
Vibrio alginolyticus influences quorum sensing-controlled phenotypes of acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus
- Published
- Accepted
- Received
- Academic Editor
- Marcelino Suzuki
- Subject Areas
- Aquaculture, Fisheries and Fish Science, Microbiology, Molecular Biology, Zoology, Freshwater Biology
- Keywords
- AHPND, Biofilms, Flagella, LuxR, Motility, OpaR, Probiotic, Quorum sensing, V. alginolyticus, V. parahaemolyticus
- Copyright
- © 2021 Paopradit et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. Vibrio alginolyticus influences quorum sensing-controlled phenotypes of acute hepatopancreatic necrosis disease-causing Vibrio parahaemolyticus. PeerJ 9:e11567 https://doi.org/10.7717/peerj.11567
Abstract
Background
Acute hepatopancreatic necrosis syndrome (AHPND) caused by Vibrio parahaemolyticus strain (VPAHPND) impacts the shrimp industry worldwide. With the increasing problem of antibiotic abuse, studies on quorum sensing (QS) system and anti-QS compounds bring potential breakthroughs for disease prevention and treatment.
Methods
In this study, the cell-free culture supernatant (CFCS) and its extract of V. alginolyticus BC25 were investigated for anti-QS activity against a reporter bacteria, Chromobacterium violaceum DMST46846. The effects of CFCS and/ or extract on motility, biofilm formation and extracellular polymeric substances (EPSs) of VPAHPND PSU5591 were evaluated. Moreover, the effects of V. alginolyticus BC25 on virulence of VPAHPND PSU5591 were investigated by shrimp challenge test. The potentially active anti-QS compounds presented in the extract and effect on gene expression of VPAHPND PSU5591 were identified.
Results
The CFCS of V. alginolyticus BC25 and its extract showed a significant anti-QS activity against the reporter bacteria as well as swimming and swarming motilities, biofilms, and EPSs production by VPAHPND PSU5591. Transcriptome analysis revealed that V. alginolyticus BC25 extract significantly reduced the flagella genes involved in biofilm formation and iron-controlled virulence regulatory gene of VPAHPND PSU5591. Whereas, the LuxR family transcriptional regulator gene, c-factor, a cell-cell signaling gene, and capsular polysaccharide were up-regulated. The potentially active anti-QS compounds identified in extract were Cyclo-(L-Leu-L-Pro), and Cyclo-(L-Phe-L-Pro). Furthermore, V. alginolyticus BC25 enhanced disease resistance against VPAHPND PSU5591 in tested shrimp larvae.
Conclusion
These findings suggest that V. alginolyticus BC25 could provide natural anti-QS and anti-biofilms compounds and has great ability to be used as biocontrol agent against VPAHPND infection in shrimp aquaculture.
Introduction
In recent years, the global shrimp aquaculture industry has suffered serious losses from acute hepatopancreatic necrotic disease (AHPND) outbreak caused by a unique Vibrio parahaemolyticus strain (VPAHPND). First mass mortality of shrimp caused by AHPND occurred in China in 2009. After that, this disease has spread to several countries including Vietnam, Malaysia, Thailand, Mexico, Philippines, and Latin America (FAO, 2013; Nunan et al., 2014; De la Pena et al., 2015). Almost 80% of the economic loss in shrimp industry was estimated in China due to the catastrophic disease (Lightner et al., 2013). Moreover, in Vietnam, the shrimp production was reduced from 70,000 tons in 2010 to 30,000 tons in 2012, respectively (Zorriehzahra & Banaedrakhshan, 2015). In Malaysia, AHPND decreased the shrimp production from 70,000 tons in 2010 to 40,000 tons in 2011 (Leano & Mohan, 2012). In Thailand, the shrimp production dropped from 600,000 tons in 2011 to less than 200,000 tons in 2014 (Thitamadee et al., 2016). Also in Mexico, the disease led to an economic loss of approximately 118 million USD (Schryver et al., 2014). To overcome the disease outbreak in shrimp aquaculture, several chemicals and antibiotics are used such as chlorine, potassium permanganate, ampicillin, streptomycin, sulfamethoxazole, fosfomycin and bicozamycin (Lai et al., 2015; Boonyawiwat et al., 2017). However, antibiotic resistance of VPAHPND could potentially be transferred among bacterial species in aquaculture systems. It has been reported that VPAHPND from penaeid shrimp in Mexico demonstrated a high level of resistance to tetracycline (≥5μg/mL) and carried the tetB gene coding for tetracycline resistance (Han et al., 2015). In the environment, multiple bacterial species coexist as communities. Bacteria have a mechanism for cell to cell communication as a quorum sensing system (QS) through the use of small signaling molecule called autoinducer. They use signaling molecule to coordinate their behavior such as bioluminescence, biofilm formation, spore formation, motility and developing antibiotic resistance (Ryan & Dow, 2008). QS system in V. parahaemolyticus is similar with that of V. harveyi (Zhang et al., 2012). V. parahaemolyticus produces harveyi autoinducer 1 (HAI-1), autoinducer 2 (AI-2) and cholerae autoinducer 1 (CAI-1). At low cell density, a phosphate group from the receptor transferred to LuxU and LuxO regulator. The phosphorylated LuxO activates the small quorum regulatory RNAs (Qrrs) transcription and promotes the translation of AphA master regulator and inhibit the mRNA of LuxR or OpaR regulator. At high cell density, the autoinducers are bound to receptors lead to dephosphorylation of LuxU and LuxO regulator. The Qrr genes are not transcribed, resulting in an expressed LuxR or OpaR regulator (Gode-Potratz & McCarter, 2011; Zhang et al., 2012). It has been reported that OpaR regulator regulates colony morphology, motility, capsule polysaccharide (CPS) production, type III and VI secretion system production (T3SS, T6SS) (Gode-Potratz & McCarter, 2011; Zhang et al., 2012; Wang et al., 2013; Salomon, Klimko & Orth, 2014; Kernell Burke et al., 2015; Wu et al., 2019). Therefore, QS is one of the alternative strategies for controlling virulence and behavior of bacteria.
V. alginolyticus is closely related to V. parahaemolyticus due to their similar biochemical characteristics and flagellar systems (Kawagishi et al., 1995; Montieri et al., 2010). In 1965, it has been reported from a taxonomic numerical study that V. alginolyticus is regarded as biotype 2 V. parahaemolyticus. Vibrio parahaemoltyicus historically had included two biotypes. The similarity value between two biotypes was approximately 80%, but the differentiating characters was sucrose fermentation, Voges-Proskauer reaction and growth in the peptone water containing 10% NaCl. Thus, the second biotype be moved and put into a new species to which the name Vibrio alginolyticus (Zen-Yoji et al., 1973). In addition, V. alginolyticus has long been employed as a probiotic in various shrimp hatcheries. It can be isolated from shrimp culture water and reportedly associates with healthy larvae and juvenile shrimp (Rengpipat et al., 2000). It is also found in the hepatopancreas and intestine of healthy Penaeus monodon and Litopenaeus vannamei (Gomez-Gil et al., 1998; Vandenberghe et al., 1999). V. alginolyticus effectively stimulates the shrimp immune response (Gullian, Thompson & Rodrýguez, 2004). In addition, it has been reported that V. alginolyticus UTM102 isolated from the gastrointestinal tract of adult shrimp L. vannamei showed antagonism against shrimp-pathogenic V. parahaemolyticus PS-017 (Balcázar, Rojas-Luna & Cunningham, 2007). For this reason, the aims of this study were to analyze the efficacy of V. alginolyticus BC25 isolated from the microbiota of marine mollusk for its anti-quorum sensing controlled phenotypes of VPAHPND and ability to enhance disease resistance against this shrimp pathogen. The active fractions of V. alginolyticus BC25 extract were identified and effects on gene expression of VPAHPND PSU5591 were also studied.
Materials & Methods
Bacterial strains and culture conditions
The natural marine bacterium V. alginolyticus BC25 was isolated from the microbiota of marine blood cockle collected in Songkhla province, Thailand and is deposited in the Thailand Bioresource Research Center (TBRC), Pathum Thani, Thailand with the Accession Numbers TBRC12771. VPAHPND PSU5591 was isolated from the hepatopancreas of VPAHPND-infected shrimp in Songkla, Southern Thailand. Chromobacterium violaceum DMST46846 used in this study was purchase from DMST Culture Collection, Department of Medical Sciences Thailand. Bacteria were cultured at 30 °C in Luria-Bertani (LB) or LB containing 1% NaCl medium.
Screening for anti-QS activity
The cell-free culture supernatant (CFCS) of V. alginolyticus was screened for its anti-QS activity by testing with C. violaceum. To prepare the CFCS of V. alginolyticus, the bacteria were cultured in LB broth with 1% NaCl, and incubated at 30 °C for 24 h. Cells were removed by centrifugation at 12,000 × G for 15 min. The CFCS was sterilized by sonication. Briefly, CFCS was placed in 50 mL sterile conical and sonicated by submerging the probe tip of a 20 kHz (size 12.7 mm), 45% amplitude with pulse mode 15 s and a 5 s rest for 15 min (Joyce, Al-Hashimi & Mason, 2011). The inhibition of violacein production in C. violaceum was analyzed as described previously with slight modification (Fitriani, Ayuningtyas & Kusnadi, 2016). In brief, C. violaceum was cultured in LB broth, incubated at 30 °C for 4 h, and adjusted to 0.5 McFarland (1.0 × 108 CFU/mL). Before treatment, 30 µL of freshly grown C. violaceum culture was mixed with different concentrations (6.25, 25, 50, and 100%, v/v) of CFCS and incubated at 30 °C with shaking 150 rpm for 24 h. The total of viable bacteria was counted on LB agar. All treatments were centrifuged at 12,000 × G for 10 min and the pellets C. violaceum were resuspended in two mL dimethylsulfoxide (DMSO) to extract the violacein. Extracts were centrifuged at 12,000 × G for 10 min and 200 µL of CFCS was read at 585 nm using a microplate reader (LUMIstar Omega, BMG LABTECH, Germany) to check for the presence (or absence) of violacein.
Preparation of Vibrio alginolyticus extract and anti-QS assay
V. alginolyticus was cultured for 24 h at 30 °C in LB broth with 1% NaCl. The culture was centrifuged for 15 min at 12,000 × G. The sterilized CFCS was extracted using an equal volume of ethyl acetate (CH3COOCH2CH3), concentrated using a rotary evaporator at 45 °C, dissolved in DMSO, and stored at −20 °C (Nithya, Begum & Pandian, 2010c). The anti-QS activity of different concentration of V. alginolyticus extract (25, 50, and 100%, v/v) was re-tested by an agar well assay using C. violaceum as a reporter strain. Serratia marcescens and DMSO were considered as positive and negative control, respectively. After 24 h of incubation, the transparent halo of violacein inhibition indicates anti-QS activity.
