
Up-regulated IL-17 and Tnf signaling in bone marrow cells of young male osteogenesis imperfecta mice
- Published
- Accepted
- Received
- Academic Editor
- Eva Mezey
- Subject Areas
- Biochemistry, Bioinformatics, Cell Biology, Molecular Biology
- Keywords
- Osteogenesis imperfecta, Bone resorption, RNA sequencing, Bone marrow, Inflammation
- Copyright
- © 2022 Shao et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2022. Up-regulated IL-17 and Tnf signaling in bone marrow cells of young male osteogenesis imperfecta mice. PeerJ 10:e13963 https://doi.org/10.7717/peerj.13963
Abstract
Osteogenesis imperfecta (OI) is a congenital bone dysplasia mainly caused by either defective production or assembly of type I collagen. The skeletal phenotypes especially fractures are often seen in OI adolescents. Studies have found that an increased number of osteoclasts and excessive bone resorption existed in collagen-related OI, which has not been well understood. Emerging evidence has suggested that inflammation may be associated with OI. We speculated that the bone marrow (BM) niche had similar inflammatory changes and performed RNA-sequencing (RNA-seq) in BM cells derived from young male mice to analyze the related differentially expressed genes (DEGs) and pathways. Data showed that there were 117 shared DEGs (Q ≤ 0.05, |log2FC| ≥ 1) in BM cells isolated from two types of OI murine models that respectively simulate different OI types. Gene Ontology (GO) (Q ≤ 0.05) analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) (Q ≤ 0.05) analysis and real-time PCR validation indicated the dysregulated biology process of cellular response to interferon (Ifn) together with upregulated IL-17 signaling, tumor necrosis factor (Tnf) signaling and osteoclast differentiation in OI BM niche. Either defective collagen production or abnormal collagen assembly shared similar alterations in gene profiles and pathways involving inflammation and osteoclast activation. Data presented here not only contributed to understanding of the mechanism of the enhanced bone absorption in the bones of OI, but also provided more evidence to develop potential anti-inflammation therapies.
Highlight
There were 117 shared differentially expressed genes in bone marrow (BM) cells isolated from two types of OI young male murine models.
The upregulated IL-17 signaling, Tnf signaling and osteoclast differentiation were significantly enriched in OI BM cells.
These dysregulated DEGs and pathways in BM cells might be associated with the excessive bone resorption of the OI mice.
Introduction
Osteogenesis imperfecta (OI) is a congenital disorder characterized by bone fragility (Forlino et al., 2011). The patients generally suffer from recurrent bone fractures and deformities. Up to 21 genes have been associated with OI, but most of the total cases are caused by heterozygous mutation in either of the genes coding for the type I collagen alpha chains, COL1A1 or COL1A2 (Forlino et al., 2011). The pathogenic variants of the two genes often cause either insufficient synthesis or abnormal structure of skeletal collagen, which is correlated with mild OI type I and more severe type II–IV respectively (Saito & Marumo, 2015; Nijhuis et al., 2019).
Multiple reports have highlighted the impaired bone formation and excessive bone absorption in defective collagen-related OI (Takeyari et al., 2021; Iwamoto, Takeda & Ichimura, 2002), which are the main targets in OI treatments. Type I collagen fibrils are produced by bone-forming osteoblasts, making the dysfunctional osteoblasts and their weakened mineralization the main focus of OI etiological research. Conversely, the mechanism of enhanced bone destruction in OI bones has not been well understood, although anti-resorptive drugs have been widely applied in clinical interventions.
Some recent studies have indicated the contribution of inflammation and inflammatory factors in OI bone phenotype. Increased transformation growth factor-beta (TGF β) and excessive TGF signaling have been regarded as a promising target to enhance bone formation in some OI models and patients (Grafe et al., 2014; Marom, Rabenhorst & Morello, 2020). Also, the elevated serum level of interferon (IFN) and tumor necrosis factor (TNFα) have been found in patients and mouse models separately, suggesting the chronic inflammation state in OI (Zhytnik et al., 2020). Inflammation has been closely linked to bone biochemical changes and bone loss in rheumatoid arthritis (RA) and osteoporosis (Weyand & Goronzy, 2021; Tilg et al., 2008). The progenitor cells of osteoblasts and osteoclasts derived from hematopoietic stem cell (HSC) lineage are both situated in the same bone marrow (BM) niche (Ono & Nakashima, 2018). The inflammatory alterations of the BM niche that have not been well explored may directly or indirectly act on the bone manifestations of OI.
In the process of bone marrow development, the transformation of red bone marrow to yellow bone marrow is most obvious in infancy and childhood (Berg, 2021). Compared with the old bone marrow, the level of ROS in the young bone marrow is lower, and the cells are less affected by aging, which can better reflect the initial bone marrow state under genetic defects (Yao et al., 2021). Also, the skeletal symptoms of OI are most obvious in childhood and can be relieved in adulthood (Paterson, McAllion & Stellman, 1984). Studying the bone marrow of young mice will help to understand the cause and process of OI. Here, we performed RNA-sequencing (RNA-seq) and differential gene expression analyses in femoral BM cells isolated from young mice of two types of OI mouse models and wild-type mice to explore the changes in the BM niche. Both the Col1a1+/−365 mice and heterozygous oim mice (Col1a2oim/+) display osteogenesis deficiency accompanied by an increased number of osteoclasts. The differentially expressed genes (DEGs) shared by the two OI models were involved in IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation. The DEGs related to the three pathways were all upregulated in OI BM cells when compared with normal cells, suggesting the activation of these signalings. These dysregulated genes and pathways in mutant BM cells are likely to play an important role in the pathological changes of OI bones.
Materials and Methods
OI mice model
Adult wild-type (wt) C57BL/6 mice were purchased from the Laboratory Animal Center of the Academy of Military Medical Science (China). The B6C3Fe a/a-Col1a2oim/J mice (#001815) were bought from the Jackson Laboratory (Bar Harbor, ME, USA) and maintained on a congenic C57BL/6 background. The Col1a2oim/+ mice were heterozygotes carrying a single mutant Col1a2 allele and performed a mild form of OI. The Col1a1+/−365 mice with a Col1a1 gene knock-down can simulate OI type I. Heterozygous mice were bred by crossing heterozygous individuals and genotyped as previously described (Chipman et al., 1993; Liu et al., 2019). All mice were housed in specific pathogen-free conditions and sacrificed by CO2 in 4 weeks old to sample. Mice were fed with full-price diet and autoclaved water. The number of mice per cage did not exceed five. All experiments were performed following the approval of Animal Care and Use Committee of Tianjin Medical University (TMUaMEC 2017012). Only male mice were used in the present study.
Cell isolation
The femurs of 4-weeks and 12-weeks old male mice were harvested and the bone marrow (BM) cells were flushed out with a syringe. The wt and OI modeled mice-derived BM cells were marked as BMwt, BMoim/+ and BM+/−365 respectively.
RNA extraction and RNA sequencing (RNA-seq)
Total RNA of freshly isolated BM cells was extracted using Trizol reagents (Invitrogen, Waltham, MA, USA). Each type of cells isolated from one mouse was seen as one sample and each group contained three samples. RNA samples (n = 3/genotype from young animals) with RNA integrity number ≥ 7.0 and 28S/18S ratios ≥ 1.5 were sequenced. Another 18 RNA samples (n = 6/genotype) were used in quantitative PCR verification.
