
The role of Rnf in ion gradient formation in Desulfovibrio alaskensis
- Published
- Accepted
- Received
- Academic Editor
- Chandan Goswami
- Subject Areas
- Biochemistry, Microbiology
- Keywords
- Desulfovibrio, Rnf, Proton motive force, ionophore, Energetics
- Copyright
- © 2016 Wang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. The role of Rnf in ion gradient formation in Desulfovibrio alaskensis. PeerJ 4:e1919 https://doi.org/10.7717/peerj.1919
Abstract
Rnf is a membrane protein complex that has been shown to be important in energy conservation. Here, Desulfovibrio alaskensis G20 and Rnf mutants of G20 were grown with different electron donor and acceptor combinations to determine the importance of Rnf in energy conservation and the type of ion gradient generated. The addition of the protonophore TCS strongly inhibited lactate-sulfate dependent growth whereas the sodium ionophore ETH2120 had no effect, indicating a role for the proton gradient during growth. Mutants in rnfA and rnfD were more sensitive to the protonophore at 5 µM than the parental strain, suggesting the importance of Rnf in the generation of a proton gradient. The electrical potential (ΔΨ), ΔpH and proton motive force were lower in the rnfA mutant than in the parental strain of D.alaskensis G20. These results provide evidence that the Rnf complex in D. alaskensis functions as a primary proton pump whose activity is important for growth.
The Rnf complex was first discovered in Rhodobacter capsulatus and is thought to be transcribed in one cluster of seven genes rnfABCDGEH (Jouanneau et al., 1998). Evidence that depleting one subunit could destabilize others indicated the formation of a complex (Kumagai et al., 1997) which was later shown to be a protein complex involved in the Rhodobacter nitrogen fixation process (Schmehl et al., 1993). The Rnf complex has been differentiated into three major groups based on gene organization (Biegel et al., 2011). The first group found in R. capsulatus, Pseudomonas stutzeri (Yan et al., 2008) and Azotobacter vinelandii (Curatti et al., 2005) with rnfABCDGE. Another group with the gene order rnfCDGEAB is found in A.woodii (Biegel, Schmidt & Müller, 2009) and Clostridium kluyveri (Seedorf et al., 2008). Lastly rnfBCDGEA is found in Chlorobium limicola, Bacteroides vulgatus and Prosthecochloris aestuarii (Biegel et al., 2011).
Rnf complex has been shown to be a novel type of ferredoxin-dependent enzyme, catalyzing the oxidation–reduction reaction between reduced ferredoxin and NAD+ (Biegel, Schmidt & Müller, 2009; Boiangiu et al., 2005). Because of the high sequence similarity to the Na+-translocating NADH:ubiquinone oxidoreductase (Nqr) (Kumagai et al., 1997), the Rnf complex in R. capsulatus was suggested to be involved in ion translocation using the energy from the exergonic reduction of NAD+ coupled to the oxidation of reduced ferredoxin (Müller et al., 2008). Based on the above similarity, Rnf was originally proposed to be a sodium pump (Biegel, Schmidt & Müller, 2009). This theory was strengthened after the Rnf complex in Acetobacterium woodii was confirmed to generate a Na+ gradient when carrying out the redox reaction between reduced ferredoxin and NAD+ (Biegel & Müller, 2010). However, the Rnf complex was suggested to be a proton-translocating ferredoxin:NAD+ oxidoreductase in Clostridium ljungdahlii (Kopke et al., 2010) and was more recently shown to produce a proton gradient (Tremblay et al., 2013). It therefore appears that Rnf may pump different ions in different bacteria (Hess et al., 2016).
Desulfovibrio alaskensis G20 encodes the Rnf complex with a unique gene arrangement (rnfCDGEABF); however, a similar Rnf complex is present in most sulfate-reducing bacteria for which genome sequences are available (Pereira et al., 2011). Directly preceding the operon is a decaheme cyctochrome (dhcA) which is also present in many but not all sulfate reducing bacteria. This decaheme cytochrome belongs to cytochrome c3 family (Pereira et al., 2011). The final gene in the operon, rnfF is most similar to apbE, a membrane associated lipoprotein (Beck & Downs, 1999).
Recently, it was reported that mutants in Rnf in D. alaskensis are unable to grow using H2 or during syntrophic growth conditions with Syntrophus aciditrophicus or Syntrophomonas wolfeii (Krumholz et al., 2015; Price et al., 2014). By investigating gene expression levels of D.alaskensis G20 grown in pure culture or under syntrophic conditions (Krumholz et al., 2015), it was suggested that both formate and H2 can act as electron shuttles during syntrophic growth. Those experiments also showed that expression of rnf genes are upregulated under syntrophic conditions and during H2 dependent growth confirming the role of the Rnf complex in H2 metabolism.