Effect of V. alginolyticus on QS-controlled phenotypes of VPAHPND
Swimming and swarming motility assay
Motility tests was performed as described previously with slight modifications (Zhang et al., 2016). These assays were conducted on LB agar with 1% NaCl supplemented with or without different concentrations of CFCS (40, 60, and 80%) and its extract. For swimming motility test, VPAHPND was cultured in LB broth, and 3 µL aliquots (0.5 McFarland) was spotted on the center of 0.3% (w/v) agar plates. The diameter zone of the motility were measured and photographed. For swarming motility test, VPAHPND was streaked as a straight line on 1.5% (w/v) agar and incubated at 30 °C for 18 h. The diffuse zone surrounding the inoculum lines were measured and photographed.
Biofilm formation assay
The anti-biofilm activity assay of VPAHPND was determined as described previously with slight modifications (Nesper et al., 2001). Briefly, an overnight VPAHPND culture in LB broth was added to a 96-well microtiter plate with or without V. alginolyticus extract (2.5, 5, and 10%) and incubated at 30 °C for 24 h. Planktonic cells were removed by washing the wells with sterile water, fixed with 2.5% glutaraldehyde, and stained with 0.4% (w/v) crystal violet. After that ethanol-acetone (80:20, v/v) was added. The OD was determined at 570 nm in a microplate reader (LUMIstar Omega, BMG LABTECH, Germany).
The effect of V. alginolyticus extract on EPSs in biofilm was visualized by scanning electron microscopy (SEM) as described previously with minor modifications (Singh, Mishra & Jha, 2017). Briefly, biofilms of VPAHPND grown on glass coverslips (1.0 × 1.0 cm) submerged in LB broth with or without V. alginolyticus extract in 24-well plate were gently washed with phosphate buffer saline (PBS) to remove planktonic cells. Afterwards the cells were fixed with 2.5% glutaraldehyde (v/v) at 4 °C for 12 h. After dehydrating in a gradient ethanol series (30–100%, v/v) for 15 min each, the dried samples were coated with gold and investigated under a SEM (Quanta 400, Thermo Fisher Scientific, USA).
Gene expression analysis
The Effect of V. alginolyticus extract on gene expression of VPAHPND was analyzed using RNA sequencing (RNA-seq) analysis. VPAHPND was treated with or without V. alginolyticus extract as a control at 30 °C for 24 h. The bacterial culture was centrifuged at 8,000 × G for 10 min, and RNA extraction was initially done by stabilizing RNA using RNAprotect Bacteria Reagent (Qiagen, Germany). Total RNA from the two samples was extracted using the TRIzol™ Reagent (Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA) following the manufacturer’s protocol. All RNAs were stored at −80 °C immediately until use. RNA sequencing was performed using a NovaSeq 6000 system (Macrogen, Korea). Sequence data was mapped to reference genome with Bowtie software 1.1.2 (Bolger, Lohse & Usadel, 2014). Read count per gene was extracted from known gene annotations with HTSeq program version 0.10.0 (Anders, Theoder Pyl & Huber, 2015). The evaluating gene expression profiles were performed using FPKM method (fragments per kilobase of transcript per million mapped reads) as a normalization value. Statistical analysis was performed using fold change, exactTest using edgeR per comparion pair (Robinson, McCarthy & Smyth, 2010). The significant results are selected on conditions of lfcl ≥ 2 and exactTest raw p-value < 0.05
Separation and identification of V. alginolyticus extract
V. alginolyticus extract was fractionated by high-performance liquid chromatography (HPLC) 1260 (Agilent Technologies, USA). One hundred microliter of extract was injected onto a non-polar reverse-phase C18 column (50 × 2.1 mm, Waters, CA, USA). The mobile phase was methanol and water (20:80, v/v) at 30 °C at a flow rate of 0.2 mL/min. Each fraction was collected, concentrated by rotary evaporator, and re-tested for anti-QS activity as described above. The positive fractions were further analyzed by gas chromatograph-mass spectrometer (GC-MS) (Agilent Technologies 5977A, USA) as described previously (Savka et al., 2015). The spectra were matching with the National Institute of Standards and Technology (NIST) reference library to identify the anti-QS compounds based on retention indices. The molecular structure of active compound was confirmed by nuclear magnetic resonance spectroscopy (NMR). The active compound was dissolved in deuterated MeOH (CD3OD), and subjected to 1H and 13C NMR analyses using a NMR Bruker/Avance (500 MHz for 1H and 125MHz for 13C).
V. alginolyticus treatment and challenge test
Litopenaeus vannamei were obtained from a shrimp farm in Songkhla province, Thailand. The shrimp larvae (postlarval age = 25 days) were acclimatized for 3 day before transferring to glass tanks containing 3 L artificial sea water (Marinum, Thailand) (15 shrimp/tank). The experiment consisted of four groups: group 1 (treated with V. alginolyticus at a concentration of 105 CFU/mL to investigate the possible virulence of V. alginolyticus); group 2 (treated with VPAHPND at a concentration of 106 CFU/mL as positive control) and group 3 (treated with V. alginolyticus three days before infection with VPAHPND to test the ability of V. alginolyticus for the control of VPAHPND disease in shrimp) and only shrimp larvae (no any treatment) as negative control , as described previously (Rodriguez et al., 2011; Joshi et al., 2014). Each treatment was carried out in triplicates. Mortality was recorded until 72 h post infection.
Statistical analysis
Effects of different treatments were tested by one-way ANOVA analyses using the SPSS software version 14 (SPSS Inc., USA). Differences supported by a p-value of less than 0.05 were considered as significant.
Results
V. alginolyticus showed anti-QS activity by inhibiting violacein production
A lack of the purple pigment violacein from indicator strain indicates a potential anti-QS result. Our results revealed that CFCS of V. alginolyticus and its extract showed anti-QS activity. The reduction of violacein production in C. violaceum are CFCS concentration-dependent manner. About 79.7% inhibition (P < 0.001) was observed with the addition 50% (v/v) CFCS and without affecting to C. violaceum growth (Fig. 1A). Different concentrations of V. alginolyticus extract on violacein production were re-tested by agar well diffusion assay. As expected, the violacein production reduced with the increasing concentration of V. alginolyticus extract. However, V. alginolyticus extract inhibited the growth of C. violaceum at high concentration (Fig. 1B).
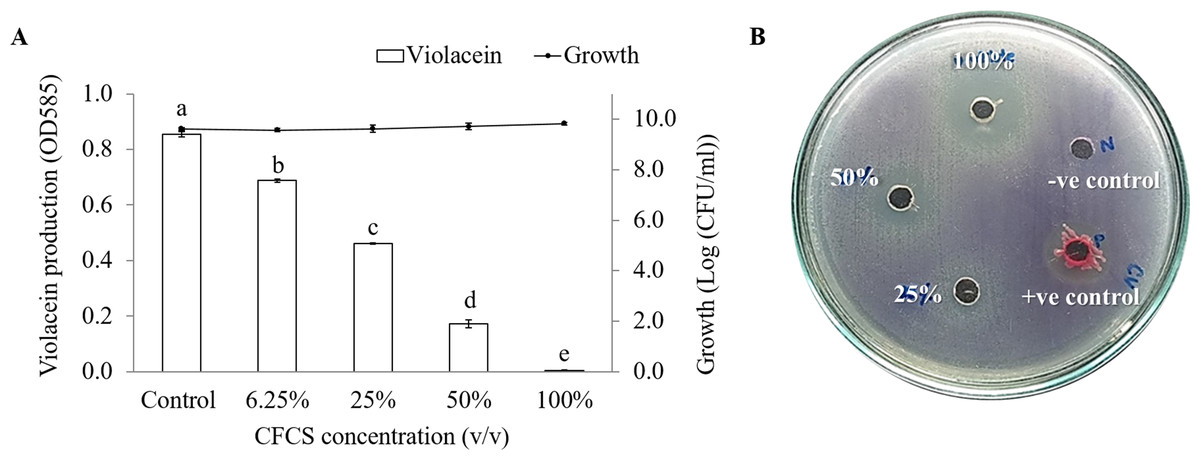
Figure 1: The inhibition of violacein production in C. violaceum DMST46846.
(A) Different concentration of CFCS of V. alginolyticus BC25 was used to detect the reduction of purple pigment as indicator for QS activity. Different letters (a, b, c, d, and e) are significant difference (p < 0.001) among the treatments. (B) Screening of anti-QS activity of V. alginolyticus BC25 extract by agar well diffusion method. DMSO solvent and S. marcescens were used as negative and positive control, respectively.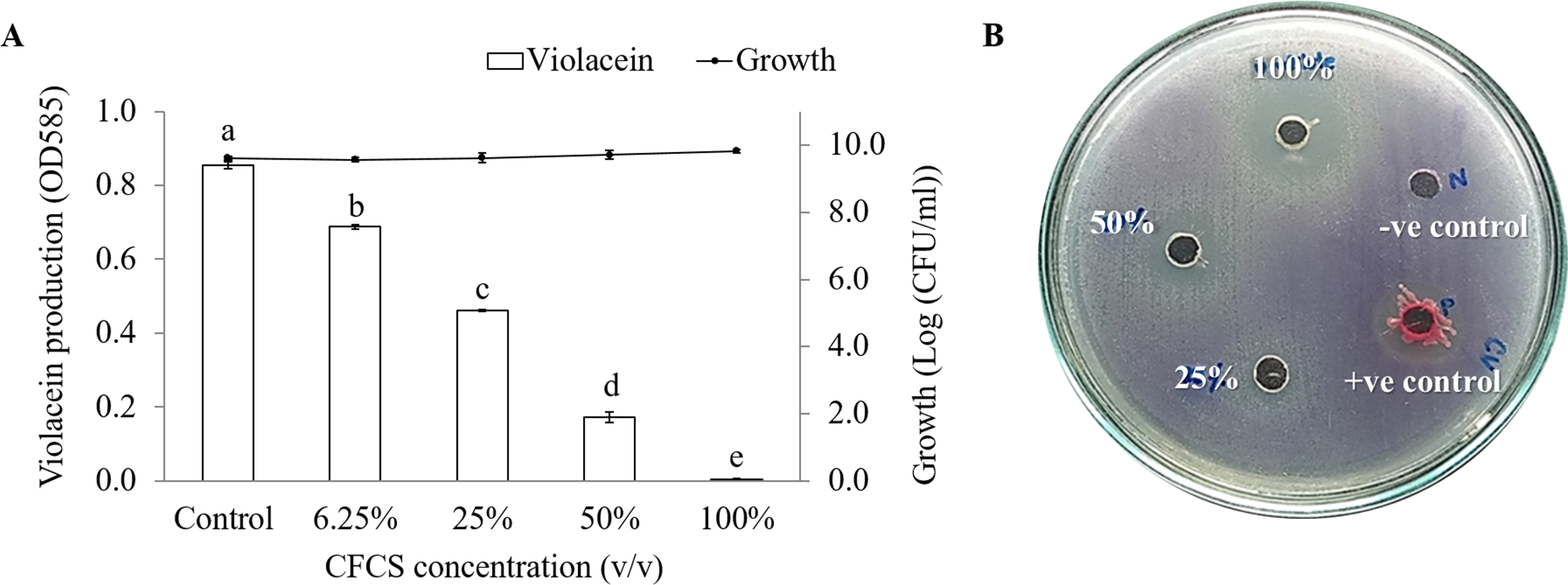{kind=link}
V. alginolyticus inhibited motilities and biofilms of VPAHPND
It was observed that CFCS of V. alginolyticus reduced the swimming and swarming motilities of VPAHPND (Fig. 2). The CFCS at 80% significantly decreased the diameter of the swimming and at 40 to 80% CFCS also decreased the swarming area, which significantly differs (p < 0.01) when compared with control as shown in Table 1. As expected that V. alginolyticus extract greatly reduced the swimming and swarming motilities of VPAHPND at 2.5% (v/v) with significant differences (P < 0.001) (Fig. 3, Table 2). The anti-biofilm activity of V. alginolyticus extract with a different concentration (2.5, 5, and 10%) was tested against VPAHPND. About 36.6% inhibition of biofilm formation with P <0.001 was noticed with 2.5% (v/v) extract without significant effect on cell growth. However, the growth inhibition was observed with high extract concentrations used (Fig. 4). Thus, similar concentration of V. alginolyticus extract was employed to test for a possible effect on the architecture of the VPAHPND biofilm by SEM analysis. A thick and dense biofilm along with EPSs production was observed in control after 24 and 48 h incubation (Figs. 5A, 5C), whereas the reduction in VPAHPND EPSs was observed in treated samples (Figs. 5B, 5D).
