
Ectopic expression of Jatropha curcas APETALA1 (JcAP1) caused early flowering in Arabidopsis, but not in Jatropha
- Published
- Accepted
- Received
- Academic Editor
- Sheila McCormick
- Subject Areas
- Molecular Biology, Plant Science
- Keywords
- Jatropha, Flower identity, Arabidopsis, APETALA1, Physic nut, Flowering
- Copyright
- © 2016 Tang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. Ectopic expression of Jatropha curcas APETALA1 (JcAP1) caused early flowering in Arabidopsis, but not in Jatropha. PeerJ 4:e1969 https://doi.org/10.7717/peerj.1969
Abstract
Jatropha curcas is a promising feedstock for biofuel production because Jatropha oil is highly suitable for the production of biodiesel and bio-jet fuels. However, Jatropha exhibits a low seed yield as a result of unreliable and poor flowering. APETALA1 (AP1) is a floral meristem and organ identity gene in higher plants. The flower meristem identity genes of Jatropha have not yet been identified or characterized. To better understand the genetic control of flowering in Jatropha, an AP1 homolog (JcAP1) was isolated from Jatropha. An amino acid sequence analysis of JcAP1 revealed a high similarity to the AP1 proteins of other perennial plants. JcAP1 was expressed in inflorescence buds, flower buds, sepals and petals. The highest expression level was observed during the early developmental stage of the flower buds. The overexpression of JcAP1 using the cauliflower mosaic virus (CaMV) 35S promoter resulted in extremely early flowering and abnormal flowers in transgenic Arabidopsis plants. Several flowering genes downstream of AP1 were up-regulated in the JcAP1-overexpressing transgenic plant lines. Furthermore, JcAP1 overexpression rescued the phenotype caused by the Arabidopsis AP1 loss-of-function mutant ap1-11. Therefore, JcAP1 is an ortholog of AtAP1, which plays a similar role in the regulation of flowering in Arabidopsis. However, the overexpression of JcAP1 in Jatropha using the same promoter resulted in little variation in the flowering time and floral organs, indicating that JcAP1 may be insufficient to regulate flowering by itself in Jatropha. This study helps to elucidate the function of JcAP1 and contributes to the understanding of the molecular mechanisms of flower development in Jatropha.
Introduction
With the decreasing availability of fossil fuels and the deteriorating trend of environmental pollution, biodiesel has garnered significant attention as an alternative fuel (Mofijur et al., 2016). Physic nut (Jatropha curcas L.) is a perennial plant that belongs to the Euphorbiaceae family. Jatropha is monoecious, with male and female flowers borne on the same inflorescence (Divakara et al., 2010; Pandey et al., 2012; Wu et al., 2011). The potential benefit of growing Jatropha as a cash crop for biofuel in tropical and sub-tropical countries is now widely recognized (Akashi, 2012; Khalil et al., 2013; Pua et al., 2011). Jatropha has been propagated as a unique plant with biodiesel potential because of its multipurpose value, high oil content, high biomass productivity, adaptability to marginal land under a variety of agro-climatic conditions, and non-competitiveness with food production (Akashi, 2012; Khalil et al., 2013; Pandey et al., 2012; Pua et al., 2011). The oil content of Jatropha seeds and kernels ranges from 30 to 40% and 40 to 50% by weight, respectively (Pan & Xu, 2011; Sinha et al., 2015). Oil from Jatropha contains high levels of polyunsaturated fatty acids; therefore, Jatropha is suitable as a feedstock for the production of biodiesel and bio-jet fuel (Ong et al., 2011; Pramanik, 2003). The whole-genome sequence and genetic mapping of Jatropha have been reported (Hirakawa et al., 2012; Wu et al., 2015), and several genetic transformation methods have been established (Fu et al., 2015; Gu et al., 2015; Kajikawa et al., 2012; Kumar et al., 2010; Mao, Ye & Chua, 2013; Misra et al., 2012; Pan, Fu & Xu, 2010). Consequently, it is convenient to clone Jatropha genes and analyze their functions. However, the potential of Jatropha as a biofuel plant is limited by its low seed production (King et al., 2015). Despite the clear evidence of the abundant biomass generated by Jatropha, these data are not indicative of high seed productivity (Ghosh et al., 2007). Jatropha exhibits an overabundance of vegetative shoots and leaves in that could develop into reproductive shoots under suitable conditions. Thus, a reduction of undesired vegetative growth is imperative (Ghosh et al., 2010; Tjeuw, Slingerland & Giller, 2015). In addition, unreliable and poor flowering is an important factor that contributes to low seed productivity in Jatropha (Divakara et al., 2010). Therefore the elucidation of the genetic basis of flowering in Jatropha would be helpful for the molecular breeding of high-yielding Jatropha cultivars.
The APETALA1 (AP1) was identified as a floral meristem identity gene to regulate flowering in many plant species. Mandel et al. (1992) reported that AP1 encodes a putative transcription factor containing a MADS domain. This gene acts locally to specify the identity of the floral meristem and determine sepal and petal development. AP1 and LEAFY (LFY) are pivotal in the switch to the reproductive phase. During floral initiation, a positive feedback loop between AP1 and LFY is mediated by direct interactions (Kaufmann et al., 2010; Liljegren et al., 1999; William et al., 2004). After transition to flowering, the expression of AP1 appears to be only indirectly affected by LFY (Wagner, Sablowski & Meyerowitz, 1999). The AP1, FRUITFULL (FUL) and CAULIFLOWER (CAL) genes act redundantly to control the flower meristem identity and inflorescence architecture by affecting LFY and TFL1 expression levels (Ferrándiz et al., 2000). AP1 and CAL are expressed in floral meristems and developing sepal and petal primordial cells (Blazquez et al., 2006; Mandel et al., 1992).
In Arabidopsis ap1 mutants, the sepals are converted to bract-like structures, the petals are absent, the bract-like organs of the first whorl subtend secondary flowers in the second whorl, and tertiary flowers can also form (Bowman et al., 1993; Irish & Sussex, 1990; Mandel et al., 1992; Ng & Yanofsky, 2001). Recent studies have indicated that AP1 can regulate cytokinin levels through the suppression of cytokinin biosynthesis and the activation of cytokinin degradation. These effects mediate the function of AP1 in establishing determinate floral meristems in Arabidopsis (Han et al., 2014). In 35S:AP1 Arabidopsis plants, extremely early flowering occurs after the production of five leaves, and the primary shoot meristem has been converted into a compound terminal flower. The secondary shoot meristems present in the axils of cauline leaves have been transformed into solitary flowers. In addition, 35S:AP1 Arabidopsis can partially complement the later flowering phenotype of a lfy mutant (Liljegren et al., 1999). Constitutive expression of the Arabidopsis AP1 gene in juvenile citrus seedlings resulted in transgenic citrus plants with fertile flowers and fruits after just one year of growth. The transgenic citrus exhibited an appreciably shortened juvenile phase (Peña et al., 2001). In Populus, the overexpression of the Populus ortholog of APETALA1 (LAP1) produced a novel function in photoperiodic regulation of seasonal growth, the LAP1 overexpression resulted in severe attenuation of SD-mediated growth cessation in hybrid aspen (Azeez et al., 2014).
However, the function analysis of AP1 gene in Jatropha has not been reported. Currently, only one flowering-related gene, Jatropha FLOWERING LOCUS T (JcFT), has been functionally analyzed in Jatropha (Li et al., 2014). Overexpression of JcFT can produce more seeds in a shorter time frame by shortening the flowering time in Jatropha, suggesting the possibility to increase seed yield by manipulating the flowering time (Chen et al., 2014). Therefore, in this study, we cloned and characterized a Jatropha AP1 homolog, JcAP1, through genetic complementation of the Arabidopsis AP1 loss-of-function mutant ap1-11. We analyzed the function of JcAP1 in flowering induction and floral organ specification using transgenic Arabidopsis and Jatropha plants.
Materials and Methods
Plant materials and growth conditions
The roots, stems, mature leaves, inflorescence buds, flower buds, male flowers, female flowers and fruits of Jatropha were collected during the summer from Xishuangbanna, Yunnan Province, China. All of the tissues to be prepared for qRT-PCR were immediately frozen in liquid N2 and stored at −80°C until use. The wild-type (WT) Arabidopsis thaliana Columbia ecotype (Col-0) and the ap1-11 mutant of the same ecotype were purchased from The Arabidopsis Information Resource (TAIR) website (http://www.arabidopsis.org/). The seeds of the Arabidopsis plants were germinated on 1/2 MS medium for one-week. Then, the seedlings were transferred to peat soil in plant growth chambers maintained at 22 ± 2 °C under long-day (16 light/8 h dark) or short-day (8 light/16 h dark) conditions. Phenotype analysis was performed on homozygous (T2) Arabidopsis plants and heterozygous (T0) Jatropha plants. More than 20 plants were used for the characterization of each Arabidopsis genotype. The number of rosette leaves and the number of days between transplantation to soil and appearance of the first visible flower bud were recorded. The aboveground tissues of 15-day-old Arabidopsis seedlings were harvested to analyze mRNA transcription levels.
