
Enrichment effects on growth, health, and reproduction in a single clone of the asexual Amazon molly, Poecilia formosa
- Published
- Accepted
- Received
- Academic Editor
- María Ángeles Esteban
- Subject Areas
- Animal Behavior, Aquaculture, Fisheries and Fish Science, Ecology, Environmental Impacts
- Keywords
- Welfare, Social and physical enrichment, Laboratory fish, Poeciliid
- Copyright
- © 2024 Cunningham et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits using, remixing, and building upon the work non-commercially, as long as it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2024. Enrichment effects on growth, health, and reproduction in a single clone of the asexual Amazon molly, Poecilia formosa. PeerJ 12:e18734 https://doi.org/10.7717/peerj.18734
Abstract
This study examines the impact of two types of environmental enrichment on the growth of the Amazon molly (Poecilia formosa), a clonal fish species. We investigated the effects of two welfare-related enrichment factors: physical enrichment (presence of a halfpipe PVC and gravel substrate, E) and social enrichment (presence of visible neighbor fish, N). Fish were divided into four treatment groups: (1) both physical and social enrichment (EN), (2) no physical enrichment but social enrichment (nEN), (3) physical enrichment without social enrichment (EnN,), and (4) no enrichment (nEnN). Results showed that individuals, which received both types of enrichment, exhibited faster growth rates and lower early life mortality compared to the other groups, indicating improved health and welfare. However, this initial advantage did not persist into adulthood, suggesting that the benefits of enrichment may be stage specific. These findings highlight the importance of environmental factors in promoting welfare in a particular species, especially during early developmental stages. This study may have implications for early life-stage fish research indicating that some environmental factors may play a role in enhancing growth, reproductive competence, and reduced mortality. Enrichment effects on growth, health, and reproduction in a single clone of the asexual Amazon molly, Poecilia formosa.
Introduction
The Five Freedoms, universally recognized as minimum standards of care for animals under human supervision, originated from an inquiry into the treatment of farm animals (Brambell, 1965). The United Kingdom’s Farm Animal Welfare Council later formalized these principles. In 2016, Mellor noted that while the Five Freedoms focused on alleviating negative experiences (‘freedom from’) and providing measures to address them, this approach mainly supported animal survival without promoting thriving (Mellor, 2017). The current model, The Five Domains, expands this standard by recognizing the interaction of physiological function and mental well-being and includes the potential consequences of various conditions. Mellor further expounded on each domain in 2020, changed Domain 2 from Environment to Physical Environment, and expanded Domain 4 from Behavior to Behavioral Interaction (Mellor et al., 2020). In their current conception, each Domain is described as follows: Domain 1, Nutrition considers the nature, quantity, and quality of the food and water available to the animals. Domain 2, Physical Environment refers to the physical and environmental/climatic conditions surrounding the animal and includes shelter, space cleanliness, temperature, substrate, odor, noise. Domain 3, Health focuses on the impacts of physical health, injury, and physical fitness on animal welfare. Domain 4, Behavioral Interactions emphasizes the animal’s conscious pursuit of specific goals when behaviorally interacting with the environment, other non-human animals, humans, and includes the display of natural behaviors of that species such as foraging, exploring, and playing, as well as interactions with conspecifics, other animals, and caretakers. Domain 5, Mental state focuses on the psychological state of the animal that is influenced by its subjective experiences from other domains. The interrelation of these domains cannot be understated, and by considering all domains together, animal welfare can be assessed and improved in a holistic manner. In the animal research community, significant attention is given to animal welfare, guided by the principles of the 3Rs, first introduced in 1959 (Russell & Burch, 1959). These principles include replacement, which advocates for the use of non-animal alternatives when possible; reduction, which focuses on minimizing the number of animals used while maximizing the quality and quantity of data obtained; and refinement, which involves modifying husbandry and research practices to reduce or eliminate pain, distress, and discomfort in animals, thereby improving their overall welfare. One key aspect of refinement is environmental enrichment-the practice of enhancing animals’ surroundings to promote their psychological well-being. Over time, environmental enrichment has become a well-studied and widely applied strategy in research settings. Although many studies lacked vigorous design, there was evidence that enrichment activities did have health and wellbeing benefits (Lovell et al., 2015). Providing environmental enrichment has been shown to decrease abnormal behaviors associated with stress, fear, and anxiety (Bayne, 2018). Evidence suggests that the brain functions responsible for abnormal repetitive behaviors (ARB) impact behavioral studies, and incorporating enrichment strategies could enhance their reliability, validity, and reproducibility (Garner, 2005). In 2013, a study assessing pain in zebrafish revealed two key findings. First, when given a choice between two environments-one enriched and the other barren with bright lights-the zebrafish consistently chose the enriched environment. Within this cohort, a smaller group of zebrafish was injected subcutaneously with a noxious substance, while another group was injected with saline. Both groups continued to prefer the enriched environment. However, when an analgesic was introduced to the barren chamber, only the fish experiencing pain shifted their preference to the barren environment thus forfeiting the enriched environment for pain relief (Sneddon, 2013, 2015). Although the provision of environmental enrichment has been shown to lower stress, increase immunocompetency, and lower cortisol in captive salmonids (Webster et al., 2020), most work has focused on traditional animal models such as mice (Bayne, 2018; Kappel, Hawkins & Mendl, 2017), rats, and zebrafish (Schroeder et al., 2014). Poor environmental conditions have been shown to result in reduced weight and poor body condition in production animals (Li et al., 2024; Albernaz-Goncalves, Antillon & Hotzel, 2022), decreased poultry egg production (Bist et al., 2023), and impaired weight gain and broiler chicken performance (Harris et al., 1975). Studies of environmental enrichment on hatchery-reared fish demonstrating improvements in various physiological and behavioral traits including growth performance are well documented (Li et al., 2023; Kientz & Barnes, 2016). In a study investigating the effects of social rearing environments on the growth and behavior of juvenile Pelvicachromus taeniatus, it was found that juveniles reared in groups were, on average, larger than those reared in isolation (Hess & Thünken, 2014). Furthermore, Kasper–Hauser experiments in another species of molly (Poecilia mexicana) showed that males raised in complete isolation exhibited typical male behaviors (Parzefall, 1969), showing that lack of enrichment does not always have strong negative effects. Clearly, the effectiveness of environmental enrichment and social environment is partly dependent on the fish species, life stage, and specific enrichment structure used. Since animal welfare itself is complex and direct measurement not feasible, using physiological parameters as indicators of animal welfare is a widely accepted practice (Zhang et al., 2023). The view that animals in a good welfare state are more likely to explore the unknown aligns with the idea that animals experiencing stress or discomfort may exhibit reduced exploration and activity levels. This concept could provide valuable cues for assessing animal welfare in various contexts. Preference testing, a more recent approach, may offer insight by allowing animals choice assuming they will select what is in their best interests. Of all fields of inquiry, good animal welfare is particularly important in studies of animal behavior. The underlying assumption is intuitive: if animals are not experiencing wellbeing, they will not perform natural behaviors, thus rendering studies with such animals problematic and potentially misleading.