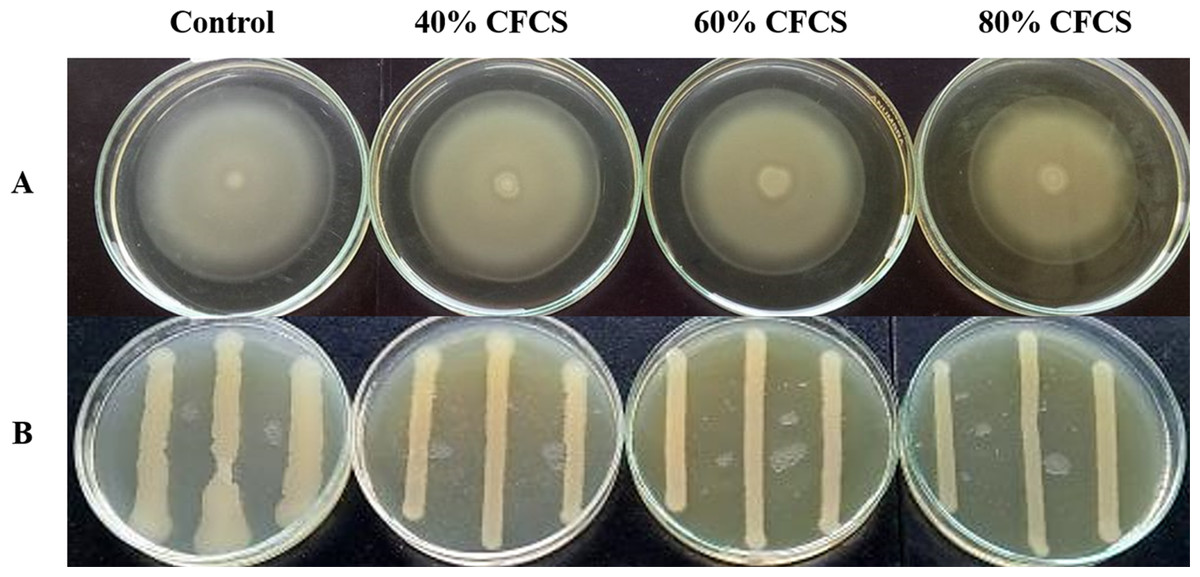
Figure 2: Motility of VPAHPND PSU5591 in different concentration of CFCS of V. alginolyticus BC25.
(A) Swimming motility of VPAHPND on 0.3% (w/v) LB agar by spot method. (B) Swarming motility of VPAHPND on 1.5% (w/v) LB agar by streak method. The motility zone was measured and provided as mean ± standard deviation as described in Table 1.{kind=link}
| Motility | CFCS concentration (%, v/v) | Motility zone (mm) |
|---|---|---|
| Swimming | Control | 66.20 ± 0.21a |
| 40 | 64.70 ± 0.04a | |
| 60 | 61.78 ± 0.41a | |
| 80 | 55.28 ± 0.65b | |
| Swarming | Control | 10.68 ± 1.08a |
| 40 | 7.12 ± 0.42b | |
| 60 | 6.87 ± 0.46b | |
| 80 | 6.12 ± 0.40b | |

Figure 3: Motility of VPAHPND PSU5591 in the presence of V. alginolyticus extract.
(A) Swimming motility of VPAHPND on 0.3% (w/v) LB agar by spot method. (B) Swarming motility of VPAHPND on 1.5% (w/v) LB agar by streak method. The motility zone was measured and provided as mean ± standard deviation as described in Table 2.{kind=link}
| Motility | Concentration (%, v/v) | Motility zone (mm) |
|---|---|---|
| Swimming | Control | 46.52 ± 1.32b |
| DMSO | 51.10 ± 0.92b | |
| 2.5% extract | 4.60 ± 0.41a | |
| Swarming | Control | 7.15 ± 0.69b |
| DMSO | 5.38 ± 0.12b | |
| 2.5% extract | 0.97 ± 0.16a |
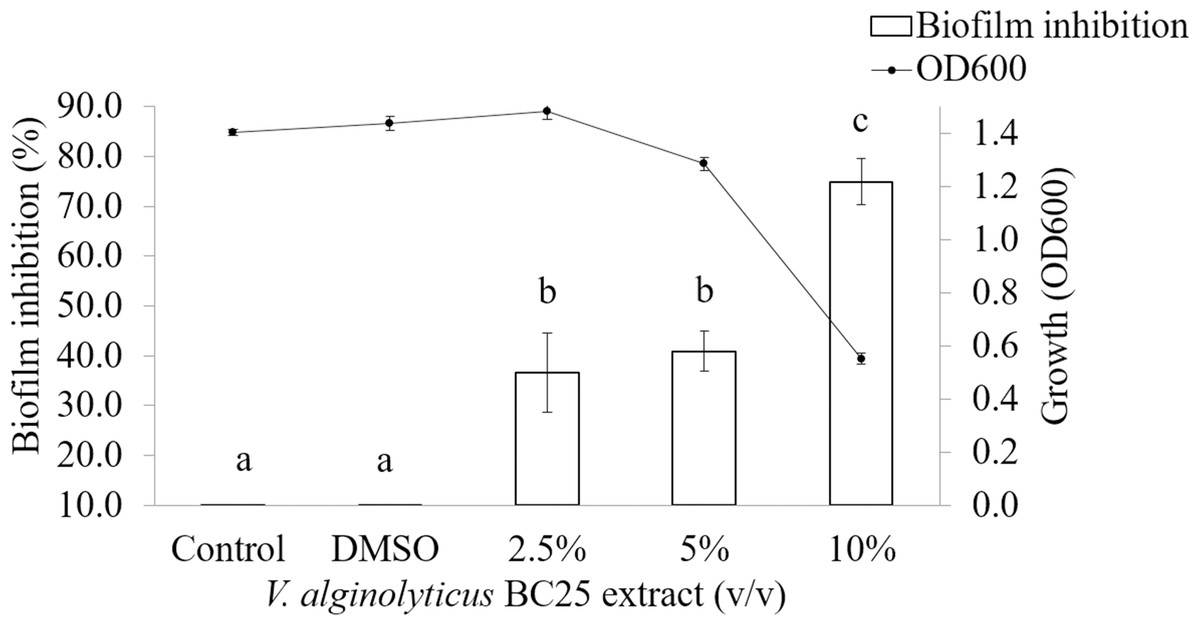
Figure 4: Inhibition of biofilm formation in VPAHPND PSU5591 by different concentration of the V. alginolyticus BC25 extract.
Different letters (a, b, and c) are significant difference (p < 0.001) among the treatments.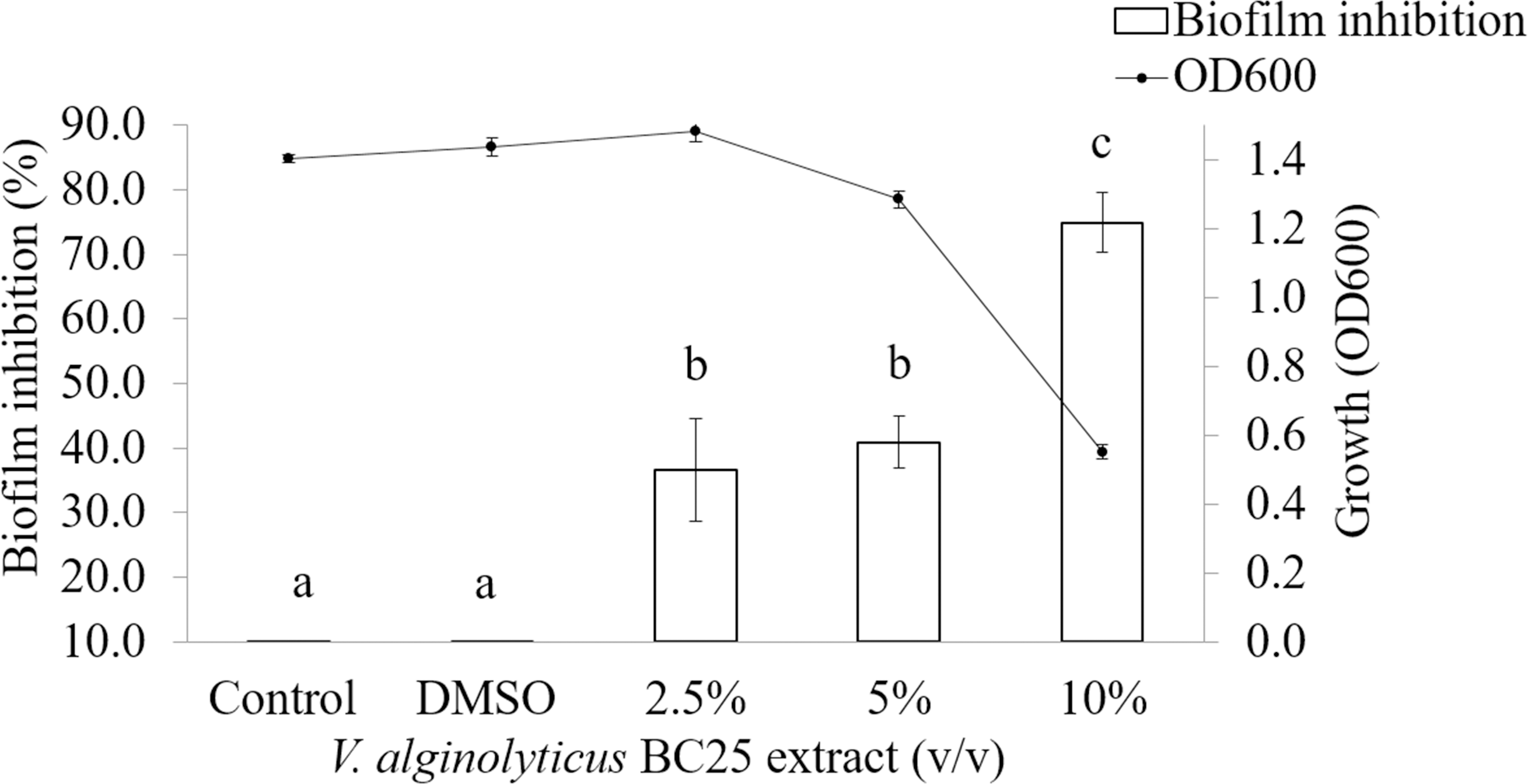{kind=link}

Figure 5: Inhibition of EPSs in biofilm of VPAHPND PSU5591 by 2.5% (v/v) V. alginolyticus BC25 extract in 24 and 48 h cultivation.