Cloning of JcAP1 cDNA
Total RNA was extracted from the Jatropha flowers using the protocol described by Ding et al. (2008). First-strand cDNA was synthesized using M-MLV-reverse transcriptase according to the manufacturer’s instructions (TAKARA, Dalian, China). The full-length JcAP1 genomic DNA sequence (Sato et al., 2011) (http://www.kazusa.or.jp/jatropha/) was amplified via PCR using the primers XK928 and XK929 (Table S1), which introduced KpnI and SalI recognition sites, respectively. The JcAP1 cDNA containing full-length coding sequence was amplified from flower cDNA with the same primers. The PCR products were subsequently cloned into the pGEM-T vector (Promega Corporation, Madison, Wisconsin, USA) and sequenced. All primers used in this research were listed in Table S1.
Sequence and phylogenetic analyses
JcAP1 amino acid sequence was deduced according to the coding sequence. Related sequences were identified through a BLAST search (http://www.ncbi.nlm.nih.gov/BLAST/). To determine the amino acid identities, the alignment results were subjected to pairwise comparisons using DNAMAN 6.0. A phylogenetic tree based on the protein sequences was constructed with MEGA 5.0 (http://www.megasoftware.net). The amino acid sequences of AP1s, FULs, and CALs were assembled using ClustalX. A neighbor-joining phylogenetic tree was generated with MEGA 5.0 using the Poisson model, with gamma-distributed rates and 1,000 bootstrap replicates.
Plant expression vector construction and Arabidopsis and Jatropha transformation
To construct the 35S:JcAP1 plant overexpression vector, the JcAP1 sequence was excised from the pGEM-T vector (Promega, Corporation, Madison, Wisconsin, USA) using the restriction enzymes KpnI and SalI. Next, JcAP1 was cloned into the pOCA30 vector containing the CaMV 35S promoter. Transformation of Arabidopsis WT and ap1-11 mutant plants with the Agrobacterium strain EHA105 carrying the 35S:JcAP1 construct was performed using the floral dip method (Clough & Bent, 1998). Transformation of Jatropha with the Agrobacterium strain EHA105 carrying the same construct was performed according to the protocol described by Pan, Fu & Xu (2010) and Fu et al. (2015). All of the transgenic plants were confirmed using genomic PCR and RT-PCR.
Expression analysis via quantitative RT-PCR (qRT-PCR)
The roots, stems, mature leaves, inflorescence buds, flower buds, male flowers, female flowers and fruits of mature Jatropha plants and the aboveground tissues of 15 days Arabidopsis seedlings were collected for qRT-RCR detection. Total RNA was extracted from frozen Jatropha tissues as described by Ding et al. (2008). Total RNA was extracted from frozen Arabidopsis tissues using TRIzol reagent (Transgene, China). First-strand cDNA was synthesized with the PrimeScript® RT Reagent Kit with gDNA Eraser (TAKARA, Dalian, China). The cDNA templates of first-strand cDNA were diluted 5-fold with sterilized double-distilled water. qRT-PCR was performed using SYBR® Premix Ex Taq™ II (TAKARA, Dalian, China) on a Roche 480 Real-Time PCR Detection System (Roche, Mannheim, Germany). The primers employed for qRT-PCR are listed in Table S1. qRT-PCR was conducted with three independent biological replicates and three technical replicates for each sample. The data were analyzed using the 2 method described by Livak & Schmittgen (2001). The transcript levels of specific genes were normalized using Jatropha ACTIN1 or Arabidopsis ACTIN2.
Results
Cloning and sequence analysis of JcAP1
A combined reverse transcriptase-polymerase chain reaction (RT-PCR) strategy was used to isolate AP1-like cDNA (JcAP1) from Jatropha. The JcAP1 coding sequence (CDS) (GenBank accession no. KR013222) is comprised of 732 bp and encodes a 243-amino acid protein showing 81%, 79%, 75%, and 71% sequence identity to Vitis vinifera VvAP1 (Calonje et al., 2004), Populus trichocarpa PtAP1 (Tuskan et al., 2006), Coffea arabica CaAP1 (De Oliveira et al., 2014), and AtAP1 (Mandel et al., 1992), respectively.
The genomic sequence of JcAP1 was 4,928 bp and consisted of eight exons and seven introns, which resembles the genomic structure of the Arabidopsis AP1 gene (Mandel et al., 1992). Multiple alignments were performed using the JcAP1 sequence and the sequences of AP1 homologs from other species. The MADS-box domain, K-Box domain, and euAP1 motif were determined (Fig. 1A). Since AP1 is closely related to CAL and FUL, and three genes exhibit high similarity and share redundant functions for floral meristem specification (Bowman et al., 1993), we undertook a phylogenetic analysis of the AP1/FUL MADS-box gene lineage (Fig. 1B), which is also called the SQUA lineage (Krogan & Ashton, 2000). The phylogenetic tree is divided into two clades, AP1 clade and FUL clade. CAL, which appears only in Brassicaceae, is clustered in the AP1 clade (Litt & Irish, 2003). Because CAL and AP1 originated from a recent duplication event <60 million years ago (Alvarez-Buylla, Garcia-Ponce & Garay-Arroyo, 2006). JcAP1 isolated in this study is clustered in the AP1 clade, while the JcFUL is clustered in the FUL clade. Moreover, JcAP1 is more closely related to AP1s of the Euphorbiaceae plants, such as Ricinus communis RcAP1, Manihot esculenta MeAP1 and Plukenetia volubilis PvAP1. In addition, the JcFUL is also closely related to Ricinus communis RcFUL (Fig. 1B). The results indicated that the AP1/FUL MADS-box gene phylogeny follows species phylogeny to interact with other MADS box proteins to confer sepal and petal identity (Mandel et al., 1992).
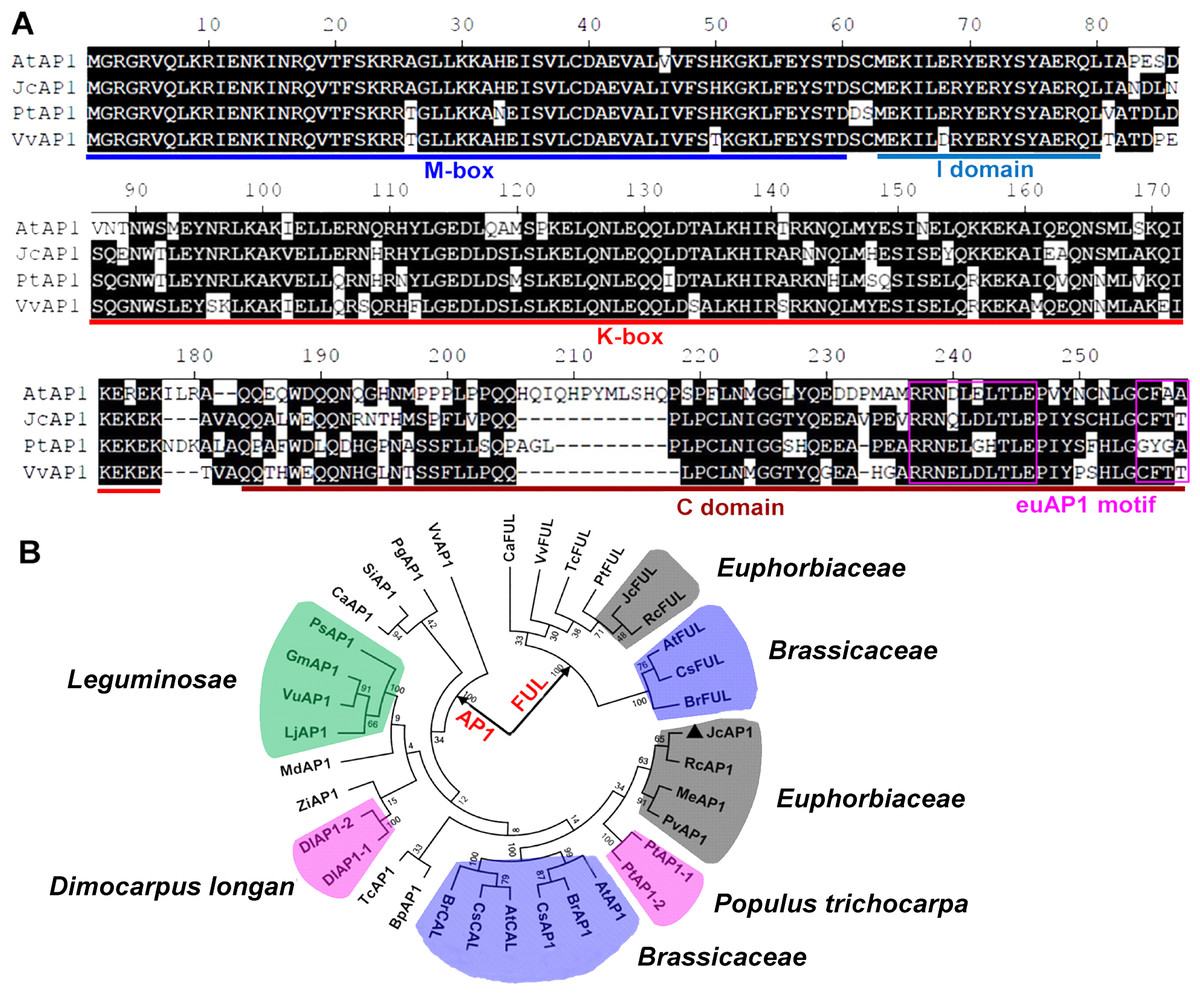
Figure 1: Comparison and phylogenetic analysis of JcAP1 and other AP1 genes.
