Exploring the transcriptome of luxI- and ΔainS mutants and the impact of N-3-oxo-hexanoyl-L- and N-3-hydroxy-decanoyl-L-homoserine lactones on biofilm formation in Aliivibrio salmonicida
- Published
- Accepted
- Subject Areas
- Microbiology, Molecular Biology
- Keywords
- Quorum sensing, Acyl homoserine lactone, Aliivibrio salmonicida, Biofilm, Exopolysccharides
- Copyright
- © 2018 Khider et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ Preprints) and either DOI or URL of the article must be cited.
- Cite this article
- 2018. Exploring the transcriptome of luxI- and ΔainS mutants and the impact of N-3-oxo-hexanoyl-L- and N-3-hydroxy-decanoyl-L-homoserine lactones on biofilm formation in Aliivibrio salmonicida. PeerJ Preprints 6:e27443v1 https://doi.org/10.7287/peerj.preprints.27443v1
Abstract
Background. The marine bacterium A. salmonicida uses the quorum sensing (QS) systems, AinS/R and LuxI/R to produce eight acyl-homoserine lactones (AHLs) in a cell density dependent manner. Biofilm formation is one of the QS regulated phenotypes, which requires the expression of exopolysaccharides (EPS).We previously demonstrated that inactivation of LitR, the master regulator of QS in A. salmonicida resulted in biofilm formation, which was, similar to the biofilm formed by the AHL deficient mutant ΔainSluxI-.In this work, we have identified genes regulated by AinS and LuxI using RNA sequensing (RNA-Seq), and studied their role in biofilm formation, colony morphology and motility. We have also studied the effect of two AHLs on the biofilm formation.
Results.The transcriptome profiling of ΔainS and luxI- mutants allowed us to identify essential genes regulated by QS in A. salmonicida. Relative to the wild-type, the ΔainS and luxI- mutants revealed 40 and 500 differentially expressed genes (DEGs), respectively. The functional analysis demonstrated that the most pronounced DEGs were involved in bacterial motility and chemotaxis, exopolysaccharide production, and surface structures related to adhesion. Inactivation of luxI but not ainS genes resulted in wrinkled colony morphology. While inactivation of both genes (ΔainSluxI-) resulted in strains able to form wrinkled colonies and mushroom structured biofilm. Moreover, when the ΔainSluxI- mutant was supplemented with N-3-oxo-hexanoyl-L- homoserine lactone (3OC6-HSL) and N-3-hydroxy-decanoyl-L-homoserine lactone(3OHC10-HSL), the biofilm did not develop. We also show that LuxI is needed for motility and repression for EPS production, where repression of EPS is likely operated through the RpoQ-sigma factor.
Conclusion.These findings imply that LuxI and AinS synthases have a critical contribution to the QS-dependent regulation on gene expression and the phenotypic traits related to it.
Author Comment
This is a submission to PeerJ for review.