Representative photographs by SEM. (A) control at 24 h, (B) treated at 24 h, (C) control at 48 h and (D) treated at 48 h.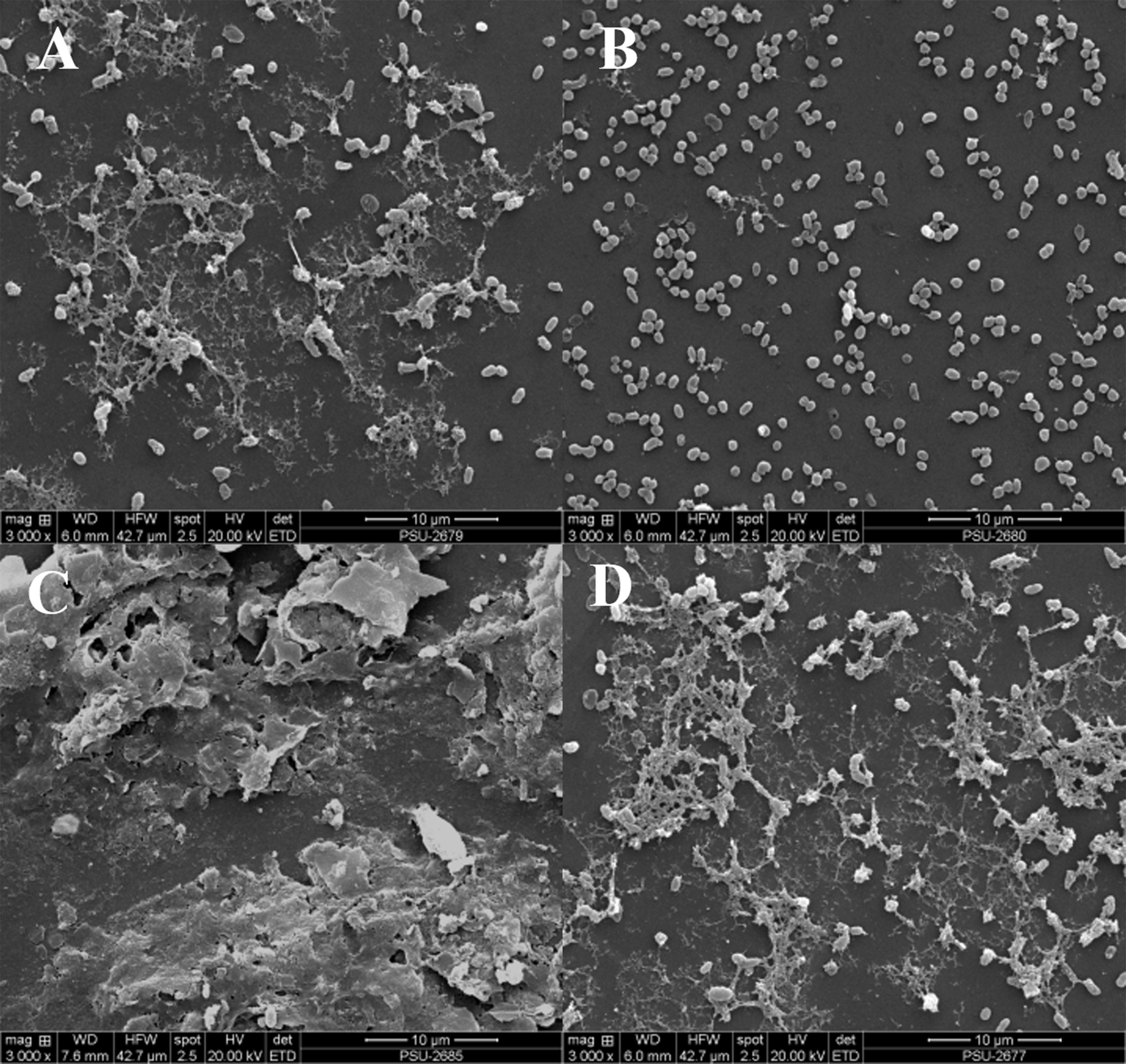{kind=link}
V. alginolyticus extract affected the expression of QS regulatory genes of VPAHPND
For better understanding of the effect of V. alginolyticus extract on global gene expression of VPAHPND, cultures grown with or without 2.5% (v/v) extract were analyzed using RNA-Seq. A total of 6.3 Gb sequence data, including 62,669,812 raw reads and 48,830,934 mapped reads were obtained. The sequencing raw reads were deposited in the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) with GCA_000196095.1 reference genome and accession number SRR12398138, SRR12459902. The Q20 and Q30 percentages were higher than 98% and 95%, respectively (Table 3). A p-value less than 0.05 were chosen as limitation for screening of differentially expressed genes (DEGs). The RNA-seq data showed that 524 genes were differentially expressed, of which 259 were up-regulated and 265 were down-regulated. The volcano plot of DEGs showed the genes affected by V. alginolyticus extract were either down-regulated (blue dots, 265 genes) or up-regulated (yellow dots, 259 genes) (Fig. 6). Of the DEGs, three domains including biological process (14 subcategories): cellular process, metabolic process, localization, and biological regulation; cellular component (10 subcategories): cell part, membrane part, and organelle part; and molecular function (12 subcategories): catalytic activity, transporter activity, binding, signal transducer activity, and structural molecule activity were identified (Fig. 7). Interestingly, the expression of motility genes were highly affected by V. alginolyticus extract (Table S1).
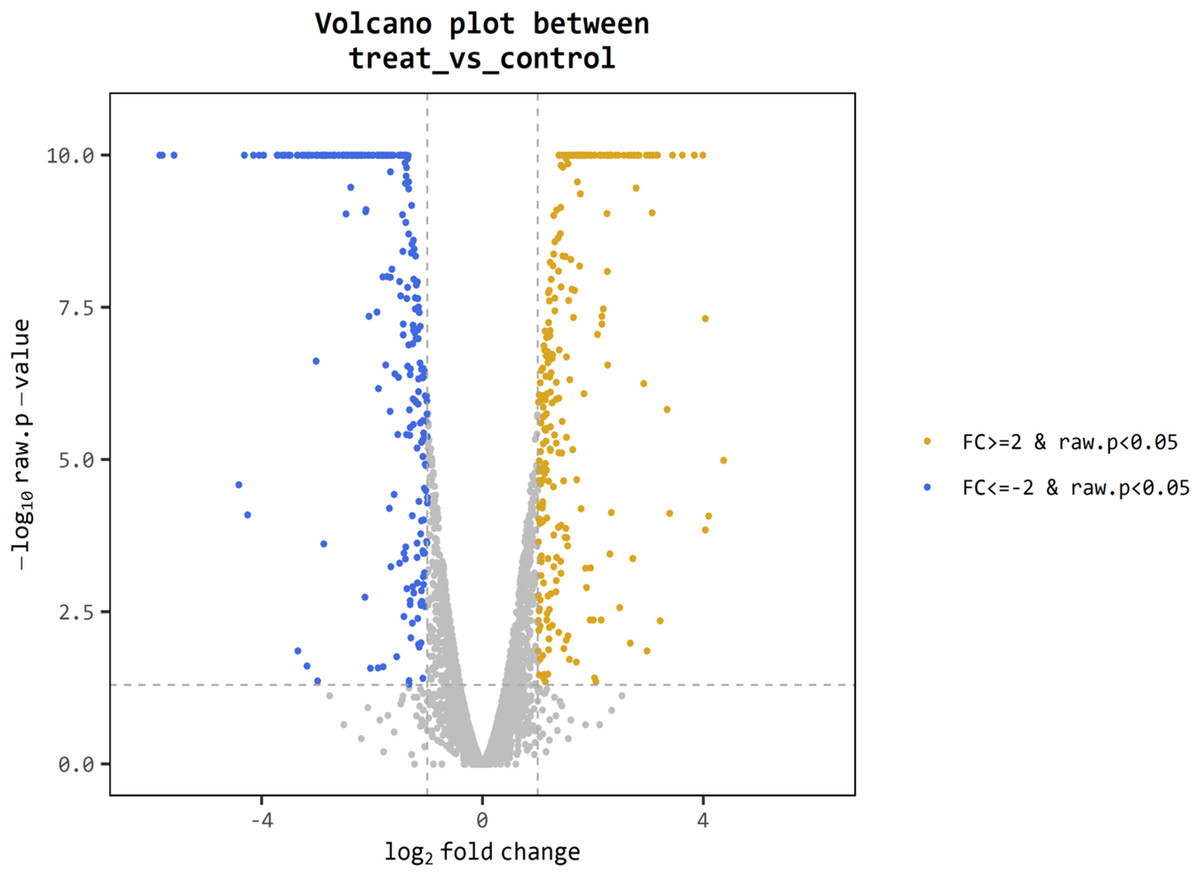
Figure 6: Differentially expressed genes (DEGs) of VPAHPND PSU5591 by volcano plot.