(A) Sequence alignment of the JcAP1, AtAP1, PtAP1, and VvAP1 amino acid sequences. Similar amino acid residues are shaded in black. Dots denote gaps. Each colored line under the alignment indicates a different domain of an AP1 homolog. “M-box” indicates the highly conserved MADS-box domain; “I domain” indicates the intervening domain; “K-box” indicates the conserved keratin-like domain; “C domain” indicates the C-terminal domain; and “euAP1 motif” indicates the euAP1 motif. (B) Phylogenetic analysis of AP1 homologs from different plant species: AtAP1, Arabidopsis thaliana AP1 ( NP_177074); BpAP1, Betula platyphylla AP1 ( AFV92462); BrAP1, Brassica rapa AP1 ( XP_009105460.1); DlAP1-1, Dimocarpus longan AP1-1 ( AEZ63951); DlAP1-2, Dimocarpus longan AP1-2 ( AGC13077); JcAP1, Jatropha curcas AP1 ( KR013222); PtAP1-1, Populus trichocarpa AP1-1 ( XP_002311353); PtAP1-2, Populus trichocarpa AP1-2 ( XP_002316076); RcAP1, Ricinus communis AP1 ( XP_002514623); CaAP1, Coffea arabica AP1 ( AHW58038); MdAP1, Malus domestica AP1 ( ACD69426); SiAP1, Sesamum indicum AP1 ( AIS82596); VvAP1, Vitis vinifera AP1 ( NP_001268210); PgAP1, Panax ginseng AP1 ( BAK20019); LjAP1, Lotus japonicus AP1 ( AAX13296); VuAP1, Vigna unguiculata AP1 ( BAJ22385); CsAP1, Camelina sativa AP1 ( XP_010415539); TcAP1, Theobroma cacao AP1 ( XP_007045796); PvAP1, Plukenetia volubilis AP1( KU942379); MeAP1, Manihot esculenta AP1(_029935m http://treetfdb.bmep.riken.jp); PsAP1, Pisum sativum AP1 ( AAL66379); GmAP1, Glycine max AP1 ( XP_003531957); ZjAP1, Ziziphus jujube AP1 ( ACG70964); AtCAL, Arabidopsis thaliana CAL ( NP_564243); BrCAL, Brassica rapa CAL ( XP_009109914); CsCAL, Camelina sativa CAL ( XP_010477869); AtFUL, Arabidopsis thaliana FUL ( NP_568929); CaFUL, Coffea arabica FUL ( AHW58040); JcFUL, Jatropha curcas FUL ( KDP31379); PtFUL, Populus trichocarpa FUL ( ABK92820); VvFUL, Vitis vinifera ( XP_002263017); RcFUL, Ricinus communis FUL ( KDP31379); BrFUL, Brassica rapa FUL ( XP_009130138); CsFUL, Camelina sativa FUL ( XP_010443902); TcFUL, Theobroma cacao FUL ( XP_007037634); The phylogeny of these AP1 homologs was determined based on their amino acid sequences using MEGA5 and the neighbor-joining method. Bootstrap values were obtained using 1,000 bootstrap replicates.{kind=link}
Expression pattern of JcAP1 in Jatropha
To assess the expression pattern of JcAP1 in Jatropha, we performed a qRT-PCR analysis using RNA extracted from the roots, stems, mature leaves, inflorescence buds, flower buds, male flowers, female flowers and fruits. The morphologies of different developmental stages of flower were shown in Fig. S3. JcAP1 was expressed in the inflorescence buds, flower buds, flowers, and fruits but showed very low expression in the roots, shoots and leaves. The expression profiles revealed that JcAP1 was highly expressed during the later stages of inflorescence buds (IB3) and early stages of flower buds (FB1). During the development of inflorescences, the expression levels of JcAP1 increased, whereas JcAP1 expression decreased during the development of flower organs. In the reproductive organs, the fruits showed the lowest expression level of JcAP1 (Fig. 2A). JcAP1 was expressed in all floral organs, particularly in the sepals and petals (Fig. 2B).

Figure 2: Expression of JcAP1 in various adult Jatropha organs.
(A) The expression level of JcAP1 in the roots, stems, leaves, inflorescence buds, flower buds, flowers and fruits. (B) The expression level of JcAP1 in different whorls of male and female flowers. The qRT-PCR results were obtained from two independent biological replicates and three technical replicates for each sample. The error bars represent the standard deviation. R, roots; S, stems; L, mature leaves; IB1, inflorescence bud stage 1 (0–5 days, inflorescence buds are visible); IB2, inflorescence bud stage 2 (1 week after IB1); IB3, inflorescence bud stage 3 (1 week after IB2); FB1, flower bud stage 1 (1 week after IB3); FB2, flower bud stage 2 (1 week after FB1, the male flower bud (MFB) and female flower bud (FFB) are identifiable); MF, male flower (1 week after MFB); FF, female flower (1 week after FFB). Fruits (Fr) were harvested 15 days after fertilization. Male sepals (MS), male petals (MP), stamens (St), female sepals (FS), female petals (FP), and carpels (Ca) were harvested 1 or 2 days before the male and female flowers bloomed. The levels of the detected amplicons were normalized using the amplified product of JcACTIN1. The mRNA levels in the IB1 and male petal tissues were used as standards, with a set value of 1.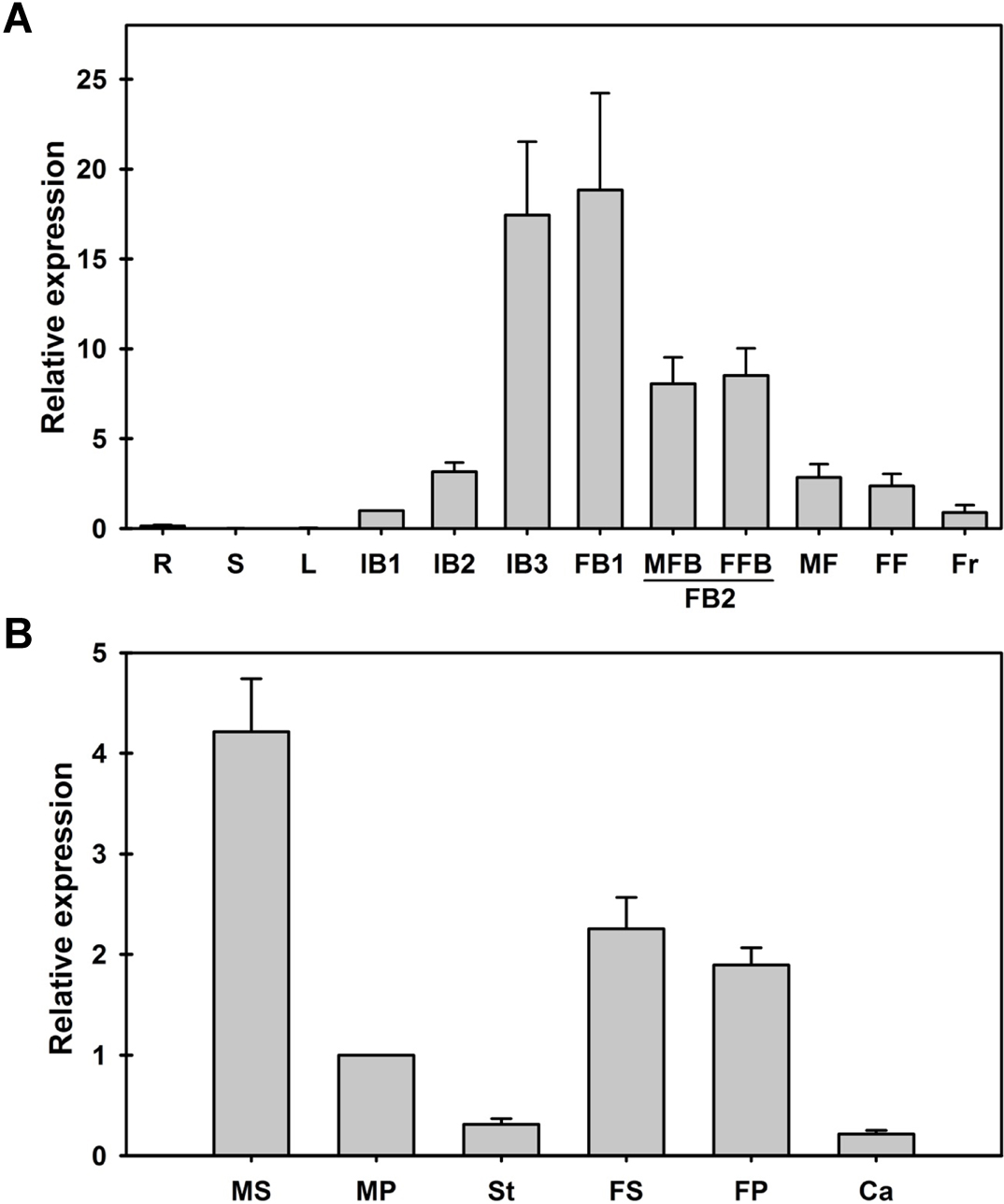{kind=link}
Constitutive overexpression of JcAP1 in Arabidopsis induces early flowering and abnormal flowers
To determine whether JcAP1 is involved in the regulation of flowering time, JcAP1 cDNA driven by the CaMV 35S promoter (Fig. 3A) was transformed into WT Arabidopsis. WT plants under the same growth conditions were used as a control. Transgenic plants were confirmed via qRT-PCR analysis of JcAP1 expression using the aboveground tissues of 15-day-old Arabidopsis seedlings. Thirty-four independent T0 transgenic lines were generated with the 35S:JcAP1 construct. Transgenic plants showed high JcAP1 expression level (Fig. S1A). In the majority of the transgenic lines, bolting occurred notably earlier than in WT plants under both long-day (LD) and short-day (SD) conditions.

