
Investigation of the effects of estrogen on skeletal gene expression during zebrafish larval head development
- Published
- Accepted
- Received
- Academic Editor
- Joan Heath
- Subject Areas
- Aquaculture, Fisheries and Fish Science, Biodiversity, Developmental Biology, Molecular Biology, Toxicology
- Keywords
- Craniofacial skeleton, Development, Estrogen, Gene expression, Zebrafish larvae, qPCR, Reference genes, 17-β estradiol
- Copyright
- © 2016 Pashay Ahi et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. Investigation of the effects of estrogen on skeletal gene expression during zebrafish larval head development. PeerJ 4:e1878 https://doi.org/10.7717/peerj.1878
Abstract
The development of craniofacial skeletal structures requires well-orchestrated tissue interactions controlled by distinct molecular signals. Disruptions in normal function of these molecular signals have been associated with a wide range of craniofacial malformations. A pathway mediated by estrogens is one of those molecular signals that plays role in formation of bone and cartilage including craniofacial skeletogenesis. Studies in zebrafish have shown that while higher concentrations of 17-β estradiol (E2) cause severe craniofacial defects, treatment with lower concentrations result in subtle changes in head morphology characterized with shorter snouts and flatter faces. The molecular basis for these morphological changes, particularly the subtle skeletal effects mediated by lower E2 concentrations, remains unexplored. In the present study we address these effects at a molecular level by quantitative expression analysis of sets of candidate genes in developing heads of zebrafish larvae treated with two different E2 concentrations. To this end, we first validated three suitable reference genes, ppia2, rpl8 and tbp, to permit sensitive quantitative real-time PCR analysis. Next, we profiled the expression of 28 skeletogenesis-associated genes that potentially respond to estrogen signals and play role in craniofacial development. We found E2 mediated differential expression of genes involved in extracellular matrix (ECM) remodelling, mmp2/9/13, sparc and timp2a, as well as components of skeletogenic pathways, bmp2a, erf, ptch1/2, rankl, rarab and sfrp1a. Furthermore, we identified a co-expressed network of genes, including cpn1, dnajc3, esr1, lman1, rrbp1a, ssr1 and tram1 with a stronger inductive response to a lower dose of E2 during larval head development.
Introduction
Craniofacial development is a critical part of embryogenesis and identification of molecular mechanisms underlying this process is important in gaining a better understanding of morphological diversity in vertebrates (Szabo-Rogers et al., 2010) as well as human health (Oginni & Adenekan, 2012). The viscerocranium in humans is of interest because of oro-facial clefts and associated malformations (Marazita, 2012). The vertebrate craniofacial skeleton, including the viscerocranium, is built from neural-crest derived tissues. Changes in these tissues over evolutionary time have given rise to a wide diversity of facial morphologies among vertebrate species (Trainor, Melton & Manzanares, 2003; Bronner & LeDouarin, 2012).
Estrogens, steroid hormones synthesized by aromatase from androgenic precursors, have recently been shown to affect craniofacial development (Fushimi et al., 2009; Cohen et al., 2014). Though estrogens are normally thought of as sex hormones, they affect a variety of tissues including the cardiovascular and skeletal systems (Hall, Couse & Korach, 2001; Allgood et al., 2013; Cohen et al., 2014). Estrogens signal through classical nuclear receptors (ER-alpha and ER-beta) (Jia, Dahlman-Wright & Gustafsson, 2015) and a G-protein coupled receptor, GPR-30 (Jenei-Lanzl et al., 2010). These signaling proteins are found in the chondrocytes of many vertebrate species (Tankó et al., 2008) and are present during chondrogenesis (Jenei-Lanzl et al., 2010; Elbaradie et al., 2013). Among vertebrates, teleost fish are highly diversified in craniofacial morphology and estrogen has been shown to greatly affect chondrogenesis in many of the fish species studied so far, including tilapia, fathead minnow, and zebrafish (Ng, Datuin & Bern, 2001; Warner & Jenkins, 2007; Cohen et al., 2014). Furthermore, the teleost zebrafish is a well-developed model system for studying both embryonic development and human disease and it can be useful in understanding the development of the human viscerocranial skeleton as these processes are well-conserved among vertebrates (Kuratani, Matsuo & Aizawa, 1997).
Low concentrations of estrogen cause subtle changes in craniofacial morphogenesis during zebrafish larval development (Cohen et al., 2014). These changes are characterized by a shorter snout, flatter face and wider angles of cartilage elements in the viscerocranium (Cohen et al., 2014). Another recent study has also demonstrated that an estrogen mediated signal underlies the evolution of shorter snouts and flatter faces in females of some reptilian species (Sanger et al., 2014). These observations suggest that similar mechanisms might underpin hormone-based phenotypic plasticity and diversity (Dufty, 2002), as well as subtle differences in head/skeletal morphology of dimorphic sexes (Loth & Henneberg, 2001; Fujita et al., 2004; Callewaert et al., 2010). Therefore, studies offering insights into molecular mechanisms rendering the observed phenotypes caused by hormonal changes would be called for.
| Gene symbol | Related function | Viscerocranial expression during zebrafish development | Potential estrogen responsive* | Craniofacial skeletogenesis | References | |
|---|---|---|---|---|---|---|
| Shortened snout** | Other effects | |||||
| alx4 | Patterning and development of craniofacial skeleton | + | + | + | + | (Qu et al., 1999; Joshi, Chang & Hamel, 2006; Lours-Calet et al., 2014) |
| bmp2 (a/b) | Induction of bone and cartilage formation | + | + | ? | + | Thisse et al. (2001), Thisse et al. (2004), Zhou et al. (2003), Nie, Luukko & Kettunen (2006), Hu, Colnot & Marcucio (2008), and Yamamoto, Saatcioglu & Matsuda (2013) |
| col2a1a | Extracellular matrix formation in cartilaginous tissues | + | + | + | + | Maddox et al. (1997), Eames et al. (2010) and Maneix et al. (2014) |
| ctsk | Bone remodelling and resorption | + | + | + | + | Thisse et al. (2004), Troen (2006), Petrey et al. (2012) and Ahi et al. (2014) |
| dlk1 | Differentiation of skeletal cells | ? | + | + | + | Abdallah et al. (2011) |
| erf | Regulation of cellular senescence | ? | + | + | + | Frasor et al. (2003) and Twigg et al. (2013) |
| esrra | Regulation of estrogen mediated pathway | + | + | ? | + | Bonnelye & Aubin (2005), Bonnelye et al. (2007) and Auld et al. (2012) |
| esr1 | A ligand-activated receptor for estrogen | ? | + | ? | + | O’Lone et al. (2004) and Syed et al. (2005) |
| ets2 | Regulation of developmental genes and apoptosis | ? | + | + | + | Sumarsono et al. (1996), Deblois et al. (2009) and Ahi et al. (2014) |
| mmp (2/9/13) | Extracellular matrix formation and signal transduction | + | + | + | + | Breckon et al. (1999), Tüshaus et al. (2003), Marin-Castaño et al. (2003), Lu et al. (2006), Hillegass et al. (2007a), Hillegass et al. (2007b), Mosig et al. (2007) and Nilsson, Garvin & Dabrosin (2007) |
| opg | Negative regulation of bone resorption | ? | + | ? | + | Bord et al. (2003) and Whyte et al. (2002) |
| pbx1 (a/b) | Co-ordination of chondrocyte proliferation and differentiation | + | + | + | + | Selleri et al. (2001), Thisse & Thisse (2005) and Magnani et al. (2011) |
| ptch (1/2) | Receptors for hedgehog signalling pathway | + | + | + | + | (Fushimi et al., 2009; Roberts et al., 2011) |
| rankl | Osteoclast differentiation and activation | ? | + | ? | + | Bord et al. (2003) and Lézot et al. (2015) |
| rarab | A receptor for retinoic acid signalling pathway | + | + | + | + | (Lohnes et al., 1994; O’Lone et al., 2004; Linville et al., 2009) |
| runx2b | Osteoblast differentiation and skeletal morphogenesis | + | + | + | + | Sears et al. (2007), McCarthy et al. (2003) and Flores et al. (2006) |
| sfrp1a | A soluble modulator of Wnt signalling pathway | ? | + | + | + | Satoh et al. (2006), Trevant et al. (2008), Yokota et al. (2008), Fukuhara et al. (2013) and Ahi et al. (2014) |
| Shh (a/b) | Activators of hedgehog signalling pathway | + | ? | ? | + | Hu & Helms (1999) and Swartz et al. (2012) |
| sox9b | Chondrocyte differentiation | + | + | ? | + | Yan et al. (2005), Bonnelye et al. (2007) and Lee & Saint-Jeannet (2011) |
| sparc | Extracellular matrix synthesis and regulation of cell growth | + | + | + | + | Lehane et al. (1999), Renn et al. (2006) and Rotllant et al. (2008) |
| spp1 | Attachment of osteoclasts to ECM in bone | + | + | + | + | Craig & Denhardt (1991), Vanacker et al. (1998) and Venkatesh et al. (2014) |
| timp2a | Inhibition of mmps and regulation of tissue homeostasis | ? | + | + | ? | Dew et al. (2000), Lam et al. (2009), Letra et al. (2012), Wang & Ma (2012) and Ahi et al. (2014) |
Notes:
The subtle changes in craniofacial skeletogenesis mediated by low concentrations of Estradiol (E2) are likely to be a result of differences in level and timing of the expression of skeletogenesis-associated genes during head development (Albertson et al., 2010; Ahi et al., 2014; Gunter, Koppermann & Meyer, 2014; Powder et al., 2015). These morphological changes were only revealed by careful measurements of skeletal elements at zebrafish larval stages (Cohen et al., 2014), therefore the identification of responsible genes might also require precise expression studies in developing heads of zebrafish larvae using a sensitive tool such as quantitative real-time PCR (qPCR) (Bustin, 2000; Kubista et al., 2006). In the present study, we set out to quantitatively assess the effects of estrogen on the expression dynamics of candidate genes which are known as potential targets of estrogen pathway and also involved in craniofacial skeletogenesis in different vertebrate species (Table 1). We hypothesized that these genes may be critical to the estrogen modulation of craniofacial skeletogenesis. We first identified the most stably expressed reference genes in developing heads of zebrafish treated with two doses of estrogen (2 µM and 5 µM) across five stages in larval development. Then, we accurately measured small changes in the expression levels of the candidate genes. In addition, we have used available co-expression data from zebrafish to identify a co-expressed network of genes with greater transcriptional response to the lower dose of estrogen (2 µM) during larval head development.
