
Leishmania amazonensis promastigotes in 3D Collagen I culture: an in vitro physiological environment for the study of extracellular matrix and host cell interactions
- Published
- Accepted
- Received
- Academic Editor
- Barbara Chan
- Subject Areas
- Cell Biology, Parasitology
- Keywords
- Leishmania , Extracellular matrix, 3D migration, Collagen matrix, Co-culture, Protease, Matrix remodelling, Parasite, Leishmania–macrophage interaction
- Copyright
- © 2014 Petropolis et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
- Cite this article
- 2014. Leishmania amazonensis promastigotes in 3D Collagen I culture: an in vitro physiological environment for the study of extracellular matrix and host cell interactions. PeerJ 2:e317 https://doi.org/10.7717/peerj.317
Abstract
Leishmania amazonensis is the causative agent of American cutaneous leishmaniasis, an important neglected tropical disease. Once Leishmania amazonensis is inoculated into the human host, promastigotes are exposed to the extracellular matrix (ECM) of the dermis. However, little is known about the interaction between the ECM and Leishmania promastigotes. In this study we established L. amazonensis promastigote culture in a three-dimensional (3D) environment mainly composed of Collagen I (COL I). This 3D culture recreates in vitro some aspects of the human host infection site, enabling the study of the interaction mechanisms of L. amazonensis with the host ECM. Promastigotes exhibited “freeze and run” migration in the 3D COL I matrix, which is completely different from the conventional in vitro swimming mode of migration. Moreover, L. amazonensis promastigotes were able to invade, migrate inside, and remodel the 3D COL I matrix. Promastigote trans-matrix invasion and the freeze and run migration mode were also observed when macrophages were present in the matrix. At least two classes of proteases, metallo- and cysteine proteases, are involved in the 3D COL I matrix degradation caused by Leishmania. Treatment with a mixture of protease inhibitors significantly reduced promastigote invasion and migration through this matrix. Together our results demonstrate that L. amazonensis promastigotes release proteases and actively remodel their 3D environment, facilitating their migration. This raises the possibility that promastigotes actively interact with their 3D environment during the search for their cellular “home”—macrophages. Supporting this hypothesis, promastigotes migrated faster than macrophages in a novel 3D co-culture model.
Introduction
The promastigote form of Leishmania amazonensis, causative agent of American cutaneous leishmaniasis (Barral et al., 1991; Murray et al., 2005), is transmitted through the bite of infected sand flies from the genus Lutzomyia (Kaye & Scott, 2011; Neuber, 2008). Once inoculated, the promastigotes are exposed to the dermis microenvironment, which is composed of extracellular matrix (ECM) proteins organized in a fibrilar network (Rogers, 2012). Because L. amazonensis is an intracellular parasite that only proliferates inside a cellular host in the mammalian host, its interaction with the host ECM has been neglected. However, in order to establish an intracellular infection, the promastigote form must overcome the obstacles presented by the dermis ECM (Chang & McGwire, 2002). Except for broad ECM alterations during experimental Leishmania infection (Abreu-Silva et al., 2004; Melo et al., 2009; Silva-Almeida et al., 2012a) little is known about the interaction of L. amazonensis with the host ECM (Kulkarni et al., 2008; McGwire, Chang & Engman, 2003). Although long viewed only as a supportive structure, the ECM is an essential part of the cell’s milieu that, through direct or indirect means, regulates almost all cellular behavior (Hynes, 2009), including inflammatory signaling (Larsen et al., 2006).
The glycoprotein gp63 present on parasite surfaces is a broad-spectrum zinc-dependent metalloprotease that works as an important virulence factor during Leishmania infection (McMaster et al., 1994; Silva-Almeida et al., 2012a; Yao et al., 2002). gp63 can degrade ECM components such as fibronectin and collagen IV and seems to enhance L. amazonensis migration in Matrigel, a commercial 3D gelatin of basement membrane ECM proteins (Kulkarni et al., 2008; McGwire, Chang & Engman, 2003). However, collagen I (COL I) is the main component of the dermis ECM, and for the moment little is known about the interaction of L. amazonensis with COL I rich environments (Lira, Rosales-Encina & Arguello, 1997).
Here, we introduce a new model for culture of L. amazonensis promastigotes in an in vitro 3D COL I matrix, which represents a more physiological in vitro culture because it better mimics many extracellular aspects of the Leishmania environment in the human host. The objective of this work was to characterize L. amazonensis invasion, migration, and matrix remodeling in 3D COL I matrices, focusing on the functions of Leishmania proteases. The establishment of this model enables evaluation of the mechanisms of Leishmania interaction with host cells in a 3D environment.
Methods
Ethics statement
The use of animal models was approved by the Ethics Committee for Animal Experimentation of the Health Sciences Centre, Federal University of Rio de Janeiro (Protocols n. IBCCF 096/097), according to the Brazilian federal law. All animals received humane care in compliance with the “Principles of Laboratory Animal Care” formulated by the National Society for Medical Research and the “Guide for the Care and Use of Laboratory Animals” prepared by the National Academy of Sciences, USA.
Leishmania amazonensis promastigotes culture
The MHOM/BR/75/Josefa strain of L. amazonensis used in this study is an anonymized strain that was isolated in 1975 from a patient with diffuse cutaneous leishmaniasis by Dr. Cesar A. Cuba-Cuba (Brasilia University, Brazil) and kindly provided by the Leishmania Collection of the Instituto Oswaldo Cruz (Code IOCL 0071–FIOCRUZ). It has been maintained by BALB/c footpad inoculation. Amastigote forms were obtained from these mice, and transformed into promastigotes that were axenically cultured in Warren’s medium (brain and heart infusion with 20 µg/ml hemin and 10 µg/ml folic acid) supplemented with 10% fetal bovine serum at 25 °C. Stationary-phase promastigotes were obtained from 5- to 6-day-old cultures and used throughout.
L. amazonensis promastigote cultivation on 3D COL I matrices
Rat tail extracted COL I solution was diluted with 5-times concentrated DMEM media and completed to the desired density (1.5 or 3.0 mg/ml) with DMEM. 0.1 M NaOH was used to neutralize the solution pH. 107 promastigotes were mixed with 1 ml of diluted and neutralized COL I solution and incubated at 37 °C for a 1 h polymerization. After complete polymerization, serum-free RPMI medium was added to the top of the matrices to feed the cells and hydrate the COL I matrix. The culture was then left at 27 °C for no more than 72 h. Viability was measured using a fluorescent Live/Dead assay (Invitrogen, L3224) and only samples with viability scores higher than 90% were used.
Scanning electron microscopy
COL I matrix samples cultivated with L. amazonensis promastigotes were fixed with 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.2) for 1 h and then washed overnight with PBS. After fixation, promastigotes in the COL I matrix were postfixed for 30 min in a solution containing 1% OsO4, 1.25% potassium ferrocyanide and 5 mM CaCl2 in 0.1 cacodylate buffer, washed in the same buffer and then dehydrated in an ethanol series from 30% to 100%. Finally, samples were critical point dried, coated with a thin gold layer in a Balzers gold sputtering system, and observed in a FEI-Quanta scanning electron microscope.
Protease inhibitors
Metalloprotease inhibitors included 200 nM Marimastat or 5 mM o-phenantroline, cysteine protease inhibitors included 20 ng/ml Cystatin or 100 µM of trans-epoxysuccinyl-L-leucylamido-(4-Guanidino)butane (E-64) and the serine protease inhibitor used was 1 mM 4-(2-aminoethyl)-benzenesulfonylfluoride (AEBSF), (all protease inhibitors were purchased from Sigma-Aldrich). Protease inhibitors (PI) mix was used in some experiments. The PI mix used was composed of AEBSF, E-64, Cystatin and Marimastat.
Migration assay
The L. amazonensis promastigote migration was characterized for all experimental conditions. Samples were placed in a culture chamber, with controlled temperature (27 °C), adapted to an inverted microscope Nikon Eclipse TE 300 (Nikon, Melville, NY). Brightfield images were captured for 5 min with a Hamamatsu C2400 CCD camera (Hamamatsu, Japan) and digitalized by a SCION FG7 frame grabber (Scion Corporation, Torrance, CA) using a frame rate of 2 frames/s. The process was repeated every 24 h for a total period of 72 h. For each movie 15 different cells were marked with black dots. The distance covered by the black dot trajectories were then obtained by image analysis using the ImageJ software (National Institutes of Health, USA) and the indirect promastigote migration rates were determined by dividing the distance covered (µm) by the time in seconds (sec). Three different movies were made and analyzed for each condition.
Transmatrix migration (invasion) assay
For the transmatrix migration assay, the COL I matrix was previously prepared without L. amazonensis in a 96 well plate. Then, 100 µl serum free RPMI media containing 106 promastigotes was added to the top of a 150 µl polymerized matrix. After 48 h, the RPMI media was washed out and the samples were fixed and processed for histology. The quantity of promastigotes inside the matrix (complete transmatrix process) and the distance migrated from the top to the bottom of the COL I matrix (transmatrix invasion distance) was used to determine the invasion ability of L. amazonensis. The transmatrix invasion distance was normalized considering 100% as the total distance between the top and the bottom of each COL I matrix histological cut. Similar results were obtained when the data were analyzed without normalization.