In this experiment, mutants of RnfA (Dde_0585) and RnfD (Dde_0582) were studied and compared with the G20 parent strain to determine whether Rnf in D. alaskensis is involved in generation of a proton motive force (PMF). These two mutants of Desulfovibro alaskensis G20 were obtained by using mini Tn10 transposon (Groh et al., 2005) and identified individually during syntrophic growth with Syntrophomonas wolfeii (Krumholz et al., 2015).
Materials and Methods
Growth of cultures
Desulfovibrio alaskensis G20 and rnf mutants were routinely cultured anaerobically (N2-CO2, 80:20) in basal medium with lactate-sulfate (50 mM each) as the electron donor and electron acceptor, respectively and containing 0.1% yeast extract (Krumholz et al., 2015). The medium was prepared under a headspace of N2∕CO2(80/20, vol/vol) with sodium bicarbonate (3.5 g/L) added as a buffer. Before inoculation, the medium was reduced with 0.025% each cysteine and sulfide. Kanamycin (1050 µg/mL) was added to the mutant cultures. Inoculum consisted of 0.2 ml transferred into 10 ml of media or the same ratio for large volume cultures. Cultures were grown in the incubator at 37 °C and growth of triplicate cultures was measured by optical density at 600 nm. The stock cultures were frozen in 20% glycerol at −20°. Growth of syntrophic cocultures was as previously described (Krumholz et al., 2015).
Growth was tested with a variety of other electron donors and acceptors: lactate (50 mM), pyruvate (25 mM), ethanol (10 mM) and formate (50 mM). Sulfate was used at 50 mM with lactate and 25 mM with other electron donors and sulfite was added at 10 mM.
The protonophore 3,3,4,5-tetrachlorosalicylanilide (TCS, 5 µM or 20 µM), or the sodium-specific ionophore N,N,N′,N′-Tetracyclohexyl-1,2-phenylenedioxydiacetamide (ETH2120, 20 µM) were added to media to test their effects on growth.
Transcriptional analysis of rnf genes
Total RNA was extracted from the parent strain, rnfA, and rnfD mutants using the Qiagen RNeasy kit as previously described (Krumholz et al., 2015). Purity and adequate yield was confirmed with a diode-array spectrophotometer. First-strand cDNA synthesis was performed using the Fermentas Revertaid kit with gene-specific primers covering the entirety of the rnf mutants’ operons. Primers were designed to span the gaps between each gene in the operon (Table 1). PCR was performed with the parent strain and mutants with each primer set followed by agarose gel analysis to determine whether all genes in the operon were expressed and whether the genes were on the same transcript.
| Gap | Forward primer | Reverse primer |
|---|---|---|
| Gap CC (cytC/rnfC) | GGCATTCACGGCCTGTGCCT | CTTGTCGCCGGGGTGATCGG |
| Gap CD (rnfC/rnfD) | CCTGCTGGGACGCTACAGCG | ATGCCGTGTATGGTGCGCCC |
| Gap DG (rnfD/rnfG) | GGCAAGGCCGCCATGGTCAT | TGTACTCGGCCCCCGTGGAG |
| Gap GE (rnfG/rnfE) | ATCGGCTGCATGGTTGCCGT | ATTGGCGGACTTGGTGACCGC |
| Gap EA (rnfE/rnfA) | GGGCTTGTTCTGGGCGCCAT | CCGCCCATGCCCAGACTGAC |
| Gap BE (rnfB/rnfF) | AAAGCCTGCCTCGCGTTCGG | AAGCCCCAGCAGACCGCATG |
Quantitative PCR analysis
Triplicate cultures of the parent strain and rnfA and rnfD mutants were grown to mid-log phase (OD600 0.3–0.45) and harvested by centrifugation at 5,000 × g for 5 min at 4 °C. The pellet was homogenized in RNAprotect Bacteria reagent (Qiagen) for 5 min at room temperature to prevent degradation of RNA transcripts. Excess liquid was removed by centrifugation and cell pellets were resuspended in 200 ul of TE buffer containing 1 mg/ml lysozyme. Total RNA was extracted using the RNeasy mini kit (Qiagen). RNA was then treated with the Ambion Turbo DNA –free kit (Thermo) to eliminate genomic DNA contamination. RNA was quantified spectrophotometrically. cDNAs were synthesized using the First Stand cDNA synthesis kit (Fermentas) as described in the manual. Control PCR reactions were done to ensure there was no genomic DNA or non-specific amplification.