Consequently, behaviorists have a strong interest in providing optimal conditions which facilitate good welfare for their study animals (Rutz & Webster, 2021; Webster & Rutz, 2020); but interestingly, due to the lack of research at the intersection of animal welfare and behavior, standardized protocols for many fish do not exist. Establishing such protocols would not only enhance the validity and reliability of research findings but also ensure ethical treatment of the animals involved. Social isolation is known to negatively impact aspects of social behavior and development (Matsuo, 2018; Schausberger, Gratzer & Strodl, 2017). This underscores the crucial link between environmental conditions, social interactions, and animal well-being, highlighting the importance of considering these factors in scientific research. The impact of enrichment upon Amazon mollies (Poecilia formosa) development has not been well studied. We postulated Amazon mollies provided with both physical and social enrichment would have a significant advantage regarding health, size, mass, and reproductive capability. This hypothesis suggests that environmental factors play a significant role in shaping the developmental trajectory and reproductive outcomes of Amazon mollies.
The Amazon molly originated from a natural hybridization event (Hubbs & Hubbs, 1932), ca. 100,000 years ago (Schartl et al., 1995), are all female, but require sperm of males from a syntopic sexual species to fertilize their diploid eggs (Schlupp, 2005, 2009). Amazon mollies are ameiotic and have no crossing over, but a recent study showed that they initiate the pachytene (Dedukh et al., 2022). Hence, they do not have functional meiosis, making them clonal and genetically relatively uniform (Schlupp, 2005), but with behavioral individuality documented even in near-identical rearing conditions (Bierbach, Laskowski & Wolf, 2017).
Materials and Methods
Animal experimentation was granted by the University of Oklahoma IACUC under protocol number R21-004. The sample fish (n = 80) for this experiment were the offspring of nine randomly selected Amazon mollies, born between May 8, 2021, and August 3, 2021. Fish were eligible for inclusion if they displayed normal swimming behavior, food consumption, and had a known birthdate. The nine selected mollies were obtained from the International Stock Center for Livebearing Fishes, part of the Aquatic Research Facility in the Department of Biology at the University of Oklahoma. The nine fish were housed individually in 40-L freshwater tanks, each with its own filtration system, fed ad libitum and were monitored daily for offspring. To eliminate genetic diversity resulting from sexual reproduction and meiosis, a single clone of the Amazon molly, identical to the one used in sequencing the Amazon molly genome was used (Warren et al., 2018). This approach ensured that any measurable differences in mass, growth, and successful reproduction between individual fish across treatment groups could be attributed to environmental experiences rather than genetic variability. The identity of mothers was not tracked because of the clonality of this species. However, the offspring were randomly assigned to the four treatment groups. The offspring were observed daily, had weekly water changes, and fed ad libitum a commercially available flake food (TetraMin), supplemented once a week with frozen brine shrimp nauplii, Daphnia and bloodworms (mosquito larvae) until they reached 30 days of age. We decided not to handle the offspring right after birth to avoid stress and potential harm to the offspring. After giving birth, the mothers were moved to a new 40 L tank, and the offspring were raised in their home tank with filtration, gravel, and plants until day 30 when they were randomly assigned to their treatment groups. The conditions until day 30 were as uniform as possible, but we did not socially isolate the fish until day 30. After 30 days, 80 sample fish were isolated and kept individually in randomly assigned 5 L freshwater transparent glass tanks with individualized filtration systems in rows across a dedicated rack in a temperature-controlled greenhouse under natural light conditions. Ambient temperatures fluctuated between 26 °C to 32 °C and since tanks did not contain heaters, water temperature corresponded closely with ambient temperatures and were similar between tanks. Amazon mollies in nature experience wide natural fluctuation in temperature (Fischer & Schlupp, 2009) and were well adapted to the temperature conditions provided.
Tanks were set up to accommodate different treatment groups and fish were divided into four treatment groups with 20 fish each. Treatment group 1 was provided physical enrichment (E) consisting of a monolayer of natural colored gravel and an 8 cm by 3 cm gray halfpipe for cover as well as social enrichment (N), which consisted of the ability to see a neighbor fish in a separate tank; (EN). Treatment group 2 did not have physical enrichment but had social enrichment; (nEN), treatment group 3 was provided physical enrichment only (EnN), and treatment group 4 had no physical or social enrichment; (nEnN). Solid dividers were placed between the tanks to block sight of adjacent tanks for treatment groups without social enrichment (EnN and nEnN). Test-fish were observed daily. Fish that were ill (n = 4) were humanely euthanized using an overdose of buffered Tricaine S and fish that died (n = 9) were removed from the sample (see Table 1 for a summary). If removal of a fish led to the neighboring tank being empty, a replacement Amazon molly was placed in the emptied tank to continue the social enrichment treatment. When a neighbor fish was needed, the tank was cleaned prior to the new fish being introduced. We measured the individuals throughout their adult life to investigate potential effects of enrichment on adults (when they are used in most studies). To obtain a tight granular timeline we measured the fish monthly. Fish size and mass were recorded every 30 days beginning on day 90 through day 300. Fish were not measured prior to day 90 due to the inherent harm associated with handling small fish. Individuals were dip-netted out of their tank and placed on a balance (Ohaus Portable Balance, PS121). After weighing (in grams), the fish was placed on a laminated millimeter grid, where the standard length (snout to the end of the spinal column) was recorded to the nearest millimeter. This was performed without anesthesia and a trained observer was able to measure an individual fish in less than 10 s. After obtaining measurements, the fish was immediately returned to its tank.
| Treatment group | Mortality rate day 90–180 | Mortality rate day 181–300 |
|---|---|---|
| EN | 1 | 1* |
| EnN | 0 | 1* |
| nEN | 3 | 0 |
| nEnN | 6 | 0 |
Note:
Reproduction
When fish surpassed 30 mm in length and were considered sexually mature, 40 randomly selected fish representing all treatment groups were paired with a male Poecilia mexicana or Poecilia latipinna. These male species are natural host species of Amazon mollies and known to successfully induce embryogenesis in Amazon mollies (Hubbs & Hubbs, 1932; Ryan et al., 1996; Cerepaka & Schlupp, 2023). Males were allocated randomly and based on availability (see Table 2 for details). All pairings lasted for at least 30 days to correspond with the sexual cycle of the mollies (Parzefall, 1973). Twenty-nine fish were paired with a P. mexicana male from the Rio Oxolatan, an allopatric population for Amazon mollies, and eleven fish were paired with a P. latipinna males from two sympatric populations. The number of broods delivered, the number of offspring per brood, and time between two broods were recorded for each fish (Table 1). The birth date of any newborn was recorded, and the newborns were removed from the parental tank and placed in holding tanks.