A total of 18 cDNA libraries were constructed and separately sequenced by Beijing Genomics Institute (BGI, Beijing, China) using the BGISEQ-500 platform. RNASeqPower Software were used to calculated the statistical power of this experimental design. Sequence data (~75 million reads) were checked for sequencing quantity by FASTQC. Reads with low quality (unknown nucleotides > 10%, or Q20 < 20%), adapter contamination and high N content of unknown bases (N > 5%) were excluded to gain clean reads. The clean reads were then mapped to the mouse reference genome (Mus_musculus, NCBI, GCF_000001635.26_GRCm38.p6) and analyzed by HISAT2 software. Read counts were normalized to TPM (transcripts per kilobase of exon model per million mapped reads). Q-value was obtained by false discovery rate (FDR) correction of the P-value. Differentially expressed genes (DEGs) (Q ≤ 0.05, |log2FC| ≥ 1) were analyzed by DEseq2 software. Gene Ontology (GO) (Q ≤ 0.05) and Kyoto Encyclopedia of Genes and Genomes (KEGG) (Q ≤ 0.05) analyses were performed in DEGs shared by the two OI models derived BM cells using BGI Dr. Tom multi-omics data mining system. Gene Set Enrichment Analysis (GSEA) based on the KEGG database was also performed using BGI Dr. Tom and results with P ≤ 0.05 and Q ≤ 0.25 were considered statistically significant.
Quantitative real-time PCR (RT-PCR)
The RNA samples from both young and adult mice were reverse transcribed to cDNA using GoScript reverse transcriptase (Promega, Madison, WI, USA) according to the manufacturer’s protocol. RT-PCR analysis was performed using the AceQ RT-PCR SYBR Green Master Mix kit (Vazyme, Nanjing, China). The cycling program referenced our previous report (Liu et al., 2020). All samples were evaluated in triplicate and normalized to mouse glyceraldehyde-3-phosphate dehydrogenase (Gapdh). All the primers were synthesized from Sangon Biotech Co., Ltd. (China), and the sequences were listed in Table 1.
| Gene | Forward (5′-3′) | Reverse (5′-3′) | Product length (bp) |
|---|---|---|---|
| Lif | TCAACTGGCACAGCTCAATGGC | GGAAGTCTGTCATGTTAGGCGC | 119 |
| Jun | CCTTCTACGACGATGCCCTC | GGTTCAAGGTCATGCTCTGTTT | 102 |
| Fosb | TTTTCCCGGAGACTACGACTC | GTGATTGCGGTGACCGTTG | 174 |
| Stat1 | TCACAGTGGTTCGAGCTTCAG | GCAAACGAGACATCATAGGCA | 155 |
| Cxcl10 | CCAAGTGCTGCCGTCATTTTC | GGCTCGCAGGGATGATTTCAA | 157 |
| Ccl2 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT | 121 |
| Ifng | ATGAACGCTACACACTGCATC | CCATCCTTTTGCCAGTTCCTC | 182 |
| Ifit1 | GCCTATCGCCAAGATTTAGATGA | TTCTGGATTTAACCGGACAGC | 75 |
| Irf7 | GAGACTGGCTATTGGGGGAG | GACCGAAATGCTTCCAGGG | 102 |
| Gapdh | CATCACTGCCACCCAGAAGACTG | ATGCCAGTGAGCTTCCCGTTCAG | 153 |
Note:
Lif, leukemia inhibitory factor; Jun, jun proto-oncogene; Fosb, FBJ osteosarcoma oncogene B; Stat1, signal transducer and activator of transcription 1; Cxcl10, chemokine (C-X-C motif) ligand 10; Ccl2, chemokine (C-C motif) ligand 2; Ifng, interferon gamma; Ifit1, interferon-induced protein with tetratricopeptide repeats 1; Irf7, interferon regulatory factor 7; Gapdh, glyceraldehyde-3-phosphate dehydrogenase.
Statistical analysis
Statistical analysis was conducted using SPSS version 17.0 software (IBM SPSS Statistics, Chicago, IL, USA). All data were presented as mean ± standard deviation (SD). Two groups were generally compared by unpaired. The D’Agostino-Pearson omnibus normality test was used to test the normal distribution of data. If the data conformed to the normal distribution (P > 0.1), the unpaired t-test was used. In the unpaired t-test, the F-test was used to analyze the homogeneity of variance for pairwise comparison. If the variance was homogenous (P > 0.05), the unpaired t-test was performed. If variances were not homogeneous (P < 0.05), Welch’s correction was used to correct them. If the data were not subject to a normal distribution, the Mann-Whitney test was used. P < 0.05 was considered statistically significant.
Results
Differential gene expression analysis
RNA-seq was performed in BM cells of the three genotypes of mice. The statistical power (depth = 100, cv = 0.1, effect = 2, α = 0.05, power = 0.9) of this experimental design, calculated in RNASeqPower was 0.0875. Compared with BMwt, a total of 279 DEGs were identified in BM+/−365 and 219 of which were upregulated and 60 were downregulated (Fig. 1). A total of 291 DEGs were differentially expressed in BMoim/+ when compared with BMwt, 237 of which were upregulated and 54 were downregulated (Fig. 1). There were 117 DEGs shared by the two types of OI mice (Fig. 1). A total of 102 of the common DEGs were concordantly upregulated and 12 were concordantly downregulated, suggesting their consistency of alteration (Data not shown).
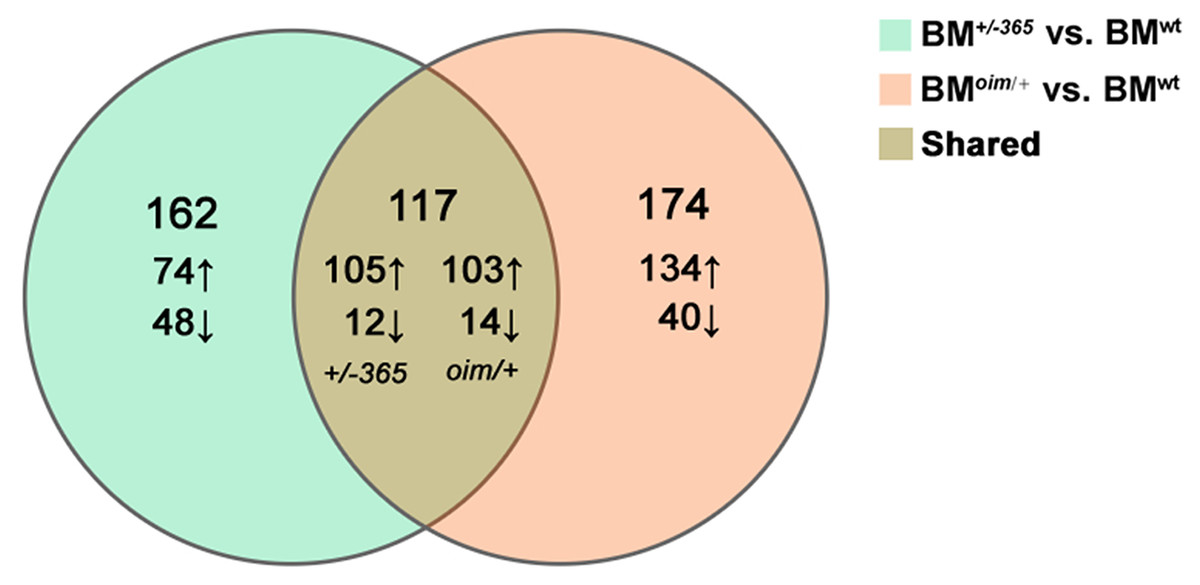
Figure 1: The results of RNA-seq of bone marrow (BM) cells.
