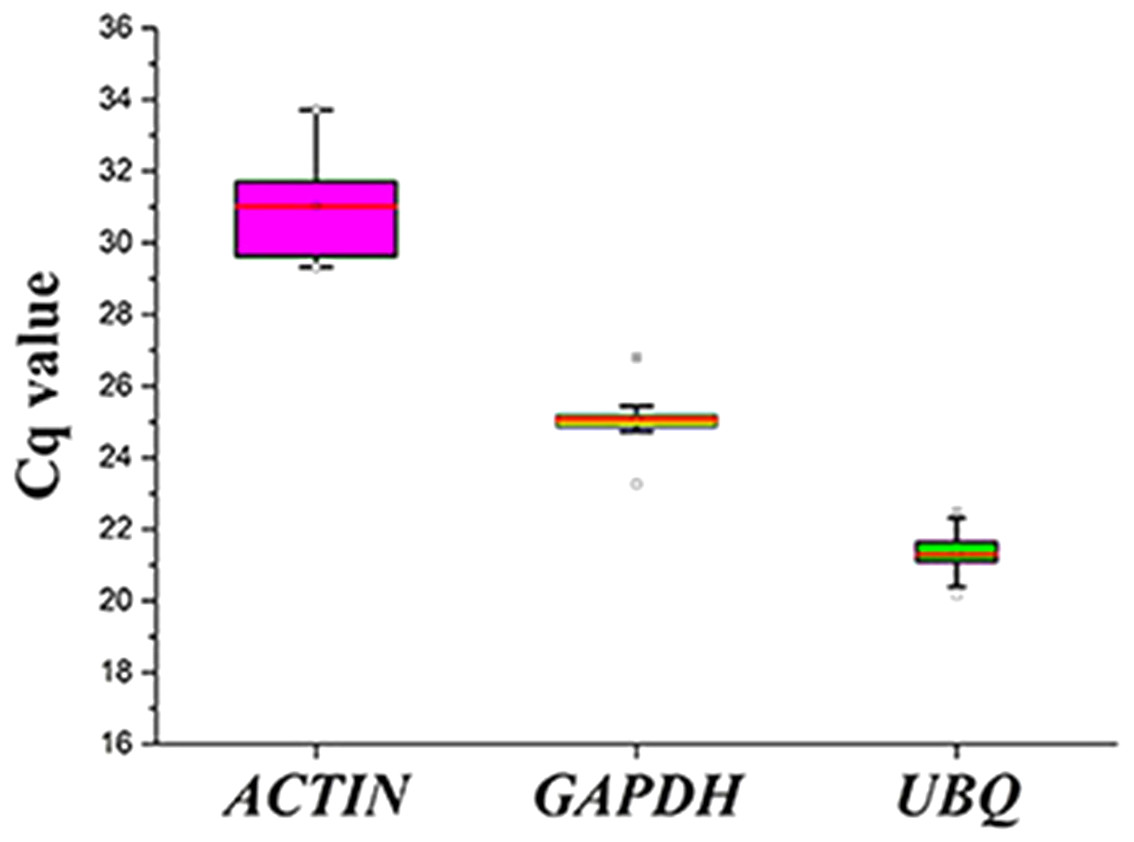
Figure 3: Cq values for the candidate reference genes in all the samples.
Expression levels of the three candidate reference genes in all the samples. The expression data are displayed as the Cq values for each reference gene in all the experimental samples (n = 15). A line across the box depicts the median values. The box indicates the 25th and 75th percentiles, and the whiskers represent the maximum and minimum values.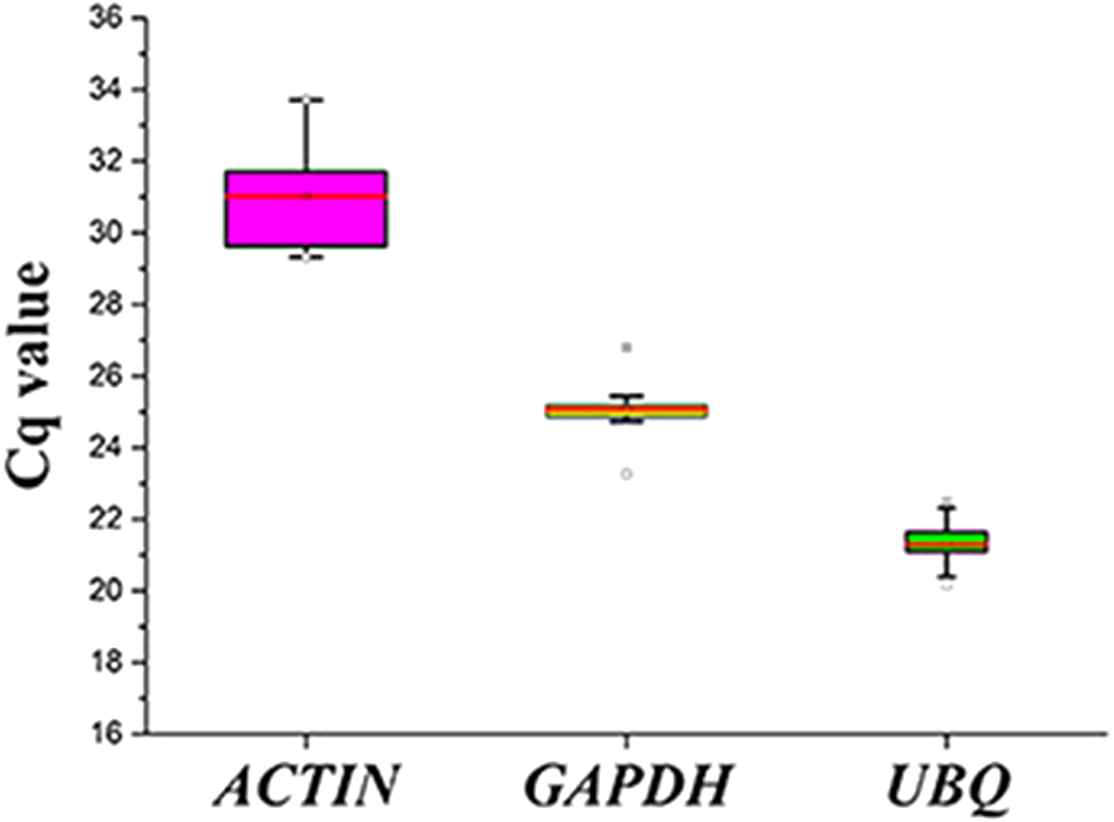{kind=link}
The expression levels of the three candidate reference genes are presented as raw Cq values. The results showed that the selected reference genes exhibited relatively wide expression abundance and variation. The mean Cq values for the reference genes ranged from 31 (SpACTIN) to 21.3 (SpUBQ) (Fig. 3). None of the candidate reference genes was consistently expressed in the tested samples. Thus, the results showed that screening for suitable reference genes is critical for the analysis of gene expression in S. pseudocapsicum.