In total, 524 genes indicate different expression. Blue shows down-regulated expression, brown shows up-regulated expression and gray shows no significant difference when comparing control and treat sample.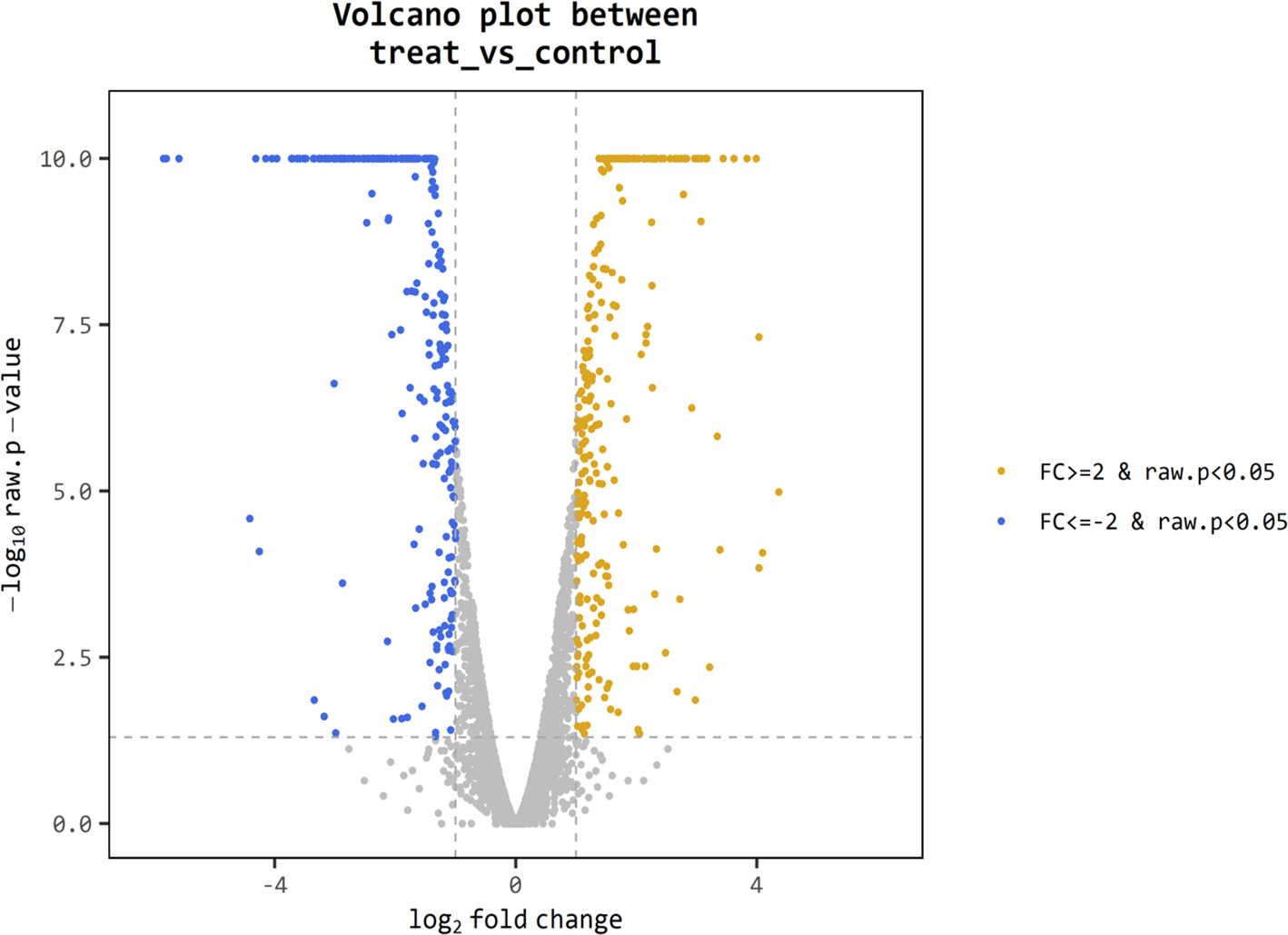{kind=link}
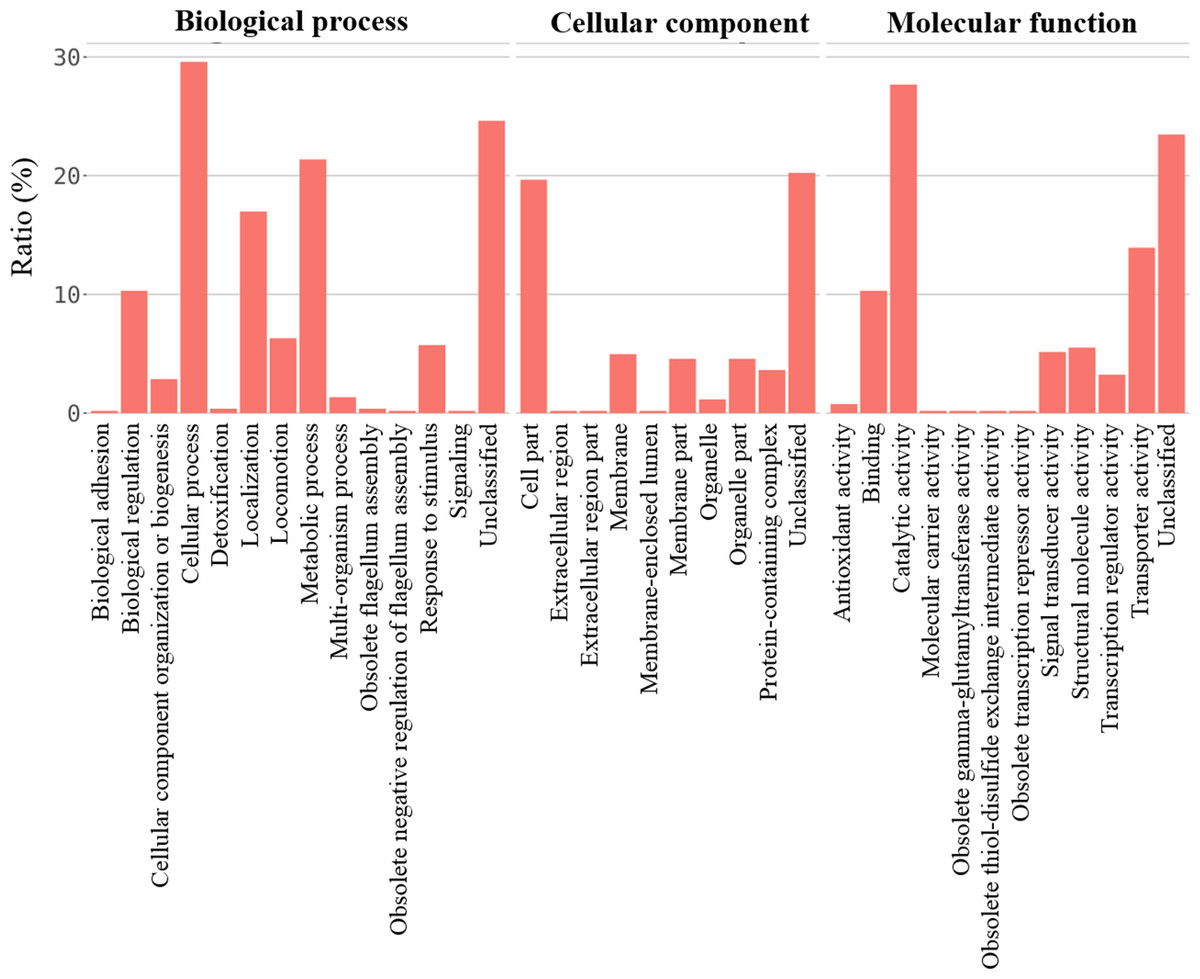
Figure 7: The gene expression associated to functional categories of VPAHPND PSU5591 culture in the presence of V. alginolyticus BC25 extract.
{kind=link}
Separation and identification of V. alginolyticus extract
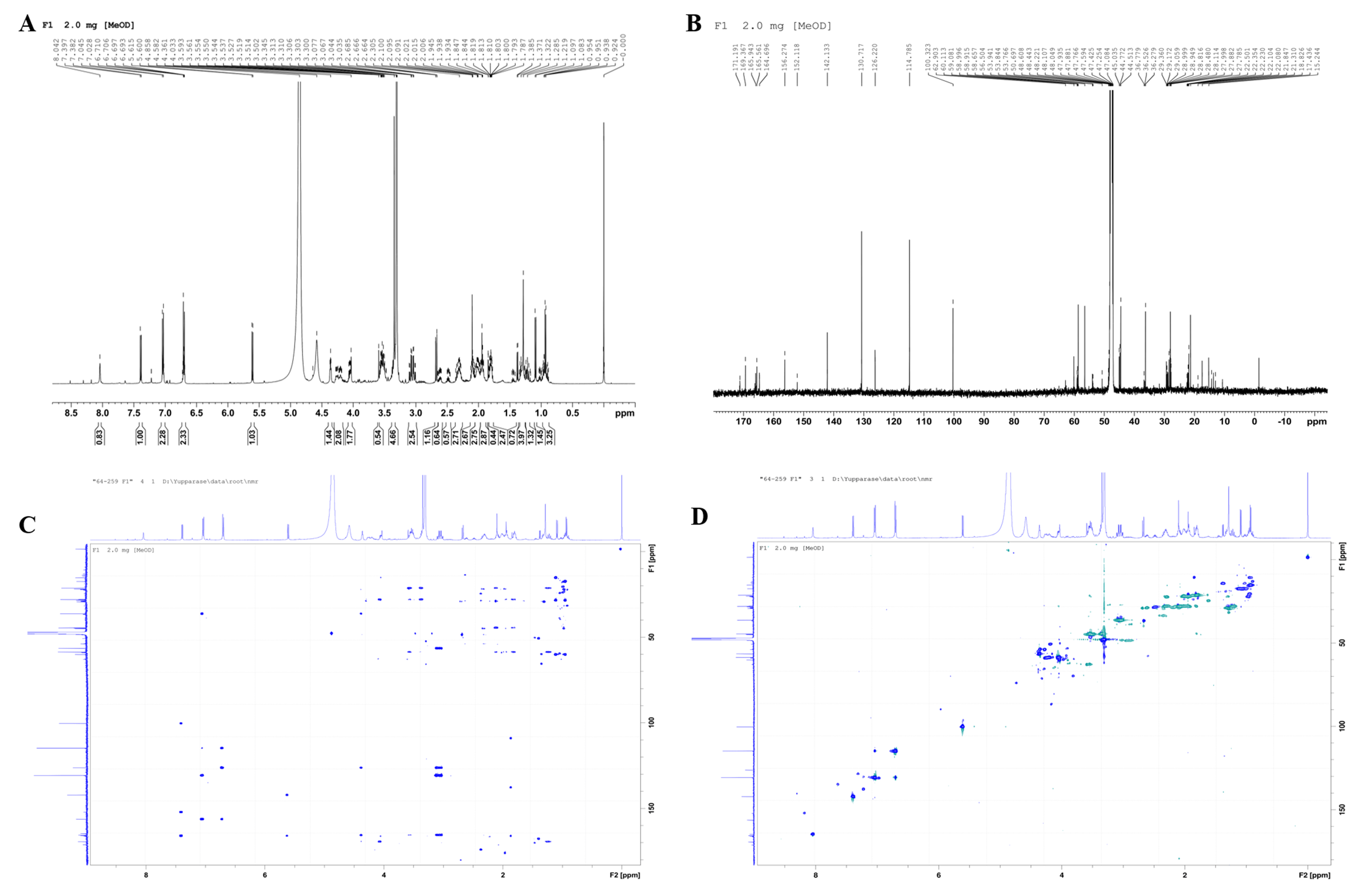
A total of seven fractions of V. alginolyticus extract were collected and individually screened for anti-QS activity using a reporter strain, C. violaceum by plate assay. The zone of QS inhibition was observed in four fractions collected (no. 1, 4, 5, and 7; Fig. S1). Therefore, these fractions were selected for further characterization by gas chromatography-mass spectrometry (GC-MS) analysis. The chromatogram showed a predominant peak at the retention time 19.44 min for fraction 1, 20.62 min for fraction 4, 27.36 min for fraction 5, and 27.38 min for fraction 7 (Dataset S1), respectively. A major mass spectral peak detected at m/z 154 for fraction 1 and 4, as well as at m/z 125 for fraction 5 and 7, was considered as active fraction. The detected mass spectra indicated some resemblance to Cyclo-(L-Leu-L-Pro) for fraction peak 1, 4 and Cyclo-(L-Phe-L-Pro) for fraction peak 5, 7 in the GC-MS library. The 1H NMR spectrum (500 MHz, MeOH, J in Hz) indicated the presence of 18 hydrogen atoms with value δ 0.92−0.95 (3H, 0.93 (d, J = 7.0 Hz)), δ 1.07−1.12 (3H, 1.09 (d, J = 7.0 Hz)), δ 1.78−1.80 (2H, m), δ 1.46 (1H, m), δ 1.83−1.98 (4H, 1.84 (d, J = 1.50 Hz), 1.94 (m), 1.95 (d, J = 1.50 Hz), 1.85 (m)), δ 4.58 (1H, (d, J = 25.0 Hz), δ 4.86 (1H, s), δ 3.47−3.61 (2H, 3.50 (m), 3.59 (m)), δ 6.69 (1H, s). The 13C NMR spectrum (125 MHz, MeOH) suggested the presence of 11 carbon atoms thus, confirming that the active compound was Cyclo-(L-Leu-L-Pro). The 13C NMR spectrum showed methane carbon in aliphatic region at δ 59.08 and 53.76 ppm. Two quaternary carbons of two C=O groups at δ 165.9 and 169.3 ppm were observed. Two methyl carbons were showed at δ 22.23 and 22.10 ppm. Furthermore, four methylene and a third methine carbons displayed at δ 47.08, 45.03, 28.48, 22.35, and 22.50 ppm, respectively. The NMR spectrum were compared with a standard compound in NMR Predict (http://www.nmrdb.org) and found to be similar to previous reported for Cyclo-(L-Leu-L-Pro) (Lalithal et al., 2016; Putra & Karim, 2019). Representative 1H-NMR, 13C-NMR spectra and 2D NMR (HMBC, HSQC) as shown in the Fig. S2.