Methods
Fish husbandry, treatment and sampling
Adult zebrafish were fed a diet of live brine shrimp supplemented with Ziegler zebrafish diet (Pentair) and maintained on a 14/10 day/night cycle. Embryos were raised in E3B (5 mM NaCl, 0.17 mM KCl, 0.33 mM CaCl2, 0.33 mM MgSO4, 0.00025% methylene blue). Embryos were treated with estrogen (17β-estradiol, E2, Sigma) dissolved in ethanol and diluted in E3B for a final ethanol concentration of 0.1%. Control fish were treated with 0.1% ethanol with no developmental malformations as described previously (Cohen et al., 2014). For each treatment group (estrogen concentration), zebrafish larva were raised in Petri dishes, and treatment solutions were refreshed daily until the stages indicated (3, 4, 5, 6 and 7 days post fertilization, dpf). Three biological replicates of 30 larva were collected at each time-point (3–7 dpf) and for each treatment group (control, 2 µM E2, and 5 µM E2) for a total of 90 larva at each time-point and treatment. The fishes were anesthetized with 0.4% tricaine (MS-222, Sigma). Isolated heads (anterior to the yolk sac) were placed into RNAlater (Qiagen) and stored frozen until RNA isolation. Zebrafish experiments were performed under the Roanoke College IRB protocol #14BIO76.
RNA isolation and cDNA synthesis
Around 30 heads of zebrafish from each treatment group and larval stage were pooled in TRI Reagent (Sigma) and homogenized with a disposable Kontes Pellet Pestle Cordless Motor tissue grinder (Kimble Kontes). RNA was prepared according to manufacturer’s instructions and dissolved in 50 µl RNase-free water. RNA samples were treated with DNase (New England Biolabs) to remove contaminating DNA. Quantity of the resulting RNA samples was assessed using a NanoDrop ND-1000 UV/Vis-Spectrophotometer (NanoDrop Technologies). The quality of the RNA samples was evaluated by agarose gel electrophoresis and all samples displayed intact 28 S and 18 S rRNA without noticable high molecular weight genomic DNA contamination. cDNA was prepared from 1000 ng of RNA using the High capacity cDNA Reverse Transcription kit (Applied Biosystems), according to manufacturer’s protocol. Several samples without addition of reverse transcriptase (-RT samples) were prepared to confirm the absence of genomic DNA. cDNA was diluted 3 fold in water for further use in quantitative real-time PCR.
Gene selection, Primer design and real-time qPCR
In order to validate suitable reference genes for accurate measurement of the transcriptional changes of candidate genes by qPCR, we selected 7 potential reference genes based on published studies in zebrafish (Table S1) (McCurley & Callard, 2008; Pelayo et al., 2012; Schiller et al., 2013), none of which have been validated during development or in zebrafish head. In addition we selected 28 target genes that are known as potential targets of the estrogen pathway in different vertebrate species, and also involved in craniofacial skeletal formation/morphogenesis (Table 1 and Table S1). Finally, we extended our list of candidates by adding more genes showing co-expression with the estrogen receptor esr1 based on the zebrafish database COXPRESdb (http://coxpresdb.jp/) version 6.0 (Obayashi & Kinoshita, 2011). To obtain the maximum number of coexpressed genes with a high degree of reliability, we filtered the genes by setting the mutual rank (MR) to the top-ranked 2000 and the Supportability score of minimum 1 (as described by Obayashi & Kinoshita, 2011). This yielded 338 candidate genes, and from them, we selected 11 genes with reported craniofacial expression during zebrafish development according to the ZFIN database (http://zfin.org) (Bradford et al., 2011) (Table S1).
Locations overlapping exon boundaries of the genes in zebrafish were determined by NCBI Spidey software (www.ncbi.nlm.nih.gov/spidey) and annotated genome sequences in the Ensembl database (http://www.ensembl.org/Danio_rerio). The qPCR Primers were designed on exon boundaries using Primer Express 3.0 software (Applied Biosystems, Foster City, CA, USA) and checked for self-annealing, hetero-dimers and hairpin structures with OligoAnalyzer 3.1 (Integrated DNA Technology) (Table S1).
Real-time PCR was performed in 96 well-PCR plates on an ABI 7500 real-time PCR System (Applied Biosystems) using Maxima SYBR Green/ROX qPCR Master Mix (2X) as recommended by the manufacturer (Thermo Fisher Scientific, St Leon-Rot, Germany). Each biological replicate was run in duplicate together with no-template control (NTC) in each run for each gene and experimental set-up per run followed the preferred sample maximization method (Hellemans et al., 2007). The qPCR was run with a 2 min hold at 50 °C and a 10 min hot start at 95 °C followed by the amplification step for 40 cycles of 15 sec denaturation at 95 °C and 1 min annealing/extension at 60 °C. A dissociation step (60 °C–95 °C) was performed at the end of the amplification phase to identify a single, specific product for each primer set (Table S1). Primer efficiency values (E) were calculated with the LinRegPCR v11.0 programme (http://LinRegPCR.nl) (Ramakers et al., 2003) analysing the background-corrected fluorescence data from the exponential phase of PCR amplification for each primer-pair and those with E less than 0.9 were discarded and new primers designed (Table S1).
Data analysis
To detect the most stably expressed reference genes, three ranking algorithms; BestKeeper (Pfaffl et al., 2004), NormFinder (Andersen, Jensen & Ørntoft, 2004) and geNorm (Vandesompele et al., 2002) were employed. The standard deviation (SD) based on Cq values of the larval stages and treatment groups was calculated by BestKeeper to determine the expression variation for each reference gene. In addition, BestKeeper determines the stability of reference genes based on correlation to other candidates through calculation of BestKeeper index (r). GeNorm measures mean pairwise variation between each gene and other candidates, the expression stability or M value, and it excludes the gene with the highest M value (least stable) from subsequent analysis in a stepwise manner. Moreover, geNorm determines the optimal number of reference genes through calculation of pairwise variation coefficient (Vn∕n + 1) between two sequential normalisation factors (NFn and NFn + 1) and extra reference genes are added until the variation drops below the recommended threshold of 0.15 (Vandesompele et al., 2002). NormFinder identifies the most stable genes (lowest expression stability values) based on analysis of the sample subgroups (stage and treatment group) and estimation of inter- and intra-group variation in expression levels.
For the analysis of qPCR data, the difference between Cq values (ΔCq) of the reference genes and the target genes was calculated for each gene; ΔCqtarget = Cqtarget − Cqreference. The geometric mean of Cq values of three best ranked reference genes, ppia2, rpl8 and tbp (see the ranking algorithms above), was used as Cq reference in the ΔCq calculations. All samples were then normalized to the ΔCq value of a calibrator sample to obtain a ΔΔCq value (ΔCqtarget − ΔCqcalibrator). For each primer pair a biological replicate in the control group at 3dpf was selected as the calibrator sample. Relative expression quantities (RQ) were calculated based on the expression level of the calibrator sample (E−ΔΔCq) (Pfaffl, 2001). The RQ values were then transformed to logarithmic base 2 values (or fold differences; FD) for statistical analysis (Bergkvist et al., 2010). A two-way analysis of variance (ANOVA) followed by post hoc Tukey’s honest significant difference (HSD) test was implemented for each reference or target gene with larval stages and treatment groups as categorical variables. To assess similarities in expression patterns of the genes Pearson correlation coefficients (r) were calculated for all gene pairs using the data from 3 treatments at 5 larval stages (degree of freedom = 13). R (http://www.r-project.org) was used for all statistical analysis.
Results
tbp, ppia2 and rpl8 are the most suitable reference genes
Real-time quantitative PCR for the 7 reference gene candidates was performed on cDNA generated from zebrafish head homogenates in three treatment groups at five larval stages. The expression levels of the candidates varied from ppia2, with the highest expression (lowest Cq) (Fig. 1A), to tbp with the lowest expression (highest Cq). Statistical analysis revealed that all of the candidates except actb1 are stably expressed between the treatment groups (Fig. 1B). However, only tbp showed constant expression during the larval stages examined. Two genes, ppia2 and rpl8, were also stably expressed in developing heads of zebrafish larvae except for the first stage (3dpf). Based on these results tbp followed by ppia2 and rpl8 were found to be the overall most stable reference genes both over time and between the treatment groups. The candidate reference genes were ranked using three algorithms, i.e., BestKeeper, geNorm and NormFinder, and based on standard deviation (SD) as described in Ahi et al. (2013) (Table 2). In all of the analyses three genes; ppia2, rpl8 and tbp, were the three highest ranking candidates, however their order varied between the rankings (Table 2). Furthermore, geNorm suggested the use of the three best ranked candidate genes as sufficient for accurate normalisation (Fig. S1). The data reflect the high expression stability of the best ranked candidate genes and suggests the combination of ppia2, rpl8 and tbp as a suitable and sufficient normalization factor to accurately quantify small differences in gene expression in developing heads of zebrafish larvae across the E2 treatment groups.
