Next-generation sequencing reveals cryptic Symbiodinium diversity within Orbicella faveolata and Orbicella franksi at the Flower Garden Banks, Gulf of Mexico
- Published
- Accepted
- Subject Areas
- Biodiversity, Genetics, Marine Biology
- Keywords
- next-generation sequencing (NGS), Flower Garden Banks, Symbiodinium, Caribbean, Orbicella faveolata, Orbicella franksi, amplicon sequencing, ITS-2, OTU
- Copyright
- © 2014 Green et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ PrePrints) and either DOI or URL of the article must be cited.
- Cite this article
- 2014. Next-generation sequencing reveals cryptic Symbiodinium diversity within Orbicella faveolata and Orbicella franksi at the Flower Garden Banks, Gulf of Mexico. PeerJ PrePrints 2:e246v1 https://doi.org/10.7287/peerj.preprints.246v1
Abstract
The genetic composition of the resident Symbiodinium endosymbionts appears to strongly modulate the physiological performance of reef-building corals. Here, we used deep amplicon sequencing to quantitatively assess Symbiodinium genetic diversity for the two mountainous star corals, Orbicella franksi and Orbicella faveolata, from two reefs separated by 19 kilometers of deep water. We aimed to determine if symbiont diversity is largely partitioned with respect to coral host species or geographic location. Our results demonstrate that across the two reefs both coral species contained only Symbiodinium identifiable as clade B type B1, represented by five distinct haplotypes. Three of these haplotypes have not been previously described and may be endemic to the Flower Garden Banks. No consistent differences in symbiont composition were detected between the two coral species. However, significant quantitative differences were observed between the east and west banks for two of the five haplotypes. These results highlight the need for consistent molecular genotyping techniques to assess local community