We would like to thank Augusta Hlin Aspar Sundbø from advanced microscopy core facility (AMCF) at UiT- the Arctic University of Tromsø for helping us with Scanning electron microscopy. We also thank Dr. Eric V. Stabb (University of Georgia) for the pVSV102 and pEVS104 plasmids
Supplemental Information
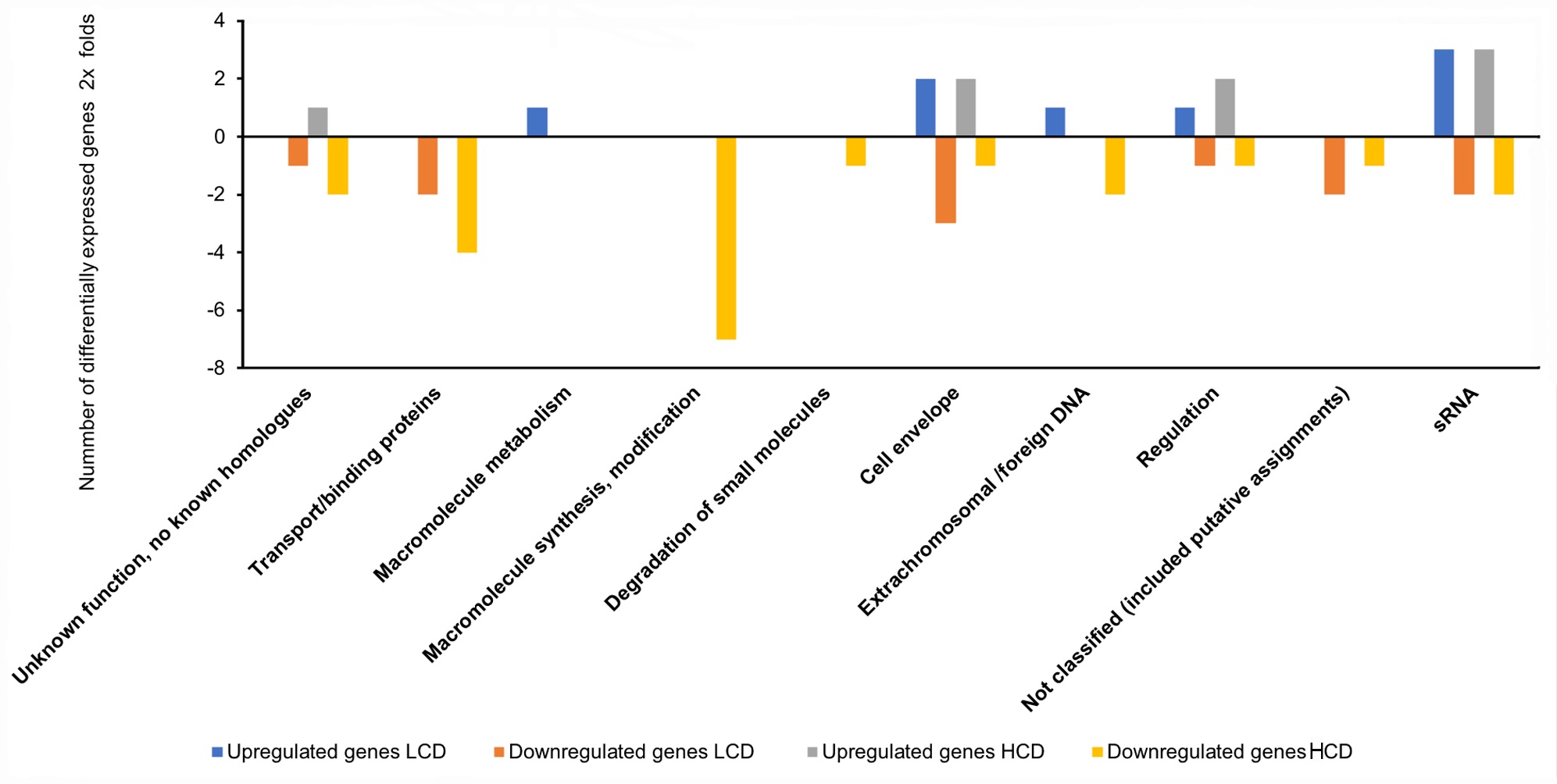
Functional distribution of genes between A. salmonicida wild-type and ΔainS mutant at HCD compared to LCD that are ≥2 × differentially expressed
{kind=link}
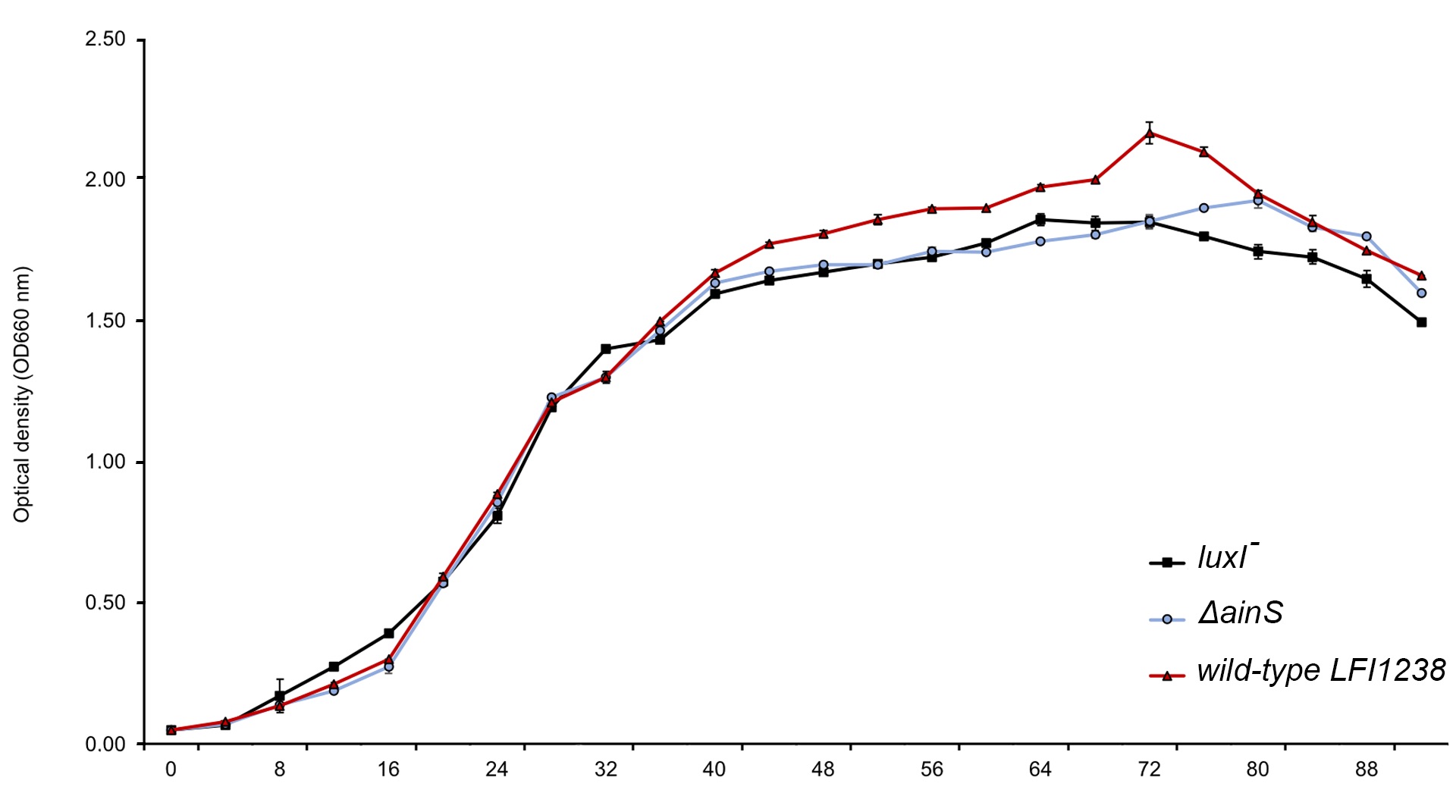
Growth curve of LFI1238, luxI-,ΔainS and ΔainSluxI-
The overnight secondary cultures were diluted to a starting OD600 of 0.05 in a total volume of 60 ml SWT. The cultures were grown further in 250 ml baffled flask at 8°C and 220 rpm. The optical density was measured every 4 hours using Ultrospec 10 cell density meter (Amersham Biosciences). The error bars represent the standard deviation of biological triplicates.
{kind=link}
Genes that are ≥2× or ≤ −2× differentially expressed in A. salmonicida luxI- mutant compared to wild-type at low cell density
Genes that are ≥2× or ≤ −2× differentially expressed in A. salmonicida luxI- mutant compared to wild-type at high cell density
Genes that are ≥2× or ≤ −2× differentially expressed in A. salmonicida ΔainS mutant compared to wild-type at low cell density
Genes that are ≥2× or ≤ −2× differentially expressed in A. salmonicida ΔainS mutant compared to wild-type at high cell density
Motility zones of LFI1238, luxI-, ΔainS and ΔainSluxI-, formed on soft agar plates
Each value represents the average (mm) of biological triplicates ± standard deviation.