The Venn diagram of differentially expressed genes (DEGs) in BM cells isolated from 4-weeks old wt mice, heterozygous Col1a1+/−365 and Col1a2oim/+ mice (Q ≤ 0.05, |log2FC| ≥ 1). There were 117 DEGs shared by the two types of OI mice.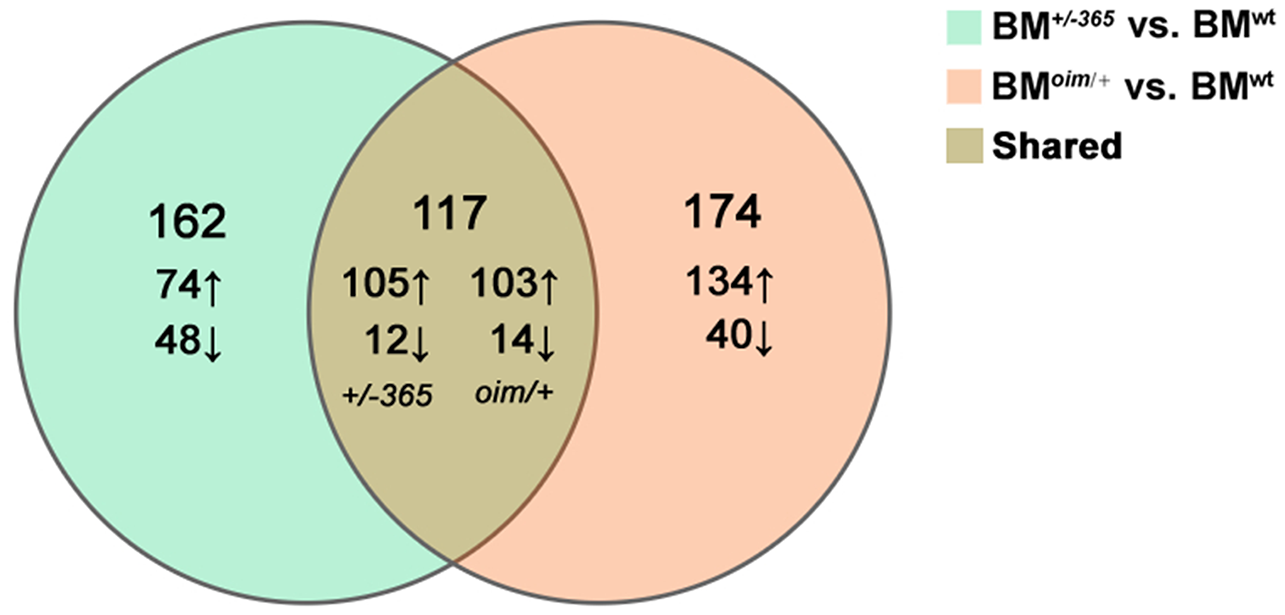{kind=link}
The top 20 upregulated shared DEGs in BM+/−365 or BMoim/+ were listed in Tables 2 and 3 respectively. A total of 17 of them were the same genes that contained several Ifn signaling-related genes (e.g., Lfit1, Lfit3, Lfi44 and Irf7) (Tables 2 and 3). Notably, the transcription of Coch, most abundantly expressed in the inner ear, was obviously increased in the two types of BM cells (Tables 2 and 3). The abnormal Coch expression might be associated with OI hearing loss.
| Gene | Description | Log2 (Fold chang) | Q-value |
|---|---|---|---|
| Fgf23 | fibroblast growth factor 23 | 5.83 | 0.0002360794650486 |
| Ptgds | prostaglandin D2 synthase (brain) | 5.54 | 0.0021177353272204 |
| Oas1g | 2’–5’ oligoadenylate synthetase 1G | 4.9 | 0.0009431000835065 |
| Apol9b | apolipoprotein L 9b | 4.6 | 0.0002133173066225 |
| Ifit1 | interferon-induced protein with tetratricopeptide repeats 1 | 4.28 | 0.0003668837536278 |
| Ifit3 | interferon-induced protein with tetratricopeptide repeats 1 | 4.25 | 0.0015924514011703 |
| Ifi44 | interferon-induced protein 44 | 4.17 | 0.003021888072243 |
| Coch | coagulation factor C homology/cochlin | 4.11 | 0.0260410468387774 |
| Isg15 | ISG15 ubiquitin-like modifier | 3.98 | 0.0000106275044397477 |
| Ifit3b | interferon-induced protein with tetratricopeptide repeats 3b | 3.91 | 0.001908267305679 |
| Rsad2 | radical S-adenosyl methionine domain containing 2 | 3.38 | 1.62699675914253e−13 |
| Usp18 | ubiquitin specific peptidase 18 | 3.24 | 0.0010491995647924 |
| Oas1a | 2’–5’ oligoadenylate synthetase 1A | 3.05 | 0.0016396718614034 |
| Oasl2 | 2’–5’ oligoadenylate synthetase 2 | 3 | 0.0031321485213441 |
| Gbp6 | guanylate binding protein 6 | 3 | 0.0010429030626467 |
| Irf7 | interferon regulatory factor 7 | 2.98 | 0.002419157920696 |
| Cfb | complement factor B | 2.97 | 0.0133696378926878 |
| Gbp10 | guanylate-binding protein 10 | 2.97 | 0.0013662770771023 |
| Zbp1 | Z-DNA binding protein 1 | 2.9 | 0.0088547659475085 |
| Rtp4 | receptor transporter protein 4 | 2.85 | 0.0013424962435546 |
| Gene | Description | Log2 (Fold chang) | Q-value |
|---|---|---|---|
| Ptgds | prostaglandin D2 synthase (brain) | 7.5 | 6.17204589560265e−9 |
| Coch | coagulation factor C homology/cochlin | 6.02 | 2.94335300566472e−8 |
| Oas1g | 2’–5’ oligoadenylate synthetase 1G | 5.18 | 9.681071247414061e−14 |
| Fgf23 | fibroblast growth factor 23 | 4.99 | 0.000606927029448 |
| Ifi44 | interferon-induced protein 44 | 4.46 | 1.23002041345624e−21 |
| Ifit3 | interferon-induced protein with tetratricopeptide repeats 3 | 4.44 | 1.08339825790861e−9 |
| Ifit1 | interferon-induced protein with tetratricopeptide repeats 1 | 4.22 | 5.155872680064089e−11 |
| Ifit3b | interferon-induced protein with tetratricopeptide repeats 3b | 4.2 | 9.503797111990189e−11 |
| Gbp6 | guanylate binding protein 6 | 3.63 | 1.1575697141787101e−74 |
| Apol9b | apolipoprotein L 9b | 3.52 | 0.0012727812836212 |
| Cfb | complement factor B | 3.47 | 9.1080396056311e−39 |
| Isg15 | ISG15 ubiquitin-like modifier | 3.37 | 7.51494012126258e−18 |
| Tgtp1 | T cell specific GTPase 1 | 3.36 | 1.28520549551154e−10 |
| Oasl2 | 2’–5’ oligoadenylate synthetase 2 | 3.19 | 1.83115647109809e−26 |
| Gbp10 | guanylate-binding protein 10 | 3.19 | 3.41807441735383e−13 |
| Oas1a | 2’–5’ oligoadenylate synthetase 1A | 3.15 | 3.8026707877775104e−15 |
| Usp18 | ubiquitin specific peptidase 18 | 3.14 | 1.29677254526075e−14 |
| Iigp1 | interferon inducible GTPase 1 | 3.14 | 0.000013763932948896 |
| Irf7 | interferon regulatory factor 7 | 3.12 | 6.5309598268511e−36 |
| Ly6i | lymphocyte antigen 6 complex, locus I | 3.1 | 3.91587391622785e−21 |
Gene Ontology biology process analysis
Gene Ontology (GE) biology process analysis indicated the enrichment of multiple immune-related genes in such as cellular response to interferon-beta (Ifnβ), immune system process, and immune response, and cellular response to interferon-alpha (Ifnα) in defective BM cells (Fig. 2A). We tested the level of Lfit1 and Irf7, and data showed that both of the two genes were significantly upregulated in mutant BM cells (Figs. 2B and 2C), suggesting the activated Ifn signaling in OI BM.