assemblages of Symbiodinium-host relationships, which could be largely irrespective of host genetic background. This deep-sequencing approach used to sensitively characterize cryptic genetic diversity of Symbiodinium will potentially contribute to the understanding of physiological variations among coral populations.
Supplemental Information
Table 1: Analysis of Molecular Variance (AMOVA) Fixation index (FST) values.
Analysis of Molecular Variance (AMOVA) Fixation index (FST) values showing no genetic differentiation among Orbicella faveolata and Orbicella franksi, among Orbicella faveolata within the two geographic locations or among Orbicella franksi within the two geographic locations.
Table 2: The sequencing coverage and mapping efficiency by geographic location and species.
Summary of sequence coverage ITS-2 amplicon sequencing of Flower Garden Banks, Gulf of Mexico. Individuals are sorted by geographic location and species using the 454 GS FLX platform.
Table 3: The significant Markov Chain Monte Carlo Generalized Linear Model results.
Only showing significant results from Poisson-lognormal Generalized Linear Models (GLMs). Haplotypes IV and V are significantly diminished at the west bank compared to the east bank (PMCMC<0.001). Haplotype V is also significantly more diminished in Orbicella faveolata than in Orbicella franksi (PMCMC=0.002).
Figure 1: The general location of the Flower Garden Banks and pictures of coral species.
A. Location of Flower Garden Banks National Marine Sanctuary, Gulf of Mexico (27°54’ N, 93°35’W for east Flower Garden Banks and 27°53’N, 93°49’W west Flower Garden Banks) Credit: USGS (http://pubs.usgs.gov/of/2003/of03-002/html/FGB_figs.htm) B. Orbicella faveolata from Panama, Credit: Mónica Medina C. Orbicella franksi from Panama, Credit: Mónica Medina
{kind=link}
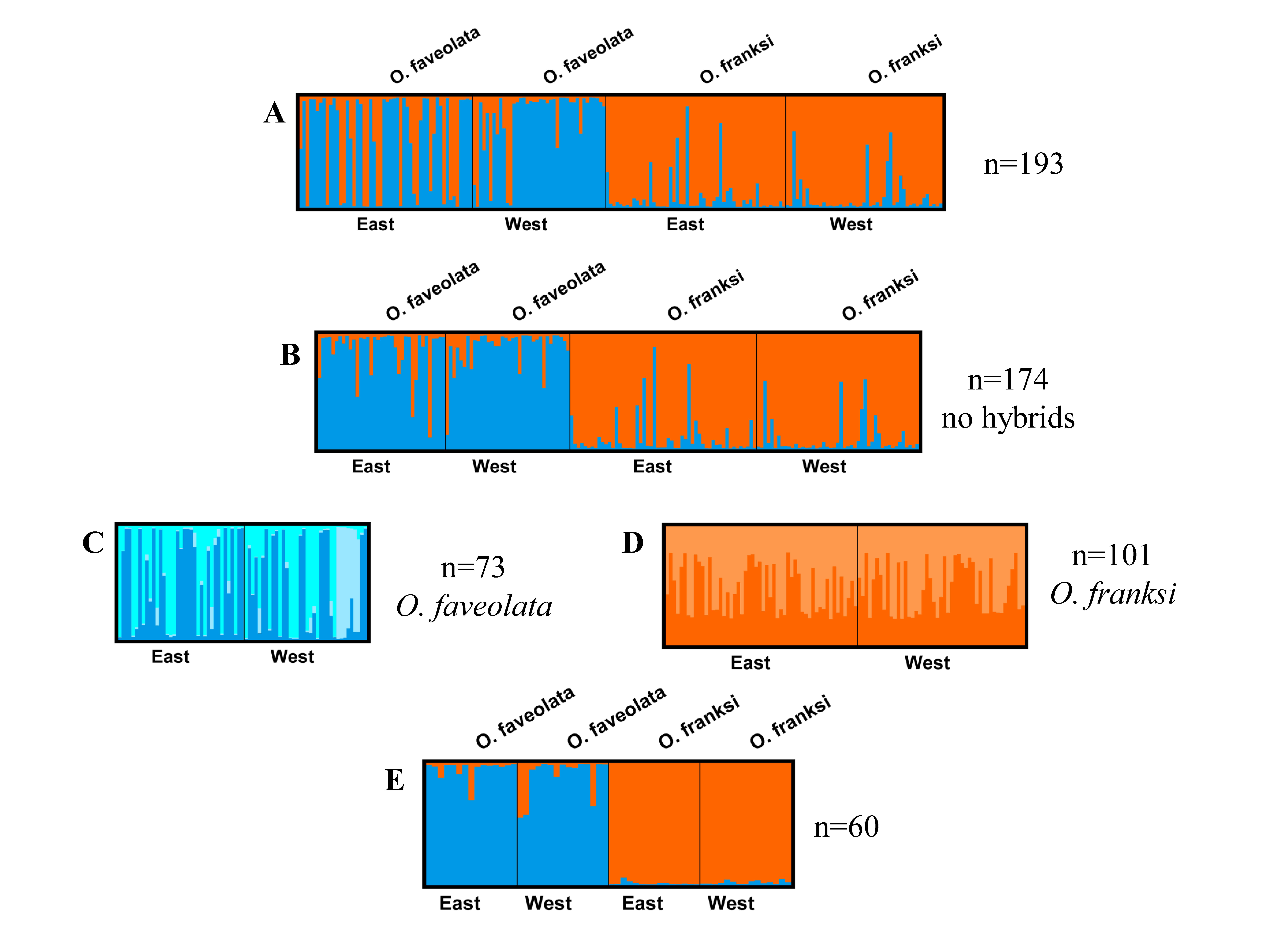
Figure 2: DISTRUCT plots of all STRUCTURE analyses.