Reference gene stability analysis
The stabilities of the reference genes were assessed using the geNorm, BestKeeper and NormFinder algorithms. The average expression stability value (M value) obtained using geNorm software was used to assess the gene expression stability. The M value of a suitable reference gene should be less than 1.5, and the gene with the lowest M value is considered to exhibit the greatest expression stability (Vandesompele et al., 2002). The results showed that the M values for the three candidate reference genes were below the geNorm threshold of 1.5. In fact, the lowest M value was 0.865 (for GAPDH), whereas the highest M value was 1.109 (for ACTIN). The ranking of the M values was GAPDH (0.865) < UBI (1.053) < ACTIN (1.109) (Fig. 4A), which indicated that GAPDH and ACTIN were the most and least stable reference genes, respectively.

Figure 4: Assessment of the expression stability of the candidate genes calculated using (A) geNorm and (B) NormFinder.
(A) The expression stability (M) of each reference gene was calculated by geNorm software using 15 samples (n = 15). A lower M value indicates more stable gene expression. (B) The stability values of the reference genes were also calculated with NormFinder using 15 samples (n = 15), and a lower stability value indicates more stable gene expression. Each experiment was repeated three times.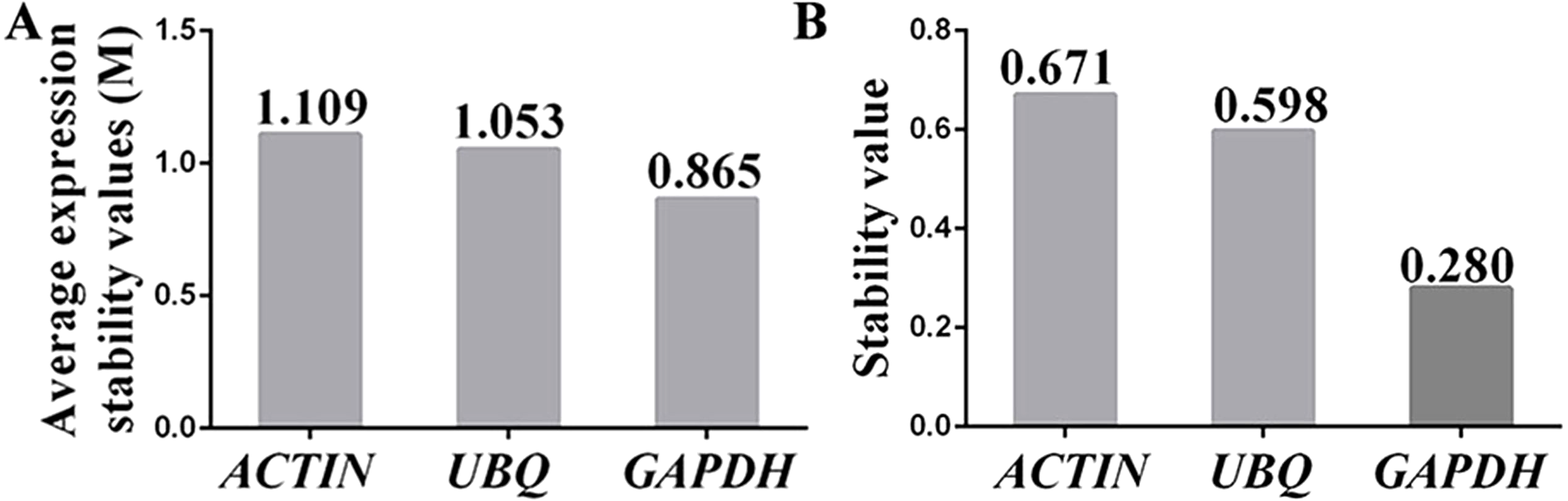{kind=link}
NormFinder software was used to further confirm the results obtained with the geNorm programme. Similar to geNorm software, the expression of the most stable gene was indicated by the lowest average expression stability value (Andersen, Jensen & Orntoft, 2004). The ranking of the stability values calculated using NormFinder was GAPDH (0.280) < UBQ (0.598) < ACTIN (0.671) (Fig. 4B). These results suggested that the genes showing the highest and lowest expression stability were GAPDH and ACTIN, respectively, which was in agreement with the results calculated using geNorm software.