Quantitative PCR, was done in triplicate from each culture replicate using 10 ng cDNA as the template and the Maxima SYBR Green qPCR master mix (Thermo) with a MyIQ Cycler (Bio-Rad). Primers were designed using Primer-BLAST (NCBI website) to specifically amplify 128- to 155-bp regions of targeted genes Dde_587, Dde_589, Dde_ 590 (Table 2). Dde_ 0587 is the final gene in the rnf operon and the other two genes are the next two ORFs directly downstream of the operon. The 16S rRNA primers were as previously described (Li et al., 2011). Reactions used the following amplification condition: 95 °C for 10 min; 40 cycles of denaturation at 95 °C for 15 s and annealing/extension at 60 °C for 1 min. mRNA expression was calculated using (Etarget)ΔCt_target(control−mutant)∕(Ereference)ΔCt_reference(control−mutant) (Pfaffl, 2001). The 16S rRNA gene was used for normalization.
| Primer | Sequence | Target gene |
|---|---|---|
| Dde_ 587 forward | 5′-AACCGTGGGTTATCGCCATT-3′ | rnfF gene |
| Dde_ 587 reverse | 5′-ATGGCTGTACATGCGTTTGC-3′ | |
| Dde_ 589 forward | 5′-AAGGGCAGGTAGTGCTGATG-3′ | Putative regulator gene |
| Dde_ 589 reverse | 5′-GGTAAACTTCACGCCTTCGC-3′ | |
| Dde_ 590 forward | 5′-GCCTGCGGCTTAATTTCGAG-3′ | Cardiolipin synthase gene |
| Dde_ 590 reverse | 5′-ACGGTTTTCCAGTTCGCTCA-3′ | |
| 16S forward | 5′-ACGGTTGGAAACGACTGCTA-3′ | 16S rRNA gene |
| 16S reverse | 5′-AGCTAATCAGACGCGGACTC-3′ |
Washed cell experiments
Washed cells were used to measure sulfide production under non-growth conditions. Briefly, 100 ml lactate-sulfate culture grown to mid log phase was harvested by centrifugation at 5,300 × g for 15 min, washed twice in buffer containing 50 mM MOPS (pH 7.2), 5 mM MgCl2 and resuspended in the same buffer. All buffers were flushed with N2 for 30 min. Assays were carried out in serum tubes containing 2 ml of buffer-cell mixture incubated at 37 °C on a shaker at 100 rpm. The assay mixture contained the washing buffer, 5 mM sodium sulfate and either 50 mM lactate, 50 mM sodium formate (N2 headspace) or H2 in the headspace. Between 50 and 200 µg of cell protein was added to each tube. Tubes were sacrificed by addition of 2 ml 10% Zinc Acetate at 60 min intervals for sulfide analysis. Sulfide was determined using the methylene blue assay (Cline, 1969).
Measurement of proton motive force
Triplicate cultures of D. alaskensis parent strain and rnfA mutant were grown in lactate–sulfate basal medium until mid exponential phase (OD600 of 0.5) in a 1 liter volume and 30 ml aliquots of cells were dispensed into 100 ml serum bottles under N2∕CO2 (80/20, vol/vol). The basal medium contains 5 mM K+, mainly added as potassium phosphate salts. Protein concentration was determined using the Bicinchoninic Acid Assay (Pierce BCA Protein Assay Kit).
Transmembrane electrical potential (ΔΨ) was measured as previously described (Tremblay et al., 2013). Briefly, cells were incubated with [3H]tetraphenylphosphonium bromide ([3H]TPP+) (ARC, 0.1 µCi/ml) for 15 min at 37 °C (Kashket & Barker, 1977; Shirvan, Schuldiner & Rottem, 1989). [3H]TPP+ was dissolved in media and filtered through 0.45 µm filters prior to adding to cells. Controls were incubated with nigericin (ACROS ORGANICS, 20 µM dissolved in ethanol at 200×) and valinomycin (ACROS), 20 µM dissolved in DMSO at 200× for 15 min to eliminate the ΔΨ. The combination of nigericin (proton/K+ antiporter) and valinomycin (K+ uncoupler) will dissipate the proton gradient (Kessler et al., 1977). Cells were then separated from the medium using silicone oil as described below and [3H]TPP+ was determined in the liquid scintillation counter (Packard TriCarb 2100TR). The uptake of [3H]TPP+ was corrected for extracellular contamination using the ratio of the intracellular to extracellular volume as described below. Non-specific binding of [3H]TPP+ was corrected by subtracting the cell associated [3H]TPP+ in the valinomycin/nigericin treatment from that in the untreated cells. The ΔΨ was calculated with the simplified Nernst equation (−2.3[RT∕F] × log[(concentration in)∕(concentration out)]).
The ΔpH was measured by testing the distribution of [14C ]benzoate (ARC, 0.4 µCi/ml) across the cell membrane after 15 min incubation at 37 °C with cells (Kashket & Barker, 1977; Shirvan, Schuldiner & Rottem, 1989). Cells were then separated from the supernatant as described below and [14C ]benzoate was measured. [14C ]benzoate uptake was corrected for extracellular contamination using the ratio of intracellular to extracellular volume described below. Tetrachlorosalicylanilide (TCS, 20 µM) was added to controls to eliminate the ΔpH and was used to calculate internal pH in the absence of a ΔpH. The external pH was measured, [14C ]benzoate uptake was quantified and intracellular pH was calculated with the Henderson–Hasselbach equation (Rottenberg, 1979). The proton motive force (PMF) was calculated using the equation: PMF = ΔΨ − zΔpH. At 37 °C , z equals to 61.48.