DISTRUCT plots from STRUCTURE for K=2 except where noted A. All samples from Orbicella faveolata and Orbicella franksi in east and west Flower Garden Banks National Marine Sanctuary, Gulf of Mexico, n=193 B. Same as A but potential hybrids removed (n=174) C. Orbicella faveolata only with potential hybrids removed (n=73, K=3) and D. Orbicella franksi only with potential hybrids removed (n=101) E. The selected 60 Orbicella faveolata (n=30) and Orbicella franksi (n=30)
{kind=link}
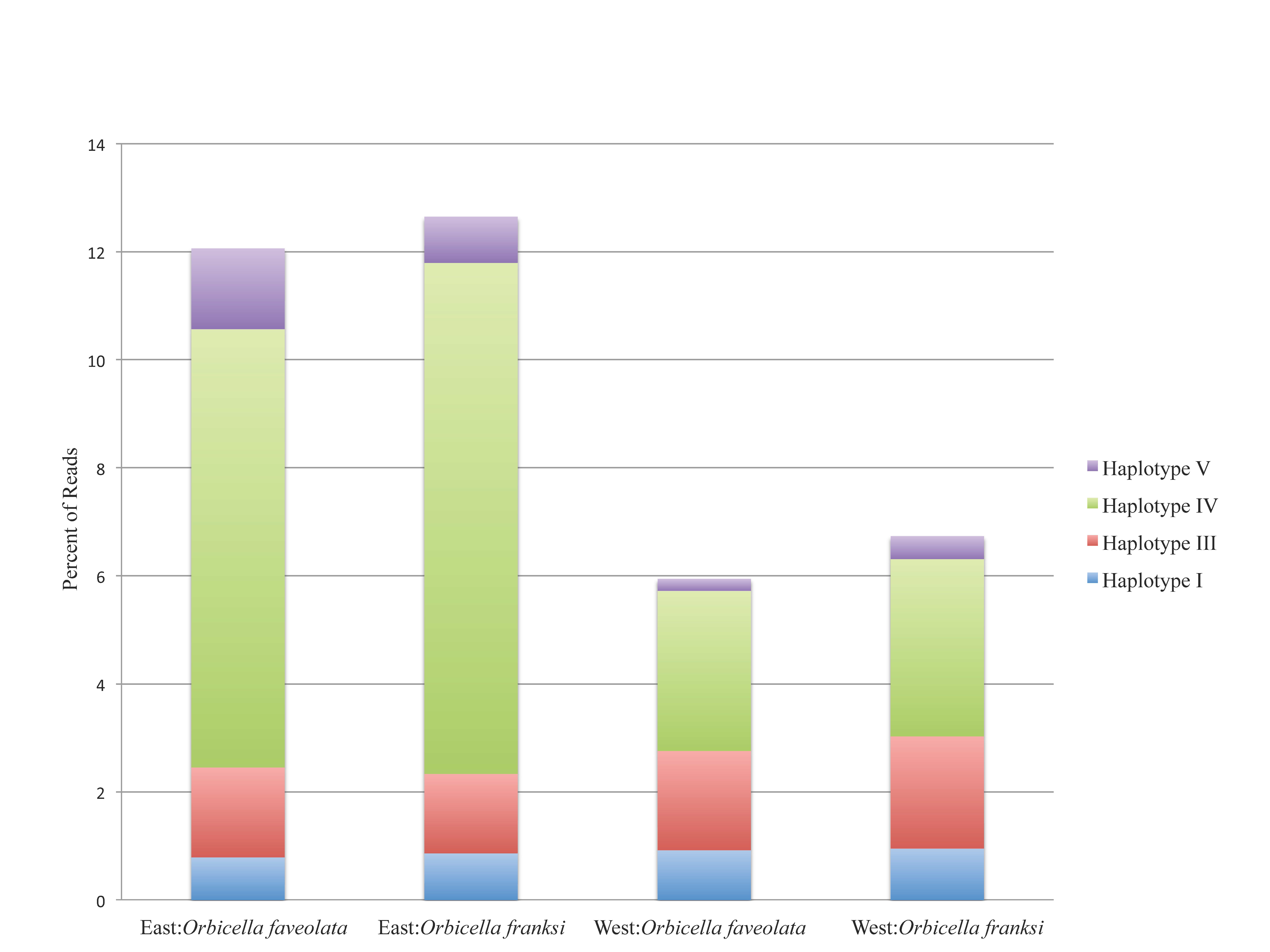
Figure 3: The percentage of reads for the four minor haplotypes by geographic location and species.
Percentage of minor Symbiodinium B1 haplotypes by geographic location and coral species. Only haplotypes I, III, IV, V are shown. The dominant Symbiodinium B1 haplotype II, used 93.26% across all individuals, is not shown. (East: Orbicella faveolata = 27,121 sequences, East: Orbicella franksi = 40,078 sequences, West: Orbicella faveolata = 26,143 sequences, West: Orbicella franksi = 27,376 sequences)
{kind=link}
Figure 4: Distribution of abundance of two haplotypes significant by site.
Distribution of abundance (log2 transformed) of Symbiodinium type B1 haplotypes IV and V in east and west Flower Garden Banks, Gulf of Mexico from the Poisson-lognormal model. Circles indicate Orbicella faveolata. Triangles indicate Orbicella franksi. Haplotypes I, II and III did not have significant effects, not shown.