COL I matrix degradation assay
L. amazonensis promastigotes were cultivated inside of 3D COL I matrices prepared with a solution containing 5% FITC-labeled type I Collagen (Invitrogen, Molecular Probes). The degradation assay was as modified method from Sugiyama et al. (1980). After 0 h, 24 h, 48 h and 72 h, solid-phase collagens were pelleted (5 min, 16,000 g) and the supernatant containing released FITC-labeled type I Collagen fragments was measured with a spectrofluorimeter (485 nm excitation, 515 nm emission). The background signal and the total degradation were obtained from a cell-free matrix and from a matrix made with the presence of 25 U/ml type II collagenase (Gibco) respectively.
Zymography
Promastigotes cultivated for 48 h inside COL I matrix were placed in an eppendorf tube and centrifuged for 7 min at 1600 g to separate the COL I matrix and cells from the supernatant. After centrifugation, supernatant was filtered to remove any remaining cells. Proteins from the supernatant were then separated by SDS–PAGE gel electrophoresis using gels containing 1.5 mg/ml COL I and 10% acrylamide and bis-acrylamide. After electrophoresis, gels were washed with 50 mM Tris 2.5% Triton-X100 buffer (pH 6.8) for 30 min and then incubated with 50 mM Tris, 10 mM CaCl2, 1 mM DTT buffer (pH 6.8) for 48 h. When indicated, the protease inhibitors o-phenantroline 5 mM or E-64 20 µM were added to the reaction buffer.
3D COL matrix model for promastigote–macrophage interaction
The murine macrophages RAW 264.7 cell line (3 × 105) were mixed with 1 ml of diluted and neutralized COL I solution and incubated at 37 °C for a 1 h polymerization. After complete polymerization, RPMI medium complemented with 5% bovine serum was added to the top of the matrices to feed the cells and hydrate the COL I matrix. The culture was then left at 37 °C for 24 h. Macrophage 3D culture viability was analyzed using a fluorescent live dye fluorescein diacetate (Sigma). After 24 h of incubation, the medium was removed and Leishmania promastigotes were added to the 3D culture in two ways: 107 promastigotes in 200 µl of RPMI were injected inside the matrix containing macrophages (injection mode) or promastigotes were added to the top of the matrix (invasion mode).
Confocal microscopy
Live promastigotes were labeled with 5 mM of cell tracker green (Invitrogen-C7025) for 15 min. Macrophages cultured in a 3D COL I matrix for 24 h were than labeled and put to interact with labeled L. amazonensis promastigotes for 4 h. After interaction the samples were fixed with 4% paraformaldehyde and than permeabilized with TritonX100 0.05% and incubated with rhodamine phalloidin (Invitrogen) overnight at 4 °C. The samples were visualized by the confocal microscopy LEICA TCS SP5 and the images were processed using the software LAS AF.
Results
L. amazonensis promastigotes extensively interact with and remodel 3D COL I matrix fibers
Scanning electron microscopy (SEM) of promastigotes cultivated in a 3D COL I matrix showed a huge number of promastigotes extensively adhered to COL I fibers (Fig. 1). The presence of promastigotes altered the organization of the COL I fibers network (Figs. 1B–1F). This matrix remodeling transformed the original homogeneous COL I matrix (Fig. 1A) into a meshwork divided into areas of high fiber density and large fiber-free channels (Figs. 1E and 1F). Promastigote cell bodies and/or flagella were observed in contact with the COL I fibers (Fig. 1C). SEM revealed promastigotes completely trapped in the high density COL I fiber areas (Fig. 1D) and promastigotes in the fiber-free channels areas. Promastigotes had their cell body elongated over time during the 3D cultures on COL I matrices (Supplemental Information 1), a feature normally associated with standard liquid cultures.

Figure 1: SEM analysis of L. amazonensis interaction with COL I matrices.
SEM of normal COL I fiber organization in the 3D matrix without promastigotes (A) and promastigotes interacting with COL I fibers of a 3D matrix (B–F). Promastigotes extensive alter COL I fiber organization (B–D). In some matrix areas the L. amazonensis promastigotes were completely trapped between COL matrix fibers (E). L. amazonensis promastigotes were able to interact with COL I fibers through their cell bodies and flagella (C).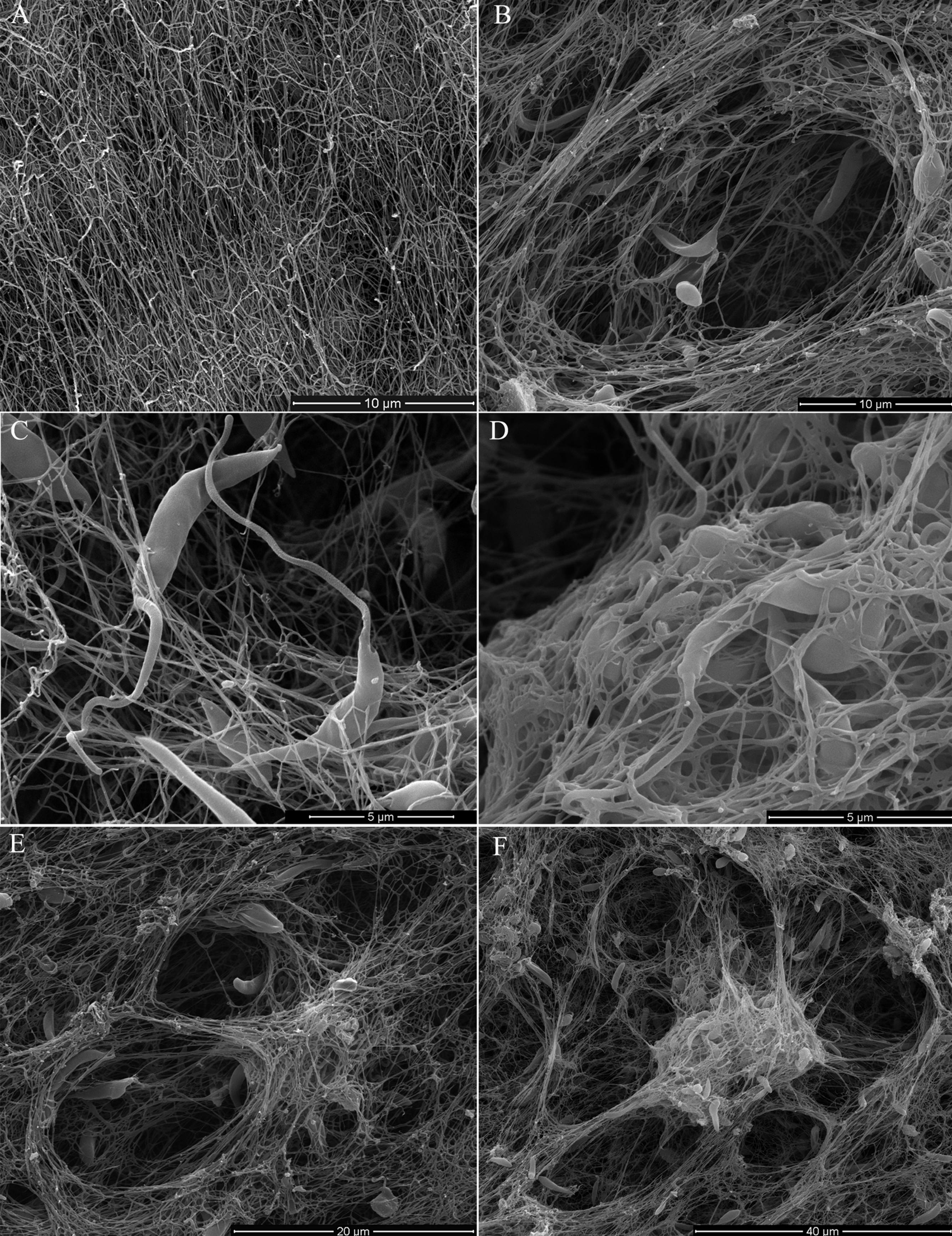{kind=link}
3D COL I matrix degradation by L. amazonensis promastigotes
Promastigotes were cultivated inside 3D FITC-COL I matrices to measure their ability to degrade this network. L. amazonensis promastigotes degraded COL I fibers in a time-dependent manner and density-independent way (Figs. 2A and 2B). After 72 h, around 23% of the COL I total matrix was degraded. This degradation was significantly reduced in the presence of the cysteine protease inhibitors cystatin or E-64 (Fig. 3). The presence of the metalloprotease inhibitor marimastat also reduced COL I degradation, but no effect was observed with the serine protease inhibitor AEBSF (Fig. 3). Incubation with anti-gp63 caused a small but significant reduction of COL I degradation, suggesting that this metalloprotease is also capable of COL I cleavage. COL degradation was not completely abolished in the presence of protease inhibitor (PI) mix (Fig. 3). Although the concentration of inhibitors used could be insufficient for complete inhibition, we chose not to use higher concentrations of this inhibitor mix because they affected cell viability (Table S1). Zymography of culture supernatant from L. amazonensis grown in a COL I matrix detected two collagenase protease bands (Supplemental Information 2). The collagenase bands completely disappeared in the presence of the metalloprotease inhibitor O-phenanthroline, demonstrating the secretion of COL I-degrading metalloproteases by L. amazonensis promastigotes (Supplemental Information 2). Other classes of proteases could not be detected by COL zymography in the conditions tested. However, reaction buffers of pH lower than 6.8 destroy the COL gels, restricting the use of this assay for Leishmania cysteine proteases.