| Treatment group | N = | Number of fish bred |
P. mexicana males |
P. latipinna males |
Brood 1 | Brood 2 | Brood 3 | Offspring |
|---|---|---|---|---|---|---|---|---|
| EN | 14 | 1 | 12 | 3 | 1 | 1 | 0 | 26 |
| EnN | 11 | 5 | 9 | 2 | 5 | 2 | 1 | 92 |
| nEN | 8 | 0 | 3 | 4 | 0 | 0 | 0 | 0 |
| nEnN | 7 | 3 | 5 | 2 | 3 | 3 | 0 | 58 |
| Total | 40 | 9 | 29 (9) | 11 (0) | 9 | 6 | 1 | 176 |
Social preference test
On day 300 we performed a standard binary choice test (e.g., Makowicz et al., 2016) to evaluate social preference. A 40-L tank with a gravel substrate was divided into three equal compartment sizes depicted with solid lines drawn on the outside of the tank. Stimulus fish were placed on opposite ends of the tank in clear perforated plexiglass columns. One end contained two stimulus fish; the other end contained a single stimulus fish. The test fish, selected randomly, was placed in a non-perforated plexiglass column in the center of the tank (Fig. 1). After allowing the molly 10-min for acclimation, the test fish column was removed allowing the test fish free association with the stimulus fish for 5 min. Using two different stopwatches, time spent in the preference zones were recorded when the test fish’s eye crossed the solid lines drawn on the front of the tank towards the stimulus fish. Only time spent in the preference zones was recorded.

Figure 1: Preference test tank image.
{kind=link}
After 5-min, the test fish was returned to the cylinder in the center, stimulus fish on each end were swapped to opposite ends and the procedure repeated. Stimulus fish were Amazon mollies from a different clonal lineage (3N) and were visually matched for size with the test fish before utilization. All trials were video recorded using a digital Nikon D5200 camera with a Nikon DX 18-55 mm lens. Videos and photographs are available on request. Stimulus fish were measured as previously described and returned to their stock tank. It is possible they may have been randomly reused in subsequent social preference tests. All surviving fish underwent the same social preference test procedure and after each trial, were humanly euthanized with an overdose of buffered Tricaine-S, weighed, photographed on a millimeter grid, and subsequently placed in formaldehyde for further postmortem analysis.
Statistical treatment