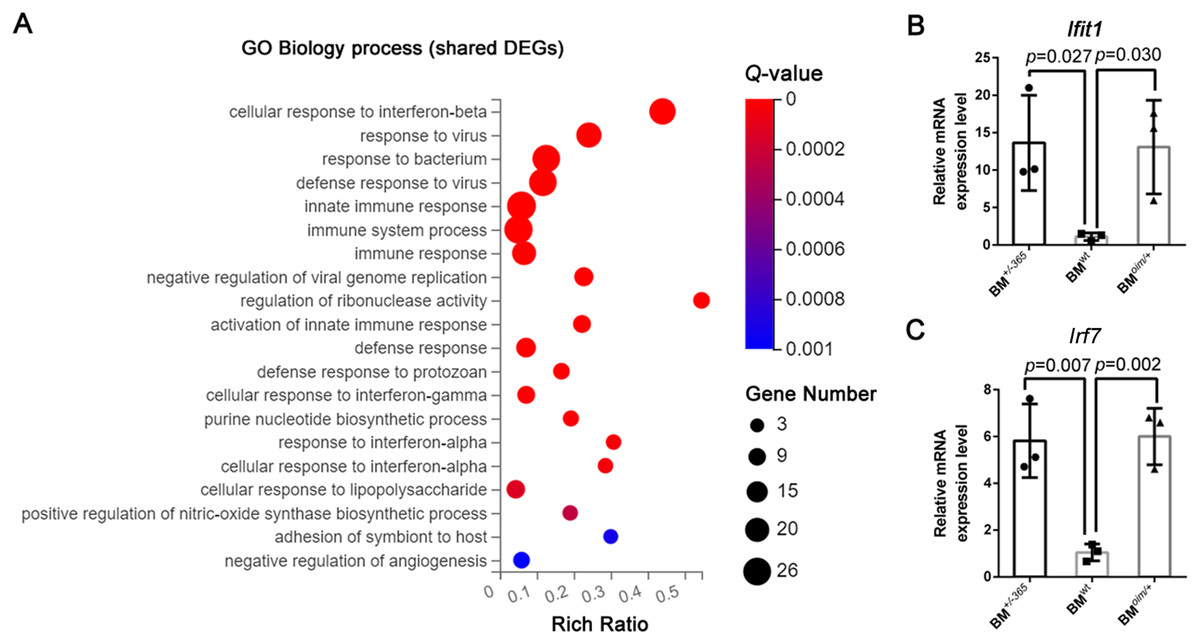
Figure 2: The results of GO Biology process analysis of the shared DEGs.
(A) Top 20 GO Biology process analysis enrichments of the shared 117 DEGs of BM+/−365 and BMoim/+ (Q ≤ 0.05). Multiple immune-related DEGs were enriched in cellular response to interferon. The size of the spot represents the number of differential genes, the color represents the Q value; (B, C) Real-time PCR identified the expression levels of Lfit1 (B) and Irf7 (C) in BM cells isolated from 4-weeks old wt, heterozygous Col1a1+/−365 and Col1a2oim/+ mice (n = 3, P < 0.05). The result indicated that both of the two genes were significantly up-regulated in BM+/−365 and BMoim/+. Data in the quantitative plots are presented as mean ± SD using upaired t-test.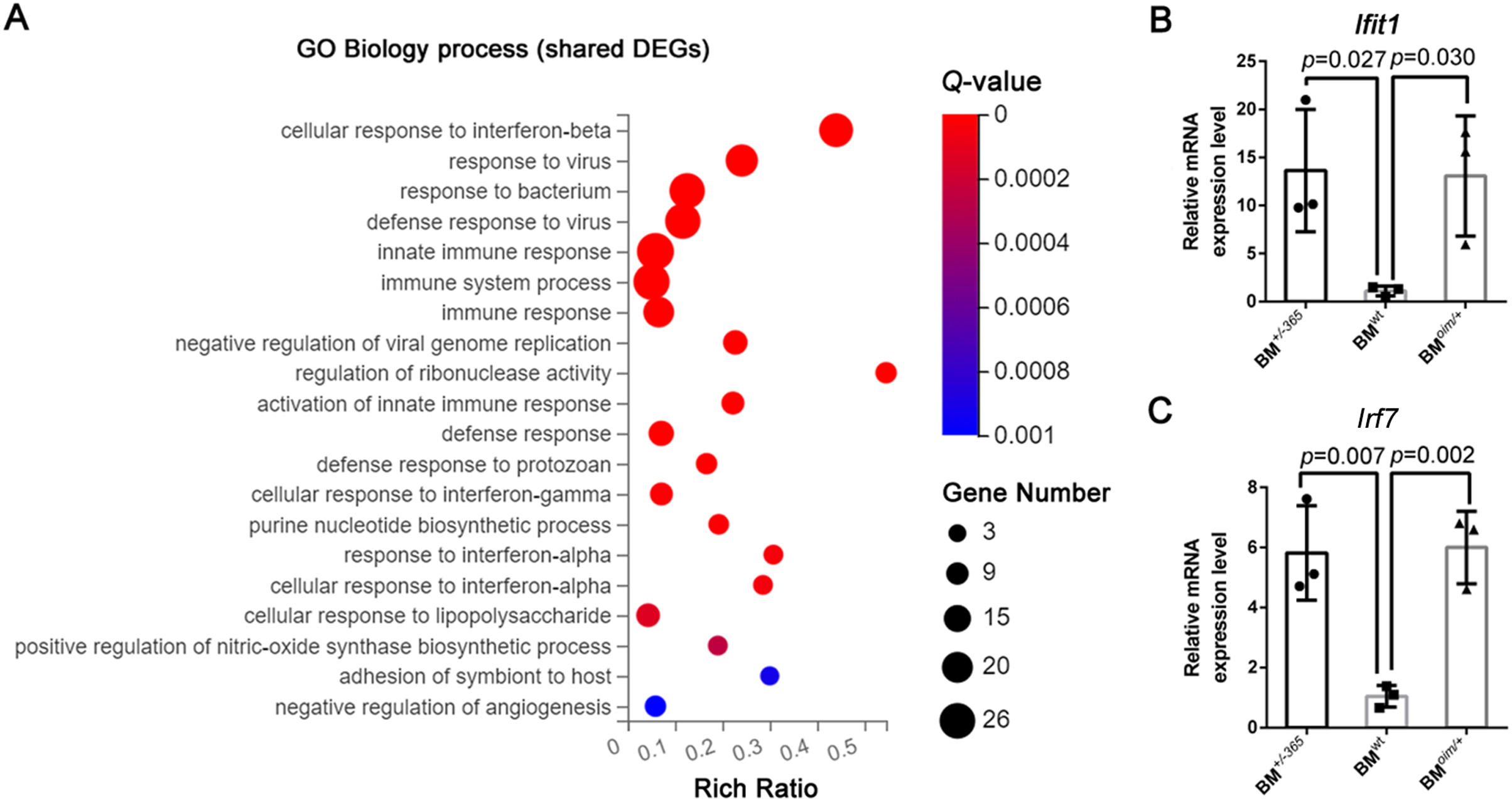{kind=link}
KEGG pathway enrichment and GSEA analysis
The significantly enriched pathways by KEGG analysis of the 117 shared DEGs included the IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation in OI BM cells (Fig. 3A). There were 11 enriched DEGs involved in the aforementioned pathways (Table 4). GSEA analysis demonstrated that all of the three pathways were markedly upregulated in OI-derived BM cells (Figs. 3B–3D). Besides, the sequencing data demonstrated that both I117a and Tnf increased more than two-fold in BMoim/+ (Table 5). The Tnf expression also got significantly elevated in Col1a1+/365 mice-derived BM cells (Table 5). In addition, the mRNA level of Rankl (Tnfsf11) rose in defective BM cells (Table 5). It has been reported that upregulated IL-17 and Tnf signalings contribute to the excessive bone resorption in multiple inflammatory bone loss diseases (Tsukasaki & Takayanagi, 2019; Weitzmann, 2017), suggesting the potential role of overactivated IL-17 and Tnf pathways in an increased number of osteoclasts under the OI background.