BestKeeper software was also used to assess the expression stability of the reference candidate genes, and this assessment was performed by calculating the standard deviation (SD), the coefficient of variation (CV) and the Pearson correlation coefficient (r) from the raw Cq values (Pfaffl et al., 2004). Specifically, the lowest SD and CV values and the highest r value indicate the most stable reference gene. The ranking of the SD and CV values for the three reference candidates was ACTIN > GAPDH > UBQ (Table 3), indicating that ACTIN was the least stable reference gene. Although the SD and CV values for UBQ were lower than those for GAPDH, the r value for GAPDH (0.918) was higher than that obtained for UBQ (0.832). Thus, GAPDH was selected as the most suitable reference gene according to BestKeeper software.
| Gene name | Standard deviation (SD) | Coefficient of variation (CV) | Pearson correlation coefficient (r) |
|---|---|---|---|
| ACTIN | 1.11 | 3.59 | 0.985 |
| GAPDH | 0.59 | 2.34 | 0.918 |
| UBQ | 0.27 | 1.29 | 0.832 |
Based on the results obtained using geNorm, NormFinder and BestKeeper software, GAPDH was selected as the most stable reference gene for detecting the expression levels of PDS and ChlH in the following experiments.
Verification of the construction of the TRV2-SpPDS and TRV2-SpChlH vectors
To investigate the accuracy of the construction of TRV2-SpPDS and TRV2-SpChlH, PCR verification was performed using TRV2-SpPDS and TRV2-SpChlH as the template, respectively. A band with the expected size of approximately 909 bp was obtained in lane 2 (using TRV2-SpPDS as template), and a 809-bp band was obtained in lane 3 (using TRV2-SpChlH as template) (Fig. 5). These findings suggested that the PDS and ChlH fragments were introduced into the TRV2 vector, respectively. To further verify the results, we sequenced the TRV2-SpPDS and TRV2-SpChlH plasmids. Alignment of the sequencing results of TRV2-SpPDS and the inserted PDS fragment showed that the two sequences shared 100% similarity (Fig. S1), and the sequencing results of TRV2-SpChlH and the inserted ChlH fragment were similar to those obtained for TRV2-SpPDS (Fig. S2). These results demonstrate that the TRV2-SpPDS and TRV2-SpChlH vectors were accurately constructed.
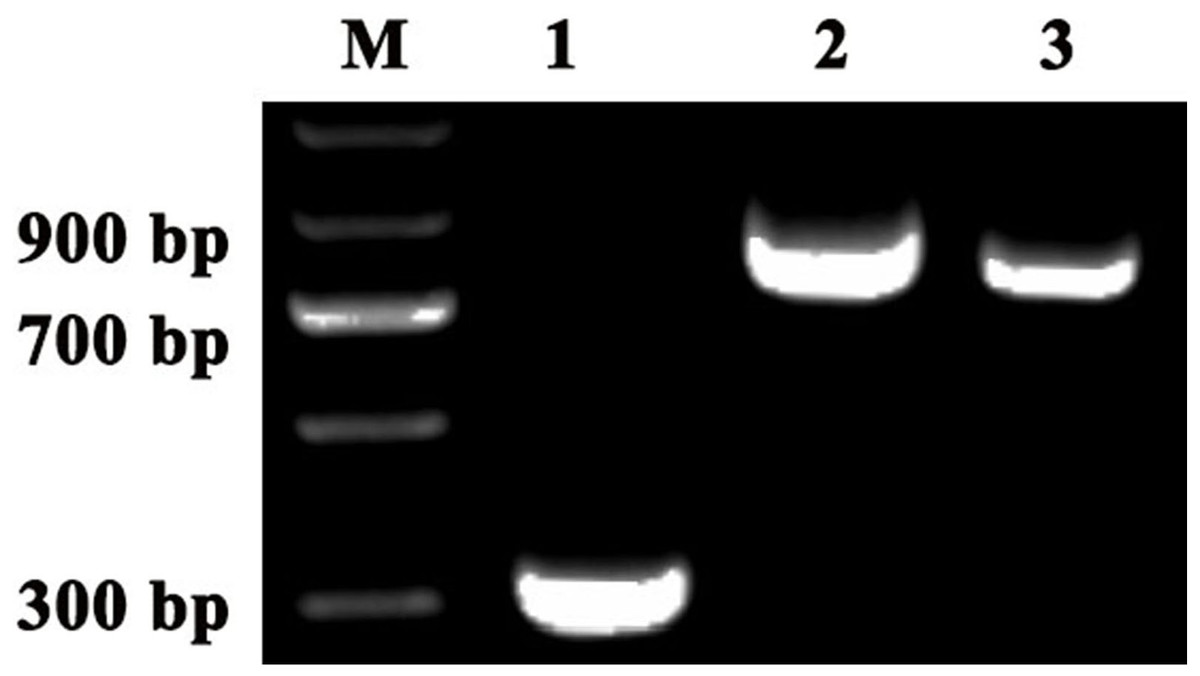
Figure 5: Detection of TRV2 and the TRV2-SpPDS and TRV2-SpChlH plasmids.