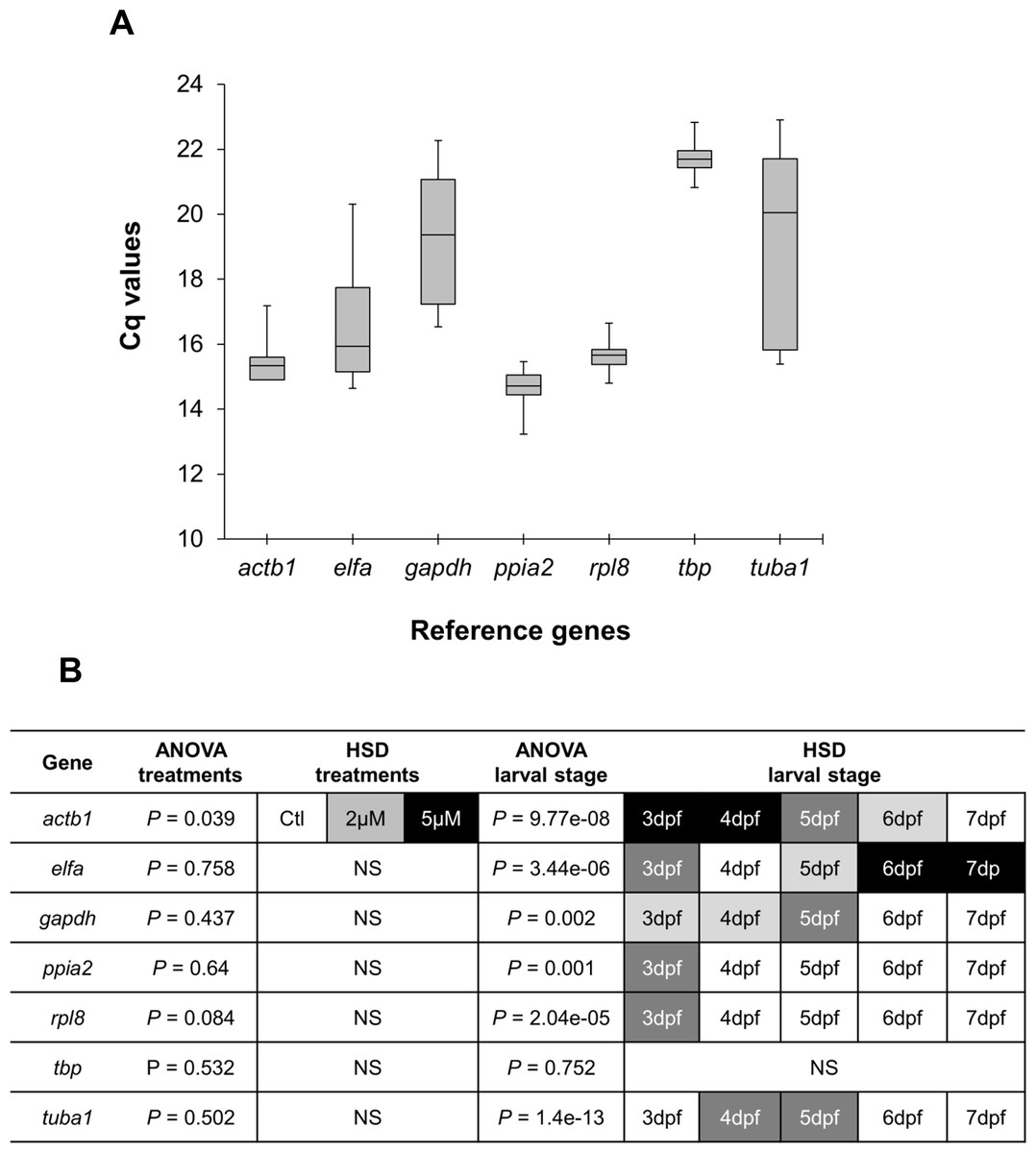
Figure 1: Expression analysis of candidate reference genes in developing heads of zebrafish larvae across control and E2 treated groups.
(A) Expression profiles of candidate reference genes in raw Cq values for all samples (3 treatments for each of 5 larval stages and with 3 biological replicates). The middle line denotes the median and boxes indicate the 25/75 percentiles. (B) Expression differences of candidate reference genes in the head of zebrafish during the larval development and three E2 treatment groups. Fold changes in expression calculated from the qPCR data, were subjected to ANOVA and Tukey’s HSD analysis to test the expression differences amongst three treatment groups (control, 2 µM and 5 µM) and across five larval stages (3 to 7dpf). White boxes represent low expression, while black boxes represent high expression. Two or more steps of shade differences in the boxes represent significantly different expression between the samples (alpha = 0.05). NS, not significant.{kind=link}
| BestKeeper | geNorm | NormFinder | |||||
|---|---|---|---|---|---|---|---|
| Ranking | r | Ranking | SD | Ranking | M | Ranking | SV |
| rpl8 | 0.908 | tbp | 0.294 | ppia2 | 0.111 | rpl8 | 0.137 |
| tbp | 0.863 | rpl8 | 0.343 | rpl8 | 0.125 | ppia2 | 0.154 |
| ppia2 | 0.862 | ppia2 | 0.350 | tbp | 0.133 | tbp | 0.157 |
| actb1 | 0.687 | actb1 | 0.396 | actb1 | 0.26 | actb1 | 0.287 |
| efl1a | 0.331 | efl1a | 1.358 | efl1a | 0.739 | efl1a | 1.128 |
| tuba1 | 0.201 | gapdh | 1.690 | gapdh | 1.084 | gapdh | 1.382 |
| gapdh | 0.148 | tuba1 | 2.773 | tuba1 | 1.482 | tuba1 | 2.088 |
Notes:
- SD
-
Standard deviation
- r
-
Pearson product-moment correlation coefficient
- SV
-
Stability value
- M
-
M value of stability
Components of different signalling pathways and skeletogenesis-associated genes are affected by estrogen during larval head development
The selected 28 candidate target genes, listed in Table 1, can be classified into distinct functional groups; (I) estrogen receptors with potential involvement in vertebrate craniofacial development (esrra and esr1); (II) components of hedgehog (Hh) signaling pathway (ptch1/2 and shha/b); (III) potential skeletogenic targets of estrogen pathway with critical roles in viscerocranial development/morphogenesis (bmp2a/b, opg, rankl, runx2b and sox9b); (IV) potential targets of estrogen pathway involved in ECM formation and associated with shortened snout morphogenesis in vertebrates (col2a1a, ctsk, mmp2/9/13, sparc, spp1 and timp2a); and (V) other potential targets of estrogen pathways with diverse functions which are also involved in viscerocranial skeletogenesis (alx4, dlk1, erf, ets2, pbx1a/b, rarab and sfrp1a). The expression levels of all candidates were measured in the three treatment groups during larval head development (Figs. 2, 3, 4 and 5). We found effects of different E2 concentrations on the expression of most of the target genes, except col2a1a and pbx1a, the effects, however, were highly variable among the genes (Figs. 2, 3, 4 and 5). For instance, while some genes, i.e., esr1, ptch1/2 and rarab displayed differential expression between the treatment groups at most of the larval stages, other genes such as alx4, bmp2b, ctsk, ets2, opg, etc., showed expression differences at only one stage. Among the more highly affected genes, erf, esrra, mmp9, rankl, shha, sfrp1a, sparc and timp2a were differentially expressed in at least three larval stages (Figs. 2, 3, 4 and 5). Although significant, most differences in expression levels of the target genes were slight between the treatment groups (RQ < 0.5), except for esr1 at the last larval stages (Fig. 2). Moreover, for all of the affected genes, except esr1 and mmp13, the different E2 treatments had mainly repressive effects on transcription. These repressive effects were not, however, increased by higher E2 concentration particularly at the last two stages when the lower E2 dose (2 µM) repressed expression of many of the genes more than the higher dose. At the last three stages, the expression of esr1 was induced at highest levels for 2 µM treatment groups (Fig. 2). The transcriptional repression by E2 was also variable between the genes and it was more pronounced for erf and ptch2 showing higher expression in control groups than both E2 treated groups at three larval stages. Taken together, these results show significant effects of low E2 concentrations on the expression of a variety of genes involved in skeletogenesis and/or craniofacial development.
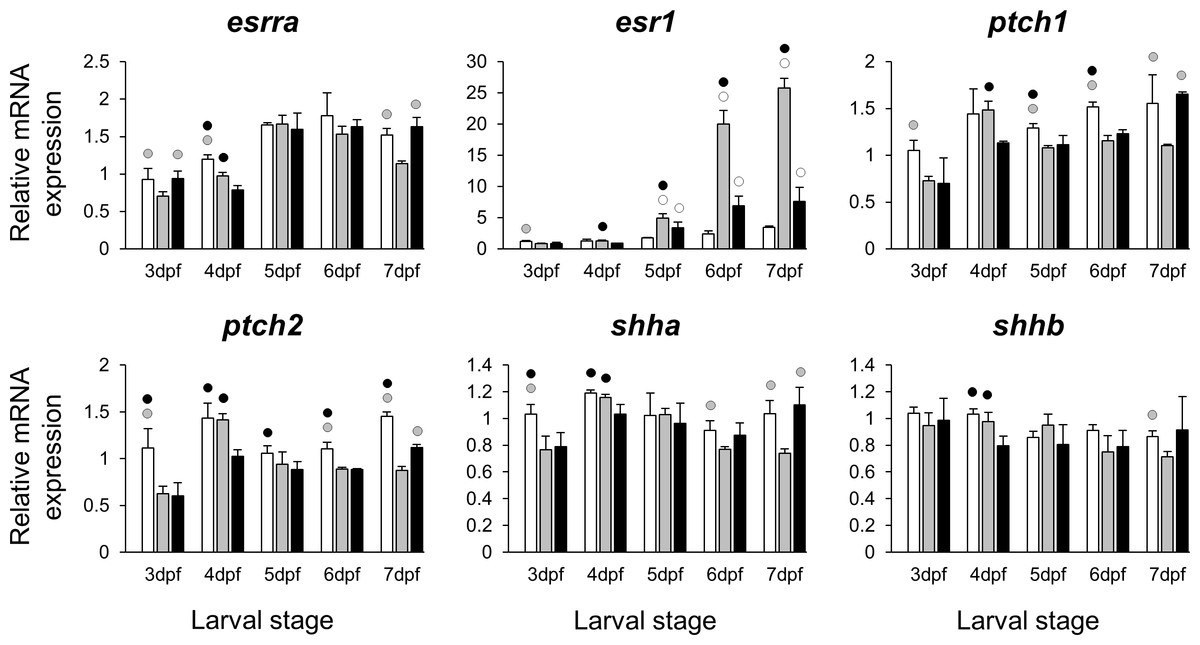
Figure 2: Expression differences of two estrogen receptors and components of hedgehog signaling pathway in developing heads of zebrafish larvae across control and E2 treated groups.
Expression of esrra, esr1, ptch1, ptch2, shha and shhb was examined with qPCR and normalised using three highest ranked reference genes (ppia2, rpl8 and tbp). For analysis of relative expression levels for each target gene a replicate of the control group at 3dpf was set to one. The white, grey, and black bars in each graph represent expression levels for control, 2 µM E2 treated and 5 µM E2 treated groups respectively. Statistical differences of each treatment group versus the others are shown in white, grey, and black circles representing higher expressed than control, 2 µM E2 treated and 5 µM E2 treated groups respectively (P < 0.05). Error bars represent standard deviation calculated from three biological replicates. Each biological replicate is from a homogenate of 30 heads.{kind=link}

Figure 3: Expression differences of six potential skeletogenic targets of estrogen pathway in developing heads of zebrafish larvae across control and E2 treated groups.