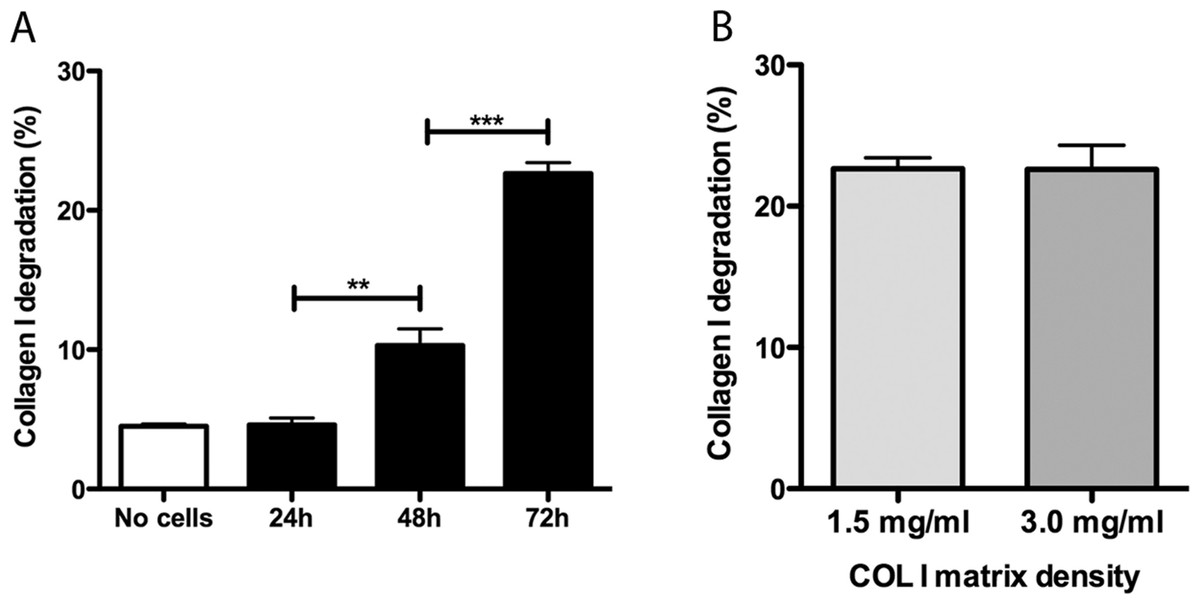
Figure 2: Collagen matrix degradation by L. amazonensis promastigotes.
COL I matrix degradation was evaluated by the amount of COL I-FITC found in the supernatant after different periods of interaction of L. amazonensis promastigotes with COL I matrix (1.5 mg/ml) (A) or after 72 h of interaction on low (1.5 mg/ml) or high (3.0 mg/ml) COL I matrix density (B). Data are expressed as the mean with whiskers representing the standard error of the mean. T test with Bonferoni’s multiple test correction ***, p < 0.0001; **, p < 0.001.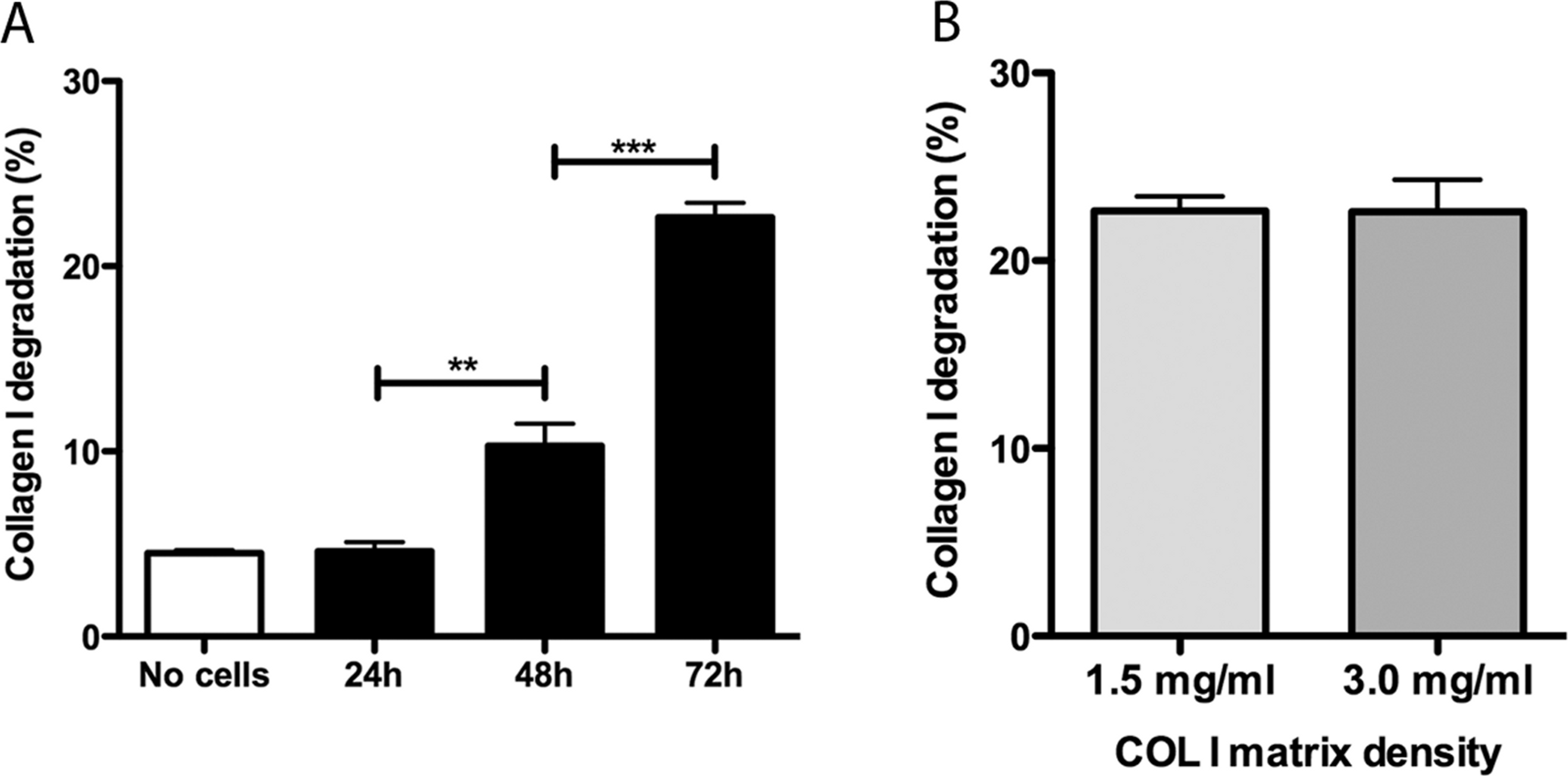{kind=link}
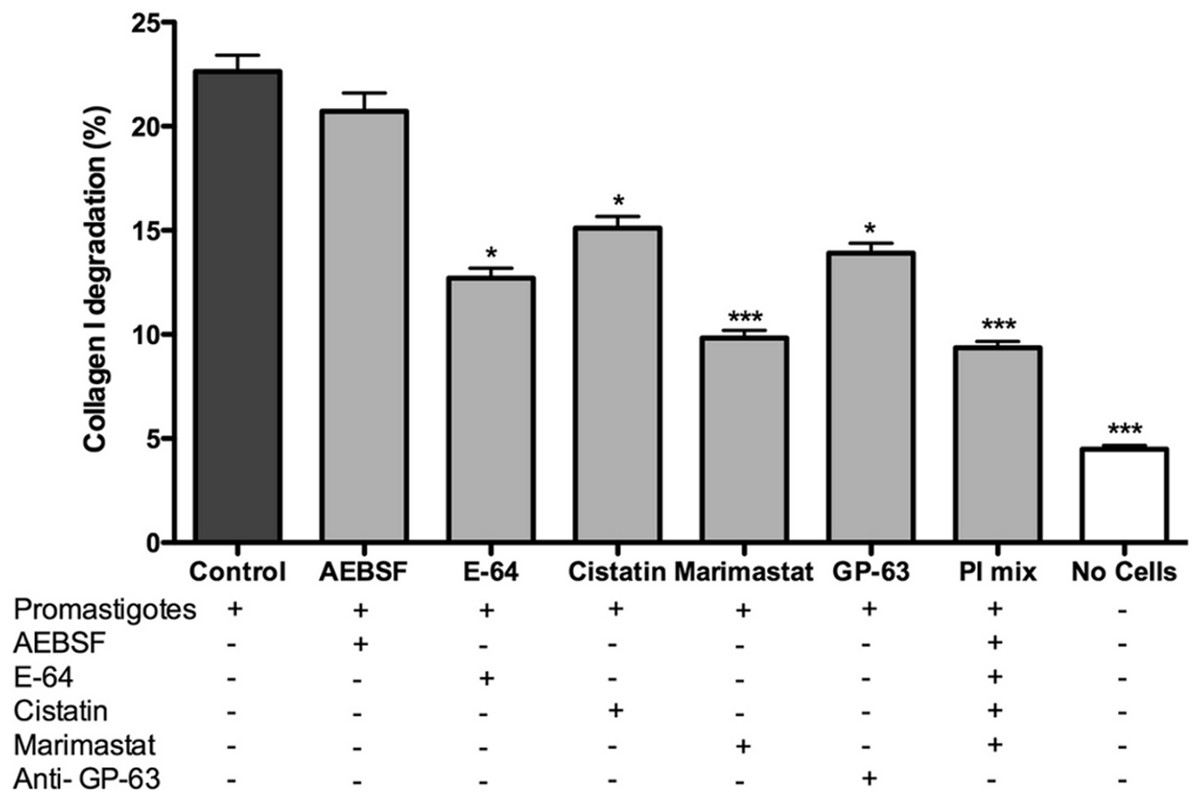
Figure 3: L. amazonensis promastigotes degrade COL I fiber by protease release.