Figure 3: Ectopic expression of JcAP1 results in early flowering and abnormal flowers in transgenic Arabidopsis.
(A) Schematic diagram of the T-DNA region of the binary plasmid used in this study. NOS-Pro, nopaline synthase promoter; 35S-Pro, CaMV 35S promoter; 35S-Enh, CaMV 35S enhancer; NOS-Ter, nopaline synthase terminator; CaMV 35S-Ter, 35S terminator; RB, right border of the T-DNA region; and LB, left border of the T-DNA region. (B–D) 15-day-old seedlings grew under LD conditions. (E–G) 60-day-old seedlings grew under SD conditions. (H) A normal flower of a wild-type plant. (I) Abnormal flowers of 35S:JcAP1 transgenic plants from L12. (J) A solitary flower appeared at the rosette and cauline leaf axils in the transgenic plants. Red arrows indicate the solitary rosette flowers; pink arrows indicate that primary shoots immediately terminated with the formation of a solitary flower; the white arrow indicates fruit formation of the abnormal terminal flowers. (B, E, and H) WT; (C, F, I, and J) transgenic L12; (D and G) transgenic Arabidopsis L30. Red bars = 1 cm, yellow bars = 1 mm.{kind=link}
We examined the phenotypes of two independent homozygous transgenic lines (L12 and L30) in the T2 generation. Arabidopsis plants ectopically expressing JcAP1 bolted 6–10 days earlier and produced 4–6 fewer rosette leaves than WT plants under LD conditions (Figs. 3B–3D and Table 1). Under SD conditions, Arabidopsis plants ectopically expressing JcAP1 flowered approximately 1–2 months earlier than WT (Figs. 3E–3G and Table 2). Therefore, the overexpression of JcAP1 in Arabidopsis significantly reduced the vegetative growth time.
| Lines | N | Rosette leaves | Flower bud formation time(day) |
|---|---|---|---|
| WT | 22 | 10.14 ± 0.89 | 20.18 ± 0.73 |
| Line 12 | 20 | 3.95 ± 0.51** | 10.60 ± 1.23** |
| Line 30 | 20 | 5.40 ± 0.99** | 14.25 ± 1.37** |
Notes:
WT plants and two independent JcAP1-overexpressing lines (L12 and L30) grown under LD conditions (16 h light/8 h dark) were subjected to the analysis of rosette leaves and flowering times. N = plant number. The rosette leaves and flowering times are presented as the mean ± standard deviation.
| Lines | N | Rosette leaves | Flower bud formation time(day) |
|---|---|---|---|
| WT | 22 | 60.41 ± 3.95 | 104.00 ± 5.83 |
| Line 12 | 25 | 14.68 ± 1.44** | 33.72 ± 3.06** |
| Line 30 | 27 | 31.63 ± 2.50** | 58.33 ± 3.95** |
Notes:
WT plants and two independent JcAP1-overexpressing lines (L12 and L30) grown under SD growing conditions (8 h light/16 h dark) were subjected to the analysis of rosette leaves and flowering times. N = plant number. The rosette leaves and flowering times are presented as the mean ± standard deviation.
In contrast to WT plants, the primary shoots of the transgenic plants were converted into compound terminal flowers consisting of two or three pistils surrounded by an abnormal number of sepals, petals, and stamens (Fig. 3I). Furthermore, the secondary shoots produced in cauline and rosette leaf axils were converted into solitary flowers. In extreme transgenic plants, all branches and inflorescences were replaced by solitary flowers (Fig. 3J).
Further analysis indicated that the promotion of flowering and abnormal terminal flowers in 35S:JcAP1 transgenic Arabidopsis was correlated with a significant up-regulation of the floral meristem identity genes AtLFY, AtFUL, AtAP1 and AtCAL and the floral organ identity genes AtAGAMOUS (AtAG), AtAP3 and AtSEPs (AtSEP1, AtSEP2, AtSEP3) (Fig. S1). The expression levels of these genes were highest in transgenic plants L12, whereas the AtTFL1 expression level was slightly down-regulated (Fig. S1). Thus, L12 also showed the most obvious changes in phenotype of extremely early flowering and solitary and terminal flowers (Fig. 3). Thus, the phenotypes of the early-flowering and abnormal terminal flowers produced due to the ectopic expression of JcAP1 in transgenic Arabidopsis were similar to those resulting from AtAP1 overexpression (Mandel & Yanofsky, 1995).
Constitutive overexpression of JcAP1 in ap1-11 mutant Arabidopsis induces early flowering and partially complements the phenotype
To further determine whether JcAP1 can function similarly to AtAP1, the 35S:JcAP1 construct (Fig. 3A) was transformed into Arabidopsis ap1-11 mutant plants. Eight independent T0 transgenic lines were generated and confirmed through qRT-PCR analysis of JcAP1 expression using RNA from aboveground tissues of 15-day-old Arabidopsis seedlings. WT and ap1-11 mutants under the same growth conditions were used as controls. Most of the transgenic lines bolted earlier than the WT and ap1-11 mutant plants under inductive LD conditions. The ap1 mutants didn’t exhibit significantly later flowering than WT (Table 3), which is because three homologous genes AP1, CAL, and FUL play redundant roles in control of flowering time. Each single mutant of ap1, cal or ful exhibited only slightly late flowering, whereas the triple mutant exhibited significantly late flowering (Ferrándiz et al., 2000).
| Lines | N | Rosette leaves | Flower bud formation time(day) |
|---|---|---|---|
| WT | 25 | 10.54 ± 0.92 | 20.18 ± 0.73 |
| ap1-11 | 20 | 11.02 ± 0.87 | 20.80 ± 1.23 |
| Line C2 | 15 | 8.40 ± 0.99** | 14.05 ± 1.45** |
| Line C5 | 15 | 3.95 ± 0.51** | 9.80 ± 1.18** |
Notes:
WT plants, the ap1-11 mutant, and two independent JcAP1-overexpressing lines (C2 and C5) grown under LD growing conditions (16 h light/8 h dark) were subjected to the analysis of rosette leaves and flowering times. N = plant number. The rosette leaves and flowering times are presented as the mean ± standard deviation.
To examine phenotypes, we selected two independent homozygous transgenic lines (C2 and C5) in the T2 generation that showed high JcAP1 expression levels: line C2 and line C5 (Fig. 4I). Complementary transgenic lines C2 and C5 bolted 6–11 days earlier and produced 2–6 fewer rosette leaves than the controls under LD conditions (Figs. 4A–4D and Table 3). In the extreme complementary transgenic line C5, solitary flowers appeared on the axils of rosette and cauline leaves and terminal flowers appeared on the primary shoots (Fig. 4D). The ap1-11 mutant flowers lacked petals, and new secondary floral buds developed on the axils of the bract-like organs present in the first whorl (Fig. 4F). The transgenic mutant C2 and C5 lines restored the development of sepals and petals, and axillary flowers were rarely seen at the bracts (Figs. 4G and 4H). Overexpression of JcAP1 in ap1 mutant leading to early flowering is because we used a strong promoter, the 35S promoter, which drives JcAP1 constitutively expressing. Similar results were found in Arabidopsis ap1 mutant overexpressing the chrysanthemum and lily AP1-like genes (Chen, Lin & Yang, 2008; Shchennikova et al., 2004). Further analysis indicated that the promotion of flowering in the 35S:JcAP1 transgenic Arabidopsis mutant was correlated with a significant up-regulation of the flower meristem identity genes AtLFY, AtSOC1 and floral organ identity genes AtSEPs (Figs. 4J and 4K).