{kind=link}
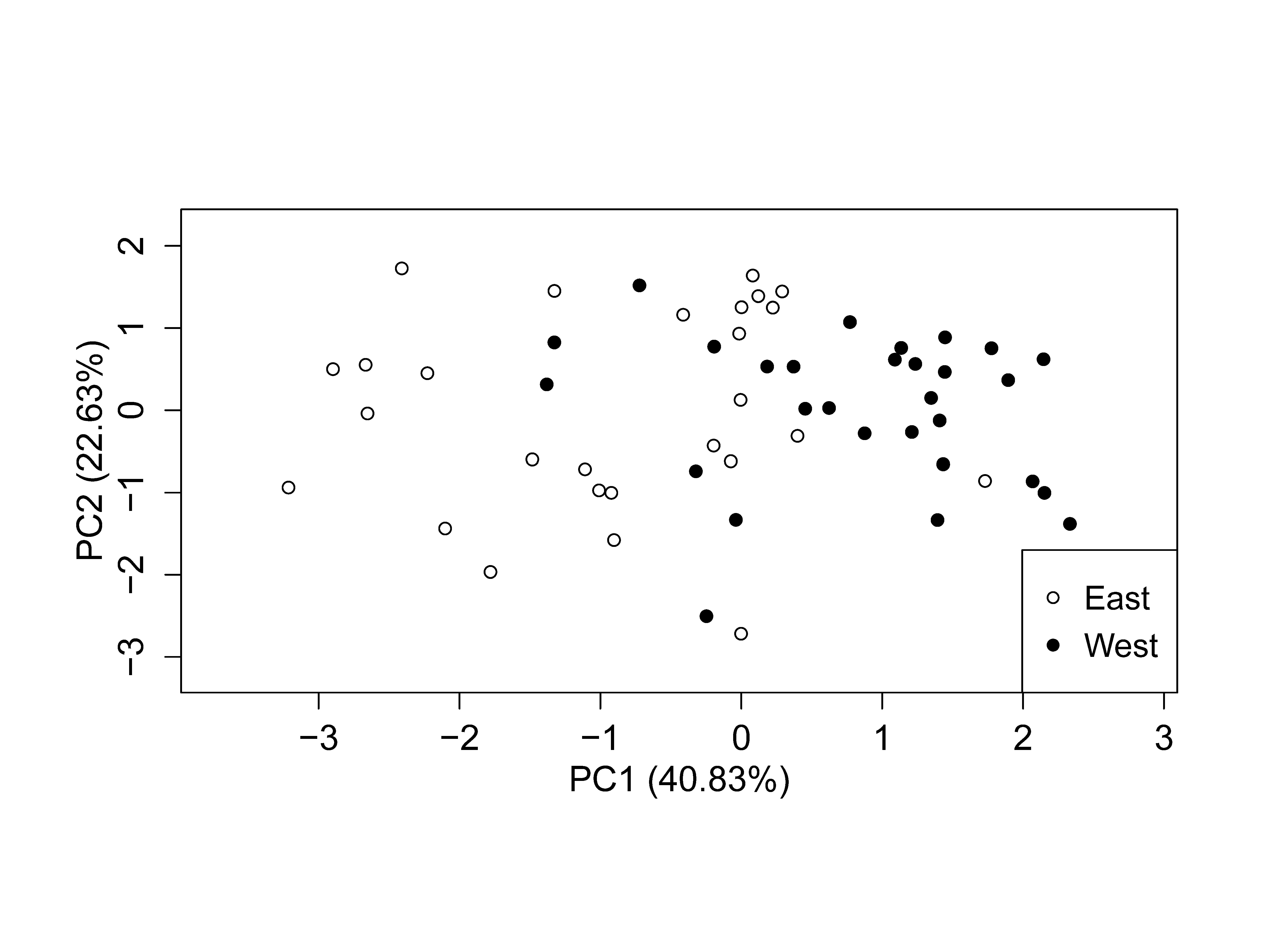
Figure 5: Principle Components Analysis (PCA) showing the first two principle components.
PCA plot of variance stabilized transformed (VST) data from a count data set showing partitioning of samples by geographic location. Principle component 1 (PC1) explains 40.83% of the variation and principle component 2 (PC2) explains 22.63% of the variation (n=56).
{kind=link}
Supplementary Data 1: The R script and input file used for MCMC GLMs and two Perl scripts used to trim and clean sequences in bioinformatics analysis.
Custom perl scripts used to remove 454 Rapid adaptors and barcodes (454trim.pl) and short reads (noshorts.pl) and R script (FGB_Symbiodinium_OTU.R).