Promastigote interaction with the COL I matrix occurred in the presence of different proteases inhibitors (PI) and PI mix. Cysteine PI, E-64 (100 µM) and cystatin (20 ng/ml), caused a significant reduction of COL I degradation, however, the metalloprotease inhibitor marimastat (200 nM) caused an even higher reduction. The presence of anti GP-63 (1:50) also reduced COL I degradation ability. Serine protease inhibitor AEBSF (1 mM) had no effect on the degradation. Whiskers represent the standard error of the mean. 1-way ANOVA, Kruskal–Wallis test, *, p < 0.05; ***, p < 0.0001.{kind=link}
L. amazonensis migration and transmatrix migration in a 3D COL I matrix
L. amazonensis promastigote migration inside of a 3D COL I matrix was observed by video microscopy (Video S1). These flagellated protozoa displayed an intermittent migration mode with displacement periods (run phase) intercalated with non-displacement periods (freeze phase). The freeze and run migration mode contrasts with the continuous swimming migration mode found in conventional in vitro liquid culture. The migration speed in 3D cultures significantly increased over time (Fig. 4), reaching the highest mean rate (6 µm/s) after 72 h. Even at the first time point, just after COL I matrix polymerization, promastigotes had a slow but non-zero migration rate (Fig. 4). Over time, the displacement periods were more frequent and lasted longer, reflecting a higher migration speed (Fig. 4). Thus, there was a correlation between time, increased speed, and COL I matrix remodeling by promastigotes.
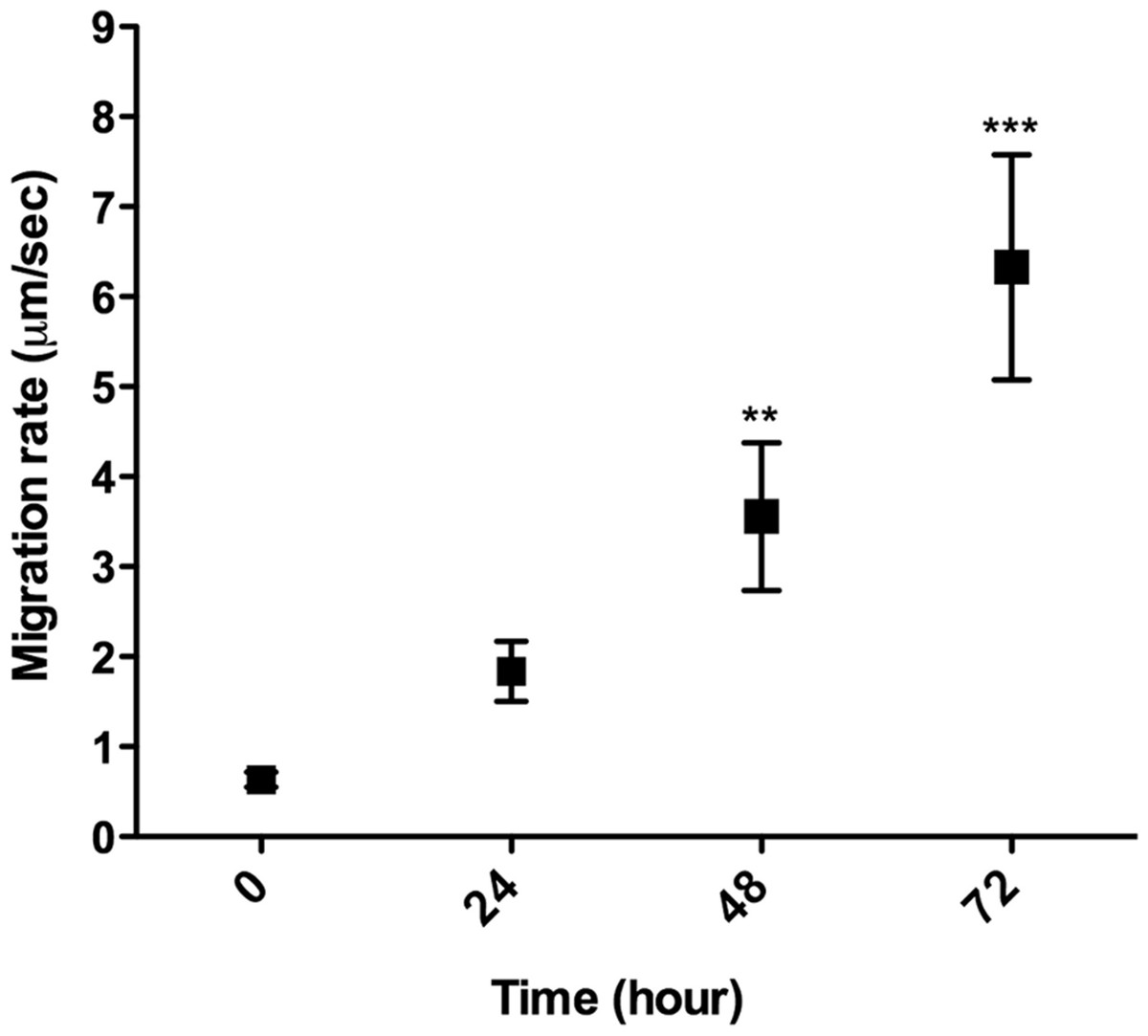
Figure 4: 3D migration rate increases over the time in COL I matrix.
L. amazonensis promastigote migration rate inside of COL I 3D matrix over time. The migration was observed by videomicroscopy and the rate analyzed by ImageJ software. Bars represent the standard error of the mean. 1-way ANOVA, **, p < 0.001; ***, p < 0.0001.{kind=link}
In addition to migration within a 3D COL I matrix, we tested the ability of promastigotes to adhere to and cross the matrix (transmatrix migration). Indeed, when promastigotes were added to the top of a polymerized 3D COL I matrix they were able to adhere to and penetrate through the COL I fiber meshwork (Fig. 5A). This matrix invasion ability (transmatrix migration) demonstrates that promastigotes are not only able to migrate inside of a 3D COL matrix (Fig. 4) but are also able to cross from a liquid environment (RPMI media) to a 3D meshwork (COL I matrix). Here it is important to note that promastigote transmatrix migration was observed without the addition of an external chemo-attractive factor, demonstrating a natural COL I matrix invasive ability for this intracellular parasite. The distance reached by promastigotes from the top of the matrix (transmatrix migration distance) was significantly reduced at higher COL I densities (3.0 mg/ml) compared to lower densities (1.5 mg/ml) (Fig. 5B).

Figure 5: Trans-matrix migration ability is dependent on the COL I matrix density.