RT-PCR was performed with locus-specific primers to detect the multiple cloning sites (MCS) using different vectors as the template. Agarose gel electrophoresis showed specific RT-PCR products of the expected size for the different vectors. Lane M, marker; Lane 1, TRV2; Lane 2, TRV2-SpPDS vector; and Lane 3, TRV2-SpChlH vector. An approximately 338-bp band (Lane 1) indicated the empty TRV2 vector, whereas a 909-bp band (Lane 2) indicated that the TRV2-SpPDS vector was successfully constructed, and a 809-bp band (Lane 3) indicated that the TRV2-SpChlH vector was also successfully constructed.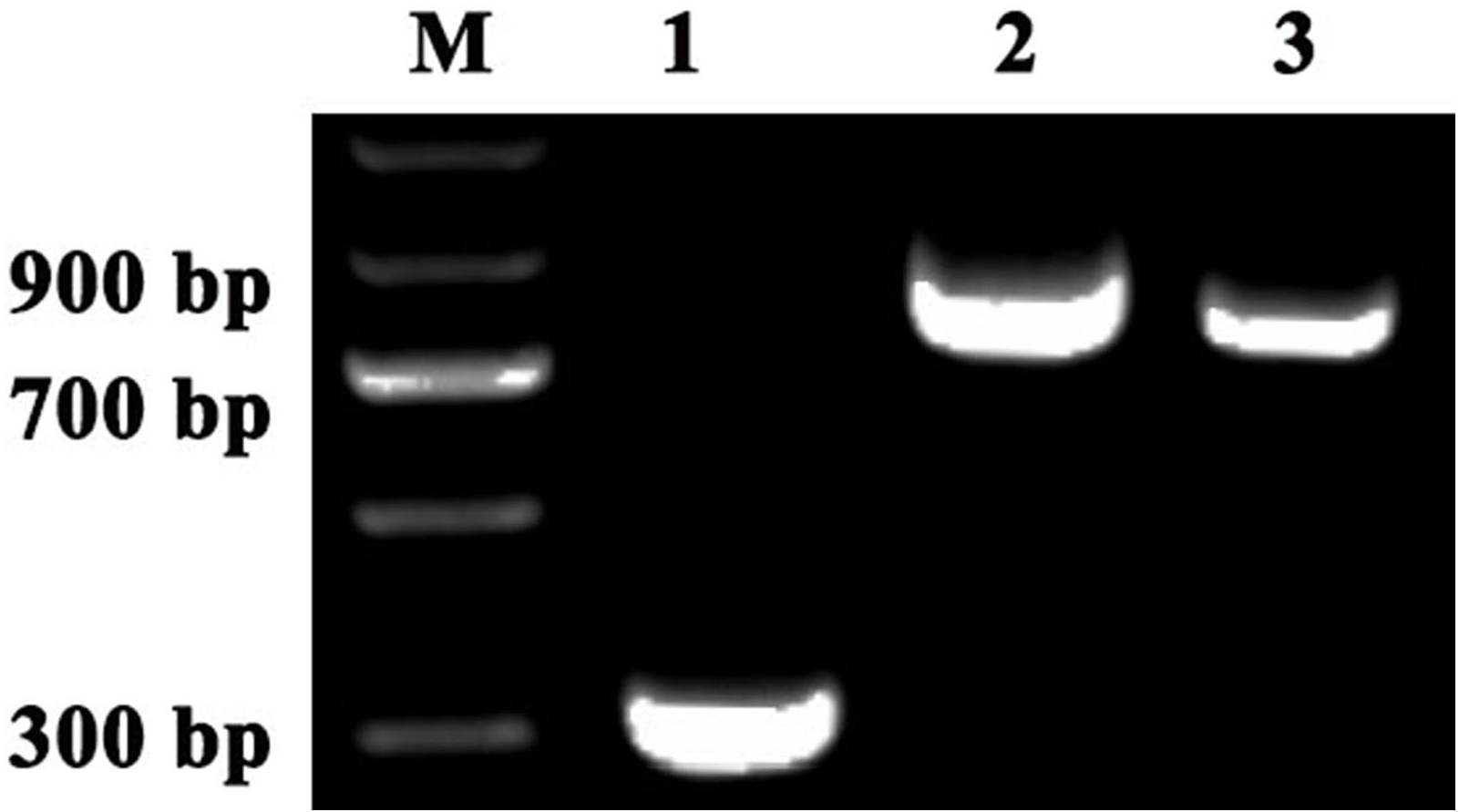{kind=link}
Silencing of the PDS and ChlH genes in S. pseudocapsicum seedlings
To investigate whether the TRV2-SpPDS and TRV2-SpChlH vectors could silence the corresponding endogenous PDS and ChlH genes in S. pseudocapsicum, the TRV2-SpPDS and TRV2-ChlH vectors were transformed into Agrobacterium strain GV3101, and the resulting strains were then inoculated into cotyledons using the leaf syringe-infiltration method. All the treated seedlings survived, indicating that the method was suitable for the tested seedlings. At 15 days post-infiltration (dpi), the photobleaching phenomenon was observed in the newly developed leaves obtained from the seedlings treated with TRV2-SpPDS but not in those obtained from the control and mock-treated seedlings (Fig. 6C, Table 4). Meanwhile, the yellow-leaf phenotype was detected in the newly developed leaves collected at 14 dpi from the seedlings treated with TRV2-SpChlH (Fig. 6K, Table 4). Moreover, the next newly developed leaves in the seedlings infiltrated with the TRV2-SpPDS and TRV2-SpChlH vectors also exhibited the photobleaching and yellow-leaf phenotypes, respectively (Figs. 6G and 6O). Additionally, we found that the silencing phenotypes obtained with the TRV2-SpPDS and TRV2-SpChlH vectors occurred along the leaf vein at the early silencing stage (Figs. 6D, 6H, 6L and 6P).