Expression of bmp2a, bmp2b, opg, rankl, runx2b and sox9b was examined with qPCR and normalised using three highest ranked reference genes (ppia2, rpl8 and tbp). For analysis of relative expression levels for each target gene a replicate of the control group at 3dpf was set to one. The white, grey, and black bars in each graph represent expression levels for control, 2 µM E2 treated and 5 µM E2 treated groups respectively. Statistical differences of each treatment group versus the others are shown in white, grey, and black circles representing higher expressed than control , 2 µM E2 treated and 5 µM E2 treated groups respectively (P < 0.05). Error bars represent standard deviation calculated from three biological replicates. Each biological replicate was made from a homogenate of 30 heads.{kind=link}
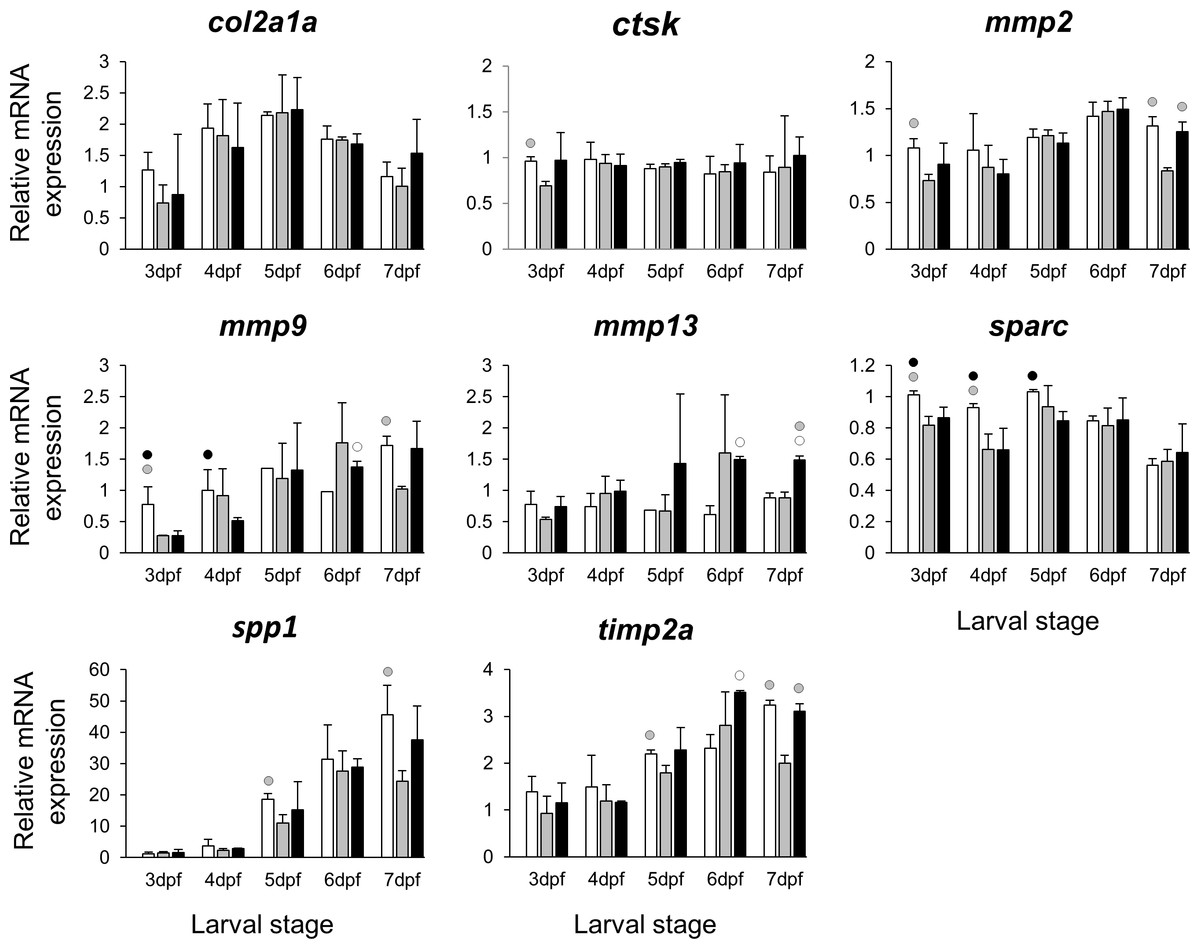
Figure 4: Expression differences of eight potential targets of estrogen pathway involved in skeletal ECM formation examined during zebrafish larval head development across control and E2 treated groups.
Expression of col2a1a, ctsk, mmp2, mmp9, mmp13, sparc, spp1 and timp2 was examined with qPCR and normalised using three highest ranked reference genes (ppia2, rpl8 and tbp). For analysis of relative expression levels for each target gene a replicate of the control group at 3dpf was set to one. The white, grey, and black bars in each graph represent expression levels for control, 2 µM E2 treated and 5 µM E2 treated groups respectively. Statistical differences of each treatment group versus the others are shown in white, grey, and black circles representing higher expressed than control, 2 µM E2 treated and 5 µM E2 treated groups respectively (P < 0.05). Error bars represent standard deviation calculated from three biological replicates. Each biological replicate was made from a homogenate of 30 heads.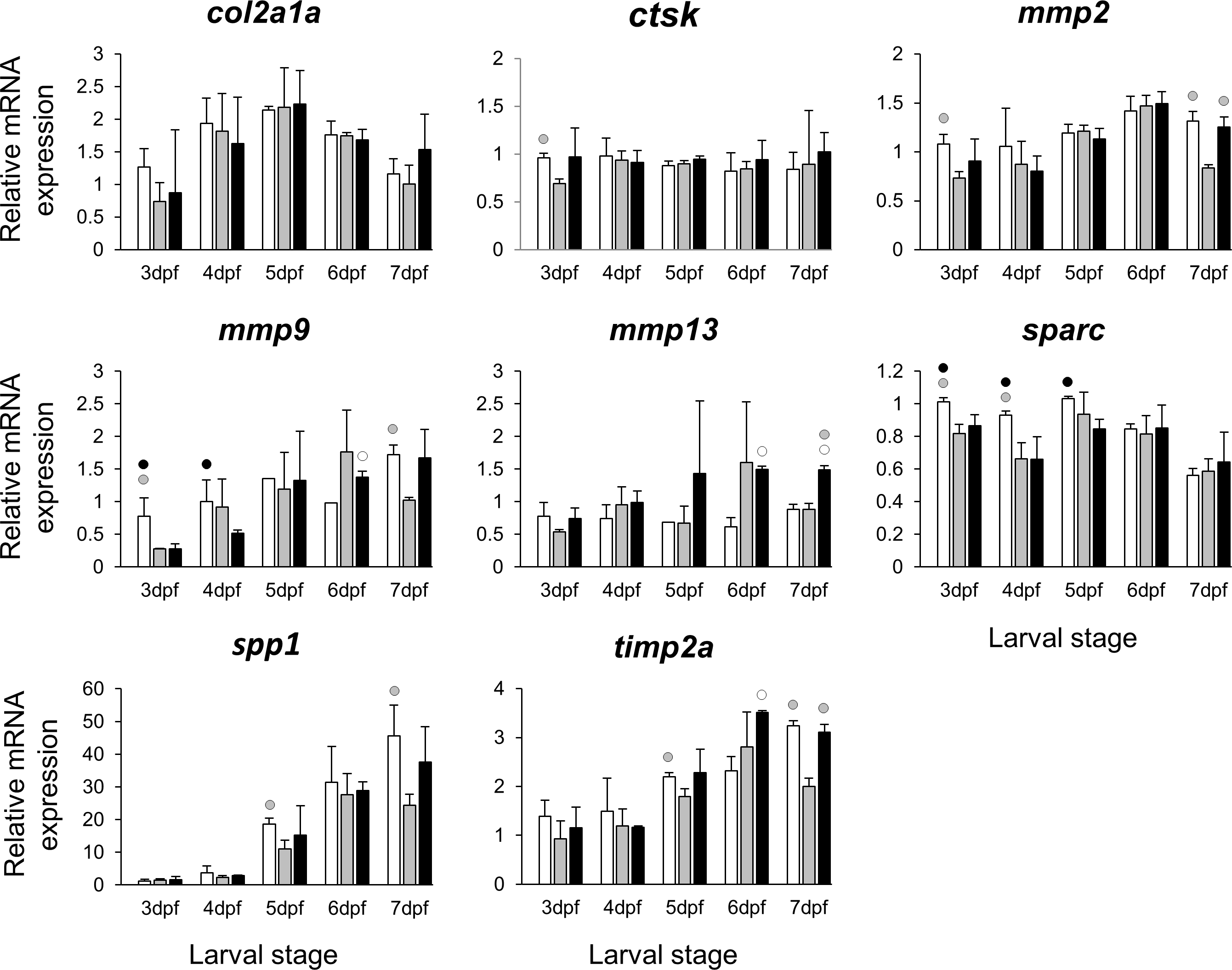{kind=link}
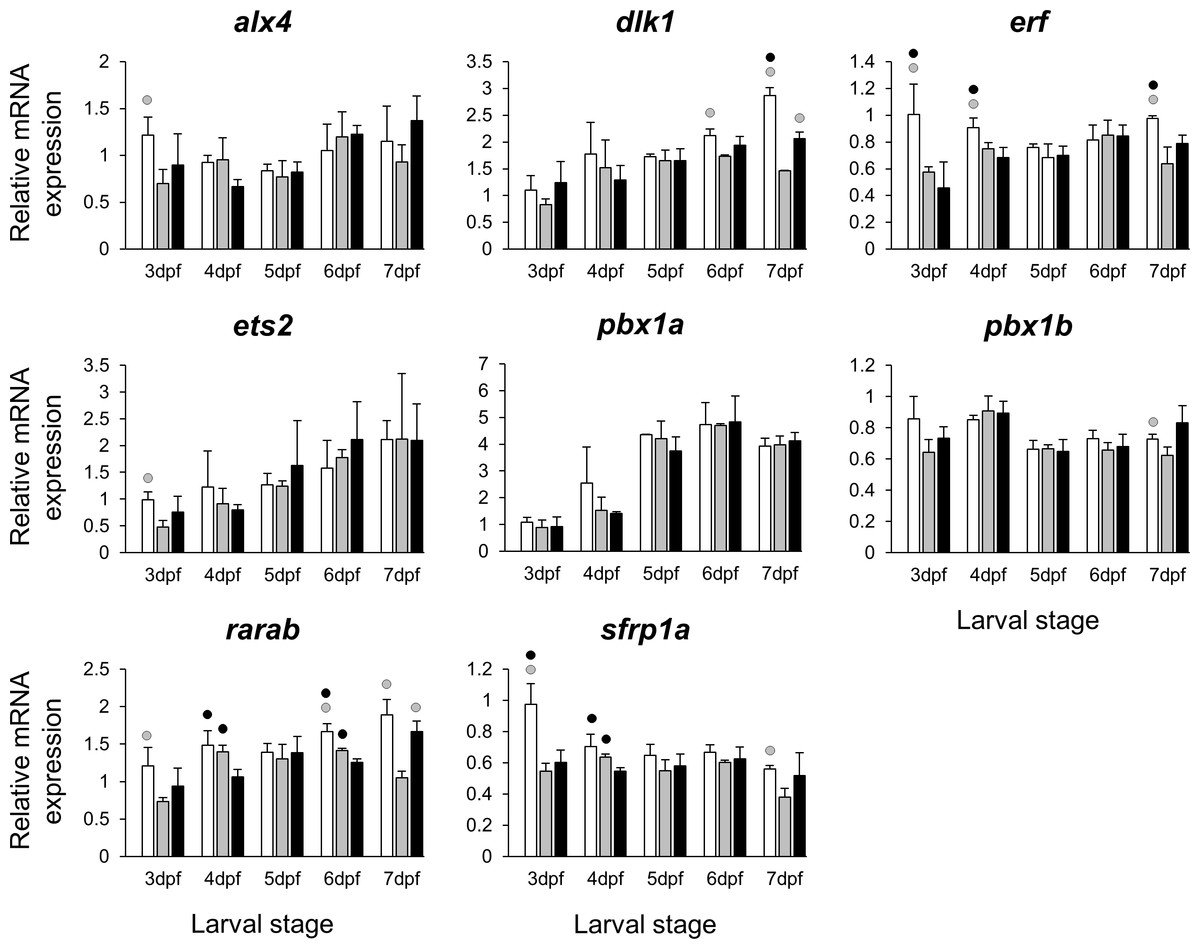
Figure 5: Expression differences of eight other potential targets of estrogen pathway involved in jaw skeletal elongation examined during zebrafish larval head development across control and E2 treated groups.