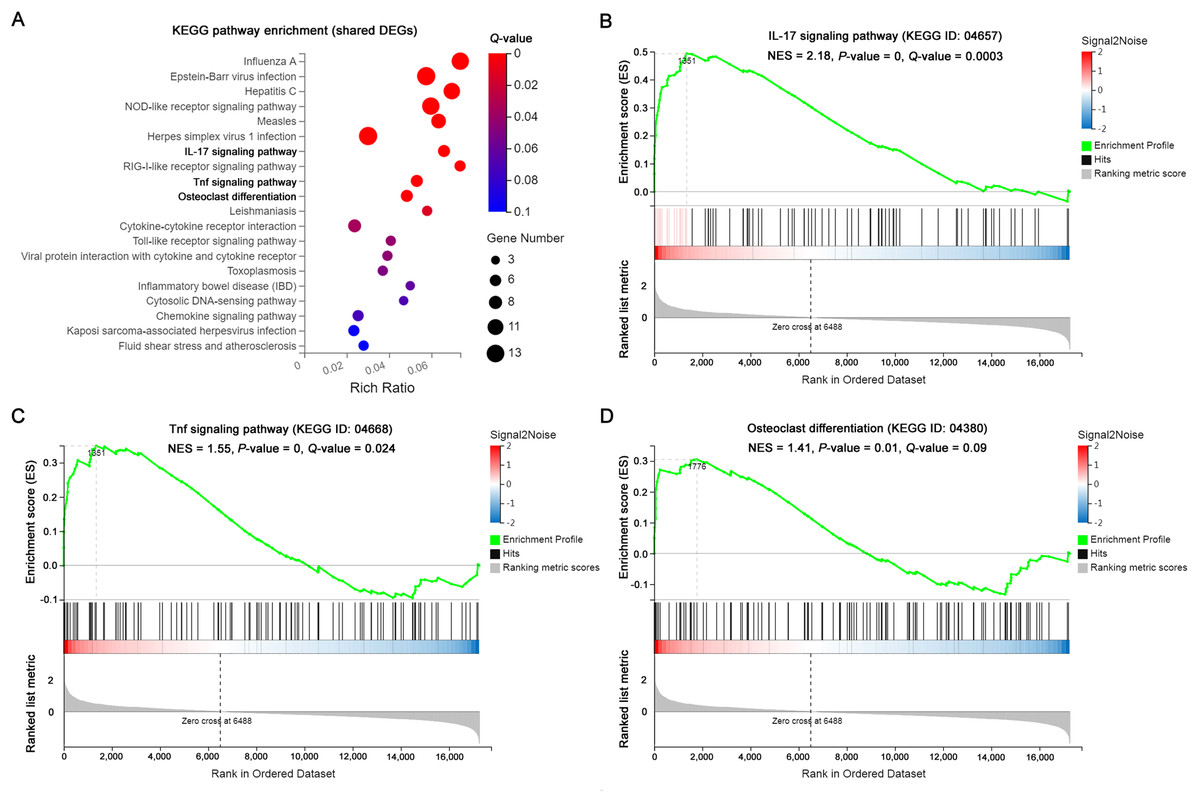
Figure 3: The results of KEGG pathway enrichment and GSEA analysis of the shared DEGs.
(A) Top 20 KEGG analysis enrichments of the shared 117 DEGs of BM+/−365 and BMoim/+. The significantly enriched pathways of the shared DEGs included the IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation in OI BM cells (Q ≤ 0.05). The size of the spot represents the number of differential genes, the color represents the Q value. (B–D) The GSEA analysis of the aforementioned three pathways (P ≤ 0.05 and Q ≤ 0.25). The results demonstrated that all of the three pathways were markedly upregulated in OI-derived BM cells.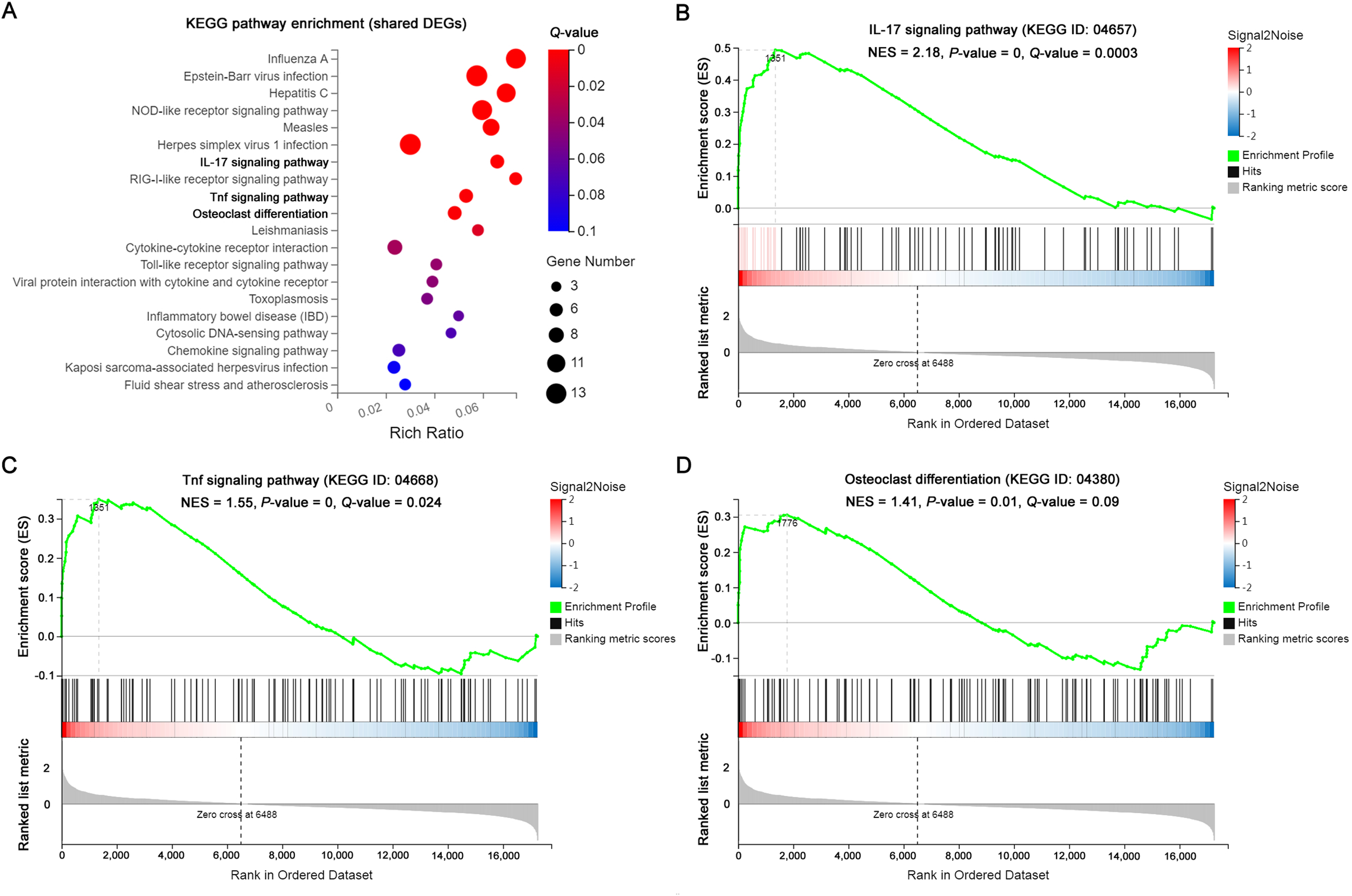{kind=link}
| Gene | Description | Log2 (Fold chang) | Q-value | ||
|---|---|---|---|---|---|
| BM+/−365 | BMoim/+ | BM+/−365 | BMoim/+ | ||
| Ccl2 | chemokine (C-C motif) ligand 2 | 2.73 | 2.92 | 5.23025917899991e−16 | 0.00000124536325640583 |
| Gm5431 | predicted gene, 45935 | 1.11 | 1.47 | 0.0421644826331762 | 4.85491962362027e−9 |
| Cxcl10 | chemokine (C-X-C motif) ligand 10 | 2.09 | 1.57 | 0.0009539010036805 | 0.0028754859114639 |
| Lif | leukemia inhibitory factor | 1.46 | 1.70 | 0.0098724762681209 | 0.0024271897572989 |
| Ifng | interferon gamma | 1.71 | 2.15 | 0.0002510762900628 | 2.01552889198521e−7 |
| Fosb | FBJ osteosarcoma oncogene B | 2.40 | 2.23 | 0.00000208273094362867 | 0.0008131293868126 |
| Stat1 | signal transducer and activator of transcription 1 | 1.27 | 1.47 | 0.0000710335367185181 | 4.59990999943035e−23 |
| Jund | jun D proto-oncogene | 1.00 | 1.04 | 0.0015850803802578 | 2.41298702546188e−14 |
| Jun | jun proto-oncogene | 1.86 | 1.66 | 4.2489456147999705e−20 | 1.75119110664004e−52 |