Figure 6: Establishment of a TRV-mediated VIGS protocol for S. pseudocapsicum using the leaf syringe-infiltration method.
All the TRV2 vectors were transformed into Agrobacterium strain GV3101 prior to its use for the treatment of S. pseudocapsicum seedlings. (A, E, I and M) CK, untreated S. pseudocapsicum seedlings. (B, F, J and N) TRV2 (mock), S. pseudocapsicum seedlings treated with TRV2. (C and G) Silenced S. pseudocapsicum seedlings treated with TRV2-SpPDS. The photobleached leaf phenotype observed in the PDS-silenced seedlings is shown in the first (C) and second (G) newly developed leaves. (K and O) Silenced S. pseudocapsicum seedlings treated with TRV2-SpChlH. The yellow-leaf phenotype observed in the ChlH-silenced seedlings is shown in the first (K) and second (O) newly developed leaves. (D, H, L and P) Silenced leaflets showing signs of the silencing phenotype along the vascular system. (D and H) Photobleached leaves of PDS-silenced seedlings. (L and P) Yellow leaves of ChlH-silenced seedlings. Photo credit: Hua Xu.{kind=link}
| Agro-inoculation method | PDS | ChlH | ||||
|---|---|---|---|---|---|---|
| NTS | NSP | DFI | NTS | NSP | DFI | |
| Seed vacuum-infiltration | 50 | 0 | – | 50 | 0 | – |
| Leaf syringe-infiltration | 50 | 25 | 15 | 50 | 26 | 14 |
| Sprout vacuum-infiltration | 50 | 4 | 13 | 50 | 5 | 12 |
Notes:
- NTS
-
number of treated seedlings
- NSP
-
number of seedlings showing a silenced phenotype
- DFI
-
first day after infiltration that the phenotype appeared
To investigate whether the photobleaching and yellow-leaf phenotypes were induced by the silencing of the corresponding endogenous PDS and ChlH genes, the PDS and ChlH expression levels in various VIGS-treated seedlings were detected by RT-qPCR using GAPDH as the reference gene. The GAPDH band was detected in all the samples, indicating that the cDNA from all the samples was of sufficient quality and could be used in a subsequent experiment (Fig. 7A). The RT-qPCR results showed that the PDS and ChlH expression levels were significantly reduced in the silenced seedling leaves compared with the leaves of the control and mock-treated seedlings. Moreover, the PDS and ChlH expression levels in the mock-infected and control leaves were similar (Figs. 7C, 7D). The above results indicated that the photobleaching and yellow-leaf phenotypes were initiated by PDS and ChlH gene silencing, respectively. Additionally, to determine whether the observed PDS and ChlH gene silencing was due to the presence of the TRV viral vector, the TRV1 and TRV2 fragments in the infected and uninfected seedlings were assessed, and both the infected and the mock-inoculated seedlings showed the presence of the TRV1 and TRV2 bands (Fig. 7B), suggesting that silencing of the PDS and ChlH genes was induced by the presence of the TRV viral vector. The above-mentioned results indicated that TRV2-SpPDS could induce photobleaching by silencing of the endogenous PDS gene and that TRV2-SpChlH could induce the yellow-leaf phenotype by silencing of the endogenous ChlH gene in S. pseudocapsicum.
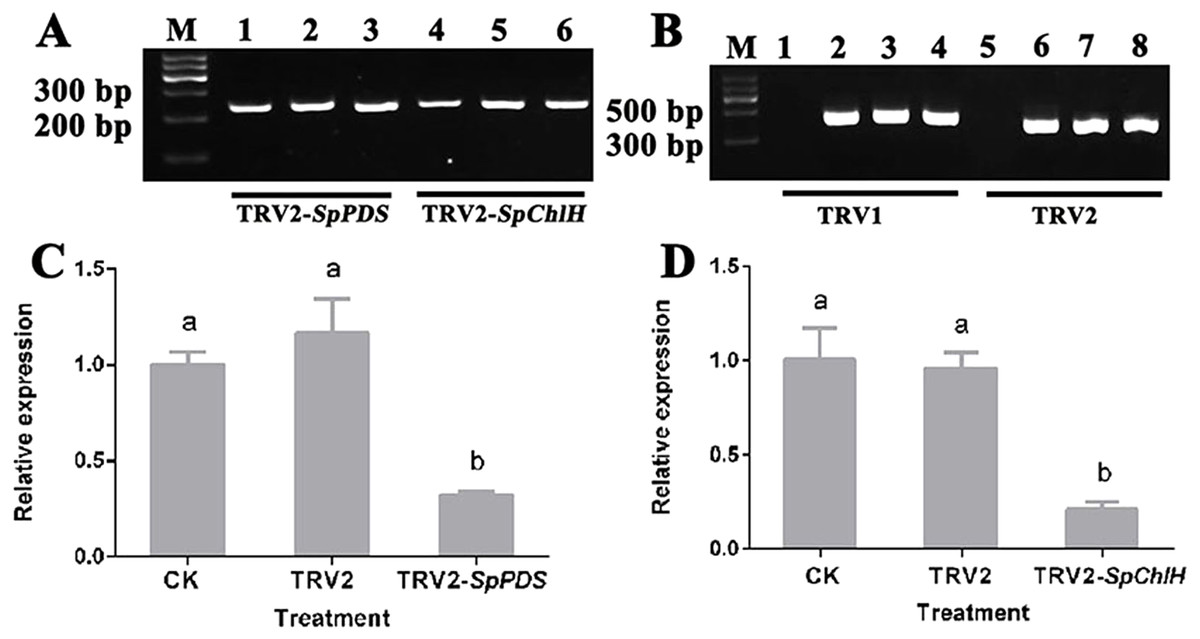
Figure 7: Detection of the SpPDS and SpChlH expression levels and TRV transcripts after VIGS.