All statistical analyses were conducted in SPSS 29. All p-values are two-tailed. For frequency data we used Chi2 tests. We also used unpaired t-tests for the data on reproduction and paired t-tests for the social choice data. Furthermore, non-parametric Kruskal-Wallis tests with treatment groups as independent variable and the size and mass measurements we obtained as dependent variables were conducted. Post-hoc pairwise comparisons between treatment groups were Bonferroni corrected for multiple comparisons.
Results
Size and mass
Fish length and mass measurements began on day 90, after 60 days in their respective assigned treatment groups. At this initial measurement, fish with physical enrichment were larger in size than fish without physical enrichment (Kruskal-Wallis test: N = 78, DF = 3, H = 20.17, p < 0.001). Pairwise post hoc comparisons (corrected for multiple comparisons) revealed that EN fish were larger than nEnN fish (p = 0.001) and nEN fish (p = 0.007); furthermore, EnN fish differed significantly from EnN fish (p = 0.018). Furthermore, the fish showed a significant treatment effect for mass (Kruskal-Wallis test: N = 78, DF = 3, H = 16.6, p < 0.001). Pairwise post hoc comparisons (corrected for multiple comparisons) revealed that EN fish differed significantly from nEnN (p = 0.005) and EnN from nEnN fish (p = 0.013). At day 300, there were no detectable differences in weight or size.
Growth rates
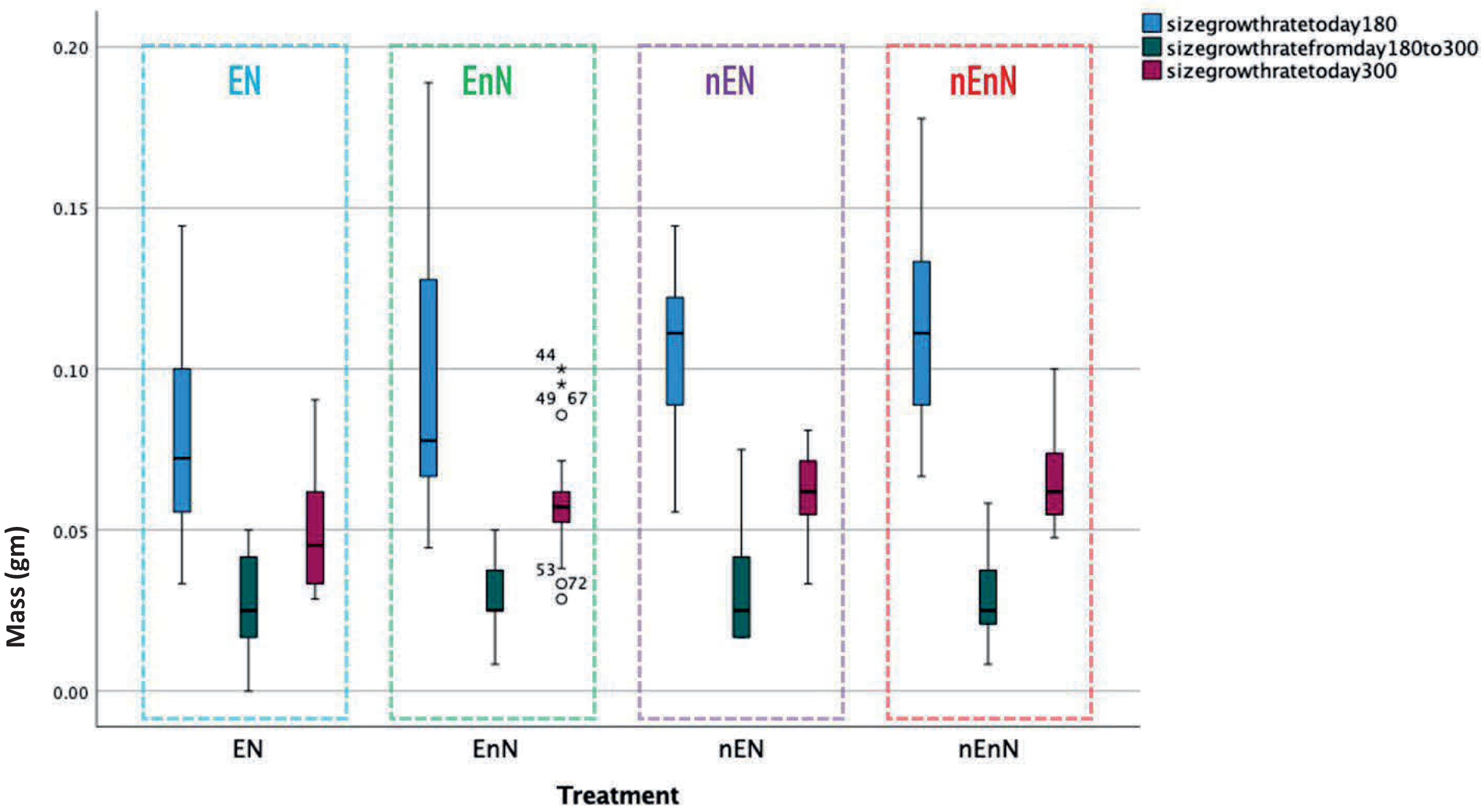
We recorded an increase in body size standard length (mm) (Fig. 2) and mass (g) (Fig. 3) over study time. In principle, fish have indeterminate growth, but independent of the treatment, the raw growth slowed down considerably after day 210. Weight initially increased but also plateaued around day 210 in two of the treatment groups, EnN and nEN. Raw growth rates reveal how the fish responded to different treatments, highlighting the impact of welfare conditions on their growth and maturation. The fish from the two treatments without social enrichment were smaller on day 90 but compensated for this with a higher growth rate after day 180, with growth rates similar for all groups from day 90 to 300 (Fig. 4).
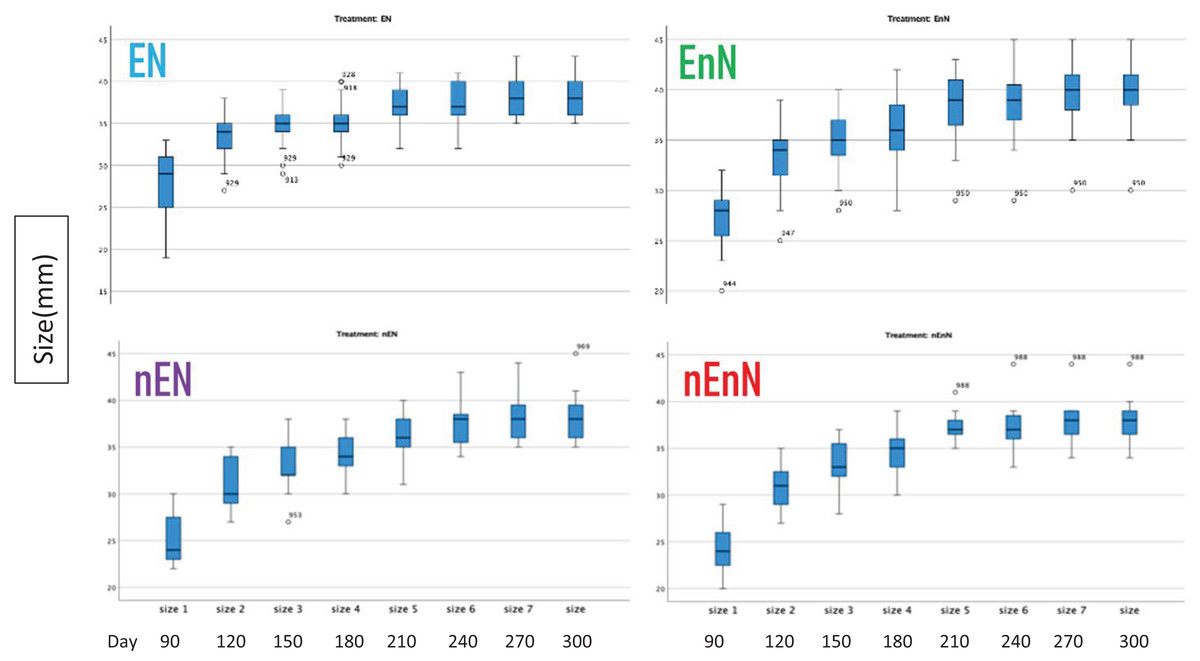
Figure 2: Increases in fish size (mm) by treatment through course of study.
{kind=link}
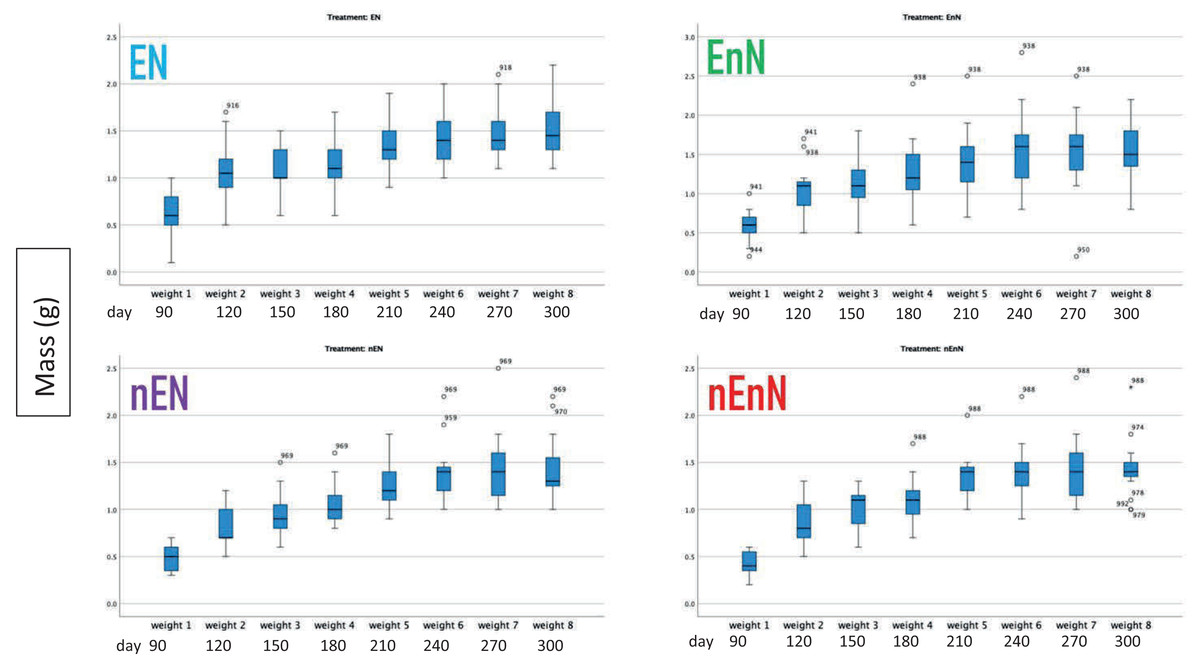
Figure 3: Increases in fish mass (g) by treatment through course of study.