Measurement of intracellular volume/total volume of cell pellet
3H2O (1 µCi/ml) was used to measure the total pellet volume while [14C ]taurine (PerkinElmer, 0.5 µCi/ml) was used to measure extracellular volume and was inferred to penetrate up to the plasma membrane (Kashket & Barker, 1977). The intracellular water volume was estimated from total pellet water volume minus extracellular water volume by measuring the difference between the distribution of 3H2O and [3H]taurine after incubating cells for 15 min at 37 °C with each isotope.
Separation of cells from the supernatant
Following the 15 min incubation with the isotope, three 10 ml aliquots of the total 30 ml solution were put into three 15 ml Falcon tubes with 3 ml of a mixture of silicone oils (25% Fluid 510, 50 centistokes, and 75% Fluid 550, 115 centistokes; Dow-Corning Corp, Serva. vol/vol). Cells were then centrifuged at 5,300 × g at 4 °C for 10 min. The aqueous layer and the silicone oil was removed with a Pasteur pipette connected to a vacuum line. The bottom of the falcon tube containing the cell pellet was cut off and moved into the scintillation vial and the cell pellet was re-suspended with 200 µl distilled-water. Liquid scintillation cocktail (Ultima Gold; PerkinElmer, Waltham, MA, USA) was added prior to scintillation counting.
Statistical analysis
Statistical analysis used either the t-test or a one-way analysis of variance (ANOVA) with the Tukey’s Range test.
Results
Mutants in the rnf operon
In a previous study, a Tn10 mutant library of D. alaskensis G20 was screened for the ability to grow on butyrate in coculture with Syntrophomonas wolfei (Krumholz et al., 2015). Seventeen mutants were obtained that were deficient in syntrophic growth. Of these, two mutants had the transposon insertions within a putative rnf operon. This operon is composed of 8 genes located on the genome as shown in Table 3 and genes appear to be co-transcribed. They are separated from the nearest upstream transcribed region by 185 bps and from the nearest downstream transcribed region by 342 bps. The two syntrophy mutants had transposon insertions within rnfA and rnfD as determined using Arbitrary PCR (Krumholz et al., 2015). Both genes encode integral membrane proteins likely located in the cytoplasmic membrane. RnfD has been previously shown to bind a flavin (Biegel et al., 2011).
| Gene | Function | Lactate/ Sulfate | H2/ Sulfate | H2/ Sulfite | SB | SW |
|---|---|---|---|---|---|---|
| Dde_ 0580 | cyt c family protein | 3.17 | 8.33 | 6.18 | 6.02 | 14.36 |
| Dde_ 0581 | RnfC subunit | 3.05 | 6.68 | 6.12 | 5.61 | 13.50 |
| Dde_ 0582 | RnfD subunit | 1.90 | 4.41 | 3.34 | 2.68 | 7.73 |
| Dde_ 0583 | RnfG subunit | 2.83 | 6.35 | 5.19 | 5.43 | 10.07 |
| Dde_ 0584 | RnfE subunit | 1.86 | 4.95 | 3.80 | 3.39 | 7.63 |
| Dde_ 0585 | RnfA subunit | 2.24 | 5.78 | 4.23 | 2.98 | 8.41 |
| Dde_ 0586 | RnfB subunit | 2.18 | 4.39 | 4.16 | 1.48 | 8.09 |
| Dde_ 0587 | RnfF subunit | 2.20 | 4.90 | 5.25 | 1.94 | 10.15 |
Transcriptional analysis of the rnf operon
The complete transcriptional analysis for strain G20 under lactate-sulfate, H2-sulfate and under syntrophic growth conditions has been previously published (Krumholz et al., 2015). Here we present a summary of normalized values for transcription of each subunit within the rnf operon (Table 3). Transcription of all genes in the operon including dchA are similar under each condition and are affected similarly by growth condition. The most highly expressed genes are the first two in the operon, the gene for the cytochrome c family protein and rnfC, both of which have 1.5–2 fold higher levels of expression than genes for the other subunits. Expression was also enhanced approximately 2-fold when cultures were grown on H2, relative to lactate, indicating the importance of this protein for H2 dependent growth. Syntrophic cultures had enhanced expression of all eight genes compared to lactate/sulfate growth with similar levels of expression in the S. aciditrophicus coculture to H2-grown cells and higher levels of expression observed in the S. wolfei coculture, indicating the importance of this protein complex under syntrophic conditions. Similar effects on transcription provide evidence that all eight genes are located in one operon.
Expression analysis of rnf operons in mutants
To determine whether the transposon insertion eliminated downstream expression of genes in the rnf operon, gap analysis was performed where all intergene regions were amplified except the region between rnfB and rnfA using cDNA prepared using mRNA. All of the intergene regions were amplified for the parent and the two mutants (Fig. S1) confirming that all 8 genes form an operon and indicating that the mutants likely have intact transcripts of the interrupted operon. In this operon, the insertions are likely only affecting the one interrupted gene.