V. alginolyticus treatment and challenge test against VPAHPND
In untreated group infected with VPAHPND alone, the mortality of shrimp started at 12 h post infection, and reached 55.56% mortality within 72 h. However, with shrimp treated with V. alginolyticus (105 CFU/mL) prior to infection with VPAHPND, the mortality of shrimp started at 24 h post infection and 80% survival was observed 72 h post-infection. During 72 h of the study, 100% survival was observed in shrimp larvae treated with V. alginolyticus suspensions (105 CFU/mL) (Fig. 8).
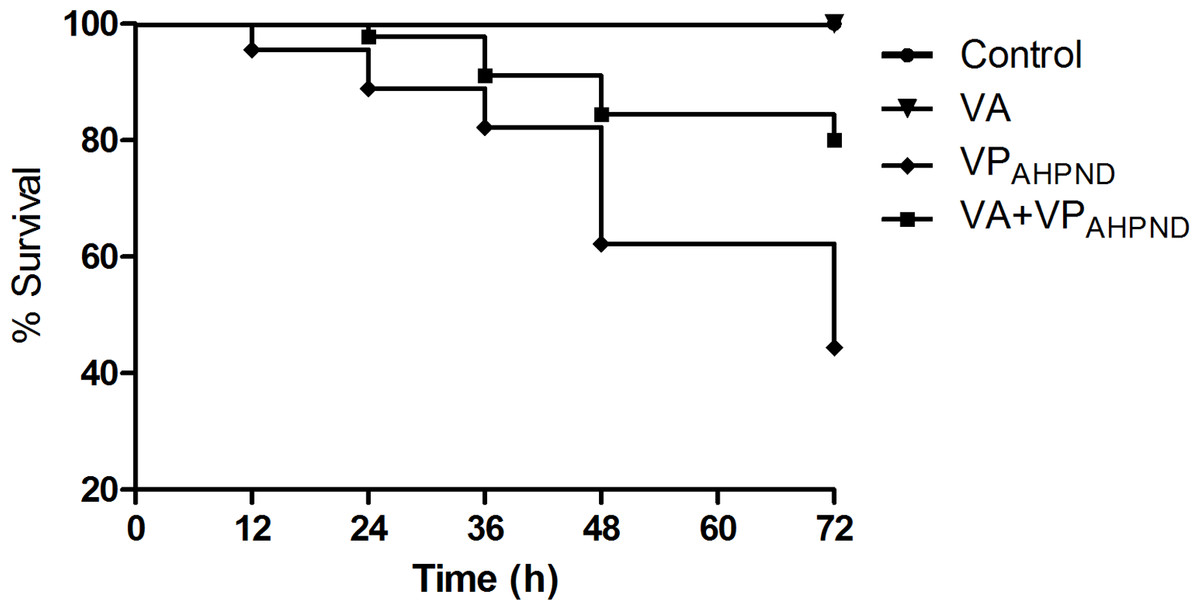
Figure 8: Kaplan–Meier survival of Litopenaeus vannamei shrimp infection assay.
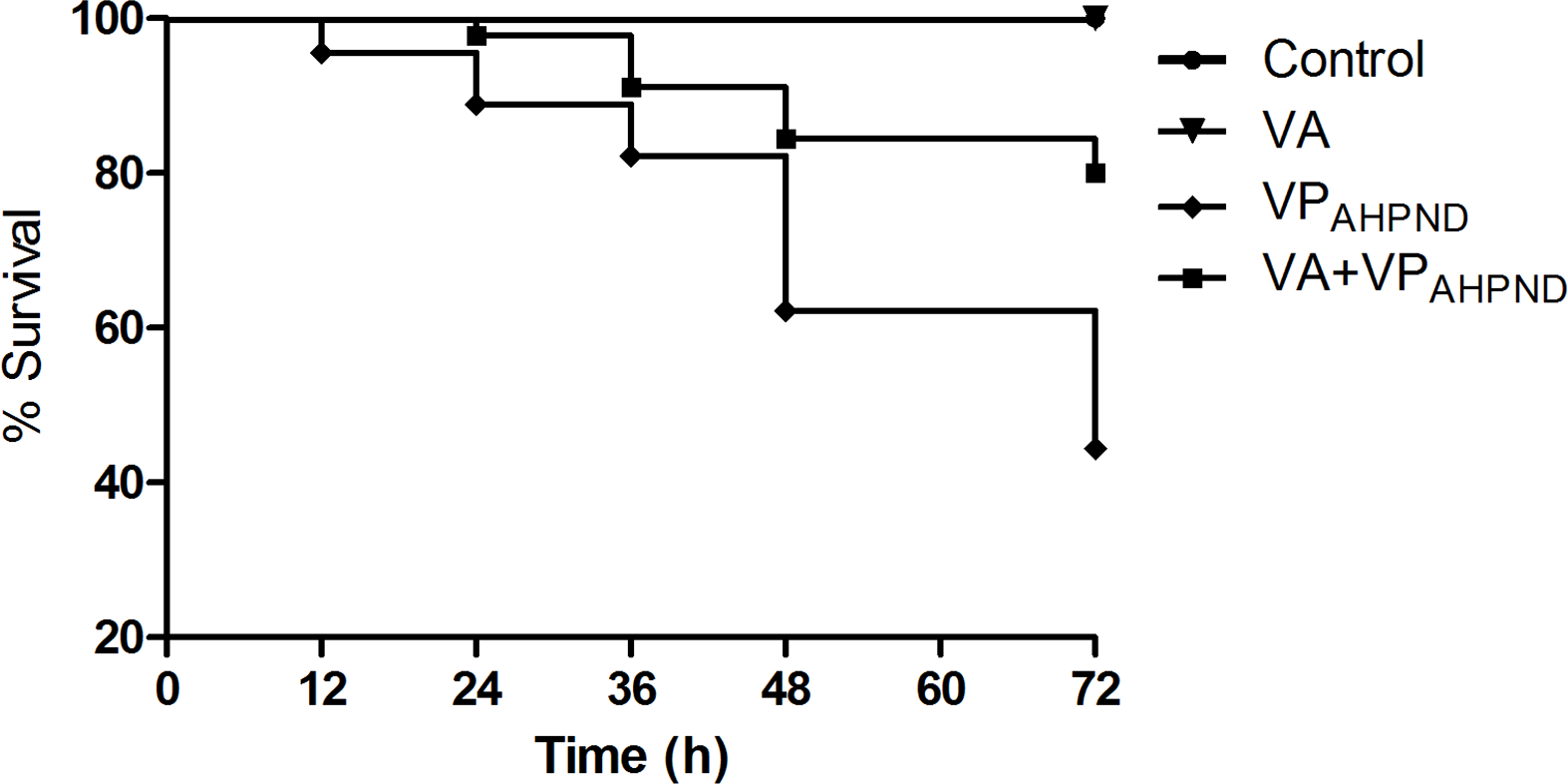{kind=link}
Discussion
Many antibiotics-resistant bacteria have occurred worldwide due to long-term use of antibiotics. There is an urgent need to develop alternative bacterial control strategies. QS inhibition has been proposed as a new strategy to control bacterial infection by interfering the expression of virulence factors and marine organisms have been proposed as an important source for the discovery of novel anti-QS compounds (Dobretsov, Dahms & Qian, 2006). Thus, this study evaluates the anti-QS activity of V. alginolyticus BC25 on reporter bacteria and investigates its effect on phenotypes of VPAHPND PSU5591.
C. violaceum was used for the preliminary screening of marine bacterium with anti-QS activity. This bacteria produce the purple pigment violacein except its QS system is obstructed (Fitriani, Ayuningtyas & Kusnadi, 2016). QS system of C. violaceum consists of two regulatory protein CviI and CviR that are homologus to LuxI and LuxR systems, respectively. The violacein production is regulated by vio operon (vioA, vioB, vioC, vioD, and vioE) through a QS system mediated by AHL (August et al., 2000). Findings from the present study revealed that CFCS of V. alginolyticus (6.25 to 50%) could have influenced QS system of C. violaceum. About 79.7% inhibition in the violacein production was detected with 50% (v/v) CFCS and without affecting to cell growth. It is possible that the addition of V. alginolyticus CFCS affected the transcription of vio gene (encoding violacein production). Similar research revealed that the cell pellet and supernatant extract of V. alginolyticus strain M3-10 at concentration 0.8 mg/mL reduced the violacein production without affecting the cell growth of C. violaceum ATCC 12472 and was therefore selected for QS inhibition assay against Pseudomonas aeruginosa (Reina et al., 2019).