| Lfi47 | interferon-induced protein 47 | 1.62 | 1.75 | 0.0247317221652343 | 5.43681272346479e−24 |
| Fcgr1 | Fc receptor, IgG, high affinity I | 2.13 | 2.63 | 0.0033203206673691 | 1.34407141389434e−33 |
| Gene | Description | Log2 (Fold Chang) | Q-value | ||
|---|---|---|---|---|---|
| BM+/−365 | BMoim/+ | BM+/−365 | BMoim/+ | ||
| Tnf | tumor necrosis factor | 0.77 | 1.21 | 0.0110345421756999 | 2.8259877145669397e−13 |
| Il17a | interleukin 17A | 2.98 | 4.62 | 0.529929360546415 | 0.0204958961911906 |
| Tnfrsf11a | tumor necrosis factor receptor superfamily, member 11a, NFKB activator/Rank | −0.004 | 0.20 | 0.996564355301463 | 0.791864645628747 |
| Tnfsf11 | tumor necrosis factor (ligand) superfamily, member 11/Rankl | 1.09 | 0.95 | 0.587237549231164 | 0.486382634245118 |
The changes of DEGs in BM cells isolated from OI modeled mice
The heatmap as shown in Fig. 4A displayed the differential expression of the 11 DEGs mentioned above. Many of them at least participate in two of the IL-17 signaling pathway, Tnf signaling pathway, and osteoclast differentiation, suggesting their complex crosstalk. RT-PCR validation indicated that Ccl2 (Fig. 4B), Jun (Fig. 4C) Cxcl10 (Fig. 4D), Lif (Fig. 4E) and Stat1 (Fig. 4F) were apparently upregulated in mutant BM cells, which was consistent with the sequencing data. And Fosb (Fig. 4G) and Ifng (Fig. 4H) expression also showed an increasing trend in OI mice-derived BM cells. These results preliminarily identified the dysregulated DEGs, which might play important roles in OI bone erosion alterations.
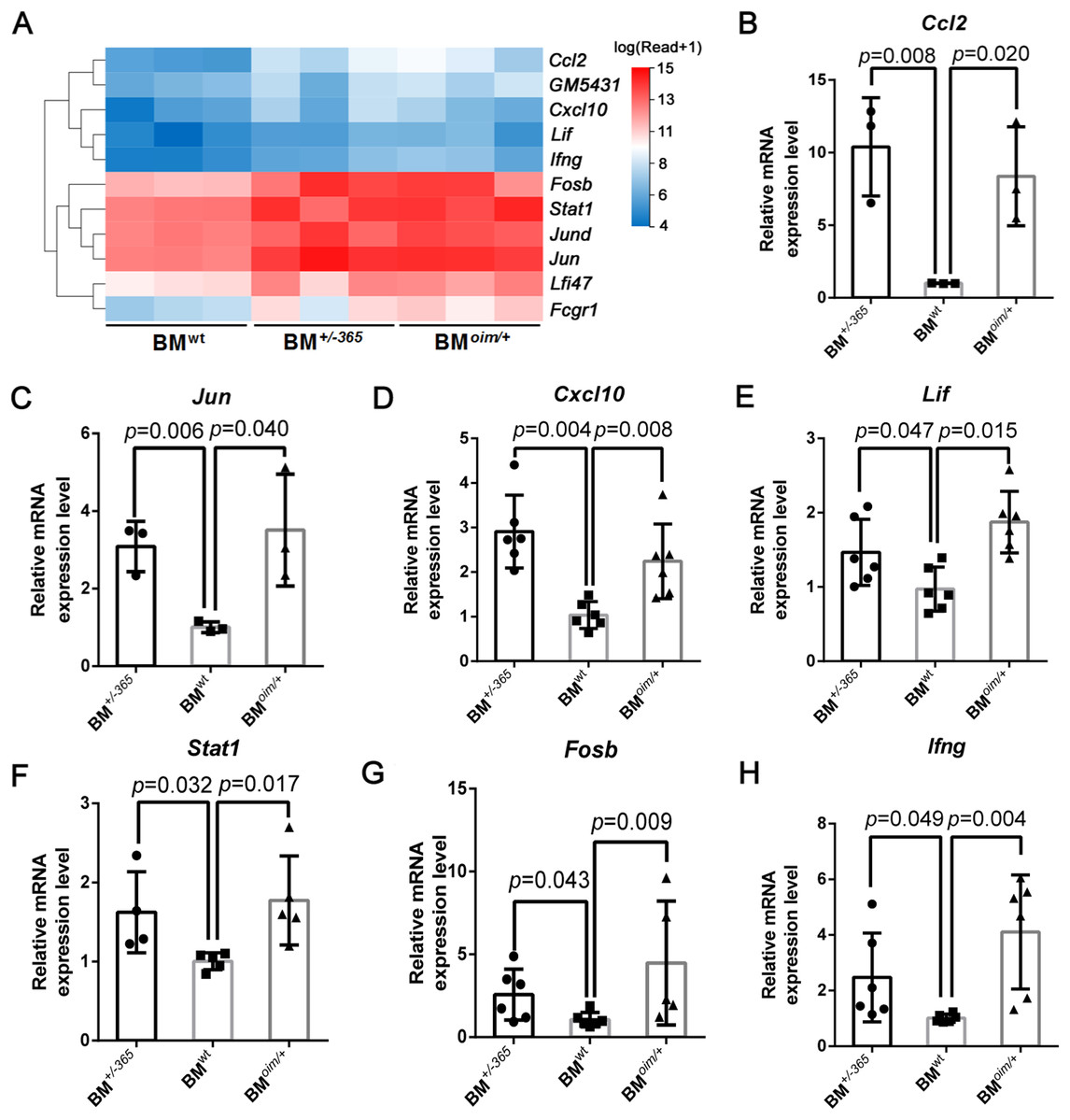
Figure 4: The expression of the shared DEGs involving IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation in BM+/−365 and BMoim/+ when compared with BMwt.