Histological transversal cuts of COL I 3D matrix at 1.5 or 3.0 mg/ml after 72 h of L. amazonensis promastigote (arrows) invasion (A). Promastigote COL I transmatrix migration was measured by the depth reached by the promastigotes divided by the total size of each histological transversal cut (B). Whiskers represent the standard error of the mean. Student’s T test ***, p < 0.0001.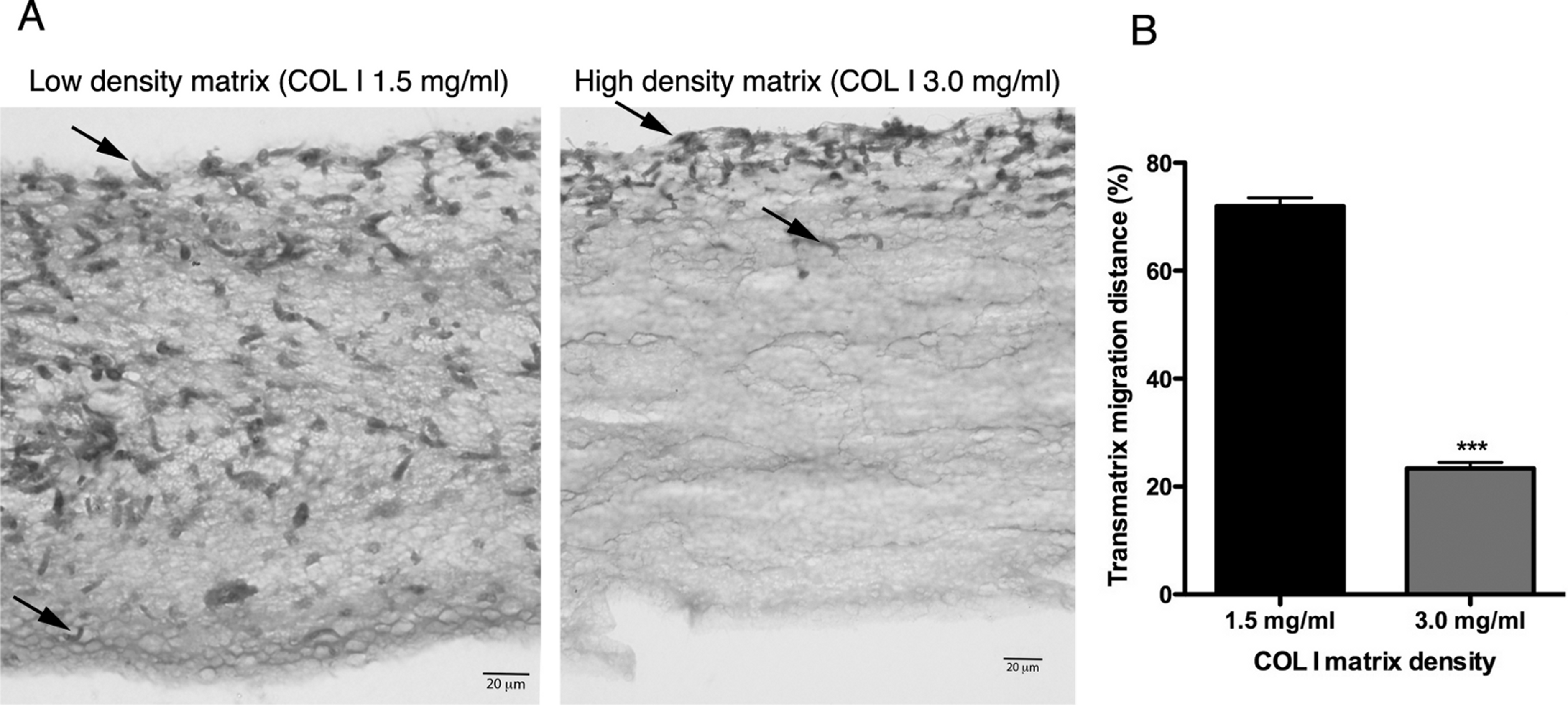{kind=link}
Extracellular protease activity promotes L. amazonensis 3D COL I migration and invasion
The correlation between protease activity and promastigote migration in 3D was analyzed by videomicroscopy in the presence or absence of a protease inhibitor (PI) cocktail. The promastigote migration rates were analyzed using the software ImageJ. In the presence of a PI mix composed of E-64, Marimastat and Cystatin, the promastigote migration rates inside the 3D COL I matrix were significantly reduced, demonstrating that protease activity increases migration rates (Fig. 6). The fraction of promastigotes with migration rates exceeding 5 µm/s was also significantly decreased by the PI mix (Fisher’s exact test, Fig. 6). The reduction of migration rates could also be observed when only the run phase was considered in the analysis, indicating an inhibition of both frequency and maximum speed of migration.
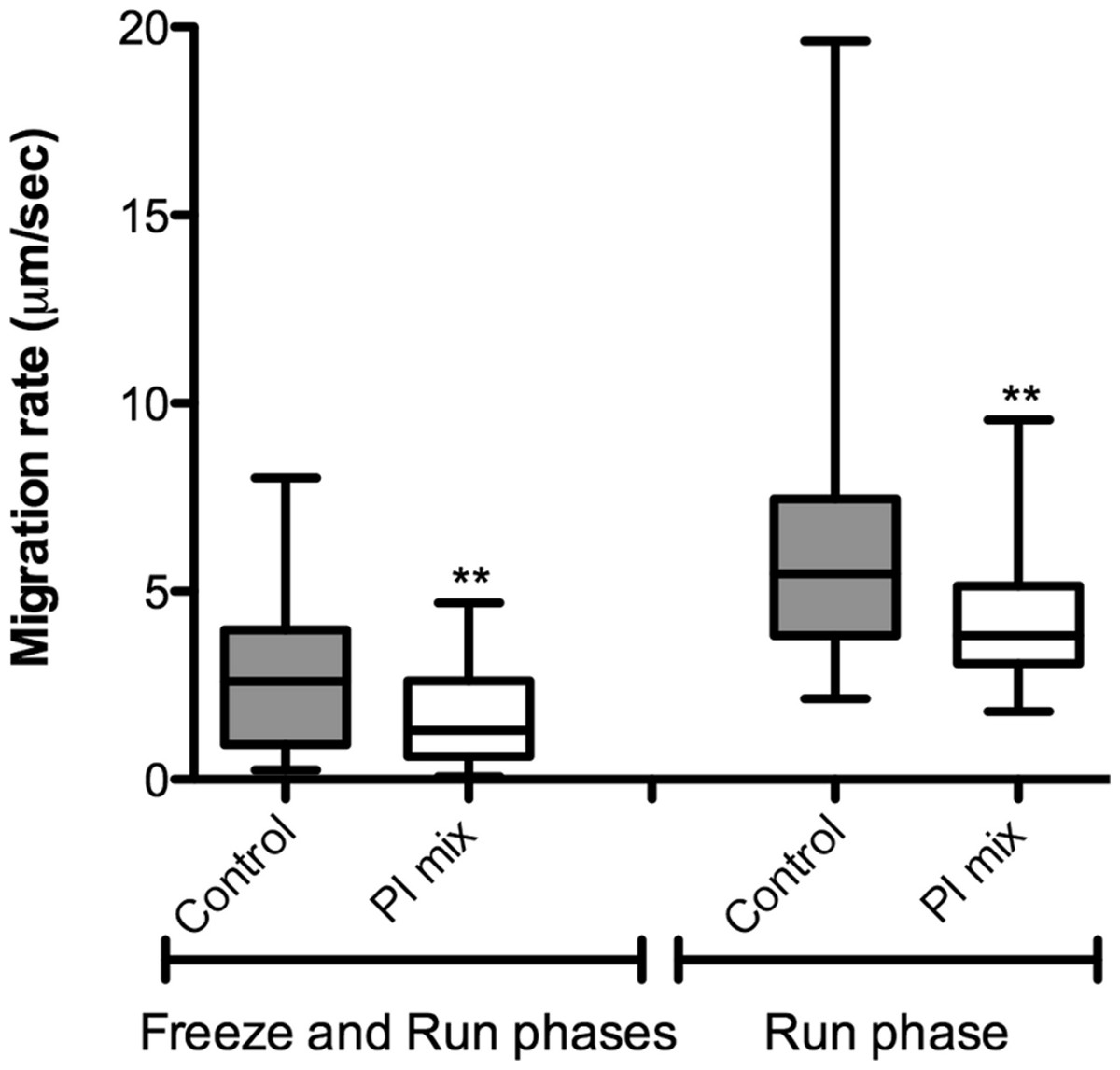
Figure 6: L. amazonensis migration in 3D cultures with and without protease inhibitors.
L. amazonensis promastigote migration rate after 24 h inside of COL I 3D matrix with or without the protease inhibitors (PI) mix. The migration was observed by videomicroscopy and analyzed by ImageJ software. The migration rate was analyzed in two ways: total migration (freeze and run phases) or during the moving phase of the migration only (run phase). Whiskers represent the minimum and maximum values. Differences in the mean value were tested using Mann–Whitney test **, p = 0.0074 and p = 0.0019; n = 45. Fisher’s exact test was used to test differences in the fraction of promastigotes with migration speed greater than 5 µm/s, p = 0.0124 (Freeze and run phases) and p = 0.0031 (run phase).{kind=link}
In addition, the role of protease activities in promastigote transmatrix migration was evaluated after 72 h, in the presence or absence of PI mix (Fig. 7). Transmatrix migration is dependent on two processes: the ability of cells to penetrate into the matrix and, once inside, the ability to migrate. The PI mix presence reduced by almost 40% the number of promastigotes inside of the COL I matrix (Fig. 7A) and also significantly reduced the maximal transmatrix migration distance (in depth) achieved by the promastigotes (Fig. 7B).