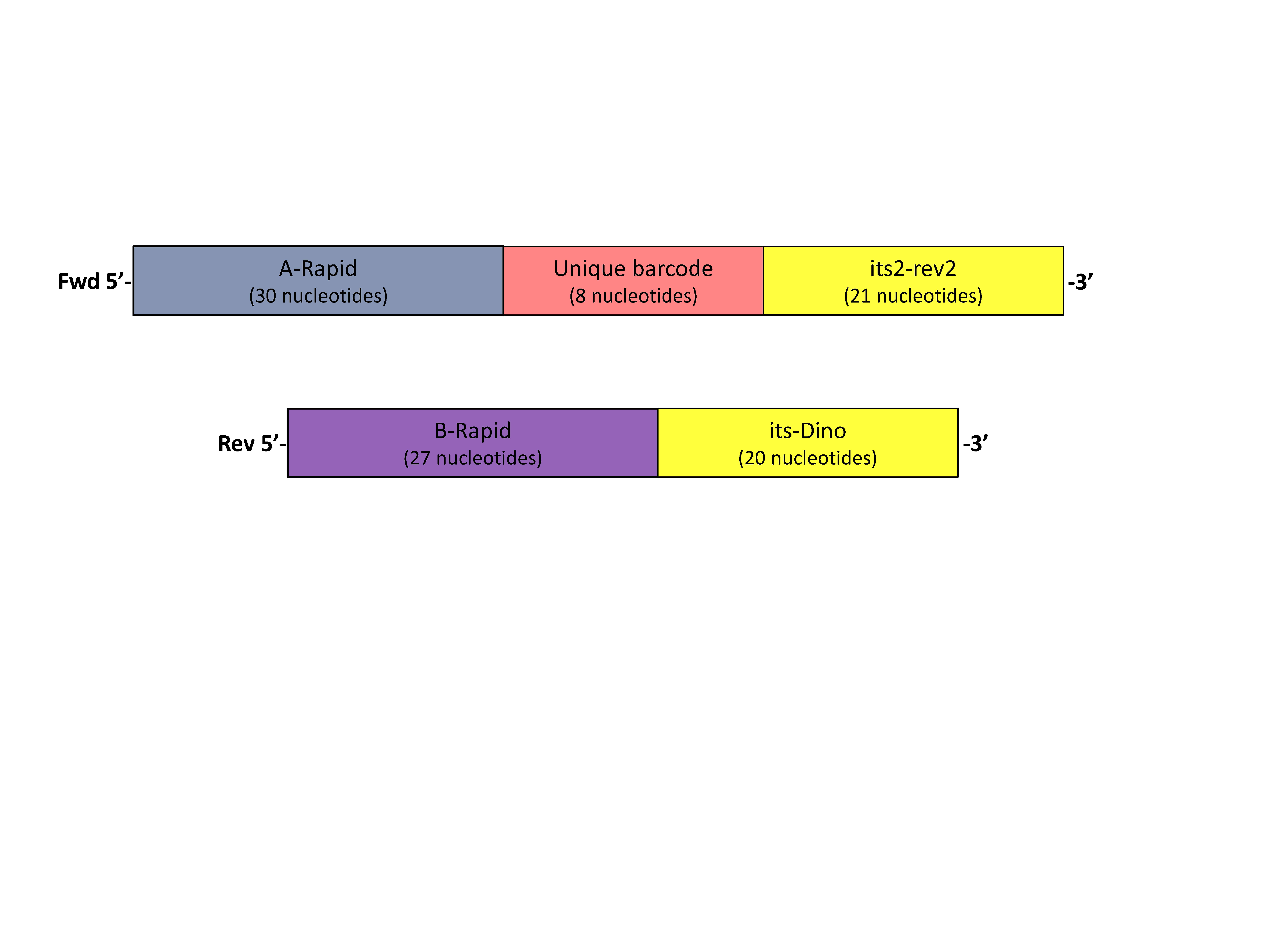
Supplementary Figure 1: The primer design to uniquely barcode individuals.
Rapid-barcode primer design annealed in second PCR to uniquely identify individuals and pool.