(A) The heatmap of the 11 DEGs involved in IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiationof 4-weeks BM cells. (B–G) Real-time PCR tested the expression levels of some DEGs. The result indicated that Ccl2 (B) and Jun (C) were apparently upregulated in mutant BM cells. The obviously increased level of Cxcl10 (D) in BM+/−365 together with Lif (E) and Stat1 (F) in BMoim/+ could be observed. Fosb (G) and Ifng (H) also showed an significantly increased in OI BM cells (n = 3 for B, C, n = 6 for D, E, G, H, n = 4 for F, P < 0.05). Data in the quantitative plots are presented as mean ± SD using upaired t-test (B–F, H) and Mann-Whitney test (G).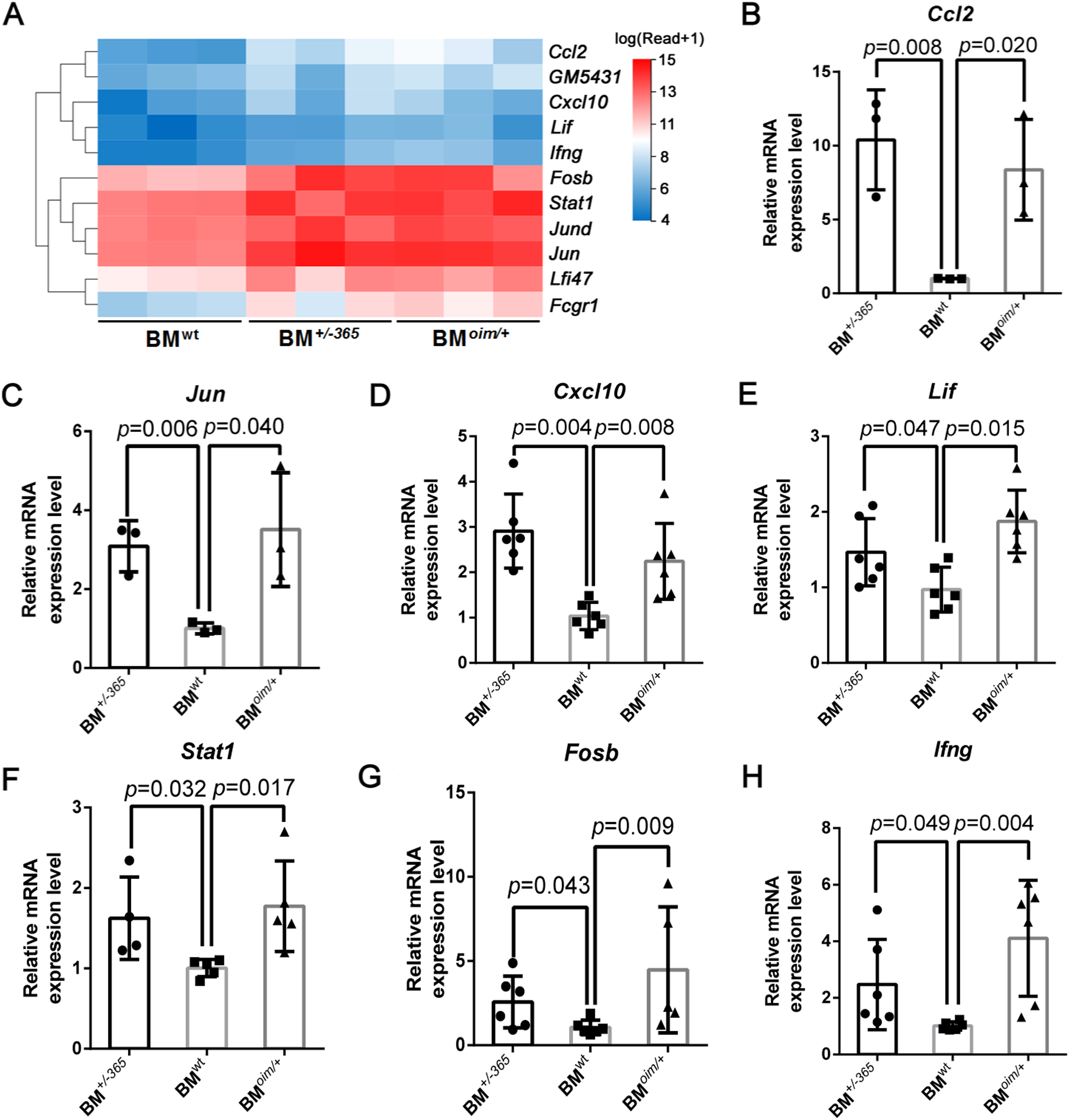{kind=link}
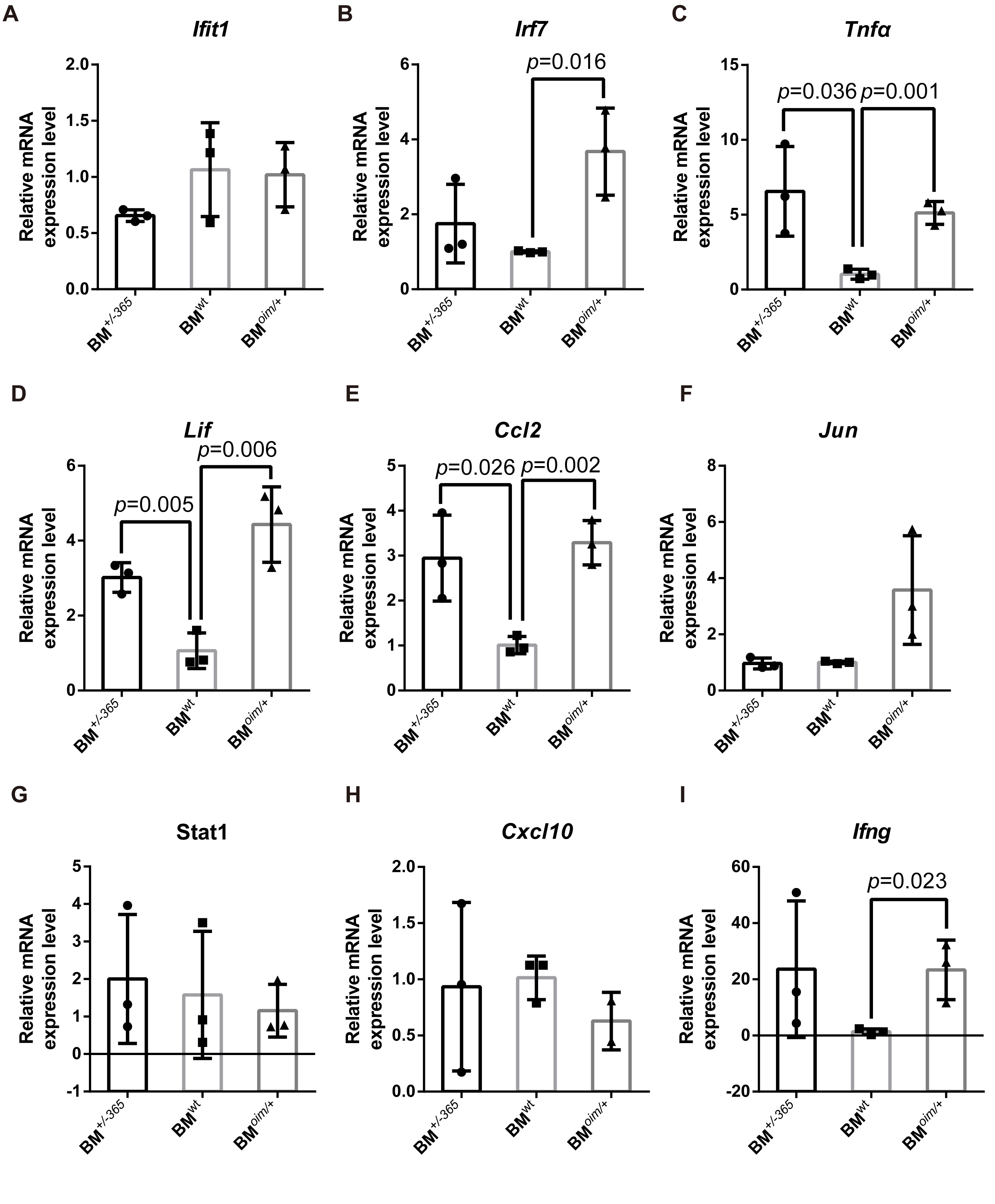
Besides, the expression of these genes in adult mice was also detected by RT-PCR. Data showed that Tnfα (Fig. S1C), Lif (Fig. S1D), and Ccl2 (Fig. S1E) were still elevated in mutant BM cells when compared to the normal cells, although there were no differences in the expression of Ifit1 (Fig. S1A), Jun (Fig. 1F), Stat1 (Fig. S1G) and Cxcl10 (Fig. S1H). The obviously increased level of Irf7 (Fig. S1B) and Ifng (Fig. S1I) in BMoim/+ instead of BM+/−365 could be observed in RT-PCR assay. The IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation under OI background remained up-regulated until adulthood.