(A) RT-PCR detection of the SpGAPDH expression levels in leaves after various treatments. Lane M, marker; Lanes 1 and 4, untreated; Lanes 2 and 5, treated with TRV2; Lane 3, treated with TRV2-SpPDS; and Lane 6, treated with TRV2-SpChlH. (B) RT-PCR detection of TRV transcripts in leaves after various treatments. Lane M, marker; Lanes 1 and 5, untreated; lanes 2 and 6, treated with TRV2; Lanes 3 and 7, treated with TRV2-SpPDS; Lanes 4 and 8, treated with TRV2-SpChlH; Lanes 1, 2, 3 and 4, detection of TRV1; and Lanes 5, 6, 7 and 8, detection of TRV2. The second newly developed leaves from untreated and mock-treated S. pseudocapsicum seedlings were compared with leaves from TRV2-SpPDS or TRV2-SpChlH-treated seedlings. RT-PCR was performed with locus-specific primers for the reference gene SpGAPDH and the viral transcripts TRV1/TRV2. (C–D) SpPDS (C) and SpChlH (D) expression levels in the leaves after VIGS treatments. The expression levels of SpPDS and SpChlH were quantified via real-time RT-PCR in samples from leaves exhibiting the silenced phenotype and CK and mock-treated leaves at the two-true-leaf stage. SpGAPDH was used as an internal control for normalization of the PCR efficiency, and the SpPDS and SpChlH expression values of the CK were set to 1. The error bars represent the ±SE of three independent experiments. Different letters indicate significant differences at P ≤ 0.05.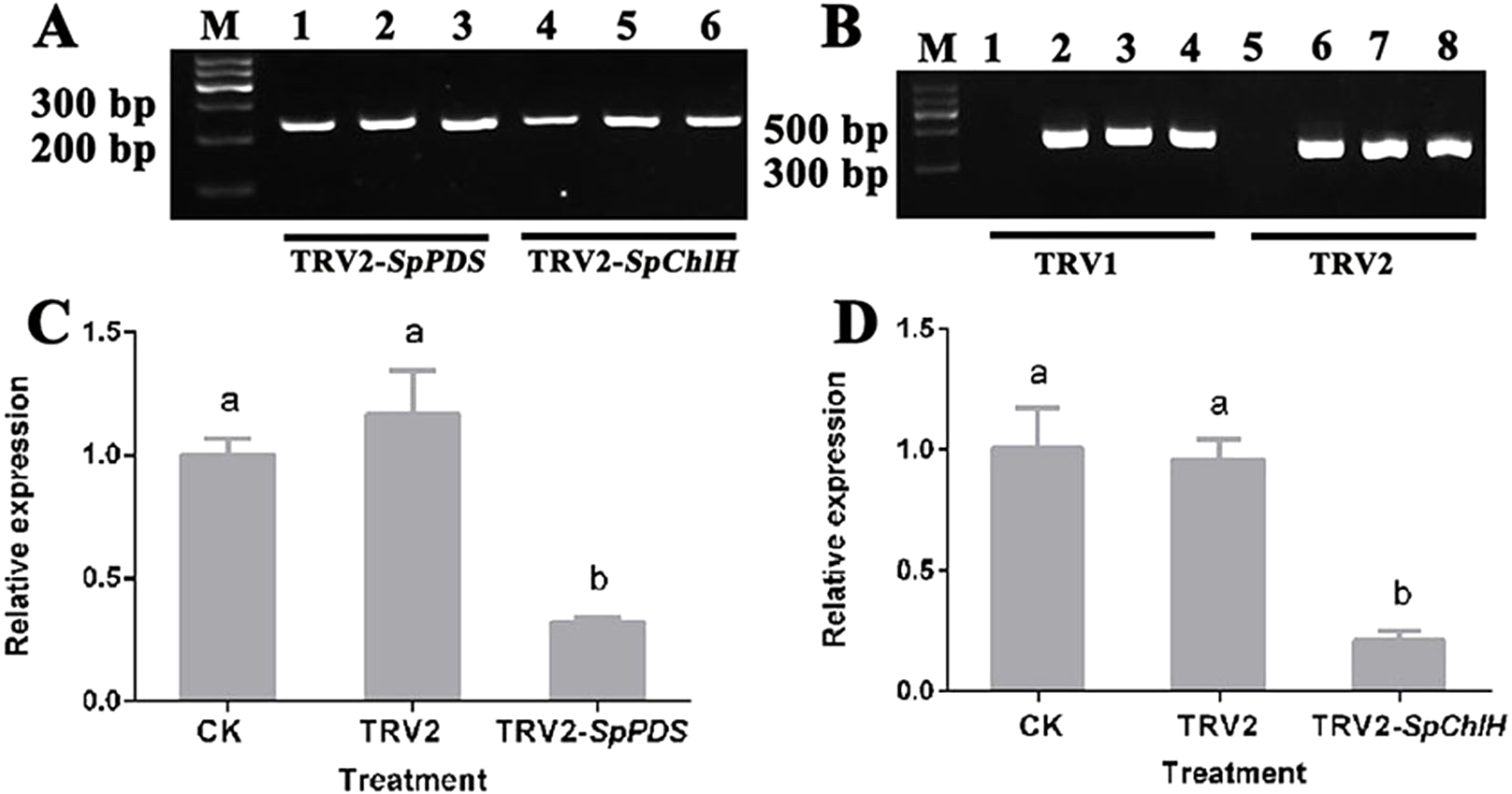{kind=link}
Effect of various infiltration methods on silencing efficiency
To test whether the infiltration method could affect the silencing efficiency of the VIGS system in S. pseudocapsicum, the effects of various inoculation methods on the silencing efficiency were compared. The seed vacuum-infiltration method failed to induce any silencing phenotypes (Table 4), and no TRV transcript was detected in the leaves of S. pseudocapsicum (data not shown). The sprout vacuum-infiltration method could induce PDS and ChlH gene silencing with a silencing efficiency of approximately 10% at 12–13 dpi, which is relatively low. In contrast, using the leaf syringe-infiltration method, the infected phenotypes were first detected at 14–15 dpi, and the TRV2-SpPDS and TRV2-SpChlH vectors induced the respective phenotypes in approximately 50% of treated seedlings (Table 4). The results indicated that the leaf syringe-infiltration method was the most effective for inducing the silencing phenotype in S. pseudocapsicum.