{kind=link}
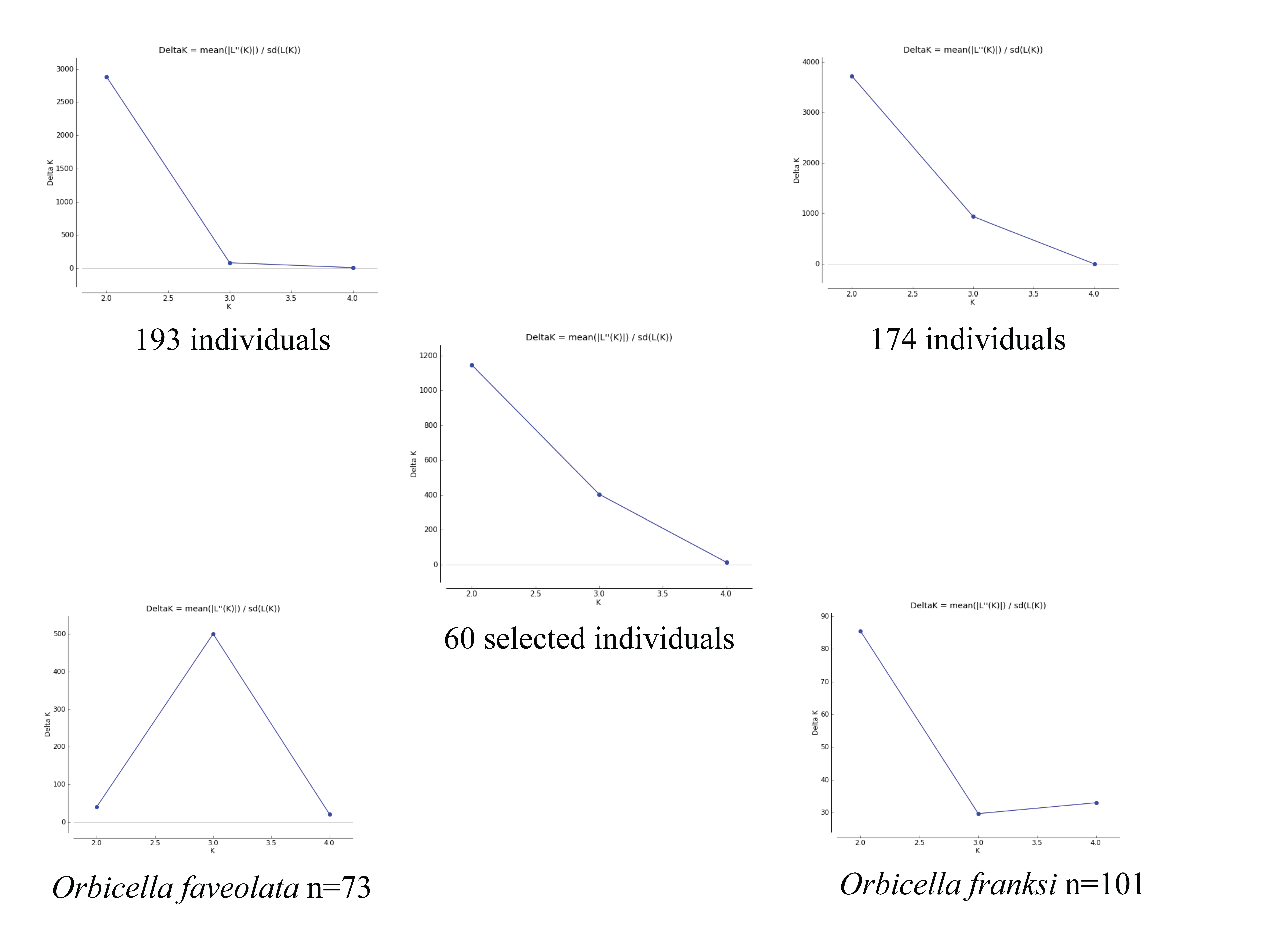
Supplementary Figure 2: The delta K figures from STRUCTURE HARVESTER.
Delta K figures from STRUCTURE HARVESTER from STRUCTURE analysis for all collected individuals (n=193), with potential hybrids removed (n=174), the selected 60 individuals, Orbicella faveolata (n=73) and Orbicella franksi (n=101).
{kind=link}
Supplementary Figure 3: The Clustal alignment of five Flower Garden Bank Symbiodinium haplotypes and previously published Symbiodinium B1.
Clustal Omega alignment of five reference haplotypes displayed in SeaView and the previously published Symbiodinium B1 (JN 558059.1).
{kind=link}