In this study, global RNA-seq was used to explore the possible effects of V. alginolyticus extract on VPAHPND. The RNA-seq data showed the differential expression of 524 genes, of which 259 were up-regulated and 265 were down-regulated. Most of these genes are involved in quorum sensing, motility, virulence, and transport. Flagella-based motility is an important mode of locomotion of bacteria. In V. parahaemolyticus, it has two kinds of flagella include polar flagella (fla gene system) and lateral flagella (laf gene system) (Kojima et al., 2011). Polar flagellum is responsible for bacterial swimming in liquid, while lateral flagellum is required for bacterial swarming in solid surface (McCarter, 2004; Jaques & McCarter, 2006). In the present study, we examined the effect of CFCS of V. alginolyticus on swimming and swarming of VPAHPND by the spot method. However, swarming zone of VPAHPNDremains unclear (data not shown). For this reason, the streak method used for the swarming test. We demonstrated that at 40 to 80% CFCS of V. alginolyticus decreased the swimming motility and also influenced the swarming motility of VPAHPND in the plate assay. As expected, a majority of flagellar genes encoded for the flagellar structure and flagellin were down-regulated in the presence of V. alginolyticus extract. Thus, it is possible that the decreased swimming of VPAHPND is caused by the regulation of PomA and flaABCFGI genes, which control the function of polar flagellar. Also, flgABCDEFGKLMN, fliADMS and flhFG genes are associated with the function of lateral flagellar.
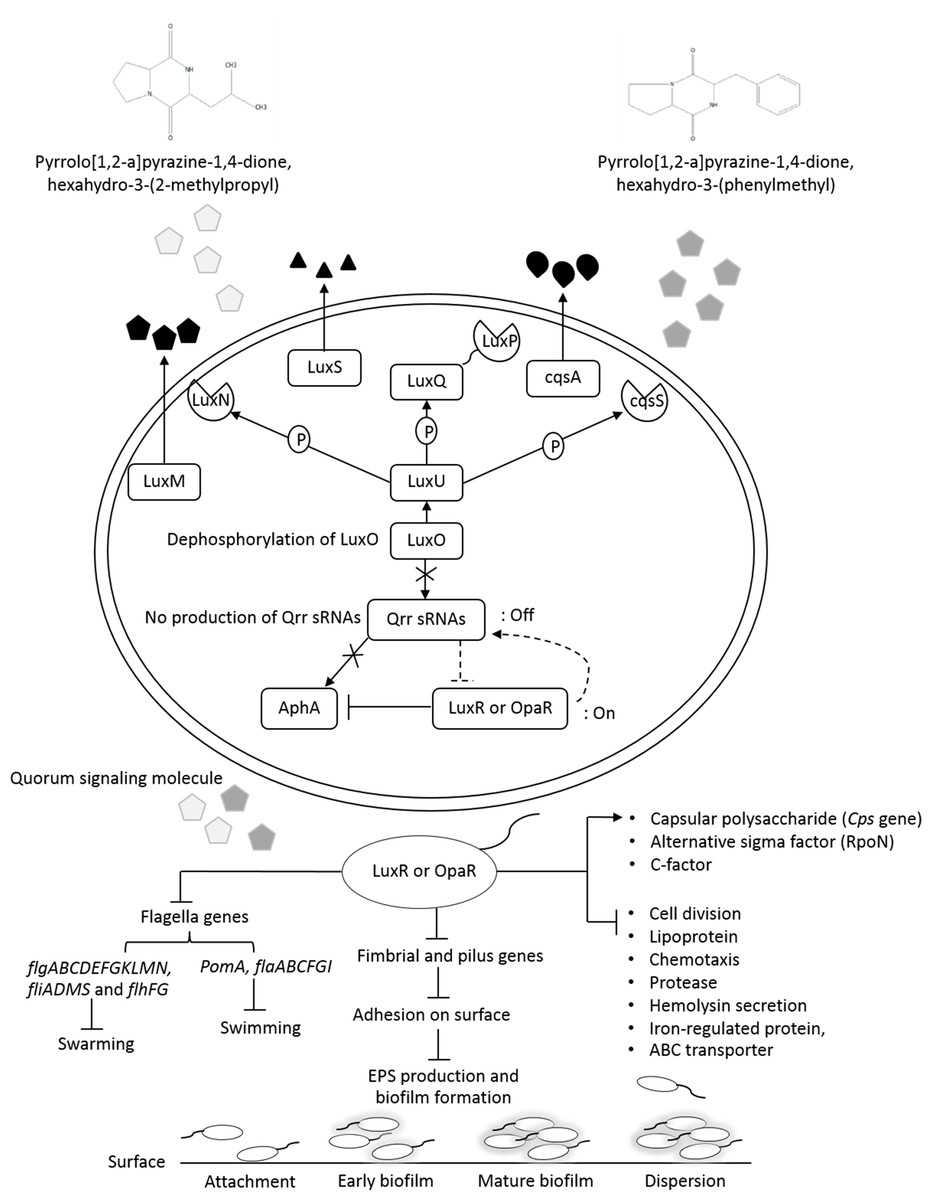
Figure 9: The transcriptional regulation model of QSI in VPAHPND PSU5591.
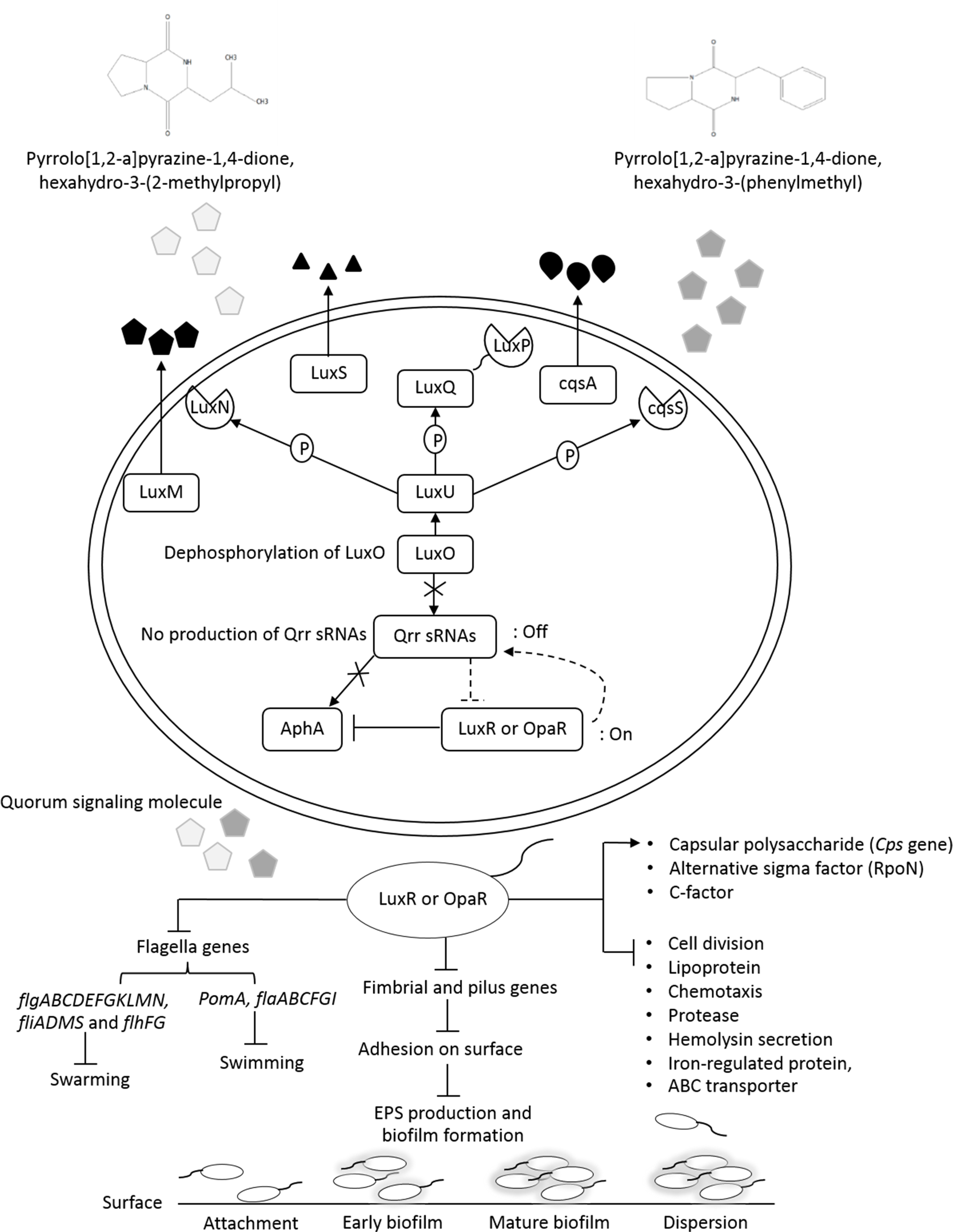{kind=link}
Additionally, about 36.6% reduction of biofilm formation was reported with 2.5% (v/v) of V. alginolyticus extract without significant effect on cell growth. We speculated that the reduction in biofilm formation by V. alginolyticus extract was associated with the loss of its ability to motility, attach to surfaces, and subsequently to form a biofilm of VPAHPND. For better understanding, the effect of V. alginolyticus extract (2.5%, v/v) was examined using SEM. The structural change in the VPAHPNDbiofilm was observed after 24 and 48 h. From the transcriptome data, we found that the genes encode fimbrial and pilus protein were down-regulated. Also, hypothetical proteins were differentially expressed by directly or indirectly. It might be play the role of biofilm formation. In the present study, we demonstrated the hypothetical model for explaining transcriptional regulation of QS in VPAHPND by V. alginolyticus extract based on the differential gene expression with available literature (Fig. 9). In general, V. parahaemolyticus produces harveyi autoinducer 1 (HAI-1) produced by LuxM and recognized by histidine kinase LuxN, autoinducer 2 (AI-2) is synthesized by LuxS and detected by histidine kinase complex LuxPQ, and cholerae autoinducer 1 (CAI-1) which is synthesized by CqsA and detected by histidine kinase CqsS. When the levels of autoinducers are high and bound to receptors lead to dephosphorylation and inactivation of the LuxO repressor. The Qrr genes are not transcribed, resulting in an expressed LuxR or OpaR regulator, whereas in turn repressed the transcription of regulator AphA (Gode-Potratz & McCarter, 2011; Zhang et al., 2012). Interestingly, transcriptome analysis revealed that the expression of LuxR transcriptional regulator or OpaR homolog (TetR family transcriptional regulator), sensor histidine kinase, sigma-54 interacting response regulator (alternative sigma factor, RpoN), C-factor and capsular polysaccharide were found to be up-regulated. Generally, QS system can be blocked by stopping of signal generator (LuxI homologue), destroying the signal molecule (autoinducer), preventing or competition signal molecule from binding with to signal receptor (LuxR homologue), and interruption with folding of some protein associated with QS such as transcriptional regulator (Rasmussen and Givskov, 2006). Hence, it perhaps suggests that the anti-QS mechanism of a marine bacterium, V. alginolyticus extract could interrupt with histidine kinase receptor and/or folding protein of LuxR and RpoN transcriptional regulator involve QS-controlled phenotypes including swimming, biofilm formation, and capsular polysaccharide. However, there was no evidence on how V. alginolyticus extract and LuxR transcriptional regulator interact in controlling VPAHPND. Moreover, genes encoding for cell division, lipoprotein, chemotaxis, biotin synthase, protease, hemolysin secretion, phage shock protein, iron-regulated protein, bacterioferritin and ABC transporter were down-regulated in the presence of V. alginolyticus extract. A previously study reported that motility, biofilm formation, type III and VI secretion system production (T3SS, T6SS), and capsule production of V. parahaemolyticus are directly controlled by the QS regulator OpaR (Gode-Potratz & McCarter, 2011; Kernell Burke et al., 2015; Salomon, Klimko & Orth, 2014; Wang et al., 2013; Wu et al., 2019; Zhang et al., 2012). OpaR has been known as a regulator for the expression of various virulence factors and represses cytotoxicity toward host cells (Gode-Potratz & McCarter, 2011). Studies have demonstrated that rpoN mutant leads to loss of motility and also reduced biofilm formation in V. parahaemolyticus, V. cholerae, V. fischeri, V. anguillarum, and P. aeruginosa (Klose & Mekalanos, 1998; O’Toole et al., 1997; Whitaker, Richards & Boyd, 2013; Wolfe et al., 2004). Moreover, the deletion of aphA leads to reduce hemolytic and cytotoxic activity in V. parahaemolyticus (Wang et al., 2013). Also, the results of this study showed that hemolysin secretion which is an important virulence factor required for pathogenesis in V. parahaemolyticus was down-regulated in the presence of V. alginolyticus extract. C-factor is an intercellular signaling protein, which is required for aggregation, sporulation, and associated with the cell surface and