Expression of alx4, dlx1, erf, ets2, pbx1a, pbx1b, rarab and sfrp1a was examined with qPCR and normalised using three best ranked reference genes (ppia2, rpl8 and tbp). For analysis of relative expression levels for each target gene a replicate of the control group at 3dpf was set to one. The white, grey, and black bars in each graph represent expression levels for control, 2 µM E2 treated and 5 µM E2 treated groups respectively. Statistical differences of each treatment group versus the others are shown in white, grey, and black circles representing higher expressed than control, 2 µM E2 treated and 5 µM E2 treated groups respectively (P < 0.05). Error bars represent standard deviation calculated from three biological replicates. Each biological replicate was made from a homogenate of 30 heads.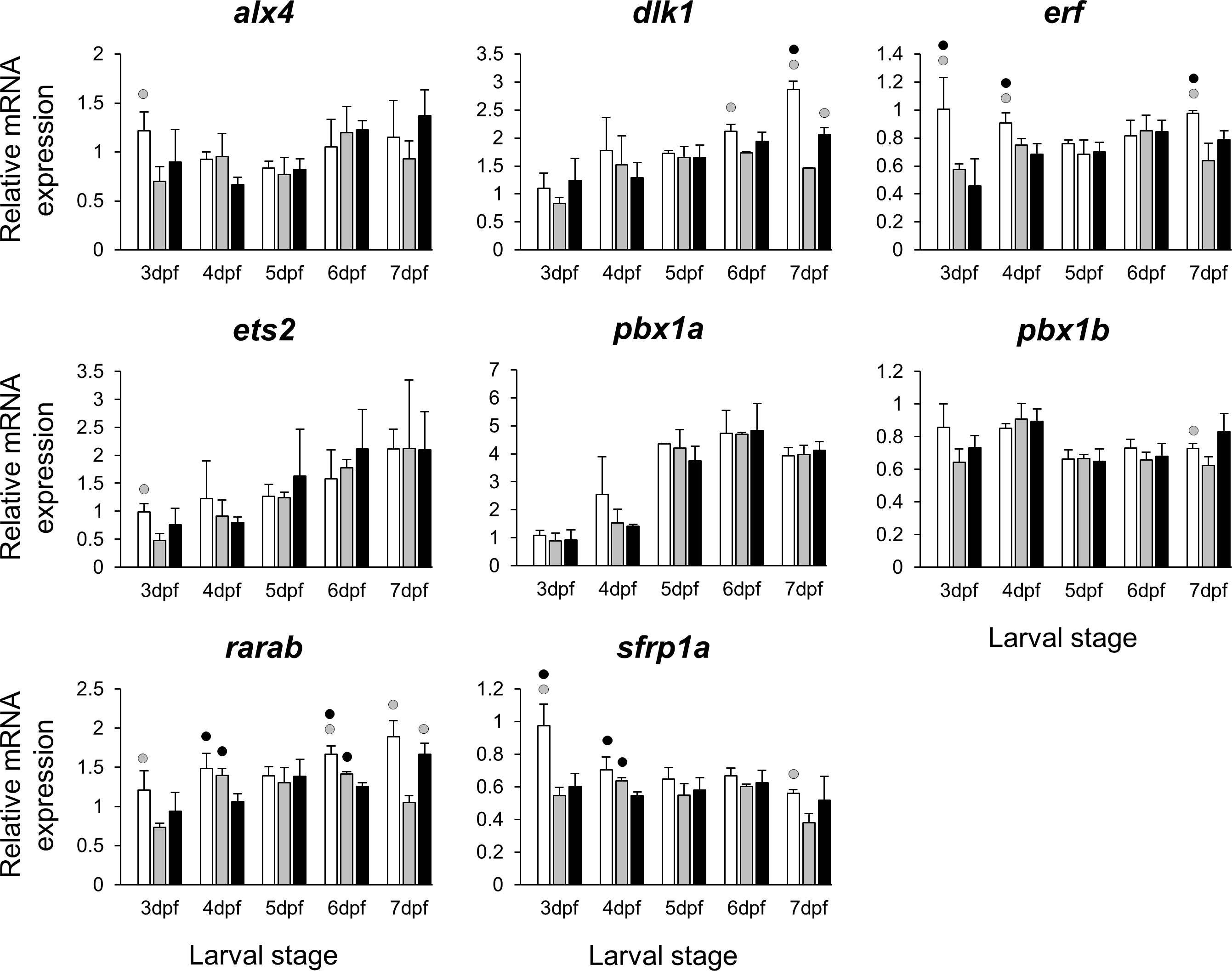{kind=link}
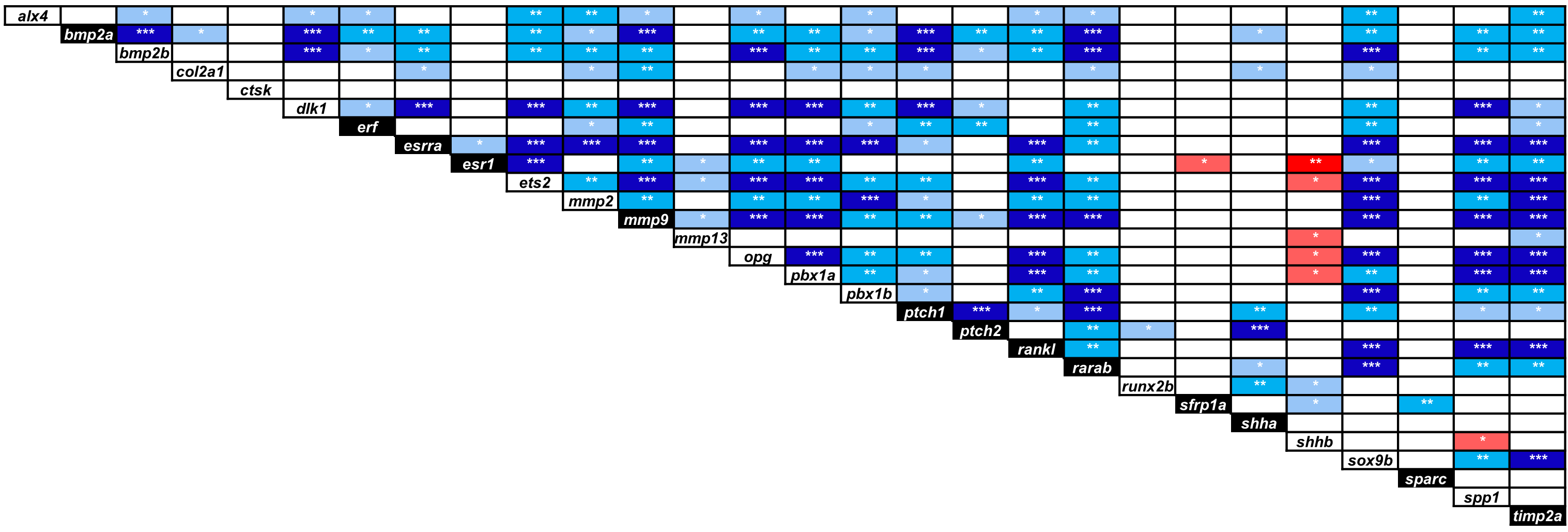
We calculated the Pearson’s correlation coefficient of the expression levels for the target genes over all treatment groups and larval stages and found positive expression correlation between many pairs of target genes (blue shadings in Fig. S2). Some of the genes i.e., mmp9, ptch1, rarab and timp2a displayed positive expression correlation with most of the genes whereas others such as mmp13, sfrp1a, shhb and sparc showed the least correlated expression. Negatively correlated expression was only seen between esr1 and sfrp1a, and between shhb and six genes including esr1, ets2, mmp13, opg, pbx1a and spp1 (red shadings in Fig. S2).