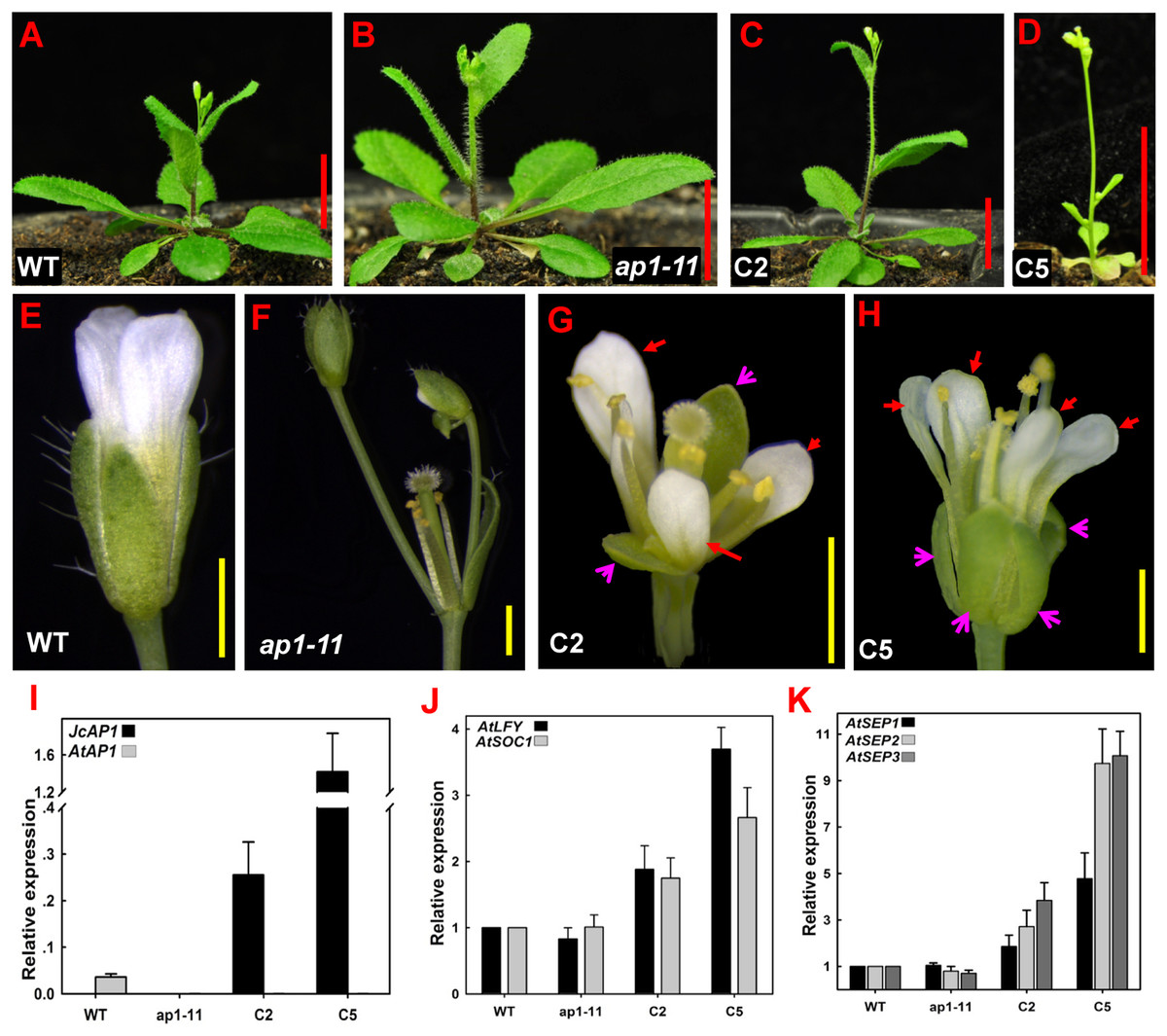
Figure 4: 35S:JcAP1 transgenic Arabidopsis recovers the phenotype of the ap1-11 mutant and causes early flowering.
(A–C) WT, ap1-11, and 35S:JcAP1 complementary ap1-11 plant line 2 (C2) growth under LD conditions at 30 days after germination. (D) 35S:JcAP1 complementary ap1-11 plant line 5 (C5) growth under LD conditions at 15 days after germination. (E–H) Florets of WT (E), ap1-11 (F), and 35S:JcAP1 complementary ap1-11 in which sepals and petals were rescued (G and H). The red arrow indicates that the petals appeared in complementary transgenic plants, and the pink arrow indicates that the sepals appeared in complementary transgenic plants. (I–K) qRT-PCR analysis of JcAP1 and other flowering-related genes including Arabidopsis APETALA1 (AtAP1), LEAFY (AtLFY), SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1 (AtSOC1), SEPALLATA 1, 2 and 3 (AtSEP1, AtSEP2, and AtSEP3) in WT, ap1-11 and transgenic Arabidopsis (C2, C5). The levels of the detected amplicons were normalized using the amplified products of AtACTIN2. The mRNA level in WT was set as the standard, with a value of 1. Red bars = 1 cm, yellow bars = 1 mm.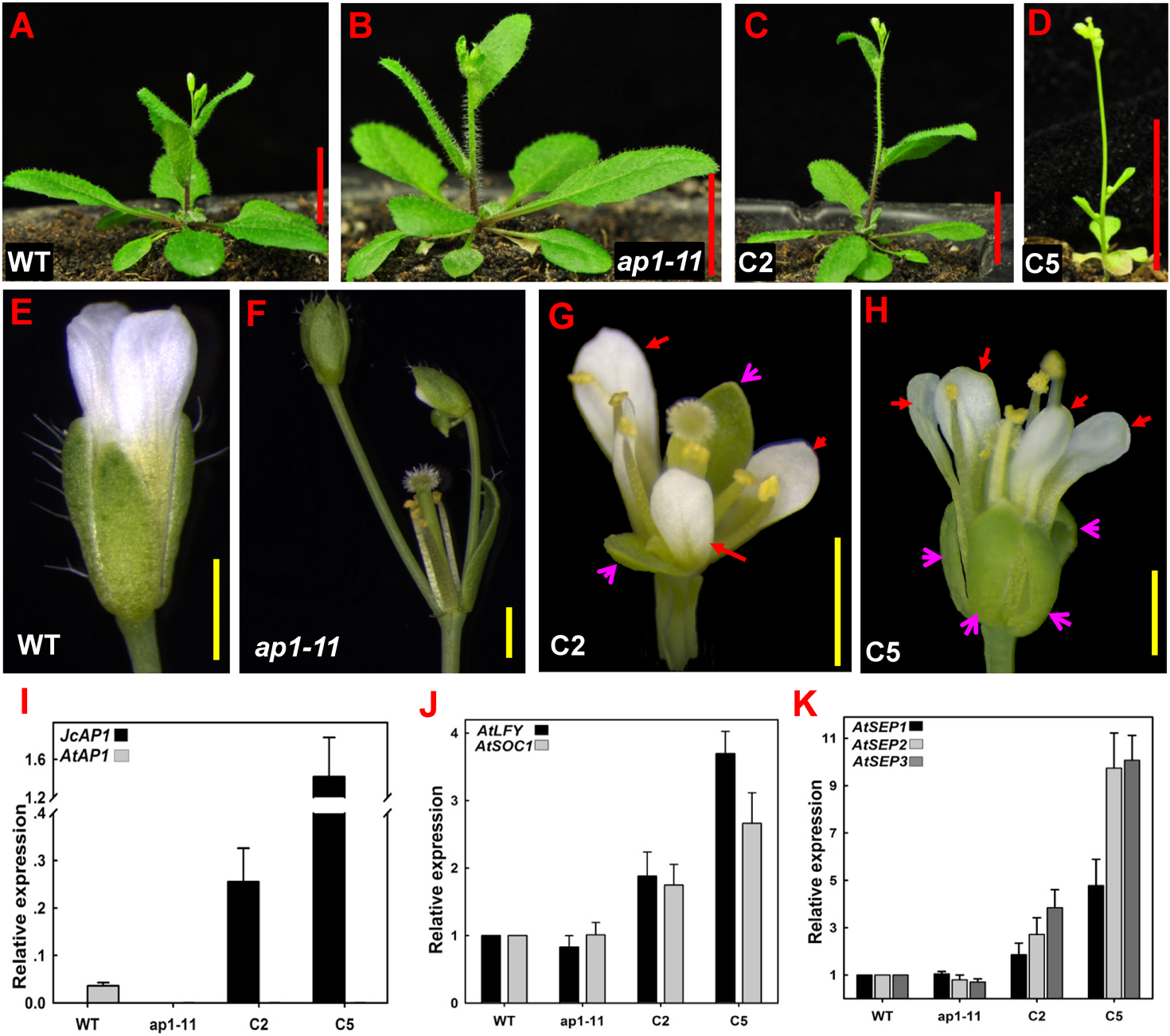{kind=link}
These results demonstrate that the constitutive expression of JcAP1 complements the defect in floral organ development observed in the ap1-11 mutant; thus, JcAP1 functions as an A-class gene in transgenic Arabidopsis.