{kind=link}
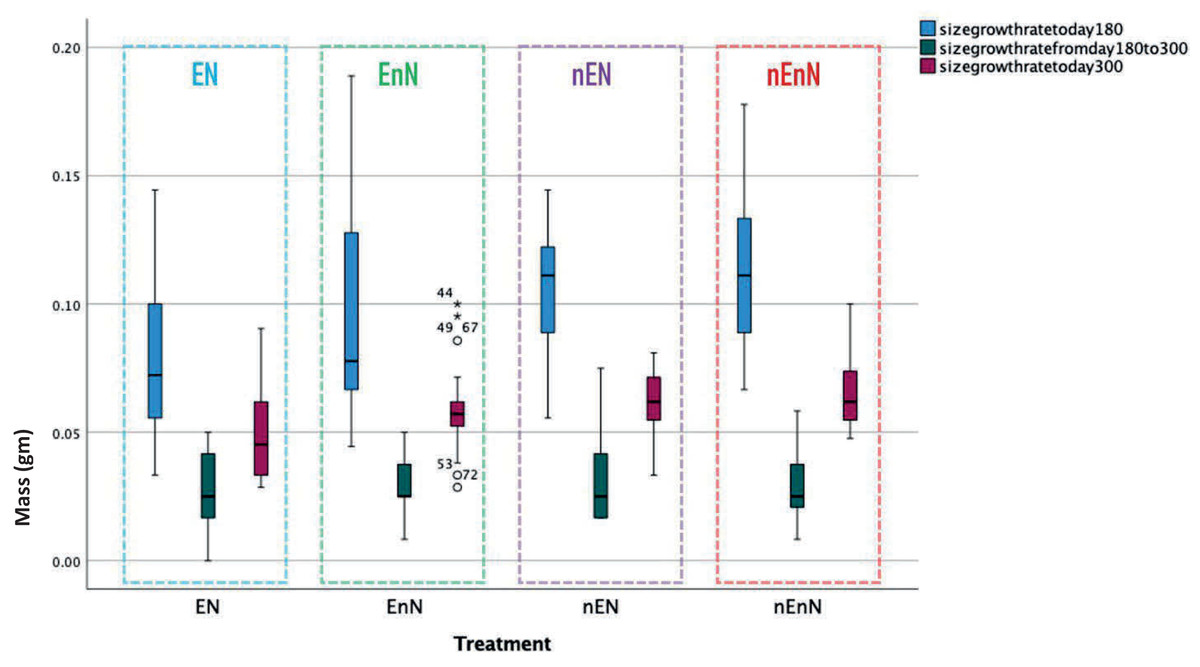
Figure 4: Comparative growth rates for early development (until day 180, blue), late development (from day 180 to day 300, green), and overall growth rate (from day 90 to day 300, mauve) for fish in different treatment groups.
{kind=link}
Since physical measurements spanned an extended period, we also express growth rates as the intrinsic growth rate for each treatment population. Intrinsic growth rate (G) was calculated as G = (logeY2-logeY1)/(t2 - t1), where (Y) is the size or mass at given time and (t) is the time in question. We used the nonparametric Kruskal-Wallis H-test to compare the treatments at an early phase of the project (day 90 to 180) and at a late phase (day 180 to 300) (Fig. 4). In the early phase of the project (day 90 to 180), there were significant differences in the intrinsic growth rate between treatments for size (Kruskal-Wallis H-test: N = 67, DF = 3, H = 9.25, p = 0.026), but not for mass (Kruskal-Wallis H-test: N = 68, DF = 3, H = 5.4, p = 0.145), (Figs. 5, 6). Post hoc tests (corrected for multiple comparisons) for size showed that the treatments EN and nEnN differed significantly (p = 0.023) from each other. Neither size nor mass intrinsic growth rate differed significantly in the late phase of the project (day 180–300).
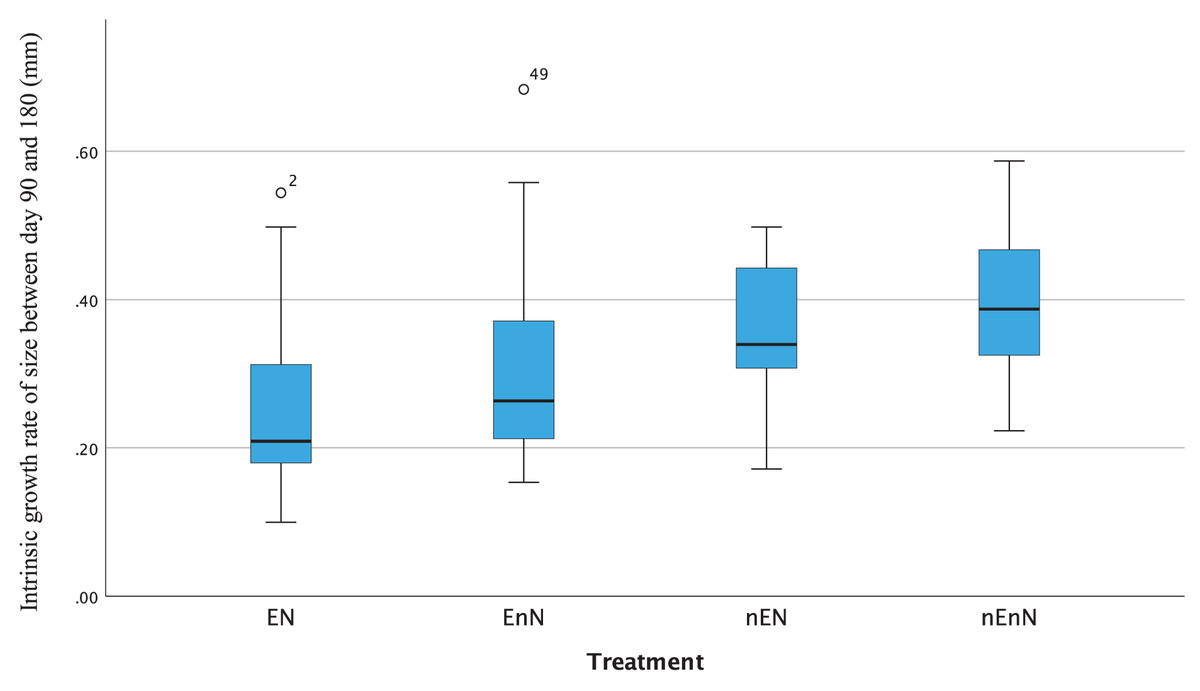
Figure 5: Significant differences in intrinsic growth rates for early development (day 90–180) based on size by treatment group.