A co-expressed network of genes shows higher expression induction in lower E2 treatment groups
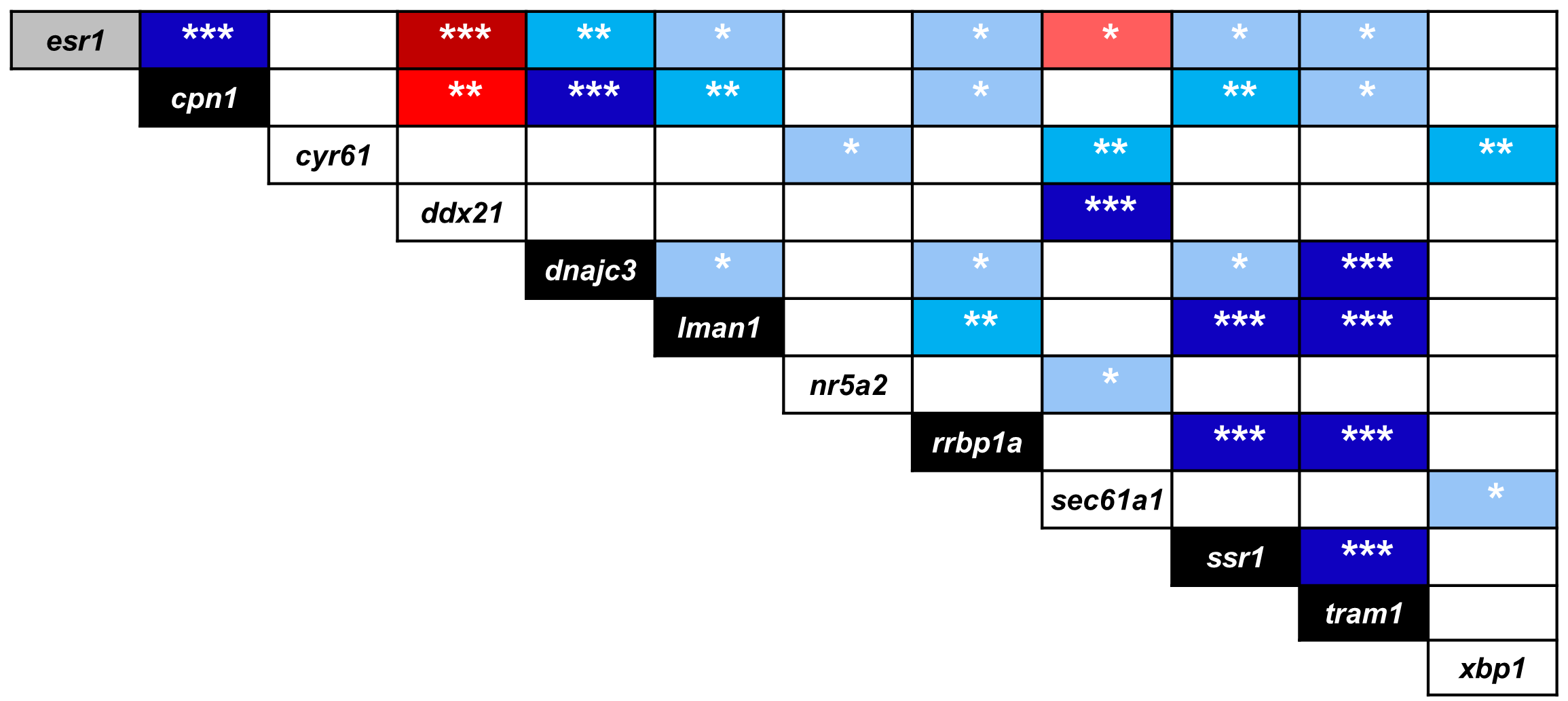
The stronger transcriptional response of esr1 to the lower E2 treatment (Fig. 2) could indicate a distinct regulatory mechanism associated with slight increase in estrogen concentration during zebrafish larval head development. In order to identify additional genes showing similar expression dynamics, we selected 11 candidate genes constructing a co-expression network with esr1 using co-expression data for zebrafish in COXPRESdb (Obayashi & Kinoshita, 2011) (Table S2). These candidates are also known to have craniofacial skeletal expression during zebrafish development based on data submitted to the ZFIN database (http://zfin.org) (Bradford et al., 2011). Strikingly, we found stronger inductive effects of the lower E2 concentration on the expression of six genes, i.e., cpn1, dnajc3, lman1, rrbp1a, ssr1 and tram1 (Fig. 6). The expression of these six genes followed a similar pattern and their higher expression levels were more pronounced at the last three stages of 2 µM treatment groups. Moreover, the gene showing strongest coexpression relationship with esr1 among the candidates, rrbp1a, had shown higher expression levels at the last four stages of 2 µM treatment groups (Table S2 and Fig. 6). Finally, we also demonstrated positive expression correlations between the six candidates and esr1, but not the rest of the non-differentially expressed genes (blue shadings in Fig. S3).
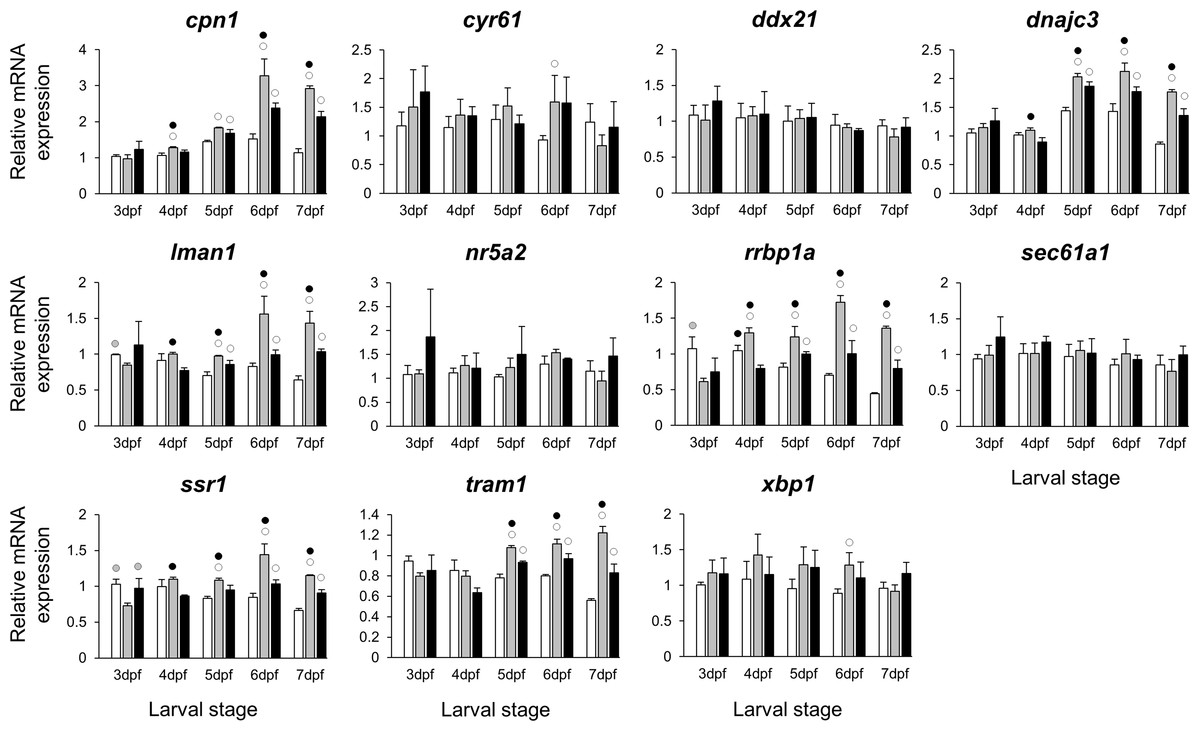
Figure 6: Expression differences of esr1 coexpressed genes in developing heads of zebrafish larvae across control and E2 treated groups.
