
Description of two new species of the genus Heterochelamon Türkay & Dai, 1997 (Crustacea: Decapoda: Brachyura: Potamidae), from southern China
- Published
- Accepted
- Received
- Academic Editor
- Mark Young
- Subject Areas
- Biodiversity, Biogeography, Taxonomy, Zoology
- Keywords
- Freshwater crab, Taxonomy, Heterochelamon, Phylogenetic
- Copyright
- © 2020 Wang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. Description of two new species of the genus Heterochelamon Türkay & Dai, 1997 (Crustacea: Decapoda: Brachyura: Potamidae), from southern China. PeerJ 8:e9565 https://doi.org/10.7717/peerj.9565
Abstract
This study describes two new species of freshwater crab of the genus Heterochelamon Türkay & Dai, 1997 from southern China, H. huidongense from Guangdong Province and H. jinxiuense from Guangxi Zhuang Autonomous Region. The two new species can be differentiated from congeners by characters derived from the shape of the epibranchial tooth, external orbital angle, cheliped proportions and structure of the male first gonopod. The present study brings the number of Heterochelamon species to seven. We used the mitochondrial 16S rRNA gene for a molecular analysis and the results are consistent with the morphological features that support the recognition of two new taxa.
Introduction
Freshwater crabs are found in the tropics and subtropics in most parts of the world, occurring in aquatic habitats ranging from clear montane streams to lowland rivers and even in caves or tree holes (Dai, 1999; Ng, Guinot & Davie, 2008). Southern China, where mountains and rivers are abundant, provides a multitude of habitats for a large number of species and the large numbers of new taxa reported in recent years indicate that research on this group remains in a “discovery” phase (Yeo et al., 2008; Zhu, Naruse & Zhou, 2010; Naruse, Zhu & Zhou, 2013; Huang, Mao & Huang, 2014; Shih & Do, 2014; Huang, Shih & Mao, 2016; Huang, Shih & Ng, 2017; Huang, Ahyong & Shih, 2017; Ng, 2017; Huang, 2018; Huang, Shih & Ahyong, 2018; Huang, Wong & Ahyong, 2018; Wang, Huang & Zou, 2019; Wang, Zhou & Zou, 2019).
Currently, the freshwater crabs distributed in China are represented by 49 genera in the families Potamidae Ortmann, 1896 and Gecarcinucidae Alcock, 1910 (Chu et al., 2018; Chu, Wang & Sun, 2018; Huang, Shih & Ahyong, 2018; Huang, Huang & Shen, 2020). The potamid genus Heterochelamon was established by Türkay & Dai (1997) to accommodate Potamon (Geothelphusa) purpureomanualis Wu, 1934 (type species), theretofore assigned to Malayapotamon, and two new species, H. guangxiense and H. yangshuoense. Naruse, Zhu & Zhou (2013) further described two new species, H. tessellatum and H. castanea. The five known species exclusively occur in Guangxi Zhuang Autonomous Region (Türkay & Dai, 1997; Naruse, Zhu & Zhou, 2013).
In 2011, we collected specimens of a Heterchelamon species from Jinxiu, Guangxi, but the crabs were not identified at the time. It was not until 2018 that our research team identified the specimens as belonging to an undescribed species. In 2019, freshwater crab enthusiast Jia-Ming Tian collected some dark colored freshwater crabs from Huidong, Guangdong. We subsequently collected specimens of this species and found that they belong to another new species of Heterochelamon. We herein describe two new species of Heterochelamon, including the one from Guangdong Province, representing the first member of the genus from outside of Guangxi.
To study the phylogenetic relationships of species within Heterochelamon, we used the mitochondrial 16S rRNA gene data of four species from this genus, including the two new species reported in this paper. Molecular data supports the recognition of the two new species and their assignment to Heterochelamon.
Material & Methods
Specimens were collected by Song-Bo Wang, Yi-Yang Xu, Jia-Ming Tian and Xi-Jiao Wei; preserved in 95% ethanol and deposited in the Department of Parasitology of the Medical College of Nanchang University, Jiangxi, China (NCU MCP), National Tropical Disease Research Center, Shanghai, China (TDRC). The abbreviations G1 and G2 are used for the male first gonopod and second gonopod, respectively. Carapace width and length are reported in millimeters. The terminology used primarily follows that of Dai (1999), Naruse, Zhu & Zhou (2013) and Davie, Guinot & Ng (2015).
Muscle tissue was extracted from the ambulatory legs and chelipeds, and total genomic DNA was extracted from the tissues using the Omega Tissue Kit following the manufacturer’s protocol. Mitochondrial 16S rRNA gene sequences were obtained by PCR amplification with the primers 1471 (5′-CCTGTTTANCAAAAACAT-3′) and 1472 (5′-AGATAGAAACCAACCTGG-3′) (Shih, Ng & Chang, 2004). The PCR procedure was as follows: 33 cycles of denaturation for 50 s at 94 °C, annealing for 40 s at 52 °C, and extension for 1 min at 72 °C, followed by a final extension for 10 min at 72 °C. Sequences were obtained by automated sequencing (ABI3730 automatic sequencer).