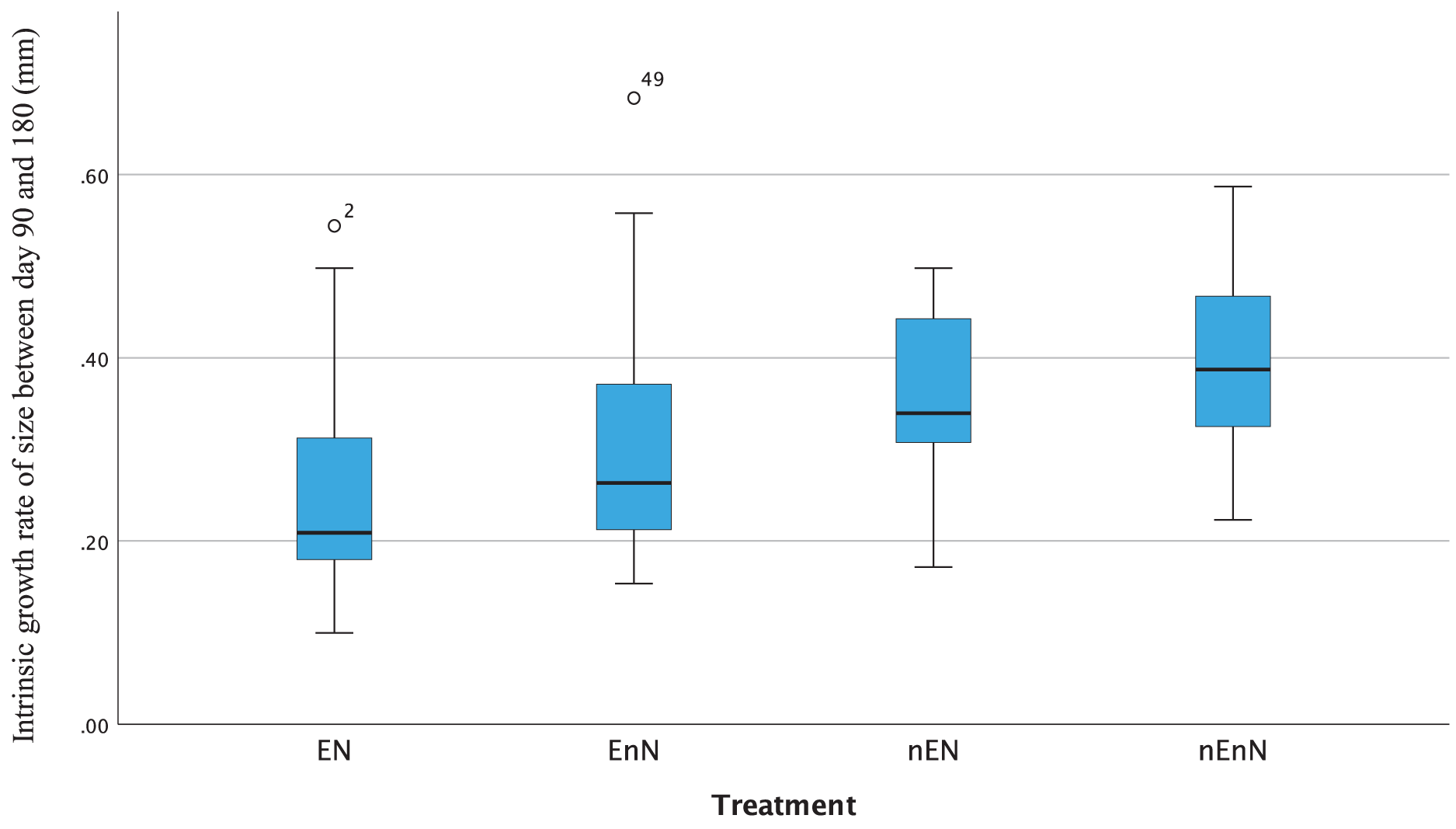{kind=link}
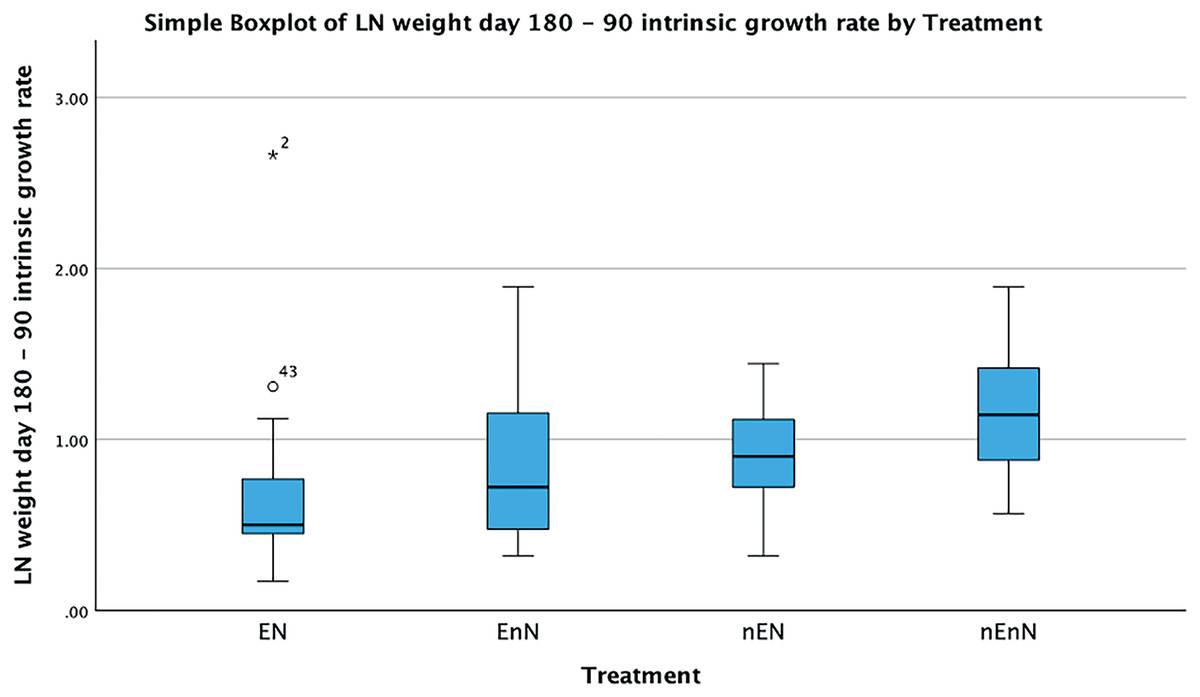
Figure 6: Significant differences in intrinsic growth rates for early development (day 90–180) based on mass by treatment group.