We carried out RT-PCR analysis of the terminal gene in the operon, Dde_ 587 (rnfF) as well as the two genes that followed that operon, Dde_ 589 (uspA) and Dde_ 590 (cls) to be certain that downstream expression was not affected by the insertions. Expression of the terminal gene in the operon was impacted in the mutants with a 4 fold decrease in expression in the rnfA mutant and a twofold increase in the rnfD mutant (Table 4). The following two genes were minimally impacted indicating that the insertions present in the mutants likely do not affect downstream expression.
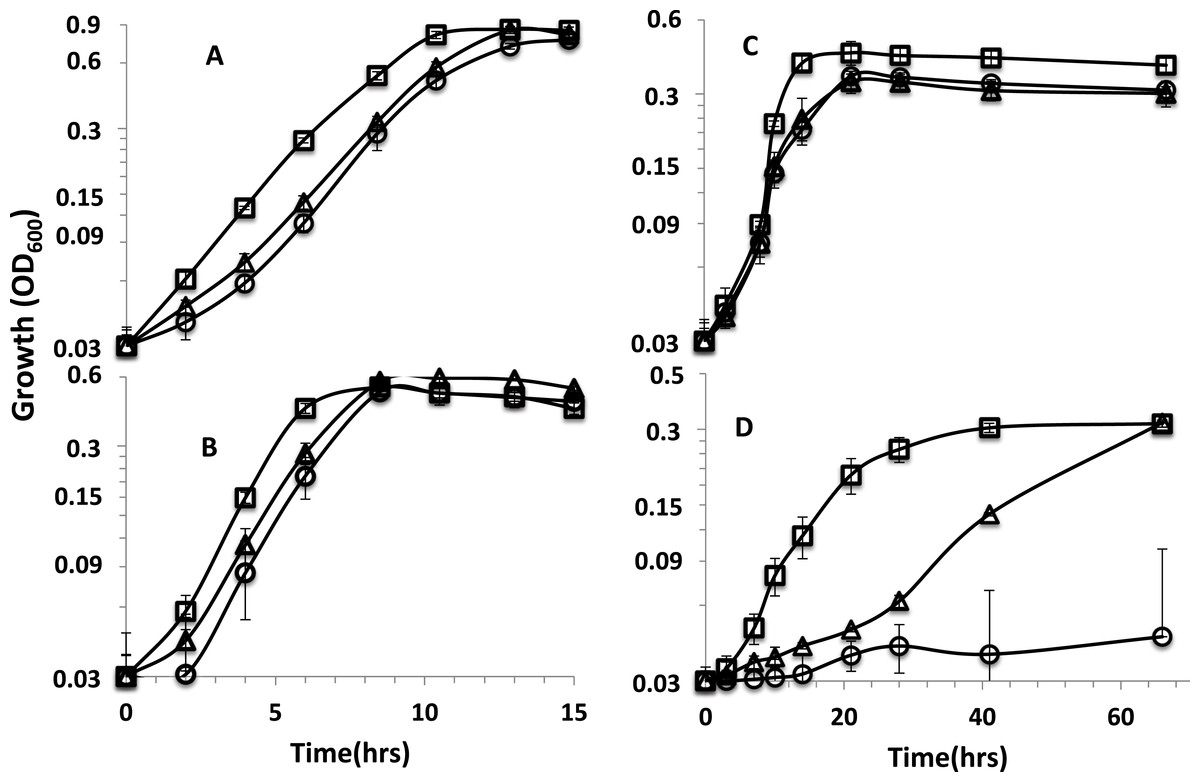
Figure 1: Growth curves of parent strain and mutants.
Growth curves on (A) lactate-sulfate (B) lactate-sulfite (C) pyruvate-sulfate and (D) formate-sulfate. Curves are for D. alaskensis parent strain (□ ) rnfA (○) and rnfD (△) mutants. Error bars show standard deviation.{kind=link}
| Mutant type | Dde_ 587 | Dde_ 589 | Dde_ 590 |
|---|---|---|---|
| rnfA | 0.227 ± 0.255 | 0.608 ± 0.097 | 0.589 ± 0.253 |
| rnfD | 1.955 ± 0.624 | 1.155 ± 0.078 | 0.972 ± 0.420 |
Growth and sulfate reduction by rnf mutants
The two mutants exhibited similar growth rates to that of the parent strain when lactate was the electron donor with sulfate (Fig. 1A) or sulfite as electron acceptor (Fig. 1B). When pyruvate was the donor, growth yields were reduced for the mutants (Fig. 1C). We previously reported that rnf mutants grew poorly (rnfD) or not at all (rnfA) on H2 (Krumholz et al., 2015) and a recent study has reported that rnf mutants grow poorly or not at all with sulfate as the electron acceptor on malate, fumarate, ethanol, hydrogen, and formate, but growth is not affected in lactate and pyruvate (Price et al., 2014). Here, it is shown that the rnfD mutant has a long lag phase with formate as the donor, but eventually grew to a similar OD to the parent strain (Fig. 1D). The rnfA mutant did not grow on formate or H2 clearly indicating the involvement of Rnf in formate/H2-dependent growth. We also confirmed that the rnfA mutant will not grow on ethanol (Fig. S3).