Expression levels of eleven candidate genes coexpresed with esr1, based on data from COXPRESdb in zebrafish, were examined with qPCR and normalised using three best ranked reference genes (ppia2, rpl8 and tbp). For analysis of relative expression levels for each target gene a replicate of the control group at 3dpf was set to one. The white, grey, and black bars in each graph represent expression levels for control, 2 µM E2 treated and 5 µM E2 treated groups respectively. Statistical differences of each treatment group versus the others are shown in white, grey, and black circles representing higher expressed than in control, 2 µM E2 treated and 5 µM E2 treated groups respectively (P < 0.05). Error bars represent standard deviation calculated from three biological replicates. Each biological replicate is based on a homogenate of 30 heads.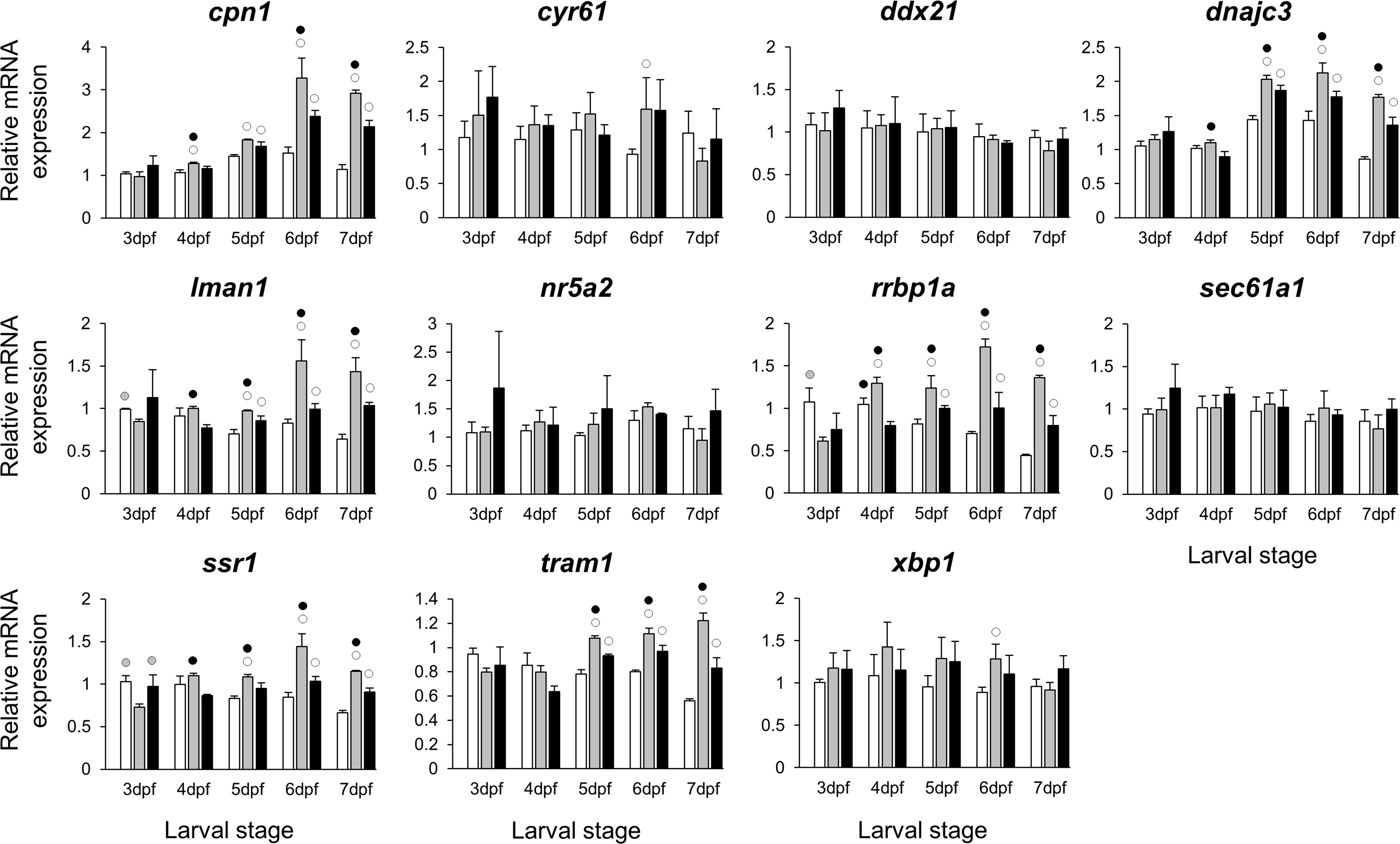{kind=link}
Discussion
Estrogen signaling, through both canonical nuclear estrogen receptors and G-protein coupled receptors, is important in embryonic development (Griffin et al., 2013; Shi et al., 2013). Estrogens can act at autocrine, paracrine, and endocrine distances in the embryo and the adult (Boon, Chow & Simpson, 2010). Aromatase, the enzyme that synthesizes estrogens, is present in the developing brain of many species, including zebrafish (Lassiter & Linney, 2007) and would be a local source of the hormone during head development. In fact, the teleost brain produces relatively high levels of estrogen compared to other vertebrates (Forlano et al., 2001). Estrogens are thus present in the cranium of developing embryos and modulate viscerocranial development (Fushimi et al., 2009; Marquez Hernandez et al., 2011; Cohen et al., 2014). Estrogen signalling has been implicated in the sexual dimorphism of cranial bones in the Anolis lizard (Sanger et al., 2014). Hence, it may play a role in craniofacial morphological divergence among species and within sexes of the same species.
A previous attempt to identify mechanisms underlying the effects of estrogen on zebrafish craniofacial development was conducted with a high concentration of 17-β estradiol (10 µM) giving rise to major disruptions of chondrogenesis followed by severe morphological defects (Fushimi et al., 2009). In the same study, analysis of gene expression after high dose estrogen treatment was limited to a semi-quantitative method (in situ hybridization) and a few chondrogenic genes belonging to only one molecular pathway (Fushimi et al., 2009). We hypothesized that many other candidate genes would be involved and hence, in the present study, we sought to quantitatively assess the expression of genes that could play role in the subtle effects of estrogen on the development of the craniofacial skeleton in zebrafish larvae (Cohen et al., 2014). Since our expression analysis depended on accurate qPCR, a prior step of careful validation of reference genes was essential to acquire reliable results (Bustin, 2000; Kubista et al., 2006). An increasing number of stably expressed reference genes have been validated for qPCR studies in a variety of fish species (Ahi et al., 2013; Fuentes et al., 2013; Liu et al., 2014; Altmann et al., 2015; Wang et al., 2015), and also in zebrafish at different developmental stages, body parts/tissues, and treatments (Tang et al., 2007; McCurley & Callard, 2008; Lin et al., 2009; Casadei et al., 2011). There is however a necessity for validation of reference genes depending on the experimental conditions under study. Here, we found three genes, ppia2, rpl8 and tbp, to be the most stably expressed candidate genes by all the methods of analysis used (Table 2 and Fig. 1) and their combination could ensure robust qPCR data normalisation (Fig. S1). We next selected candidate genes that are shown to be potential estrogen pathway targets, and at the same time, differential regulation of many of them is associated with morphological changes resembling shortened snout in different vertebrates (many found in mammalian species) (see publications referenced in Table 1). The underlying mechanisms by which these candidate genes could affect skeletogenesis are different from each other. For instance, genes like bmp2a/b, rankl, runx2b and sox9b are major factors in differentiation of skeletal cells and some others such as col2a1a, ctsk, mmp2/9/13, spp1 and sparc are critical for the formation of ECM in craniofacial skeletal structures (see Table 1).
The treatments with the two different doses of E2 (2 and 5 µM) resulted in differential expression of many of the candidates during the zebrafish larval head development (Figs. 2, 3, 4 and 5). Consistent with a previous study in zebrafish using higher E2 concentration (10 µM) (Fushimi et al., 2009), we also found significant down-regulation of ptch1 and ptch2 in the heads of fish receiving lower dose estrogen treatments during larval development. These two genes are the receptors (and the upstream mediators) of the hedgehog (Hh) signaling pathway which plays a crucial role in developmental patterning and skeletal morphogenesis (Eberhart et al., 2006; Swartz et al., 2012). Interestingly, slight changes in expression of ptch1 were shown to be associated with subtle craniofacial skeletal divergence (shorter snout and flatter face) in cichlid fish (Roberts et al., 2011; Hu & Albertson, 2014). In addition, we found a strong positive expression correlation between ptch1 and ptch2 (Fig. S2), indicating potential estrogen mediated co-regulation of the two Hh receptors. In the above mentioned study of high dose E2 treatment, the upstream activators of the Hh pathway, sonic hedegehog genes, shha and shhb (twhh), did not show significant changes in expression (Fushimi et al., 2009). However, this could be due to technical limitations such as the use of a semi-quantitative method that is unable to reveal small differences in gene expression (Fushimi et al., 2009). In this study we found small and yet significant down-regulation of shha, but not shhb, in E2 treated groups, as well as positive co-expression of only shha with the two Hh receptors. An important role of the shh in craniofacial skeletogenesis through activation of Hh signalling has been described (Hu & Helms, 1999), but it is not clear whether estrogen directly regulates its expression during development. The small reduction of shha transcripts in developing larval heads might be a result of a decreased number of cells expressing shha and not a direct estrogen mediated transcriptional regulation.
Extracellular matrix remodelling is a critical process in the developmental program of bone and cartilage differentiation and morphogenesis (Werb & Chin, 1998). The spatio-temporal expression of genes encoding matrix metalloproteinases and their tissue inhibitors plays a pivotal role in orchestrating the ECM remodelling process (Werb & Chin, 1998; Page-McCaw, Ewald & Werb, 2007). Moreover, many ECM remodelling genes are downstream targets of pathways mediated by nuclear receptors, including estrogen signalling (Cox & Helvering, 2006; Heldring et al., 2007; Ganesan et al., 2008). The selected ECM remodelling factors (mmp2/9/13, timp2a and sparc) were all reported to be regulated by estrogen signalling (Lehane et al., 1999; Tüshaus et al., 2003; Marin-Castaño et al., 2003; Lu et al., 2006; Nilsson, Garvin & Dabrosin, 2007; Lam et al., 2009; Wang & Ma, 2012) and play role in craniofacial skeletal morphogenesis (Dew et al., 2000; Renn et al., 2006; Hillegass et al., 2007a; Hillegass et al., 2007b; Mosig et al., 2007; Rotllant et al., 2008; Letra et al., 2012; Ahi et al., 2014). Our results revealed slight but significant effects of the estrogen treatments on expression of the selected ECM remodelling genes during larval head development (Fig. 4). It is interesting to note that previous investigations have shown association between differential expression of these genes and craniofacial phenotypes with flatter face and shorter snout (Hillegass et al., 2007a; Hillegass et al., 2007b; Ahi et al., 2014). The mechanism by which estrogen regulates the expression of ECM remodelling genes is not well understood. The estrogen dependent regulation might be exerted through interaction between estrogen-receptors and transcription factors that regulate ECM remodelling genes such as members of Ap-1 complex and ETS factors (Lu et al., 2006; Ahi et al., 2014; Cao et al., 2015). The binding motifs for Ap-1 and ETS transcription factors are present in the promoters of many ECM remodelling genes across vertebrate species (Ahi et al., 2014). Additionally, we found the expression of erf, an ETS repressor and estrogen target (Sgouras et al., 1995) to be down-regulated in both E2 treated groups at three larval stages. Remarkably, a recent study showed that small reduction in expression of erf causes complex craniosynostosis with shortened snout in both human and mice (Frasor et al., 2003; Twigg et al., 2013). The same study also demonstrated regulatory elements containing Ap-1, ETS and Runx motifs as preferential erf binding sites (Twigg et al., 2013). Taken together, the results of the present and previous studies suggest potential estrogen mediated regulation of ECM remodelling genes possibly through interaction with other transcription factors. Other estrogen mediated processes than direct transcriptional regulation cannot, however, be ruled out, as the slight changes in transcript levels of ECM related genes could be due to reduced proportion of skeletal cells expressing these genes in larval heads. It is also important to emphasize that the selected ECM genes can be expressed in other tissues of the head (though at considerably lower levels), thus their expression differences in other tissues might affect the overall changes in expression.