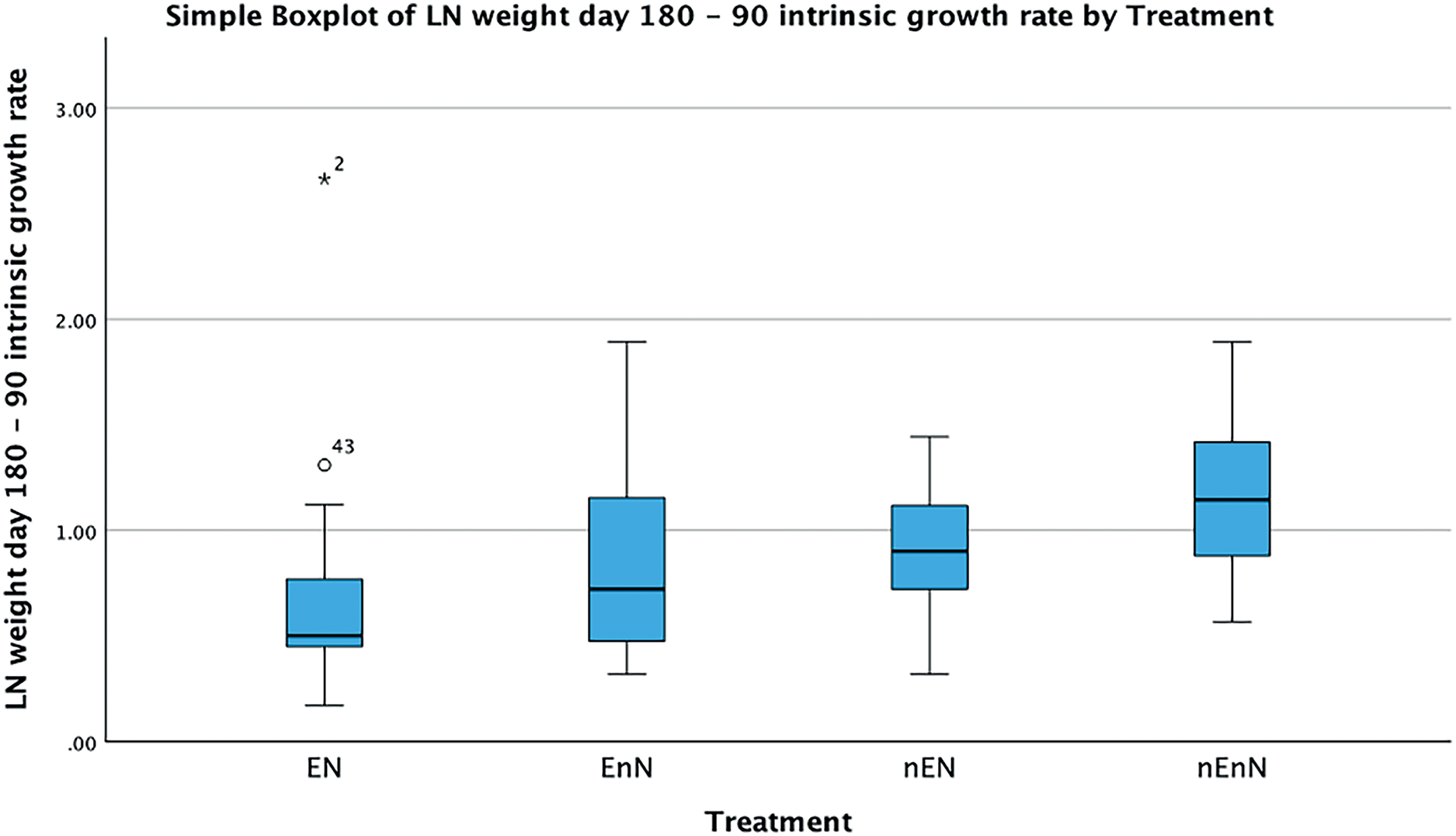{kind=link}
Reproduction
Fish paired with P. mexicana males resulted in nine fish producing at least one brood. Surprisingly, none of the P. latipinna males produced any offspring. Six of the nine fish that did reproduce were from a treatment group with physical enrichment (Table 1). Brood sizes and interbrood intervals were not analyzed due to the small sample size. Postmortem examination of all fish did not identify embryos, even those who had previously reproduced. Fish that gave birth during the project (n = 9) did not differ in final weight (unpaired t-test: t = 1.38, p = 0.072) or size (unpaired t-test: t = 1.27, p = 0.208) from fish that did not give birth (n = 31).
Social choice
Uniformly, the fish from all treatment groups preferred two fish as shoaling partners over one (paired t-test, n = 67, t = −4.6, p < 0.001; the mean time with one fish was 172 s, the mean time with two fish was 301 s) and there was no difference between the treatment groups (DF = 3, F = 0.830, p = 0.484). This is not surprising as fish usually choose shoal mates with similar physical characteristics and shoaling behavior offers protection by creating a uniform group that confuses predators making it harder to target individual fish.
Mortality
Thirteen fish deaths were recorded during the study (Table 2). One additional fish death was unrelated to study; four deaths were attributed to Ichthyophthirius multifiliis (a common fish disease) (Iaria et al., 2019). Fish that died during the study were removed from final analysis. Statistical analysis of mortality data using the Chi2 test with Yates correction (Chi2 = 0.52, n.s.) found no statistical significance. It is noteworthy, however, that nine fish deaths occurred in treatment groups nEN and nEnN. Furthermore, we identified a temporal pattern in the observed mortality. Eleven cases of mortality occurred before day 180.
Qualitative behavioral observations during the experiment
Although behavioral activity was not part of our study design, fish in the treatment group without physical or social enrichment were subjectively described as flighty, hypervigilant, and difficult to handle during weight and length measurements.