Effect of the Agrobacterium strain on the silencing efficiency
To investigate whether the Agrobacterium strains used to introduce the VIGS vectors affected the silencing efficiency, two Agrobacterium strains, GV3101 and LBA4404, were used to introduce the TRV2-SpPDS and TRV2-SpChlH vectors, respectively. The results showed that both Agrobacterium strains could introduce the VIGS vectors used for silencing of the gene of interest, but the silencing efficiency of PDS and ChlH induced by GV3101 was higher than that induced by LBA4404, regardless of whether the leaf syringe-infiltration or the sprout vacuum-infiltration method was used (Table 5). The results indicated that GV3101 was better for the introduction of the VIGS vectors into S. pseudocapsicum than LBA4404.
| Agrobacterium strain | Agro-inoculation method | Silencing efficiency | |
|---|---|---|---|
| PDS | ChlH | ||
| GV3101 | Leaf syringe-infiltration | 50% | 52% |
| Sprout vacuum-infiltration | 8% | 10% | |
| LBA4404 | Leaf syringe-infiltration | 44% | 40% |
| Sprout vacuum-infiltration | 4% | 6% | |
Effect of the growth temperature of agro-inoculated plants on the silencing efficiency
To investigate whether the growth temperature of the agro-inoculated plants affected the silencing efficiency, we compared the ChlH silencing efficiency under different growth temperatures (18 °C, 25 °C and 30 °C) after agro-inoculation using the leaf syringe-infiltration method. Analysis of the agro-inoculated seedlings showed that 52% of the seedlings that grew at 25 °C developed the yellow-leaf phenotype at 14 dpi, 12% of the seedlings that grew at 18 °C developed a yellow-leaf phenotype at 22 dpi, and 4% of the seedlings that grew at 30 °C developed the silencing phenotype at 28 dpi (Table 6). Thus, both higher and lower growth temperatures decreased the silencing efficiency and delayed the time from infection to obvious phenotype detection. The results indicated that the speed of the response and the efficiency of infection were optimal at 25 °C.
| Growth temperature (°C) | % of plants with the yellow-leaf phenotype | Days to phenotype detection |
|---|---|---|
| 18 | 12 | 22 |
| 25 | 52 | 14 |
| 30 | 4 | 28 |
Discussion
Gene function studies promote increased understanding of the molecular mechanisms in plants, and to date, few gene function studies have been performed in S. pseudocapsicum. VIGS is a recently developed powerful genetic tool for characterizing the function of plant genes (Burch-Smith et al., 2004). PDS and ChlH have been commonly used as indicator genes in VIGS systems because their resulting silencing phenotypes can be easily scored (Cunningham & Gantt, 1998; Hiriart et al., 2002; Liu et al., 2012). In the present study, we established that TRV-based VIGS could be applied to unravel the functions of the PDS and ChlH genes in S. pseudocapsicum. Systemic viral infection was essential for the VIGS system. Our results also suggested that newly developed leaves obtained after VIGS application exhibited the silencing phenotype, indicating establishment of systemic TRV viral infection in S. pseudocapsicum (Figs. 6C, 6G, 6K and 6O). Additionally, the results showed that both silencing phenotypes occurred along the leaf vein (Figs. 6D, 6H, 6L and 6P), which agrees with the results that viral propagation and the silenced systemic response occurred mainly along the vascular bundle system (Wege et al., 2007). To the best of our knowledge, this study provides the first demonstration of a successful application of TRV-based VIGS for the identification of gene function in S. pseudocapsicum.
We also compared several parameters that are likely to affect the silencing efficiency in S. pseudocapsicum, including the Agrobacterium inoculation method, the Agrobacterium strain and the growth temperature after Agrobacterium infiltration. In some previous studies, the vacuum-infiltration method was found to be more effective than other infiltration methods (Deng et al., 2012; Liu et al., 2014). However, our results suggested that the leaf syringe-infiltration method is more effective than vacuum-infiltration in S. pseudocapsicum (Table 4). This difference might have been observed due to the high hardness of the seed coat of S. pseudocapsicum, which might have not allowed infection with the virus vector.