biofilm formation (Andersen & Kaiser, 1996). Similar research revealed that the production of capsular polysaccharide was conversely related to biofilm in Pasteurella multocida. It is possible that a negatively charged capsule may interrupt the biofilm formation by preventing the EPS matrix from encasing bacterial cell or blocking adherence to surface (Petruzzi et al., 2017). The major compound presented in V. alginolyticus extract was demonstrated some resemblance with Cyclo-(L-Leu-L-Pro) and Cyclo-(L-Phe-L-Pro). Among the studies of marine isolates, the antibiotic agent pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro from Bacillus tequilensis strain MSI45 recovered from a marine sponge, Callyspongia diffusa could effectively inhibited the multi-drug resistant Staphylococcus aureus and showed high antioxidant activity (Kiran et al., 2018). Moreover, pyrrolo [1,2-a]pyrazine-1,4-dione, hexahydro- from Streptomyces mangrovisoli isolated from mangrove soil in the state of Pahang, Peninsular Malaysia exhibited strong antioxidant activity (Ser et al., 2015). In addition, Cyclo-(L-Phe-L-Pro) or pyrrolo [1,2-a]pyrazine-1,4-dione, hexahydro-3-(phenylmethyl) from Streptomyces sp. strain VITPK9 isolated from the salt spring habitat of Manipur, India was suggested as a potential anticandidal compound (Sanjenbam & Kannabiran, 2016). Findings from the present study revealed that four extract fractions of V. alginolyticus (no. 1, 4, 5, and 7) could have influenced QS system of C. violaceum at concentrations 1.6, 0.6, 0.4, and 0.8 mg/mL, respectively (Fig. S1). Similar research reported that 0.039 mg/mL (160 µM) Cyclo-(L-Phe-L-Pro) and 0.023 mg/mL (94 µM) Cyclo-(L-isoLeu-L-Pro) produced by Marinobacter sp. SK-3 inhibited QS-dependent production of violacein by C. violaceum CV017 and also reduced luminescence production of reporter E. coli pSB401 (Abed et al., 2013). Moreover, QS inhibition were achieved with 0.250 mg/mL of Cyclo-(L-Leu-L-Pro), secreted by Staphylococcus saprophyticus which isolated from marine sponges, and based on the inhibition of violacein production the inhibition of QS-controlled violacein production was observed with 0.250 mg/mL of Cyclo-(L-Leu-L-Pro), which secreted by Staphylococcus saprophyticus (Li et al., 2013). Hence, it perhaps suggests that Cyclo-(L-Leu-L-Pro) and Cyclo-(L-Phe-L-Pro) can be blocked using interference or competitive QS inhibition that outcompete autoinducer of VPAHPND for receptors resulting in LuxR or OpaR transcriptional regulator turns on which could modulate the expression of QS-controlled phenotypes including swimming, biofilm formation, EPSs production, and capsular polysaccharide. Bacteria in the genus Vibrio have been successfully used as probiotic in shrimp aquaculture (Garriques & Arevalo, 1995; Griffith, 1995). This genus is found as gut microbiota of shrimp which are associated to compete with pathogenic bacteria and to produce essential elements for host metabolism (Tzuc et al., 2014). In this study, treatment with V. alginolyticus resulted in 80% shrimp larvae survival, whereas shrimp larvae challenged with VPAHPND alone reached 55.56% mortality. Previously, a study reported that the probiotic V. alginolyticus strain Ili caused 20% increase in shrimp larvae survival and shrimp yield increased significantly 208 kg/ha (Rodriguez et al., 2011). Moreover, in other vibrios tested, the probiotic V. gazogenes has potential for the control of pathogenic bacteria in the Pacific white shrimp L. vannamei (Thompson et al., 2010). Additionally, in our experiments previously, isolation and characterization of V. alginolyticus were investigated. V. alginolyticus is isolated from seafood samples, Songkhla province, Thailand. The results revealed that 35 isolates of V. alginolyticus out of 50 isolates were identified and tested against VPAHPND by cross-streak method. A total of 5 from 35 V. alginolyticus isolates showed antagonistic activity to VPAHPND especially strain PSU5591. Interestingly, V. alginolyticus strain BC25 was high potential isolate. Therefore, we focus on V. alginolyticus BC25 against VPAHPND PSU5591 for further experiments. The data presented in “The 4th National Conference on Science and Technology of Southern Network 2019 (NSCIC 2019), Songkhla Rajabhat University”. This study demonstrates the possibility of using V. alginolyticus as a potential source for the production of anti-QS compounds or used as probiotics in aquaculture.
Conclusions
A marine V. alginolyticus BC25 exhibited anti-QS and anti-biofilm activity of VPAHPND PSU5591. Based on the results of RNA-seq, bacterial extract interfered with the expression of flagella genes involved in biofilm formation and iron-regulated virulence regulatory gene of VPAHPND. In addition, LuxR family transcriptional regulator gene, c factor cell–cell signaling gene, and capsular polysaccharide were up-regulated. The compounds Cyclo-(L-Leu-L-Pro) and Cyclo-(L-Phe-L-Pro) were identified as a potential anti-QS compound. Also, V. alginolyticus has potential for the use as a probiotic in shrimp against VPAHPND. This study represents the attractive target for alternative strategies to control VPAHPND in aquaculture.
Supplemental Information
Analysis of active fractions of Vibrio alginolyticus BC25 showing anti-QS activity
HPLC analysis of Vibrio alginolyticus BC25 extract. The chromatogram shows the seven main active peaks. Analysis was performed on a reverse-phase C18 column (50 × 2.1 mm, Waters, CA, USA). The mobile phase was methanol and water (20:80, v/v) at 30° C at a flow rate of 0.2 mL/min. The insert picture in (Fig. S1) is the re-tested of the seven peak compounds at concentrations ranging from 0.4−1.6 mg/mL against violacein production in C. violaceum DMST46846 by drop plate method.
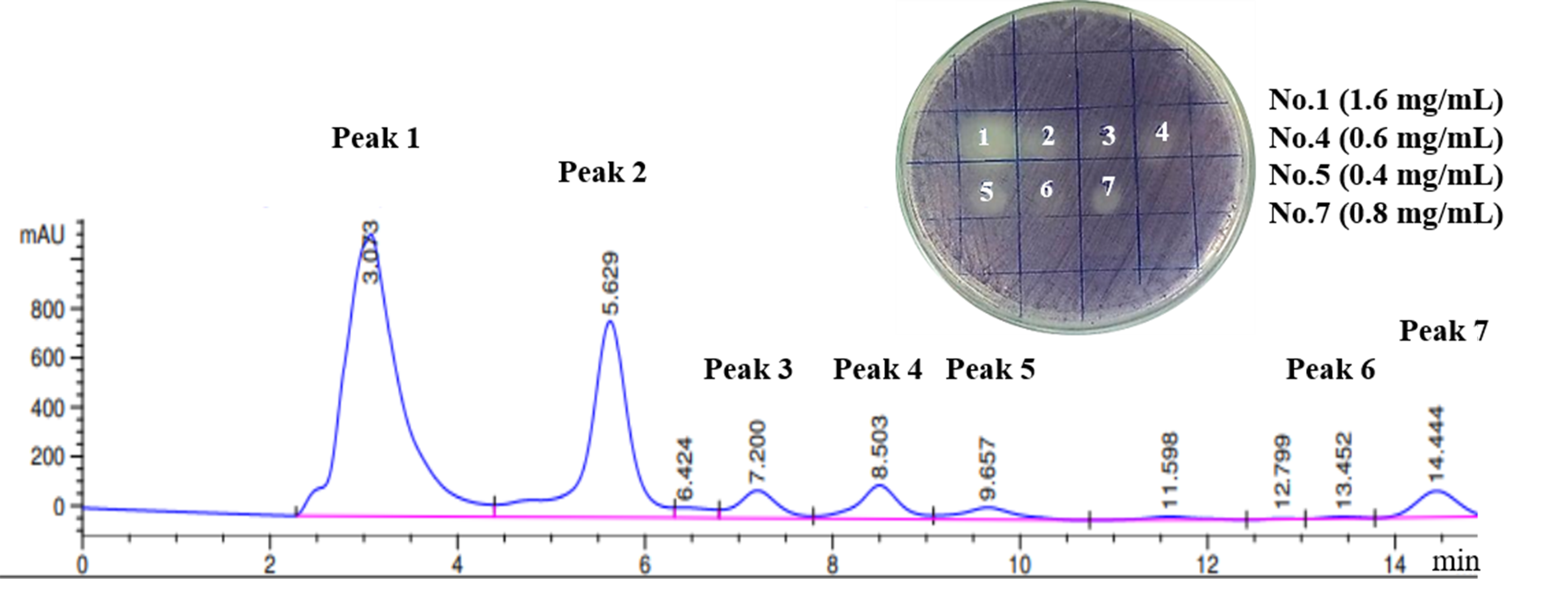{kind=link}
Representative NMR spectroscopy analysis of Vibrio alginolyticus BC25 extract
NMR spectrum analysis of active compound (Fraction 1). (A) 1H NMR spectrum at 500 MHz (B) 13C NMR spectrum at 125 MHz (C) 2D-HMBC spectrum (D) 2D-HSQC spectrum.
{kind=link}