Discussion
We studied how environmental enrichment affected physical growth, social behavior, and reproductive ability in a small cohort of the clonal Amazon mollies. To the best of our knowledge, this is the first report of environmental effects on these physiologic and social characteristics in a clonal fish species. By documenting the effects of enrichment on growth, reproduction, and social preference this study provides useful insights into how environmental conditions can influence the overall well-being and fitness of Amazon mollies. Analyzing results across five domains of animal welfare: nutrition, physical environment, health, behavioral interaction, and mental state, we attempted to correlate the effects of environmental factors on specific physical parameters.
Access to nutrition was consistent across all study groups, but their physical environment varied. For the treatment groups nEN and nEnN, this difference hindered their ability to hide. The suppression of normal activities may have increased anxiety, fear, boredom, and illness, contributing to abnormal behaviors and higher early mortality in these groups. Fish without physical enrichment (nEN) and no enrichment (nEnN) exhibited noticeable physical and behavioral differences, especially during the first 120 days of life. Although our study did not formally include observations or analysis of fish behavioral activity as part of its design, we noticed that fish in the treatment group that lacked physical and social enrichment seemed to act in a way that could be described as nervous, overly alert, and challenging to manage when they were being measured for weight and length, and while subjective, these behavioral characteristics are associated with a distressed mental state.
Water quality, water changes, and water temperature were identical for all treatment groups and the development of lchthyophthirius multifiliis suggests individuals in treatment groups nEN and nEnN may have been immunocompromised. The relationship between stress response, growth performance, reproduction, and disease resistance in fish aligns with current understanding in the field (Huntingford et al., 2006; Sloman et al., 2019; Zhang, Gao & Zhang, 2022) and chronic or recurrent stress highlights the complex nature of stress responses in fish that can have long term effects on fish performance, including growth, and disease resistance (Barreto et al., 2022). Enrichment, both physical and social, has been shown to have positive effects, including improved immunocompetence (Barnard, Behnke & Sewell, 1996).
Mortality data for the four treatments found no statistically significant difference. Ten of the cases occurred in the treatments without enrichment, and only one case in the treatment with both physical and social enrichment (EN). This data indicates that a lack of enrichment may have been associated with increased mortality before day 180.
Physical enrichment refers to any form of physical complexity added to animal housing (Jones, Webster & Salvanes, 2021) and social enrichment includes housing with same or similar fish species, the ability to school, or for Amazon mollies the ability to shoal. We recognize providing visualization of a neighbor fish did not fully provide the natural socialization of Amazon mollies who typically form socially interactive shoals (Schlupp, 2009). However, multiple studies have established that normal social behavior can be triggered by visual stimuli only (Schlupp, 2009). In our social choice test, all fish displayed the inclination toward their natural shoaling behavior by preferring association with two fish. Overall, very few test fish gave birth. This was an unexpected finding, and it is unclear if the adverse welfare early in life had a long-acting adverse impact upon fecundity. Nonetheless, an interesting pattern emerged, with only mating between males of an allopatric population with Amazon mollies, P. mexicana from the Rio Oxolatan in Tabasco, Mexico producing live offspring. For logistical reasons, these offspring and their development were not followed. The observation that some fish had a second or even third brood is consistent with known sperm storage in the family Poeciliidae. Due to the timing of the experiment most of the fish reproduced in the fall and winter when reproduction may have been suppressed due to shortening day length. Indeed, reduced reproduction attributed to seasonality is known for Amazon mollies (Fischer & Schlupp, 2009; Heubel & Schlupp, 2008). This may have been a peculiarity of our study, which took place in a greenhouse with controlled temperature but under natural light conditions. Consequently, this may be less of a concern for studies conducted indoors with controlled lighting conditions.
Two traits were measured that reflect individual growth, size as standard length (mm) and mass (g). Although these traits are correlated, the two variables were analyzed separately to facilitate detection of factors that operate on one measurement, but not the other. Near day 270, growth plateaued in all fish, reflecting the expected asymptotic growth. The box plots (Figs. 4, 5) of the increase of size and weight showed the expected shape of a diminishing return curve. At the end of the project, individuals in treatment groups without enrichment began to decrease in mass without concurrent loss in standard length. This may be interpreted as evidence of the onset of senescence and appeared to begin between day 270 and 300. Senescence has been studied extensively in natural populations of guppies (Poecilia reticulata) (Reznick, Ghalambor & Nunney, 2002; Bryant & Reznick, 2004; Reznick, Bryant & Holmes, 2006). Intrinsic growth rates show a similar pattern with significant differences in the early phase of the project for both size and mass, but no difference in the later phase of the project. We believe this is an important finding, as it shows the role of resilience in the individual development of these fish. They can compensate for poor welfare adversity early in life, although the mechanism remains unknown and may be the basis for future research. From a welfare perspective this may indicate masking of early effects later in life, but from an experimental viewpoint this indicates how important it may be to select the proper stage of life history having known welfare considerations to obtain meaningful data. It is also quite interesting to find strong resilience in a clonal fish with very limited genetic variability. Our data further shows considerable variability between groups early but also in general.
Conclusions
Our results support a broad understanding of how different environmental factors may influence the welfare and overall health of Amazon mollies. This approach is critical for both scientific research and the development of effective enrichment strategies for fish husbandry and management. We provided a genuine focus upon welfare considerations during experimental design, and such application is reflective of the definition of good welfare—allowing animals to experience good welfare by having constructs, physical environments, and social enrichments of visible neighbors to facilitate their expression of good welfare indices. Overall, this data shows a clear benefit of focal attentiveness to welfare considerations—especially with physical and social enrichment early in life for Amazon mollies. Individuals with enrichment, particularly social enrichment of a neighbor, were larger and heavier early in life. However, these differences disappeared later in life and fish from the disadvantaged enrichment treatment groups caught up to the point where all fish ended the project at day 300 with almost equal mass and size. We detected potential signs of senescence in the two treatments without a neighbor. These groups faced an earlier onset of senescence, but this requires additional studies. Based on these findings, we strongly recommend social and physical enrichment for live-bearing fish, especially early in life.
There is also evidence of increased survivability of fish with social and physical enrichment compared to conspecifics without social and physical enrichment, establishing the criticality of attentiveness to welfare concepts and a clear intention of including welfare principles during experimental design. The core reason is that animals which experience good welfare provide the most meaningful, replicative, and reliable study results. While determining the preferred environment and tools which allow this species to experience good welfare is at a minimum challenging, this study notes that what has historically been considered as ‘environmental enrichment’ is much more than just environmental enrichment but is in fact the basis of setting the stage for facilitating good welfare in research aquatic species—in this case mollies.