It is well known that susceptibility to infection by Agrobacterium varies among different plant species and cultivars. For example, in Arabidopsis, agro-inoculation with LBA4404 does not give rise to a silencing phenotype, whereas the GV3101 strain results in highly efficient VIGS (Cai et al., 2006). However, in Gossypium barbadense, agro-infiltration with the GV3101 and LBA4404 strains resulted in efficient VIGS (Pang et al., 2013). Here, we evaluated the silencing effect of two strains of Agrobacterium, GV3101 and LBA4404, in S. pseudocapsicum. The results showed that both GV3101 and LBA4404 could introduce the VIGS vector to produce the silencing phenotype (Table 5), which agrees with the results obtained in G. barbadense (Pang et al., 2013). Pang et al. (2013) found that the two Agrobacterium strains GV3101 and LBA4404 yielded no significant difference in the VIGS efficiency (Pang et al., 2013), but in this study, the silencing efficiency in S. pseudocapsicum obtained with GV3101 was slightly higher than that obtained by LBA4404, which might be due to the differences between the species.
The growth temperature conditions after inoculation have a profound effect on silencing efficiency (Burch-Smith et al., 2004). Different plant species require different temperatures for producing a good silencing phenotype after VIGS. For example, in tomato, the optimal silencing phenotype is obtained with a growth temperature of 22 °C after TRV-based VIGS (Jiang et al., 2008), whereas temperatures of approximately 25 °C are desirable for N. benthamiana (Burch-Smith et al., 2004). In the present study, we also found that the growth temperature after inoculation affected the silencing efficiency. A decreased silencing efficiency was obtained when the seedlings grew at lower (18 °C) or higher (30 °C) temperatures, and the most suitable growth temperature for obtaining a high silencing efficiency in S. pseudocapsicum was 25 °C (Table 6).
The growth stage of the agro-inoculated plants can also affect the gene silencing efficiency (Burch-Smith et al., 2004; Deng et al., 2012). In addition to at the cotyledon stage, we also inoculated the seedlings at the two-true-leaf and four-true-leaf stages with TRV2-SpPDS using the leaf syringe-infiltration method, which was predicted to initiate the photobleaching phenotype in leaves. Unfortunately, the SpPDS silencing phenomenon was not observed in the seedlings’ leaves (data not shown), indicating that the optimal VIGS inoculation stage in S. pseudocapsicum was the cotyledon stage.
RT-qPCR is an important technique for assessing gene expression levels. The reliability of RT-qPCR results is highly dependent on the suitability of the reference gene. It is thus necessary to screen for a suitable reference gene prior to the assessment of gene expression levels. To date, suitable reference genes for S. pseudocapsicum have not been well characterized. Thus, to accurately assess the gene expression levels in seedlings in which a silencing phenotype was induced with TRV2-SpPDS and TRV2-SpChlH, we screened for the most stable reference gene in this study. We compared three potential reference genes (ACTIN, GAPDH, and UBQ) for S. pseudocapsicum across five samples collected from leaves at different developmental stages and various tissues. The GAPDH gene was identified as the most stable reference gene for assessing gene expression levels in this study. To the best of our knowledge, this study constitutes the first screening for a stable reference gene in S. pseudocapsicum and lays the foundation for the characterization of gene expression levels in S. pseudocapsicum. Previous studies have shown that the expression levels of the reference genes varied under different experimental conditions (Sinha et al., 2015; Li et al., 2016; Sudhakar Reddy et al., 2016). Thus, the most suitable reference gene, which was identified as GAPDH in this study, should be subjected to further screening in S pseudocapsicum under other experiment conditions.
The rankings of the three reference genes obtained in the present study showed some discrepancies. For example, GAPDH was identified as the most stable reference gene using geNorm and NormFinder software, but the ranking order obtained for UBQ was higher than that found for GAPDH using BestKeeper. This difference might be due to the different statistical models used in the various algorithms. Differences in the ranking orders obtained with these algorithms have also been observed in other studies (Tong et al., 2009; Reddy et al., 2016; Xu et al., 2017).
Conclusions
In summary, we established a TRV-based VIGS system that can successfully induce photobleaching and yellow-leaf phenotypes by silencing SpPDS and SpChlH, respectively. Using the optimal parameters, including Agrobacterium strain GV3101, the leaf syringe-infiltration method, and a growth temperature for the agro-inoculated plants of 25 °C, the silencing efficiency can reach approximately 50%. This established system will facilitate the future characterization of gene functions in S. pseudocapsicum. Additionally, the present study constitutes the first cloning and screening of reference genes and thus lays the foundation for gene expression analysis in S. pseudocapsicum. These results provide a better understanding of the molecular and physiological mechanisms that regulate S. pseudocapsicum and its associated traits.
Supplemental Information
Supplementary materials
Fig. S1. Alignment of the sequencing results of TRV2-SpPDS and the inserted PDS fragments. C–A, sequence of TRV2-SpChl H. C-B, sequence of insert ChlH fragment.
Fig. S2. Alignment of the sequencing results of TRV2-SpChlH and the inserted ChlH fragments. P-A, sequence of TRV2-SpPDS. P-B, sequence of insert PDS fragment.
Table S1. Primers used in this study.
Raw data for box plot
Cq value of box plot for evaluating expression levels of the three candidate reference genes in all the samples.
Raw data for reference genes screening
Cq value to assess the expression stability of the candidate genes using three softwares (geNorm, NormFinder and BestKeeper).
Raw data for TRV2-ChlH-induced gene silencing
Raw data to detect the SpChlH expression levels in VIGS experiment by RT-qPCR.
Raw data for TRV2-PDS-induced gene silencing
Raw data to detect the SpPDS expression levels in VIGS experiment by RT-qPCR.