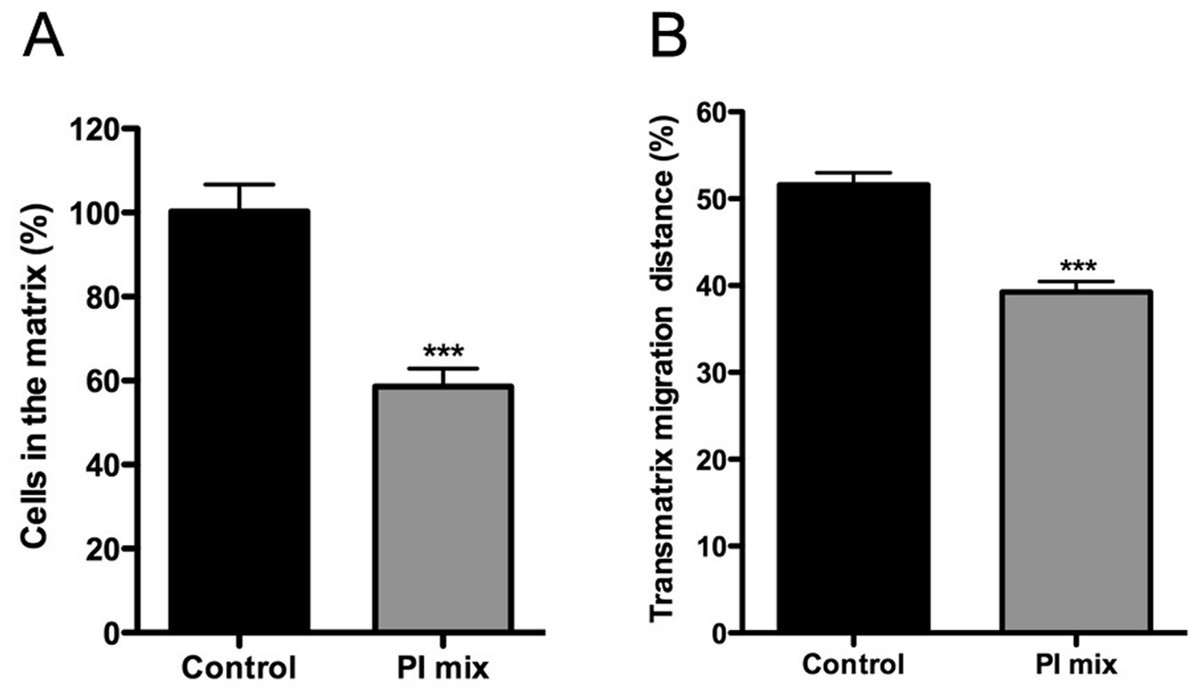
Figure 7: Trans-matrix migration ability is affected by the presence of a protease inhibitors (PI) mix.
Leishmania promastigotes were added to the top of COL I matrix of 1.5 mg/ml and the trans-matrix invasion ability in presence or absence of protease inhibitors was analyzed by the percentage of parasites inside the matrix (A) and by the maximum distance (in depth) achieved by the parasites (B). In both cases the presence of protease inhibitors mix significantly reduced the transmatrix migration ability.{kind=link}
Together the data demonstrate that extracellular protease activity is important for L. amazonensis promastigote transmatrix and migration in 3D cultures (Figs. 6 and 7), due to COL I matrix degradation/remodeling.
Interaction of macrophages and promastigotes in 3D COL I matrix
For an even better mimic of what would happen during Leishmania infection, we cultivated macrophages inside the COL I matrix for 24 h, after which L. amazonensis promastigotes were added to the top of the matrix (Fig. 8). After 6 h of interaction most of the parasites were found inside the matrix (Fig. 8), a shorter time than in the macrophage-free matrix. This suggests an influence of macrophages on the invasion ability of Leishmania promastigotes. 3D visualization by confocal microscopy of macrophage-promastigote interaction (Figs. 9A and 9B) showed macrophages possessing a round morphology with many actin-membrane filament projections (Fig. 9C). In some cases these thin actin-membrane projections were touching a promastigote’s surface (Fig. 9D). SEM microscopy observations showed the extensive contact of macrophages and promastigotes with COL I fibers in the 3D culture model (Fig. 10). The fibrotic characteristic of the ECM 3D environment seems to create a effective barrier for physical contact between Leishmania and macrophages, in contrast to 2D culture models in which macrophages become infected much earlier.
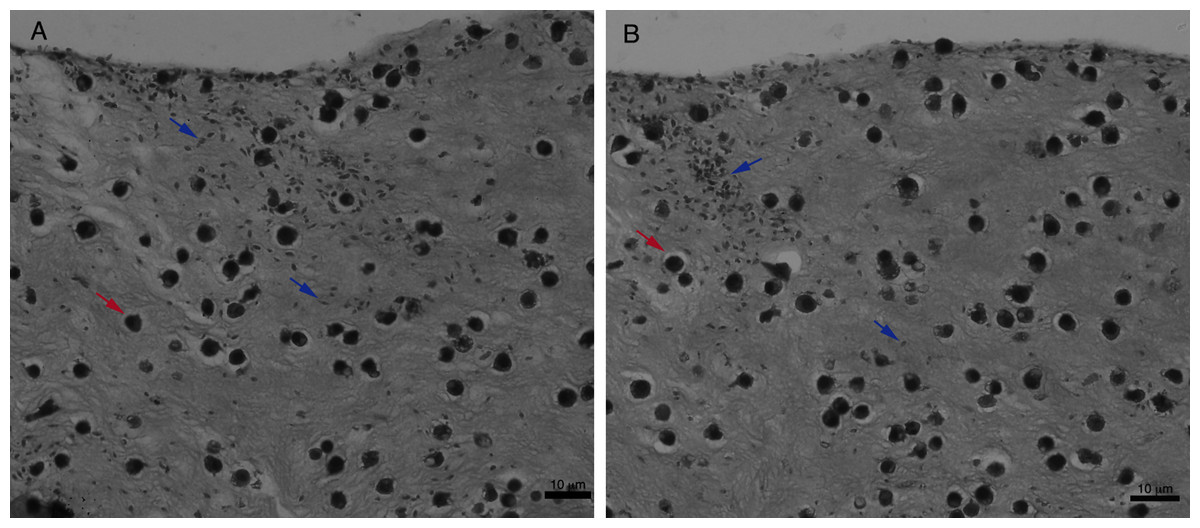
Figure 8: Promastigote trans-matrix migration ability in the presence of macrophages.
Leishmania promastigotes were added to the top of the macrophages 3D COL I culture. After 6 h the samples were fixed, prepared for histology and transversally cut. Image shows transversal cuts of COL I matrix containing macrophages (red arrows) 6 h after addition of L. amazonensis (blue arrows) promastigotes (A–B) demonstrating many parasites able to invade the COL I matrix.{kind=link}

Figure 9: Macrophage–Leishmania 3D interaction vizualization by confocal microscopy.
Leishmania interaction with macrophages in the 3D COLI matrix was visualized by confocal microscopy. Live Leishmania promastigotes were labeled with cell tracker green before interaction. Macrophage actin filaments were stained with phalloidin (red). Images are 3D reconstruction of a 11 µm deep image.{kind=link}
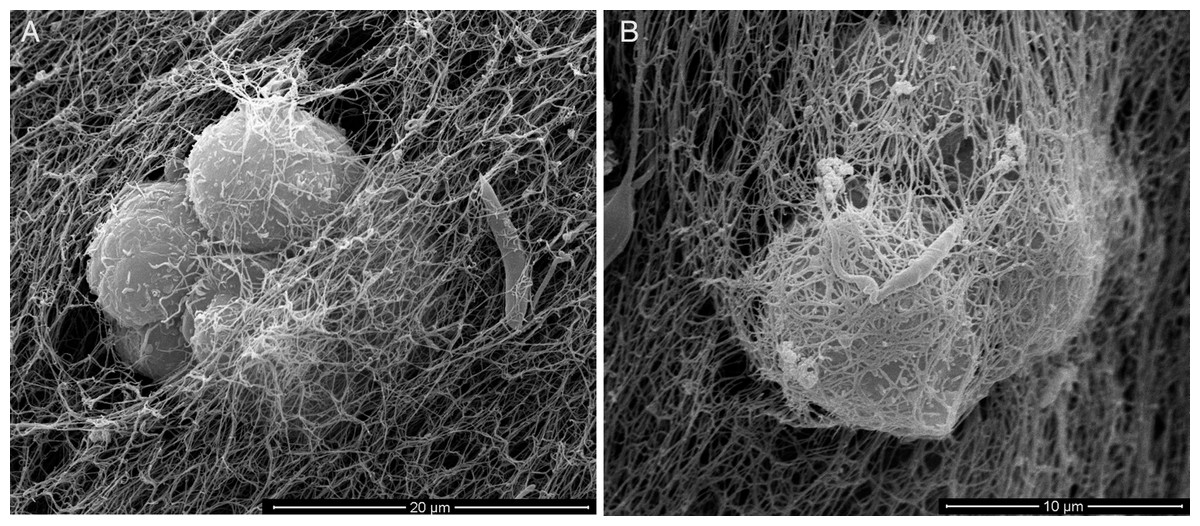
Figure 10: SEM of L. amazonensis–macrophages 3D interaction.