Sulfide production by washed cells
Washed cells were used to determine if the role for rnf was linked to biosynthesis, as mutants were able to grow better on more complex carbon compounds. Incubations of washed cells of the parent strain and the rnfA mutant produced 0.57 and 0.17 µmol sulfide.hr−1.mg−1 protein, respectively, with lactate as electron donor. Addition of either 5 µM TCS to the washed cells of the parent strain prevented 95% of the sulfide production and addition of 20 µM to washed cells inhibited sulfide production by 100%, suggesting TCS directly disrupts the proton gradient needed for respiration. With H2 as the electron donor, parent strain cells produced 0.30 µmol sulfide.hr−1.mg−1 protein whereas the rnfA mutant did not have any detectable activity. To further test whether the lack of growth on H2 was due to the mutant’s inability to biosynthesize carbon intermediates, we attempted to grow the mutant and the parent strain with H2 as the electron donor with 0.1% Casamino acids added to the media. The mutant would still not grow (Fig. S4), providing further evidence that the role for RNF was not directly linked to biosynthesis.
Growth curves with ionophores
The sodium ion ionophore ETH 2120, tested at 20 µM, did not have a significant influence on the growth of the parent strain or the rnf mutants with lactate as the electron donor whether sulfate or sulfite were present as electron acceptors (Figs. S5 and S6). Interestingly, the protonophore, TCS , at 5 µM differentially inhibited cultures (Fig. 2). TCS partially inhibited the growth of the parent strain and completely inhibited the growth of rnfmutants on lactate-sulfate (Fig. 2A and Fig. S5). Growth on lactate sulfite was then tested and 5 µM TCS was shown to have a smaller but still significant inhibitory effect on growth in the parent strain (Fig. 2B) and again completely inhibited the rnfA mutant (Fig. 2B and Fig. S6). A higher level of TCS was then tested and it was shown that 20 µM TCS almost completely inhibited the growth of the parent strain on lactate-sulfite (Fig. 2B) and again completely inhibited the mutants (Fig. S7).
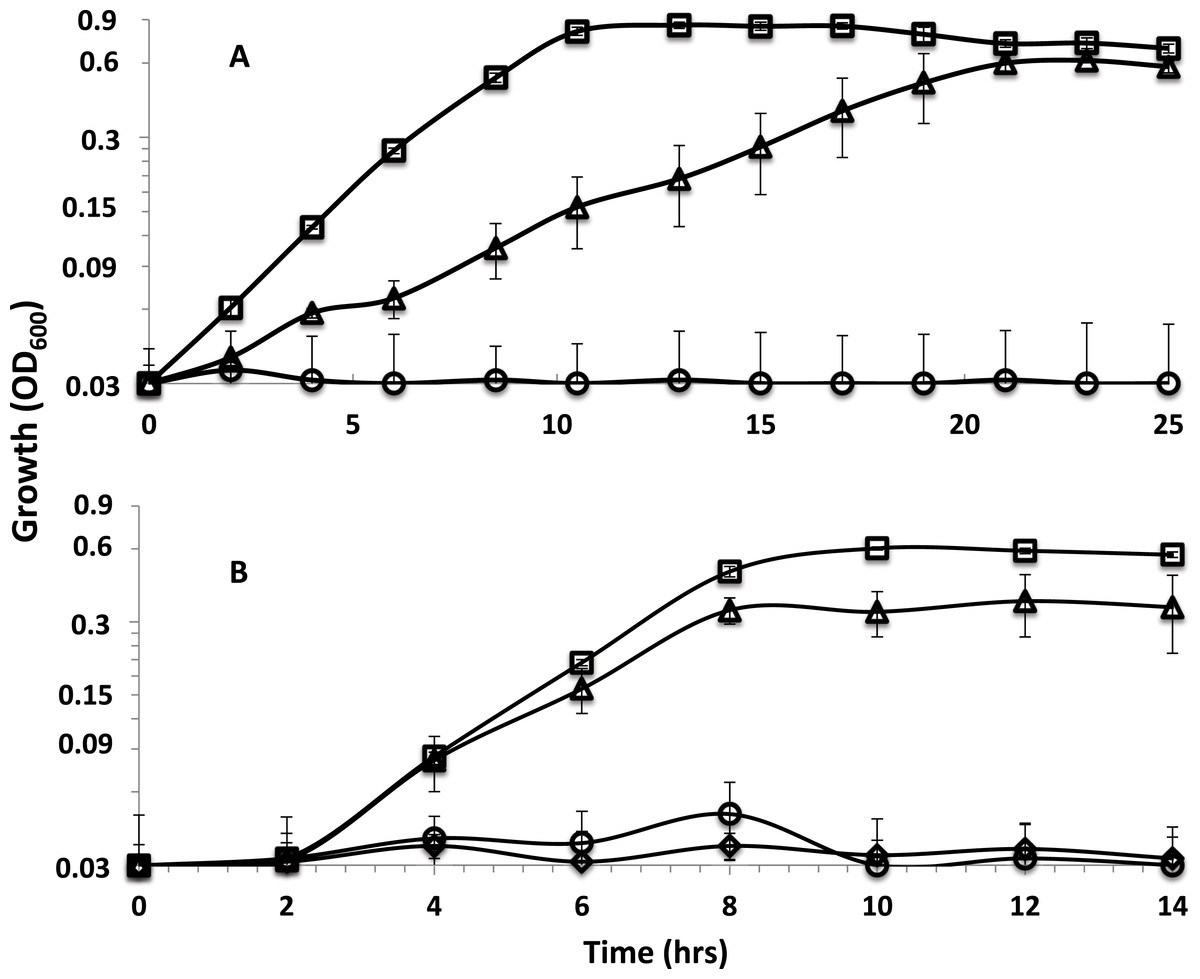
Figure 2: Effect of TCS on parent strain and mutants.