Discussion
OI is mainly caused by the defective production or assembly of type I collagen, making bone tissue severely affected (Forlino et al., 2011; Hoyer-Kuhn, Netzer & Semler, 2015). OI patients with COL1A1 or COL1A2 mutations can be classified into OI type I-IV phenotypes (Forlino et al., 2011). OI type I due to decreased synthesis of collagen fibrils shows the mildest phenotype, while structurally aberrant type I collagen can give rise to much more severe type II–IV. Despite the subtypes and caused pathogenic variants, high bone turnover represented by hypercellular osteocytes but insufficient mineralization together with enhanced bone absorption generally exist in collagen defect-related OI (Liu et al., 2019; Lopez Franco et al., 2005; McBride, Shapiro & Dunn, 1998; Saban et al., 1996). Many studies have focused on the mechanism underlying the unit bone formation insufficiency, while excessive bone erosion has not been extensively explored.
Bone resorption is mainly mediated by osteoclasts that originate from HSC-derived monocytes (Boyle, Simonet & Lacey, 2003). The BM monocytes can differentiate into mature multinucleated osteoclasts in the presence of macrophage-colony stimulating factor (M-CSF) and receptor activator of nuclear factor kappa B ligand (RANKL) (Boyle, Simonet & Lacey, 2003; Feng, Guo & Li, 2019). It has been proved that inflammatory cytokines such as TNFα can act synergistically with RANKL to promote osteoclastotogenesis (Zhao et al., 2012). Recently, the inflammatory component in OI pathogenesis has been increasingly attractive. The chronic inflammation state of OI murine models and pediatric patients has been suggested by their elevated serum levels of inflammatory cytokines including IFN and TNFα (Zhytnik et al., 2020; Brunetti et al., 2016). Many clues suggest that the cells and cytokines of the BM niche can regulate the maturation and function of osteoclasts through direct contact and the paracrine effect (Herrmann & Jakob, 2019; Ciucci et al., 2015). But the inflammatory changes in BM cells are still unclear.
OI is a type of congenital genetic disease, with the most severe symptoms in childhood (Paterson, McAllion & Stellman, 1984). Therefore, exploration of the molecular changes of bone marrow cells of OI in early childhood is helpful to understand its pathogenesis.
Here, we performed RNA-seq of whole BM cells isolated from two types of OI models that respectively stimulate abnormal quantity and structure collagen-related OI. Data showed that the BM cells from the two young murine models shared 117 DEGs (Q ≤ 0.05, |Log2FC| ≥ 1) (Fig. 1). A total of 17 of the top 20 upregulated shared DEGs were the same genes, including several Ifn signaling-related genes containing Lfit1, Lfit3, Lfi44 and Irf7 (Tables 2 and 3). Consistent with the findings of Zhytnik’s group, GO biology process analysis also enriched multiple DEGs involved in the cellular response to Ifnβ and Ifnα in defective BM cells (Fig. 2), suggesting the activated Ifn signaling in OI BM. Transcription factor STAT1 is a key effector of the IFN pathway (Michalska et al., 2018). The upregulated Stat1 in OI BM cells (Fig. 4F) further revealed the dysregulated Ifn signaling. IFNs are key cytokines for both innate and adaptive immune responses (Takayanagi et al., 2005). Some previous studies found that IFN-γ can promote osteoclastogenesis under T-cells activation and enhance the multinucleation of myeloid lineage cells in osteoporosis (Biros et al., 2022). Thus the overactivated Ifn signaling might also act on the bone resorption in the OI background.
KEGG and GSEA analysis of the 117 shared DEGs showed that the IL-17 signaling pathway, Tnf signaling pathway and osteoclast differentiation were significantly upregulated in OI BM cells (Fig. 3). A total of 11 enriched DEGs were involved in the three pathways and displayed complex crosstalk (Table 4). Il17a, Tnf and Rankl showed different degrees of upregulation (Table 5), indicating the chronic inflammation and enhanced osteoclastogenesis. RT-PCR assay confirmed the elevated expression of some DEGs in young and adult mice (Fig. 4, Fig. S1). The results of 4-weeks old mice were consistent with the results of RNA-seq (Fig. 4). While only several genes remained elevated in 12-weeks old OI mice (Tnfα, Lif, and Ccl2) (Fig. S1), which might be related to the relieved bone phenotype in adulthood. IL-17A and TNFα have been proved to play an essential role in inflammatory bone erosion by inducing the production of RANKL (Weitzmann, 2017; Zhao et al., 2012). IL-17A can also promote TNFα secretion. Matthews et al. (2017) found the increased serum level of Tnf in homozygous oim mice, however, anti-TNFα therapy failed to reduce bone absorption. These results suggested that blocking Tnf signaling alone is insufficient to effectively reverse the excessive bone resorption in OI bones. Targeting both IL-17 and Tnf signalings may be an efficient strategy for OI treatment.
Conclusions
This study preliminarily revealed the dysregulated biological process of cellular response to Ifn together with upregulated IL-17 signaling, Tnf signaling and osteoclast differentiation in young male OI BM niche by RNA-seq. Either defective collagen production or abnormal collagen assembly shared similar alterations in gene profiles and pathways involving inflammation and osteoclast activation. Data presented here not only contributed to understanding the mechanism of the enhanced bone absorption in the bone of OI, but also provided more evidence to develop potential anti-inflammation therapies.
Supplemental Information
The original ct value of mRNA expression detected by Real-time PCR.
The original ct value of mRNA expression detected by Real-time PCR.
The expression of the shared DEGs involving IL-17 signaling pathway, Tnf signaling pathway and Osteoclast differentiation in 12-weeks BM+/−365 and BMoim/+ when compared with BMwt.
(A–I) Real-time PCR tested the expression levels of some DEGs. The result indicated that Tnfα (C), Lif (D), and Ccl2 (E) were upregulated in 12-weeks of mutant BM cells (n = 3 for each group, P < 0.05). No obvious expression difference of Ifit1 (A), Jun (F), Stat1 (G) and Cxcl10 (H) in defect OI BM and normal OI BM (n = 3 for each group). Irf7 (B) and Ifng (I) were increased alone in BMoim/+ in 12-weeks (n = 3 for each group, P < 0.05). Data in the quantitative plots are presented as mean ± SD using upaired t-test.
{kind=link}