Overexpression of JcAP1 in Jatropha did not cause early flowering
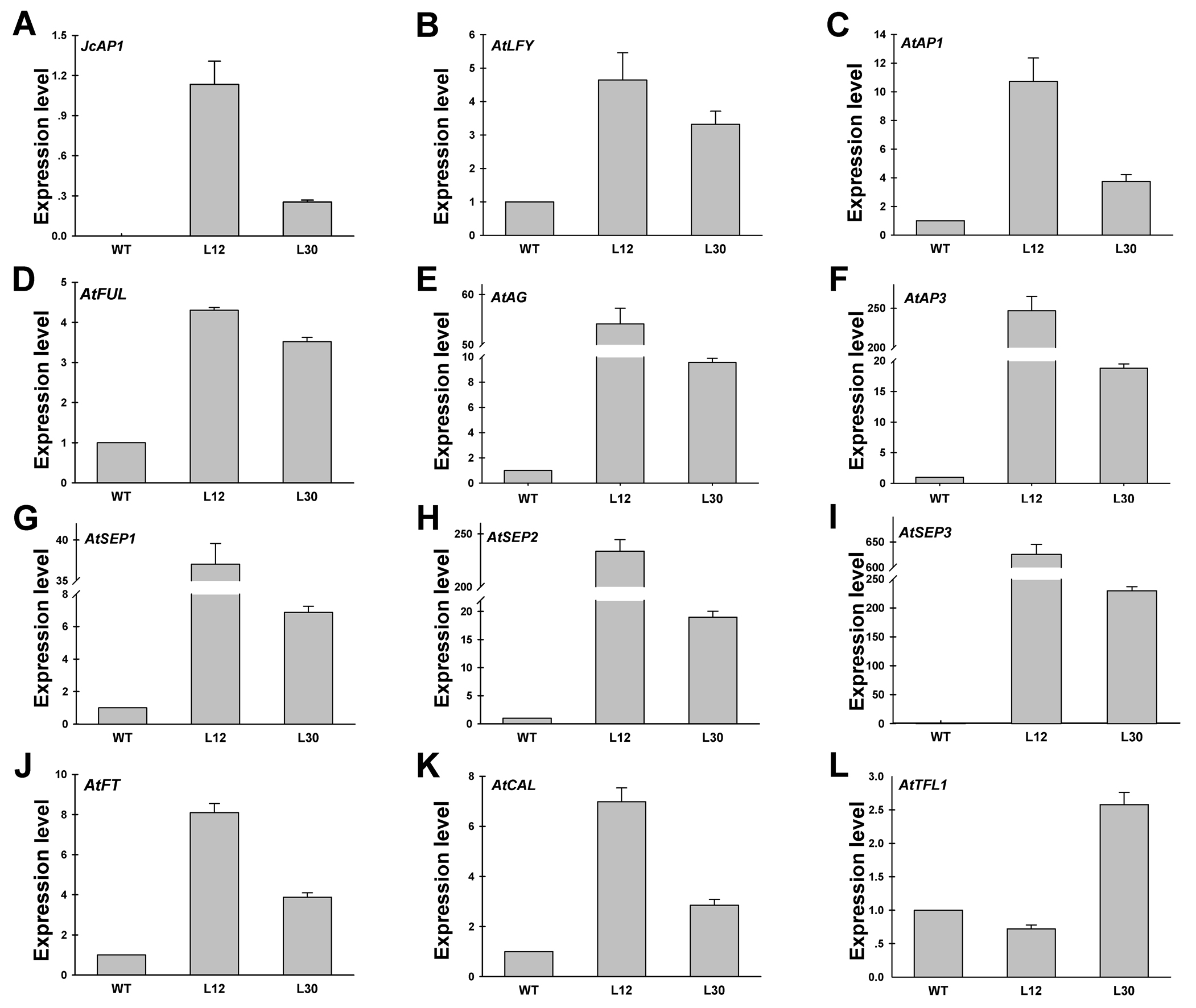
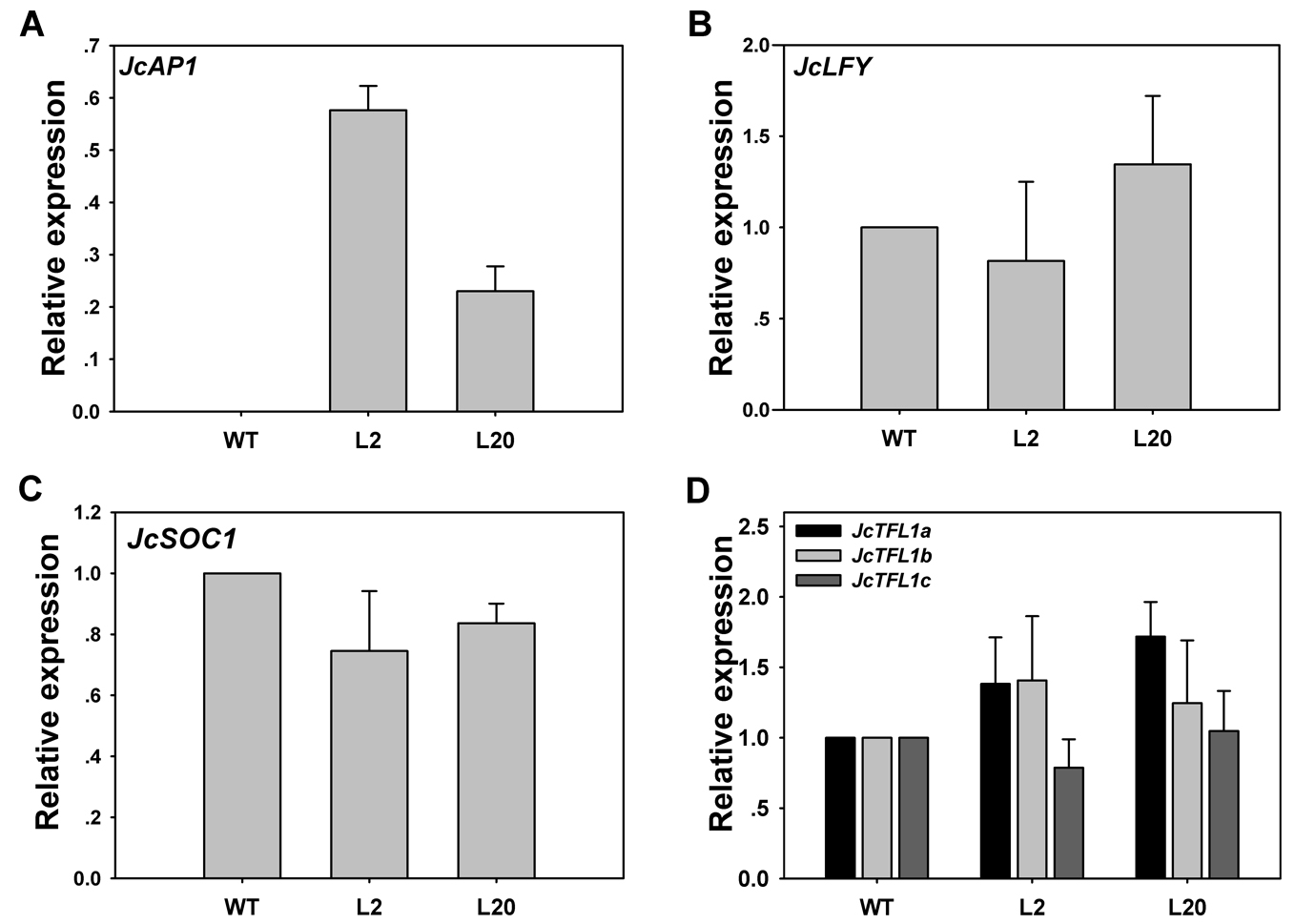
Transgenic analysis performed in Arabidopsis suggested that JcAP1 might act as a floral identity gene in Jatropha. To test this hypothesis, we generated transgenic Jatropha with the 35S:JcAP1 construct (Fig. 3A) as previously described (Pan, Fu & Xu, 2010). Non-transgenic plants were used as a control. Fifty-five independent transgenic lines were confirmed via PCR using genomic DNA isolated from leaves of 2-month-old plantlets. And the partial results are shown in Fig. 5A. Next, JcAP1 expression levels in fourteen PCR-positive lines were examined through qRT-PCR using RNA extracted from young leaves of 2-month-old plantlets (Fig. 5B). To our surprise, all of these transgenic Jatropha lacked an early-flowering phenotype (Figs. 6A and 6B). When regenerated plantlets were grown in the field for 4 months, flower buds emerged in both transgenic and control plants (Figs. 6C–6H). We chose L2 and L20, which exhibited high and intermediate expression levels, respectively (Fig. 5B), to further analyze the expression levels of several floral identity-related genes in the shoot apices of 6-month-old plantlets. The results showed that the transcript levels of JcLFY, JcSOC1 and JcTFL1s (Fig. S2) were not significantly altered in both transgenic lines. The 35S promoter was highly active in the Jatropha inflorescence buds (Tao et al., 2015), but the inflorescence structure (Figs. 6D, 6F and 6H) and floral organ pattern (Fig. 6I) were not obviously different. These results indicate that JcAP1 may be inadequate to promote flowering and floral organ development by itself in Jatropha.