Growth curves with (A) lactate-sulfate and (B) lactate sulfite for D. alaskensis parent strain no addition (□) and with 5 µM (△) or 20 µM (◇) TCS. Growth curve of the rnfA mutant with 5 µM TCS (○) is also shown. Error bars show standard deviation.{kind=link}
Measurement of ΔΨ, ΔpH and PMF
To determine whether the Rnf complex is involved in the formation of a proton gradient, the transmembrane potential (ΔΨ), ΔpH and proton motive force (PMF) were measured in cells of the G20 parent strain and the rnfA mutant growing on lactate-sulfate. The ΔΨ and ΔpH were both reduced in the mutant (Table 5), which led to a significantly lower PMF in the rnfA mutant relative to the G20 parent strain. The lower Δ pH and PMF in the rnfA mutant shows that the Rnf complex clearly had an impact on the energy conservation system by affecting the proton gradient.
| Measurement | Parent strain | rnfA mutant |
|---|---|---|
| ΔΨ | −158 mV (2.1) | −117 mV (3.2) |
| ΔpH | 0.43 (0.057) | 0.29 (0.015) |
| PMF | −185 mV (3.1) | −135 mV (4.1) |
Discussion
The Rnf complex has previously been shown to catalyze the oxidation–reduction reaction between ferredoxin and NAD+ (Biegel, Schmidt & Müller, 2009; Boiangiu et al., 2005). The redox potential of bacterial ferredoxin can vary, but has been reported within the range of −385 to −460 mV (Smith & Feinberg, 1990). That is more negative than that of NAD+/NADH couple (E′ = − 280 mV) and therefore the transfer of electrons by Rnf from reduced ferredoxin to NAD+ releases sufficient energy to generate either a proton or a sodium ion potential (Buckel & Thauer, 2013). Given that recent studies have demonstrated both proton (Tremblay et al., 2013) and Na+ translocation abilities (Biegel & Müller, 2010), it was imperative to determine which ion Rnf translocates in strain G20. Growth experiments showed that lactate-sulfate grown cells were insensitive to the Na+ ionophore, ETH2120, (Figs. S5 and S6) but were highly sensitive to the protonophore, TCS (Fig. 2). Resting cells were also shown to be highly sensitive to TCS suggesting that the proton gradient is needed for sulfate reduction. A similar growth profile was observed in C. ljungdahlii, for which this result was interpreted to infer that a proton gradient was needed for growth (Tremblay et al., 2013).
When grown on lactate-sulfate or lactate-sulfite, TCS partially inhibited the growth of the parent strain and completely inhibited the growth of rnfA and rnfD mutants (Fig. 2). The stronger effect on the mutants suggested that TCS at 5 µM was partially dissolving the proton gradient in the parent strain and this was confirmed when cells were shown to be completely inhibited at 20 µM TCS. We would therefore expect that at 5 µM TCS, growth processes requiring additional proton motive force (or ATP synthesis) would be more highly inhibited than those requiring less ATP. The use of sulfate as an electron acceptor initially requires energy to activate sulfate to adenosine-5’-phosphosulfate (Gavel et al., 1998). The requirement for energy to activate sulfate may explain why lactate-sulfate grown cells are more susceptible to the action of TCS than are lactate-sulfite-grown cells as the proton gradient generated during sulfate respiration would be needed to make ATP for sulfate activation. The inability of rnf mutants to grow in the presence of 5 uM TCS is consistent with its role in the generation of a proton motive force. In fact, the magnitude of the proton motive force in rnfmutants is much less than that in the parent strain of G20 (Table 5).