SEM visualization of macrophages cultured in a 3D COL I matrix after 3 h (A) or 4 h (B) of interaction with L. amazonensis promastigotes. In order to visualize the promastigote–macrophage interaction inside the matrix the samples were transversally cut in liquid nitrogen during SEM samples processing.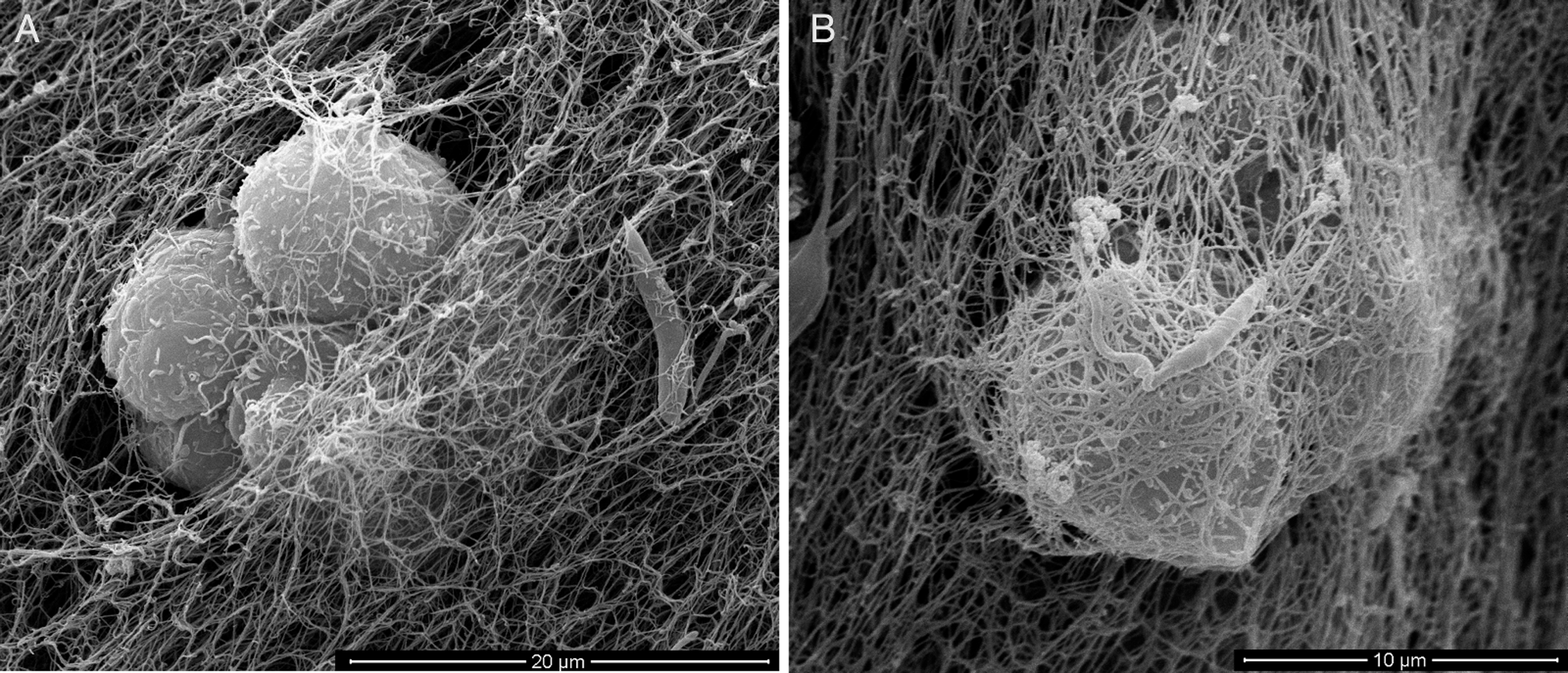{kind=link}
The injection of promastigotes directly into the matrix containing macrophages allowed a good distribution of promastigotes inside the COL meshwork and enabled the observation of macrophage-promastigote interaction in the first minutes of interaction (Video S2). That experiment demonstrated the promastigotes migrating further and faster than macrophages in the 3D culture. Additionally, the promastigotes exhibited the same freeze and run migration mode observed in the macrophage-free 3D matrix.
Discussion
In the human host, Leishmania amazonensis is an intracellular parasite that mainly proliferates inside macrophages. Therefore, the interaction of the Leishmania parasite with the host ECM has been neglected. In fact, it is critical to understand the interaction of Leishmania promastigotes with the ECM because when a host is infected, the promastigotes must pass through the dermis ECM, remaining there until the first contact with macrophages (Lira, Rosales-Encina & Arguello, 1997; McGwire, Chang & Engman, 2003) or other potential host cells. Our results demonstrated that L. amazonensis promastigotes extensively contacted the COL I fibers in a 3D matrix (Fig. 1D). Following these interactions were drastic modifications of the COL I fiber organization (Figs. 1A–1F). Many publications have shown a correlation between ECM remodeling and inflammation, and indeed, abnormal ECM organization is prominent in many diseases such as tissue fibrosis and cancer (Cox & Erler, 2011). This suggests that COL I matrix remodeling by promastigotes could play an important role during the infection process (Larsen et al., 2006; Stamenkovic, 2003).
3D culture is becoming a common approach for the study of several types of mammalian cells (Voytik-Harbin, 2001) but it is still not commonly used for pathogens or unicellular organisms (Behnsen et al., 2007). In this work we establish, for the first time, 3D in vitro cultivation of L. amazonensis promastigotes. This in vitro 3D culture mimics (at least in part) the physical dynamics of the Leishmania human host environment and makes possible the in vitro study of the mechanisms involved in the interaction of L. amazonensis with the ECM. Conventionally, promastigotes are cultivated in rich serum-containing media to maintain 90% viability. However, in our model, L. amazonensis promastigotes were able to survive (>90% viability) and proliferate for 72 h inside a 3D matrix composed of 1.5 or 3.0 mg/ml of COL I in serum-free RPMI medium, suggesting that this flagellated parasite, like some mammalian cells, required fewer survival signals when in 3D culture (Alavi & Stupack, 2007; Eke & Cordes, 2011). This could be due to the physical support presented in the 3D matrix or to the chemical signaling provided by the COL I molecules (Weaver et al., 2002).
The roles that Leishmania proteases play during inflammation and interaction with macrophages are important for infection, especially in the case of the metalloprotease gp63 (Silva-Almeida et al., 2012b; Yao, 2010). Here, we presented a new role for extracellular Leishmania proteases. Protease blockage by a mixture of inhibitors decreased matrix degradation and affected promastigote migration and transmigration among COL I fibers. The ability of promastigotes to cleave COL I matrix fibers was analyzed by a degradation assay using fluorescent COL I. We showed that promastigotes can actively degrade COL I fibers and that the degradation was independent of the matrix density (Fig. 2). Metallo- and cysteine proteases were implicated in this degradation (Fig. 3). The presence of anti-gp63 significantly reduced the COL I degradation, indicating the importance of this protease in the cleavage of COL I fibers. However, the presence of the PI mix did not completely abolish the COL I degradation. These findings suggest that either the inhibitors are not strong enough to block all the extracellular proteases without affecting the parasite survival, or that the mechanical pushing and pulling by the promastigote flagella also contribute to COL I fiber degradation.
Following the promastigotes by videomicroscopy, we showed that these parasites were able to migrate inside the 3D COL I matrix (Fig. 4). However, the migration mode in this 3D culture was completely different from the one in standard culture medium. In the 3D culture environment, the promastigote migration oscillates between periods of movement and stationary periods, which we described as a freeze and run migration mode (Video S1). This data is consistent with our SEM data (Fig. 1), showing some promastigotes completely trapped in the COL I fibers (freeze phase) and others COL I fiber-free (run phase), probably passing through the COL I matrix tunnels created by matrix remodeling (Fig. 1). The migration rate and COL I matrix degradation both increased over time (Figs. 2–4), consistent with the hypothesis that the run phase of migration is linked to the presence of COL I matrix tunnels.
In the presence of PI mix the rate of promastigote migration inside the matrix was significantly reduced (Fig. 6). This reduction was more pronounced when only the run phase was considered; the maximum speed reached in that phase was significantly slower in the presence of PIs (Fig. 6). These results suggest that the extracellular Leishmania proteases and the subsequent COL I matrix remodeling lead directly to an increased migration rate.
Transmatrix migration is an important skill of metastatic cells, invasive extracellular parasites, and of immune cells such as neutrophils and macrophages (Hagedorn & Sherwood, 2011; Schoumacher, Louvard & Vignjevic, 2011). L. amazonensis promastigotes were able to invade matrices of multiple COL I densities (Fig. 5), showing that intracellular parasites possess the ability not only to migrate inside of a COL I matrix but also to adhere to and invade these matrices (transmatrix migration), raising the unexpected possibility of invasive behavior by this intracellular parasite during infection. It is important to mention that all our experiments were made with stationary phase promastigotes (Methods) not enriched for metacyclics and we do not have access to the metacyclic/procyclic proportion of our promastigote culture. Therefore, it is possible that these forms have different behavior in 3D cultures and further work is needed to evaluate their roles.
Taken together, we showed that promastigotes degrade COL I matrix via extracellular cysteine and metalloprotease, and these proteases play a role during migration and invasion of 3D matrices. Together these results suggest that the matrix remodeling contributes to L. amazonensis migration and invasion by helping promastigotes to get free from the COL I matrix traps (reducing the freeze period), or even to open the tunnels inside of the matrix that enable a faster maximum migration speed. In both ways, protease release and the subsequent matrix remodeling could help the L. amazonensis promatigotes to create a 3D environment that facilitates access to their intracellular “home”.
Furthermore, we establish a 3D co-culture model with macrophages seeded in a COL I matrix for 24 h before the addition of L. amazonensis promastigotes. The promastigotes were added in two ways: at the top of the matrix (invasion model) or by syringe injection directly inside the matrix. Both models resulted in direct Leishmania–macrophage contact inside the 3D environment. However, the invasion model was necessary for the analysis of the trans-matrix migration of promastigotes during interaction (Fig. 8), while the injection model created a more homogenous distribution of promastigotes in the matrix, allowing videomicroscopy of Leishmania–macrophages interactions (Video S2). Surprisingly, promastigotes migrated faster than macrophages in this model, supporting an active participation of promastigotes during Leishmania infection. The Leishmania–macrophage 3D interaction assay established here will help us to better understand some aspects of Leishmania–macrophage and Leishmania-matrix interaction and could become an important model for initial drug screens.