The E2 treatments caused small and variable repressive effects on expression of other selected target genes (Figs. 2, 3, 4 and 5). The genes, bmp2a and rankl, are well characterized skeletogenic markers (Nie, Luukko & Kettunen, 2006; Hu, Colnot & Marcucio, 2008; Lézot et al., 2015) and their regulation by estrogen signalling has been reported in other vertebrate species (Bord et al., 2003; Zhou et al., 2003). It has been shown that treatment with high doses of E2 can reduce the number of skeletal cells in the craniofacial skeleton (Cohen et al., 2014), hence the small changes in transcript levels of skeletogenic markers (e.g., sox9b) may again be caused by a decreased proportion of skeletal cells in the heads. We also found components of retinoic acid and Wnt∕β-catenin signalling patways, rarab and sfrp1a, to be transcriptionally affected by E2 treatment indicating the potential crosstalk of these pathways with estrogen signalling during larval head development (Lohnes et al., 1994; O’Lone et al., 2004; Trevant et al., 2008; Yokota et al., 2008). Although, the selected components of the pathways and transcription factors in this study (Fig. 5) are known to have markedly high levels of expression in the craniofacial skeleton, they might also be expressed to a lesser extent in other tissues within the larval head. Therefore, the observed small changes in expression can not be readily attributed to viscerocranial skeletal elements and further gene expression studies using dissected skeletal elements are essential to confirm this.
In addition to skeletogenic genes, we were interested in investigating the effects of different doses of E2 on the expression of estrogen receptors. Therefore, we assessed the expression of two estrogen receptors, esrra and esr1, that could mediate estrogen signal during the development of skeletal tissues (Bonnelye & Aubin, 2005; Bonnelye et al., 2007; Auld et al., 2012). While the E2 treatments had small and variable repressive effects on expression of esrra, the increased expression of esr1 was observed in both E2 treated groups. Strikingly, the lower E2 concentration (2 µM) resulted in higher induction of esr1 expression. This suggests that the distinct effects of lower doses of estrogen on craniofacial skeletogenesis, described by Cohen et al. (2014), might be mediated by esr1, however further functional studies are required to demonstrate such a role. To identify genes sharing regulatory mechanisms in response to slight increases in estrogen levels, we further explored the expression of 11 genes constructing a co-expression network with esr1 (Table S2 and Fig. 6). These candidate genes were selected by using a vertebrate co-expression database (Obayashi & Kinoshita, 2011) which we have successfully used for identification of gene networks associated with subtle craniofacial morphological divergence in another teleost (Ahi et al., 2014; Ahi et al., 2015). Our results indicate higher transcriptional induction of six genes, i.e., cpn1, dnajc3, lman1, rrbp1a, ssr1 and tram1 in the lower (2 µM), than the moderate (5 µM) treatment groups, during craniofacial development. The genes also showed positive expression correlation with esr1 suggesting a common regulatory mechanism mediated by estrogen during head development. To our knowledge, a mechanism by which a lower concentration of estrogen can have stronger inductive effects on expression of certain genes than higher concentrations is not known. Such a mechanism might be involved in distinct regulation of estrogen receptors by different concentrations of estrogen hormone, which in turn could lead to recruitment of the receptors to distinct genomic binding sites and/or with different binding affinity (Stender et al., 2010). Among the six genes only dnajc3, a gene encoding protein kinase inhibitor P58 (P58IPK), has been shown to be involved in skeletogenesis through regulation of a cytokine-dependent cartilage degradation (Gilbert et al., 2014). Although all of the six genes have recorded developmental expression patterns in zebrafish craniofacial elements based on data in the ZFIN zebrafish database (Thisse et al., 2001; Thisse et al., 2004), their roles in craniofacial morphogenesis have yet to be investigated. Finally, an unbiased approach such as transcriptome sequencing rather than candidate gene-based study would be warranted to provide better knowledge of estrogen mediated effects on expression of genes with unknown roles in craniofacial morphogenesis as well as links between already identified genes and molecular pathways involved.
Conclusions
In this study we quantitatively assessed the effects of two doses of estrogen (2 µM and 5 µM) on gene expression during zebrafish larval head development. We performed a highly sensitive and specific qPCR analysis and carefully validated reference genes. We assessed the expression of a selected set of genes involved in craniofacial skeletal development as well as genes coexpressed with esr1, an estrogen receptor showing stronger inductive response to 2 µM than 5 µM estrogen concentration. The results implicate estrogen in the expressional regulation of genes belonging to distinct signalling pathways such as hedgehog and retinoic acid pathways, as well as genes involved in ECM remodelling during craniofacial development. Furthermore, estrogen mediated transcriptional changes in a few tested major skeletogenic factors (e.g., bmp2a and rankl), and a transcription factor, erf, with a demonstrated role in the formation of a shortened snout phenotype in human and mouse. Finally, we identified a gene network showing positive expression correlation with esr1 and higher induction in response to treatment with 2 µM than with 5 µM estrogen. This could suggest a co-regulated module of genes mediating the effects of low doses of estrogen during craniofacial development which is required to be further investigated at functional level.
Supplemental Information
Optimal number of reference genes for normalisation analysed by geNorm
The algorithm used in geNorm software determines the optimal number of reference genes in developing heads of zebrafish larvae for all the treatments together (a control and two E2 treated groups). Average pairwise variations (Vn∕n+1) were calculated using the genes ranked according to geNorm. The recommended cut-off value of 0.15 is shown by a black line. Below this line the benefit of using an extra reference gene is limited.
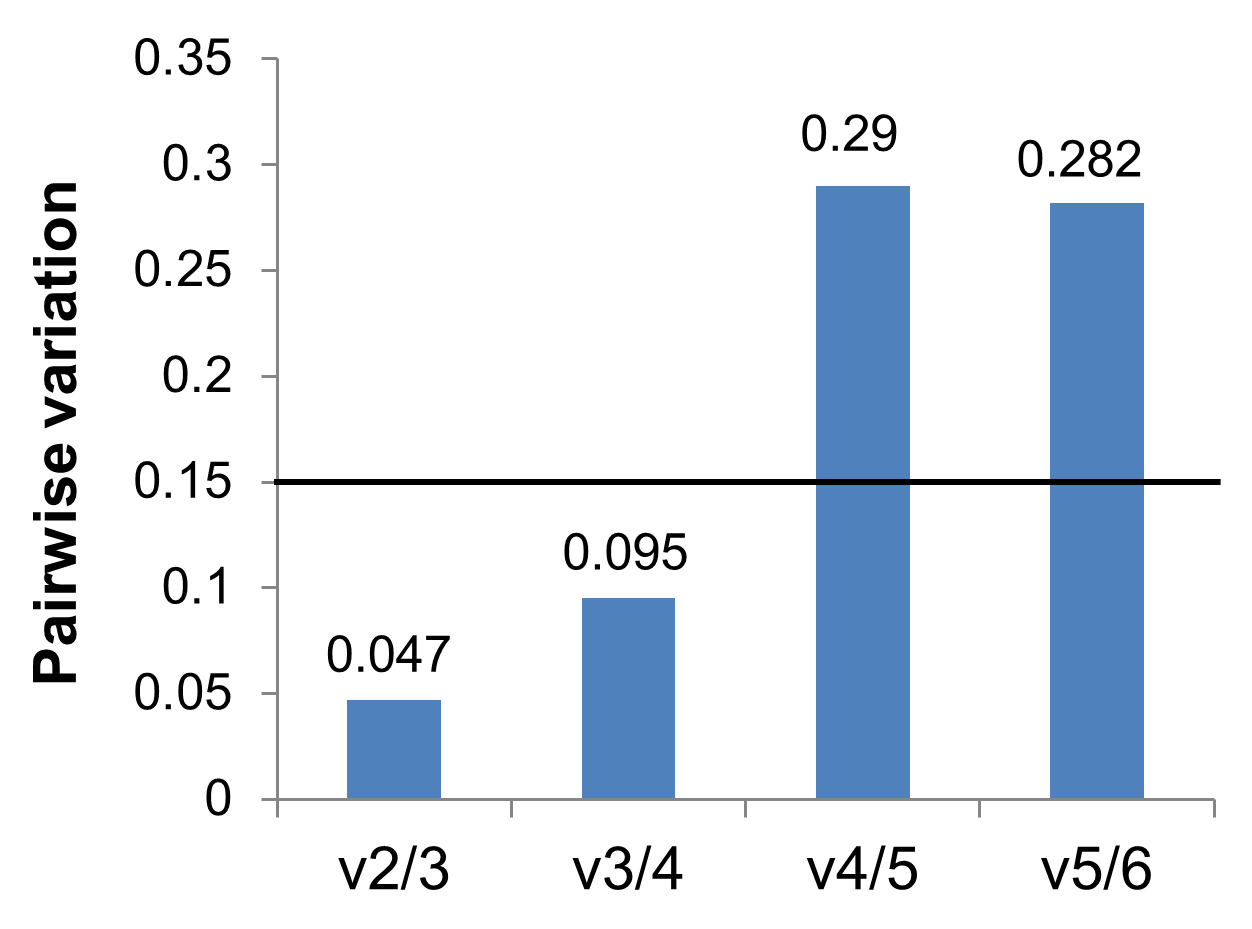{kind=link}
Significant expression correlations between selected target genes in developing heads of zebrafish larvae
Pearson correlation coefficient (r) was used to assess the pairwise expression similarity between the candidate target genes. Blue shading represents positive and red shading represents negative expression correlation and the colour gradients indicate correlation coefficients (r) above critical values (2-tail; df = 13). P-values of <0.05, 0.01 and 0.001 are indicated by one, two and three asterisks, respectively. The genes showing differential expression between the treatment groups in at least three larval stages are represented in black boxes.
{kind=link}
Significant expression correlations between selected esr1 co-expressed genes in developing heads of zebrafish larvae
Pearson correlation coefficient (r) was used to assess the pairwise expression similarity between the candidate target genes. Blue shading represents positive and red shading represents negative expression correlation and the colour gradients indicate correlation coefficients (r) above critical values (2-tail; df = 13). P-values of <0.05, 0.01 and 0.001 are indicated by one, two and three asterisks, respectively. The genes showing differential expression between the treatment groups in at least three larval stages are represented in black boxes (except esr1 gene).
{kind=link}
Zebrafish specific primers used for qPCR expression analysis
Ranking of genes coexpressed with esr1 in zebrafish
A list of genes coexpressed with esr1 in zebrafish was retrieved from COXPRESdb after they were filtered by setting the mutual rank (MR) to the top-ranked 2000 and the Supportability score of minimum 1. The expression pattern of the genes during craniofacial skeletal development was checked in a zebrafish database for gene expression (ZFIN.org).