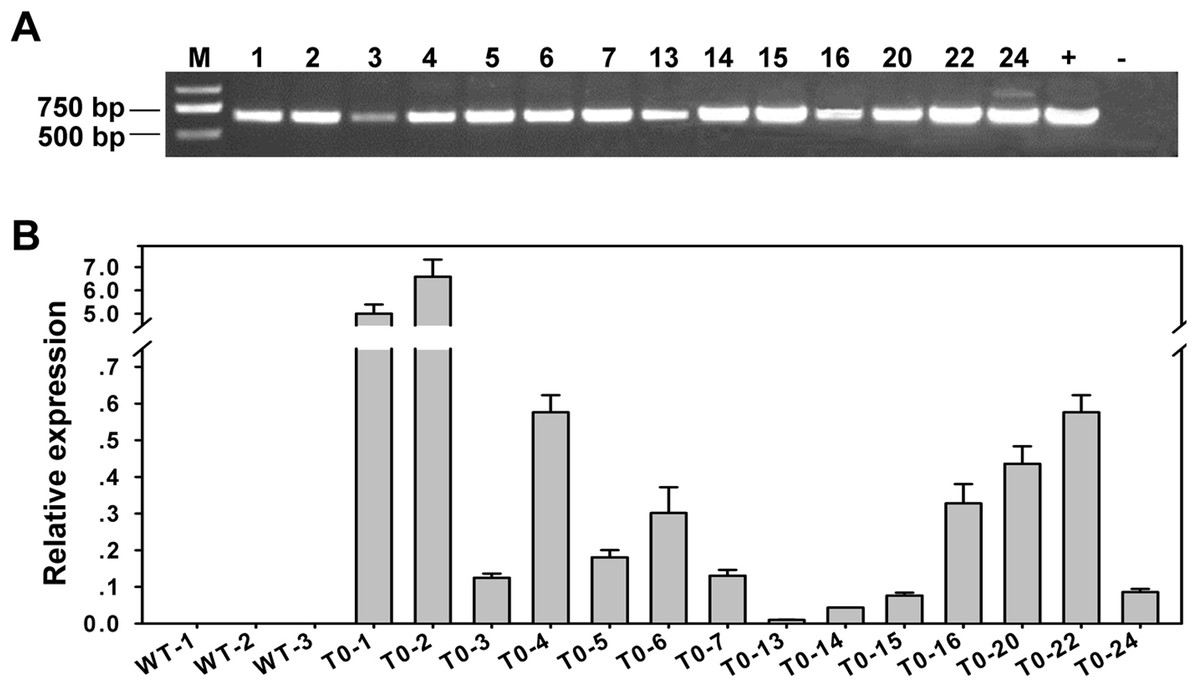
Figure 5: Analysis of the transgenic Jatropha plants.
(A) Amplification of the 600-bp fragment containing partial sequences of the 35S promoter and JcAP1 cDNA. Lanes: M, Trans 2 Kb DNA ladder; +, positive control (plasmid); −, negative control (wild type); and 14 regenerated transgenic Jatropha lines. (B) Quantitative RT-PCR analysis of 3 WT and 14 independent transgenic plants (L1, L2, L3, L4, L5, L6, L7, L13, L14, L15, L16, L20, L22, L24). Two transgenic plants, L2 and L20, showing high and intermediate expression levels, respectively, were chosen for further analysis. RNA was extracted from young leaves of 2-month-old plantlets. The transcript levels were normalized using the JcACTIN1 gene as a reference.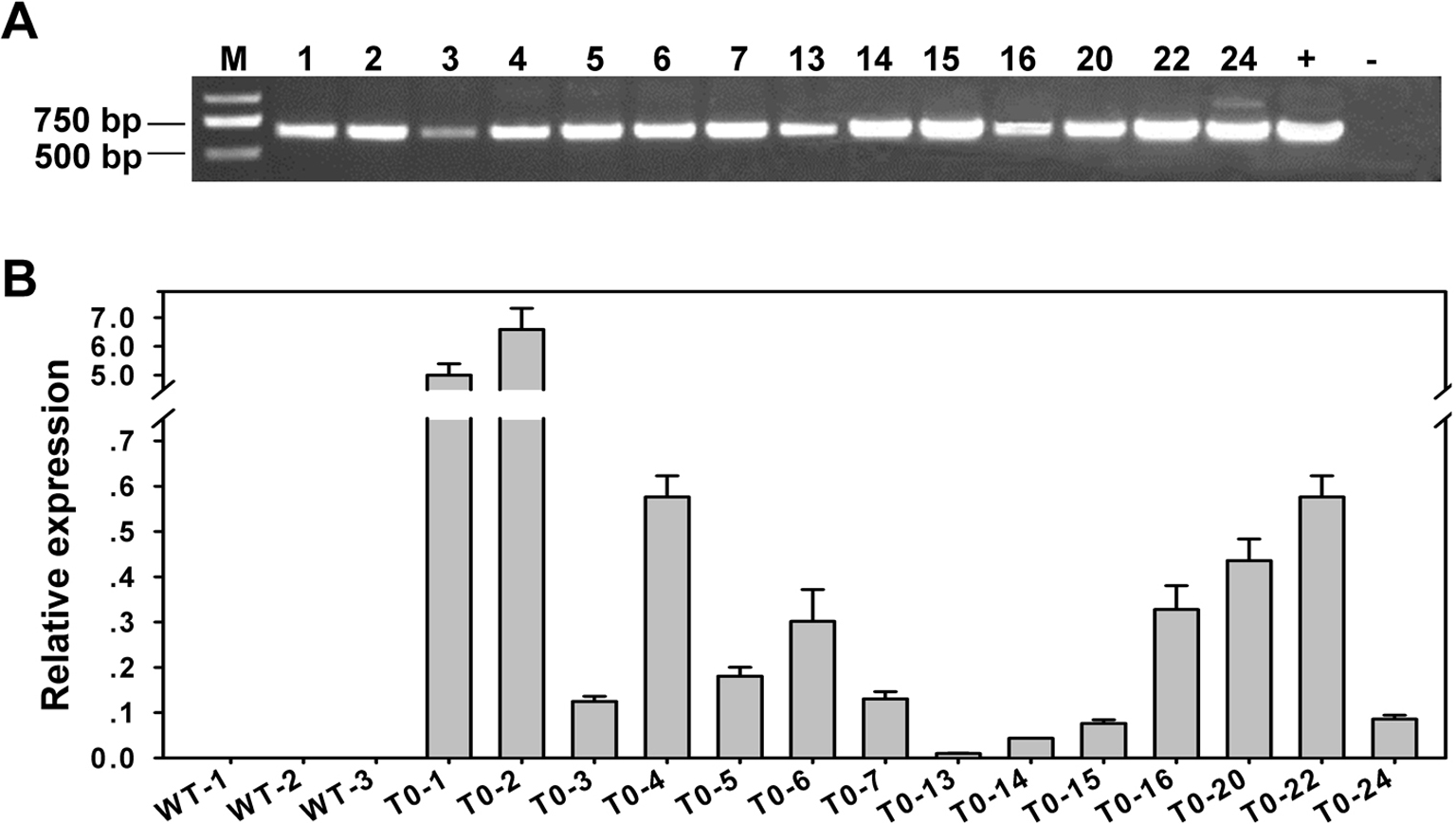{kind=link}

Figure 6: The flowering time of 35S:JcAP1 transgenic Jatropha in the field.
(A) WT plant grown in a pot for 2 months, at the vegetative growth stage; (B) 35S:JcAP1 transgenic Jatropha grown in a pot for 2 months, at the vegetative growth stage. (C) WT plants grown in the field for 4 months, at the anthesis stage; (D) The inflorescence of WT Jatropha in the field. (E, G) 35S:JcAP1 transgenic Jatropha L2 and L20 plants grown in the field for 4 months, at the anthesis stage; (F, H) The inflorescence of transgenic Jatropha in the field. (I) The flowers of WT and 35S:JcAP1 transgenic Jatropha L2 and L20 plants. Red bars = 1 cm, white bars = 10 cm, yellow bars = 50 cm.{kind=link}
Discussion
Recently, Jatropha has garnered significant attention as a potential oilseed plant for the production of renewable biofuel. Despite the interest in this woody oil plant, relatively little is known regarding the molecular biology of this species compared with more established oilseed crops, such as rapeseed (Handa, 2003; Kresovich et al., 1995) and castor bean (Chan et al., 2010).