Ideally, we would have generated complemented rnf mutants to prove that the observed insertions were not having polar effects on other genes. We have attempted to clone the rnfA and rnfD genes into Escherichia coli as the first step in complementing the mutants. Unfortunately we have not been successful. Another group has experienced similar problems with rnfAB of Clostridum ljungdahlii and suggested that rnf genes may be toxic to E. coli in some cases (Tremblay et al., 2013). Gap analysis was used to show that the insertions did not block transcription of downstream genes providing some evidence that polar effects are not important. Also, we carried out RT-PCR analysis of downstream genes. There was a decreased level of expression of the terminal gene in the Rnf operon (rnfF) by the rnfA mutant, however, there was little effect of the insertions (mutations) on expression of genes downstream of the operon.
Experiments reported here and elsewhere (Price et al., 2014) show that the rnf mutants are unable to grow on H2, formate and ethanol. These results point to a critical role for Rnf during growth on the above substrates and are consistent with a higher expression level of rnf genes when growing with H2 and sulfate relative to lactate and sulfate (Table 3).
A proton gradient is thought to be generated during H2 metabolism in Desulfovibrio (Badziong & Thauer, 1980) and used for the synthesis of ATP. Membrane vesicle experiments carried out in our lab in an attempt to demonstrate the generation of an ion gradient coupled to the oxidation of reduced ferredoxin and reduction of NAD+ have been unsuccessful. The protein product of the decaheme cytochrome that precedes the rnf operon has been proposed to accept electrons from hydrogenases and shuttle them to Rnf (Matias et al., 2005; Pereira et al., 2011). However, mutants in this gene had no effect on fitness during growth experiments on ethanol, formate or H2 (Price et al., 2014). This suggests that Rnf is not likely receiving electrons directly from H2. It is more likely that D. alaskensis relies extensively on ferredoxin oxidation by Rnf to produce a proton gradient during growth on substrates that do not yield net ATP by substrate-level phosphorylation. For those substrates that do yield ATP by substrate level phosphorylation such as malate, fumarate, pyruvate and lactate, a decreased growth rate and or yield was observed in most cases for rnf mutants (Fig. 1) (Price et al., 2014) suggesting that both Rnf and the F1Fo ATPase are involved in generating a PMF under those conditions.
We are not familiar with any studies describing mechanisms of ferredoxin reduction in Desulfovibrio, however, several possible mechanisms have been suggested (Pereira et al., 2011; Price et al., 2014). During H2 oxidation, these include a possible cytoplasmic electron-bifurcating hydrogenases-linked to a heterodisulfide reductase for which mutants grow poorly on H2 and formate (Hdr/flox-1). For ethanol oxidation, the acetaldehyde:ferredoxin oxidoreductase could be used, and with pyruvate and lactate oxidation, would involve pyruvate:ferredoxin oxidoreductase.
Results from this study are consistent with the fact that D. alaskensis Rnf complex functions as a proton rather than a sodium pump and is essential for growth on substrates that do not involve ATP synthesis by substrate-level phosphorylation. Mutation of Rnf limits the development of the PMF and, thus, affects ATP synthesis during growth.
Supplemental Information
DesulfovibrioRNF supplement
This is a complete file with all of the supplementary figures.
Figure S1
Results from PCR analysis showing the amplification of gene intervening regions on the transcripts. On the left of the gel, results from the rnfA mutant on top and the rnfD mutant are underneath. The results from the parent strain are on the right side of the gel. GN corresponds to the rnfG-rnfE gap and A corresponds to the rnfE-rnfA gap represented by primers in Table 1.
Figure S2
Growth curve on for D. alaskensis G20 parent strain (squares), the rnfA (diamonds) and the rnfD (traingles) mutant. Standard deviation is shown.
Figure S3
Growth curve on Ethanol for D. alaskensis G20 parent strain (squares) and the rnfA (traingles) mutant. Standard deviation is shown.
Figure S4
First day of growth curves on for D. alaskensis G20 parent strain (□) and rnfD mutant (△). (A) One set of cultures was incubated in the basal media described in methods (B) and the other set had 0.05% casamino acids added. Standard deviation is shown.
Figure S5
Growth curves on for D. alaskensis G20 parent strain (A), the rnfA (B) and the rnfD (C) mutant. One set of cultures was incubated with ETH 2120 (20 µM) (triangles) and the other without (squares). In plot C, the lack of growth of the rnfD mutant with 5uM TCS is shown (diamonds). Standard deviation is shown.
Figure S6
Growth curves on for D. alaskensis G20 parent strain (A), the rnfA (B) and the rnfD (C) mutant. One set of cultures was incubated with ETH 2120 (20 µM) (triangles) and the other without (squares). In plot C, the lack of growth of the rnfD mutant with 5uM TCS is shown (diamonds). Standard deviation is shown.
Figure S7
Growth curves on Lactate/SO32- for D. alaskensis G20 rnfA mutant. One set of cultures was incubated with TCS (20 µM) (triangles) and the other without (squares). Standard deviation is shown.