- COL I
Collagen I
- 3D
three-dimensional
- ECM
extracellular matrix
- FITC
fluorescein isothiocyanate
- PI
protease inhibitors
- SEM
scanning electron microscopy
Abbreviations
Supplemental Information
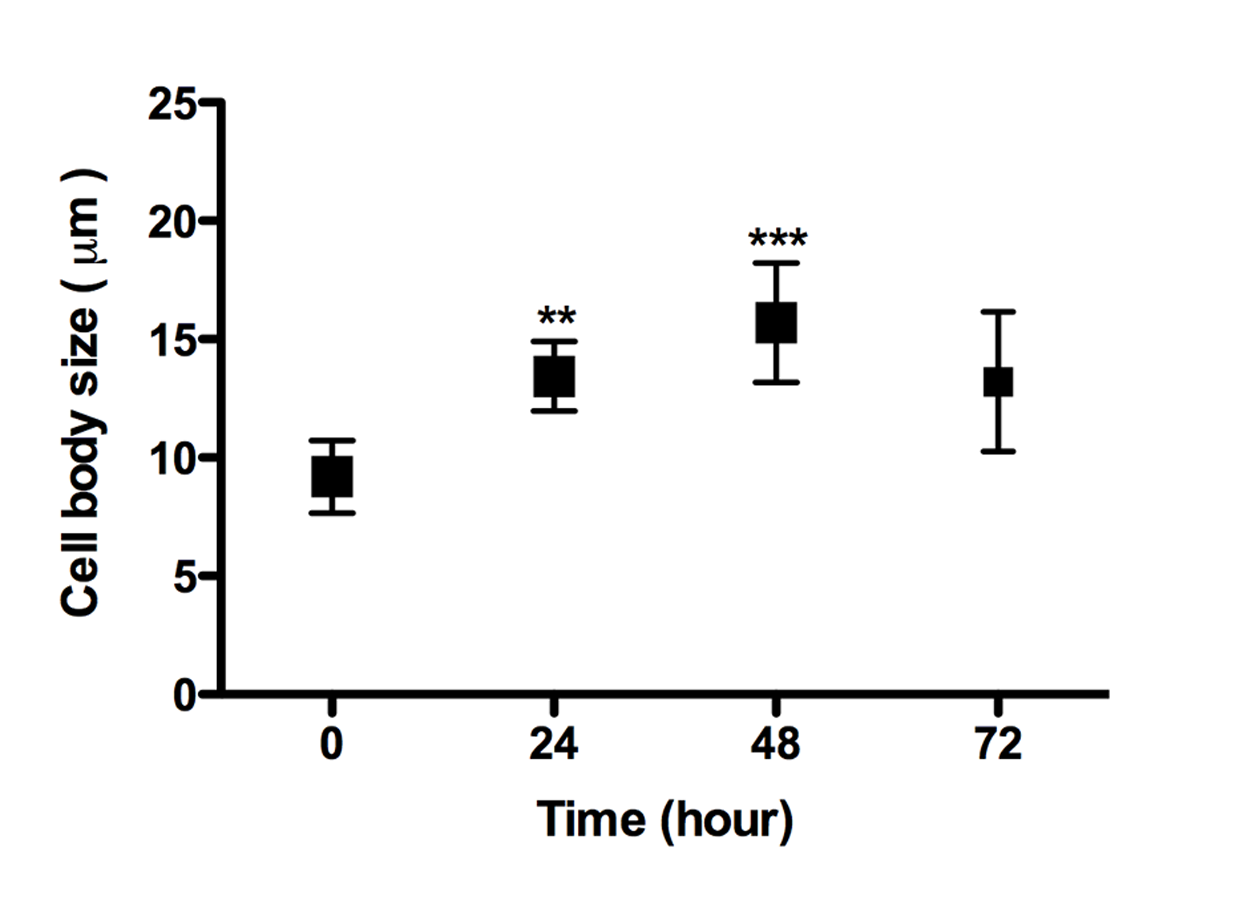
Promastigotes cell body elongation during 3D culture
Promastigote cell body size over cultivation time inside the 3D COL I matrix. Whiskers represent the standard deviation. 1-way ANOVA **, p < 0.001; ***, p < 0.0001; n = 15.
{kind=link}
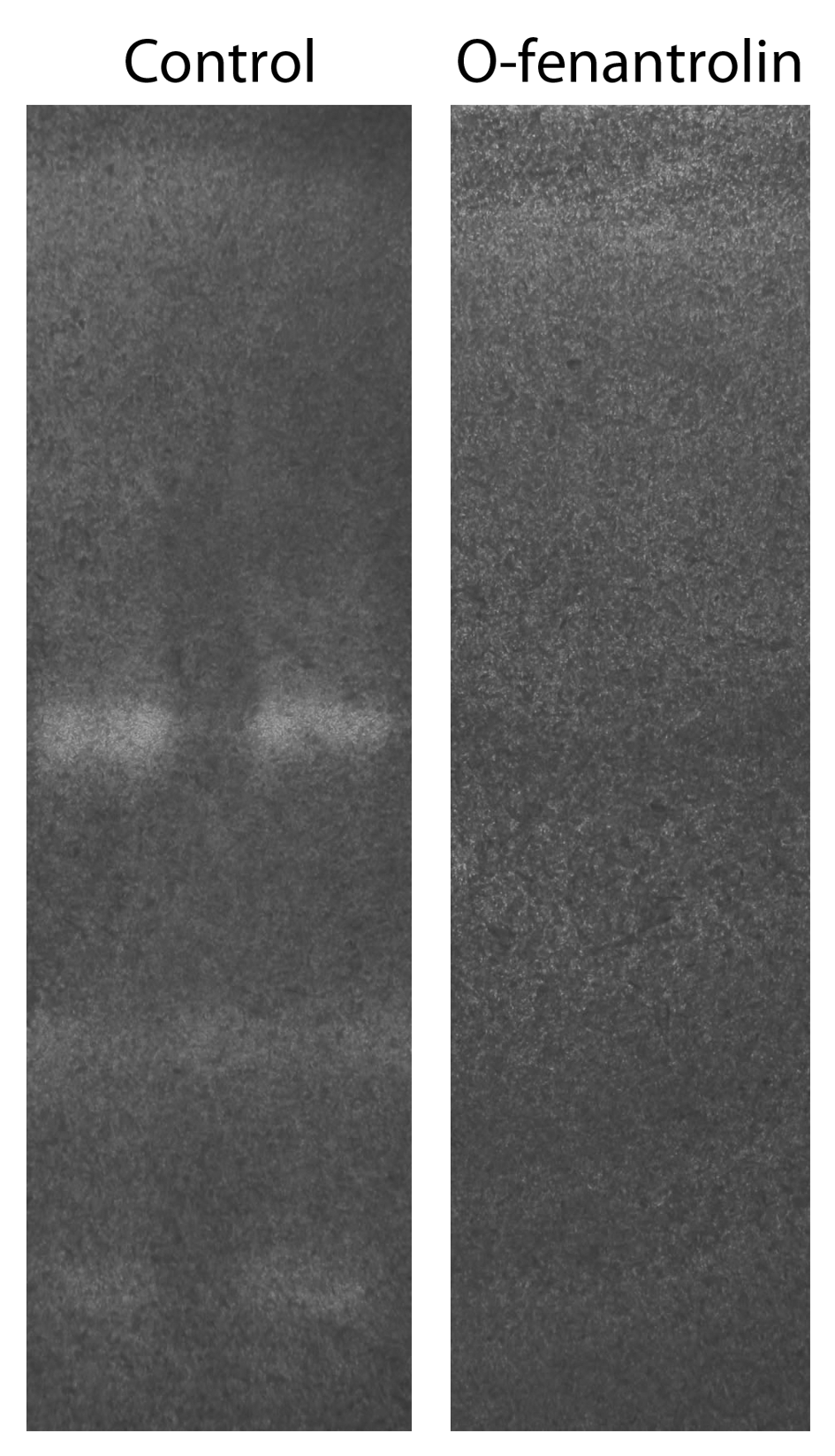
COL I zymography
Scanned pictures of COL I zymography gels. Gels containing two different samples of 72 h 3D COL I matrix promastigote cultivation supernatant. After electrophoresis the gel was cut in three parts and each part was separately incubated with 50 mM Tris, 10 mM CaCl2, 1 mM DTT buffer (pH 6.8) for 48 h with or without (control) O-phenantroline (5 mM). O-phenantroline presence in the buffer completely inhibited the COL I protease relative band.
{kind=link}
Leishmania-macrophages 3D co-culture
Videomicroscopy of macrophages 3D culture just after L. amazonensis promastigote injection into the COL I matrix. 16 frames per second of a 2 min acquisition video.
L. amazonensis 3D migration in a COL I matrix culture
Videomicroscopy of L. amazonensis promastigotes after 24 h inside of 3D COL I matrix. 16 frames per second of a 2 min acquisition video.
Leishmania amazonensis 3D COL I culture viability after PI mix treatment
Viability was measured using a fluorescent Live/Dead assay (Invitrogen) and only samples with viability scores higher than 90% (green) were used to guarantee same viability as the control samples without PI mix treatment. Important to point out that as demonstrate in that table those viabilities are related to the combination effect of the PI mix in a 3D COL I (in RPMI media) context.