Amino acid sequence similarity, protein structures, and phylogenetic analysis suggested that JcAP1 exhibits a similar function to other AP1 homologues. In Arabidopsis, AP1 functions redundantly with FUL in specifying floral meristem identity (Bowman et al., 1993). However, in the phylogenetic tree, AP1 and FUL clustered in the separate clades indicated the functional divergence between the two paralogs. Except for floral meristem determination, FUL is required for proper fruit and leaf development in Arabidopsis (Gu et al., 1998), and FUL prevents normal senescence and winter dormancy in woody Populus tremula L (Hoenicka et al., 2008). JcAP1 was clustered in the AP1 clade suggesting this gene may functions similarly as other AP1 genes in the regulation of flowering and flower organ development.
Quantitative RT-PCR results showed that JcAP1 transcripts were highly expressed in inflorescence buds, flower buds, sepals and petals (Fig. 2). This expression pattern is consistent with that of AtAP1 in Arabidopsis (Mandel et al., 1992). The expression profiles revealed that the highest JcAP1 transcript levels occurred in the earlier stages of male and female flower buds (Fig. 2A), implying that JcAP1 may play a role in maintaining the normal development of flower patterns (Collaudin, 2012). In addition, the high JcAP1 expression levels observed in sepals and petals (Fig. 2B) suggested that JcAP1 may be involved in sepal and petal development. In Arabidopsis, the function of AP1 in the regulation of sepal and petal development was revealed using an ap1 mutant (Mandel et al., 1992). The AP1 gene was identified as a member of the floral meristem identity genes, which largely acted downstream of the floral integrators FT, LFY, and SOC1 (Bowman et al., 1993; Liljegren et al., 1999; Liu et al., 2013; Riechmann, Krizek & Meyerowitz, 1996). In this study, we showed that overexpression of JcAP1 in Arabidopsis resulted in reduced vegetative growth, early flowering and the formation of terminal and solitary flowers (Fig. 3; Tables 1, and 2). These findings are similar to the phenotypic changes caused by constitutive expression of AP1 homologs in Arabidopsis (Chi et al., 2011; Kotoda et al., 2002; Liljegren et al., 1999; Liu et al., 2013; Mandel & Yanofsky, 1995; Sun et al., 2014; Wang et al., 2013; Weigel & Nilsson, 1995; Winterhagen et al., 2013). The production of terminal and solitary flowers in AP1- overexpressing plants is due to the inhibition of TFL1 expression induced by AP1 (Blazquez et al., 2006). In Arabidopsis, young seedlings showed weak TFL1 expression; the TFL1 expression increased after 8 days and young inflorescences showed the strongest TFL1 expression (Bradley et al., 1997). In previous research, TFL1 down-regulated directly by AP1 was confirmed by the CHIP-Seq analysis (Kaufmann et al., 2010). Compared with WT plants, the TFL1 expression was not detected in primary shoot apices and secondary meristems in AP1 overexpressing plants (Liljegren et al., 1999). Similarly, in our research, when JcAP1was overexpressed in Arabidopsis, the expression of TFL1 was also suppressed. The aboveground tissues of 15-day-old plants were used for detecting the TFL1 expression in our study. The inflorescence buds had appeared in JcAP1 overexpressing plants while WT plants were still in vegetative growth. According to the TFL1 expression pattern in Arabidopsis, in which the inflorescences showed the highest expression level, TFL1 expression in transgenic plant was supposed to be higher than in WT plants. However, the expression level of TFL1 was decreased in the JcAP1 highly expressing line (L12) (Fig. S1L). This result indicated that the increase of JcAP1 expression repressed the TFL1 expression. Consistently, when JcAP1 was lowly expressed (L30), the expression level of TFL1 was markedly increased (Fig. S1L). In Jatropha, nevertheless, the expression of three JcTFL1s (JcTFL1a, JcTFL1b and JcTFL1c) were not significantly altered in JcAP1 overexpressing plants (Fig. S2D). Thus, we supposed that JcTFL1 was indirectly relative to JcAP1.
Overexpression of JcAP1 in the Arabidopsis ap1-11 mutant resulted in early flowering, restoration of sepal and petal development, and repression of secondary flower formation in the bract axils (Figs. 5I–5K). These results are consistent with the phenotypic changes observed following the ectopic expression of Chrysanthemum and lily AP1-like genes in the Arabidopsis ap1 mutant (Chen, Lin & Yang, 2008; Shchennikova et al., 2004). These findings imply that JcAP1 acts as a functional homolog of AP1 in Arabidopsis.
In contrast to JcAP1-overexpressing Arabidopsis, JcAP1-overexpressing Jatropha did not exhibit early flowering (Fig. 6). But the expression profile showed that JcAP1 was predominantly expressed in inflorescence buds and flower buds (Fig. 2A). It indicates that JcAP1 may be insufficient to regulate flowering time or floral organ development by itself in Jatropha. Similar to our findings, overexpression of the Populus ortholog of APETALA1 in Arabidopsis led to early flowering whereas it failed to promote flowering in hybrid aspen (Azeez et al., 2014). In addition new functions of AP1 homologs have been identified in several species. The AP1 homolog of grapevine (VAP1) is involved in the formation of tendrils (Calonje et al., 2004). Wheat AP1 (WAP1) has no known role in flower development but is required for vernalization and phase transition (Danyluk et al., 2003; Handa, 2003; Trevaskis et al., 2003). In tomato plants, AP1/FUL MADS box genes are involved in tomato leaf development (Burko et al., 2013).
The molecular mechanisms controlling flowering in perennials have not been studied as extensively as those of annual plants (Albani & Coupland, 2010). It seems more complex in perennial plants in that a well-known identified flowering gene is unable to control the flowering time. For example, overexpression of LFY failed to cause early flowering in apple (Flachowsky et al., 2010) and hybrid populous (Rottmann et al., 2000). Overexpression of miR172, which caused extremely early flowering in Arabidopsis (Aukerman & Sakai, 2003), was unable to promote flowering in the perennial plant Cardamine flexuosa unless it was treated with vernalization simultaneously (Zhou et al., 2013). In this study, the results also suggest that JcAP1 itself may not be sufficient to promote flowering in Jatropha; perhaps it needs to be associated with other factors to function in these processes. Other important floral identity genes or environment factors need to be characterized in Jatropha in the future study.
Supplemental Information
Quantitative RT-PCR analysis of JcAP1 and other flower-related genes in WT and transgenic Arabidopsis
(A) The expression level of JcAP1 in WT and transgenic Arabidopsis L12 L30 plants; JcAP1 expression was not detected in WT; transcript levels were not normalized. (B–L) The expression levels of AtLFY, AtAP1, AtFUL, AtAG, AtAP3, AtSEP1, AtSEP2, AtSEP3, AtFT, AtCAL, and AtTFL1, respectively. RNA sample extracted from apex and rosette leaves of 35S: JcAP1 transgenic and WT plants cultured for 15 days in a pot. Transcript levels were normalized using the AtACTIN2 gene as a reference. The mRNA level in WT was set as the standard, with a value of 1.
{kind=link}
Quantitative RT-PCR analysis of JcAP1 and flower-related genes in WT and 35S:JcAP1 transgenic Jatropha
The expression levels of JcAP1, JcLFY, JcSOC1, and JcTFL1s were detected in shoot apices of 6-month-old plantlets of WT and transgenic Jatropha. The qRT-PCR results were obtained using two independent biological replicates and three technical replicates for each RNA sample extracted from the apex of the 35S: JcAP1 transgenic and WT shoots. Transcript levels were normalized using the JcACTIN1 gene as a reference. The mRNA level in WT was set as the standard, with a value of 1.
{kind=link}
Flower morphological characteristics of Jatropha in different developmental stages
(A) Inflorescence bud stage 1 (IB1): 0–5 days, inflorescence buds are visible; (B) inflorescence bud stage 2 (IB2): 1 week after IB1; (C) inflorescence bud stage 3 (IB3): 1 week after IB2; (D) flower bud stage 1 (FB1): 1 week after IB3; (E) flower bud stage 2 (FB2): male flower buds (MFB) and female flower buds (FFB) are identifiable one week after FB1; (F): male and female flower stage: male flowers (MF) and female flowers (FF) bloomed one week after FB2. In (E) and (F), red arrows indicate FFBs and FFs, respectively; and pink arrows indicate MFBs and MFs, respectively. Bars = 1 cm.
{kind=link}
Raw data
Transgenic arabidopsis flowering time and gene expression levels.