Sequences were aligned using MAFFT vers.7.355 (Nakamura et al., 2018) based on the GINS-I method and the selection of conserved regions with Gblocks 0.91b (Castresana, 2000). GTR+I+G was the best fit model for the sequence evolution of the 16S dataset, as determined with MrModeltest vers.2.2 (Nylander, 2005) and then selected based on the Akaike information criterion (AIC). MrBayes vers.3.2.6 (Ronquist et al., 2012) was used to construct a Bayesian inference (BI) tree. Four Markov chain Monte Carlo (MCMC) chains were run for 2,000,000 generations, with samples stored once every 1000 generations, discarding the first 25% as burnin. Tracer vers.1.6 (Rambaut et al., 2014) was used to check effective sample size (ESS) values (all of greater than 200). Simultaneously with the BI analysis, we used MEGA vers.X.0 (Kumar et al., 2018) to select the best evolutionary model for maximum likelihood (ML) analysis, which was the HKY+I+G model based on the Bayesian information criterion (BIC). The ML tree was built after 1000 bootstrap replicates by using MEGA vers.X.0 (Kumar et al., 2018).
The electronic version of this article in portable document format will represent a published work according to the International Commission on Zoological Nomenclature (ICZN), and hence the new names contained in the electronic version are effectively published under that Code from the electronic edition alone. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix http://zoobank.org/. The LSID for this publication is: urn:lsid:zoobank.org:pub: F680C21E-0BED-4724-BA62-1174336F433F. The online version of this work is archived and available from the following digital repositories: Peer J, PubMed Central, and CLOCKSS.
Results
Systematics
| Family Potamidae Ortmann, 1896 |
| HeterochelamonTürkay & Dai, 1997 |
| Heterochelamon huidongensen. sp. (Figs. 1–4) |
| urn:lsid:zoobank.org: act: 0F67FA91-6734-4800-9C03-76668635A1E2 |
Material examined. Holotype: male (25.8 × 23.1 mm) (NCU MCP 423601), Xinaobei Village (22°55′14.59″N 114°33′51.85″E, 47 m asl.), Baihua Town, Huidong County, Huizhou City, Guangdong Province, coll. Song-Bo Wang, Yi-Yang Xu and Jia-Ming Tian, 6th Jul. 2019. Paratypes: 3 males (30.0 × 25.8 mm, 26.2 × 23.1 mm, 21.3 × 18.6 mm) (NCU MCP 423602, TDRC 002005, TDRC 002006) and 2 females (26.1 × 23.5 mm, 20.6 × 18.5 mm) (NCU MCP 423603, TDRC 002007), same data as holotype. Others: 5 males (29.6 × 25.4 mm, 26.4 × 23.5 mm, 25.1 × 22.3 mm, 28.4 × 24.5 mm, 21.3 × 18.9 mm; NCU MCP 423606, NCU MCP 423607, NCU MCP 423608, NCU MCP 423609, NCU MCP 423610) and 2 females (21.1 × 19.1 mm, 14.5 × 12.5 mm; NCU MCP 423612, NCU MCP 423613), same data as holotype.
Comparative material. Heterochelamon yangshuoense Türkay & Dai, 1997: Holotype, male, IZCAS CB 05102, Yangshuo County, Guilin City, Guangxi Zhuang Autonomous Region, 8th May 1975. H. guangxiense Türkay & Dai, 1993: Holotype, male, IZCAS CB 01336, Guangxi Zhuang Autonomous Region, May 1974. H. purpureomanualis Türkay & Dai, 1997: not holotype, male, IZCAS CB, Yao Mountain, Xiuren Town, Lipu County, Guilin City, Guangxi Zhuang Autonomous Region, 1938. H. tessellatum Naruse, Zhu & Zhou, 2013: Holotype, male, NCU MCP 2012.0003, Dong Men Zhuang, Shuangluo Village, Sanli Town, Shanglin County, Guangxi Zhuang Autonomous Region, 14th Aug. 2006, Paratypes, 1 male, NCU MCP 2012.0004, 1 female, NCHUZOOL 13572, same data as holotype. H. castanea Naruse, Zhu & Zhou, 2013: Holotype: male, NCU MCP 2012.0001, Lituan Village, Baiming Town, Liujiang County, Liuzhou City, Guangxi Zhuang Autonomous Region, Aug. 2006, Paratype: 1 female, NCU MCP 2012. 0002, same data as holotype.
Diagnosis. Carapace subquadrate, dorsal surface covered with scattered minute pits, regions not clearly demarcated. External orbital angle triangular, sharp; postorbital cristae inconspicuous. Epibranchial tooth very sharp, distinct. Anterolateral margin cristate. Suborbital, pterygostomial regions smooth. Median lobe of epistome posterior margin protruded slightly. Third maxilliped exopod exceeding posterior margin of merus, with long slender flagellum. Chelipeds unequal, cutting edges of finger of larger cheliped with blunt teeth, with narrow gap when fingers closed. Ambulatory legs slender. Male telson triangular, lateral margins slightly concave. Female vulva located at middle of sternite 6. G1 terminal segment flat and medially bent, not tapered distally, apex rounded, reaching pleonal locking tubercle on mid length of sternite 5 but not exceeding sternites 4/5 suture.

Figure 1: Heterochelamon huidongense. n. sp. Holotype male (25.8 × 23.1 mm) (NCU MCP 423601).
(A) Overall habitus; (B) frontal view of the cephalothorax. Scales = one cm. Photo credit: Song-Bo Wang.{kind=link}
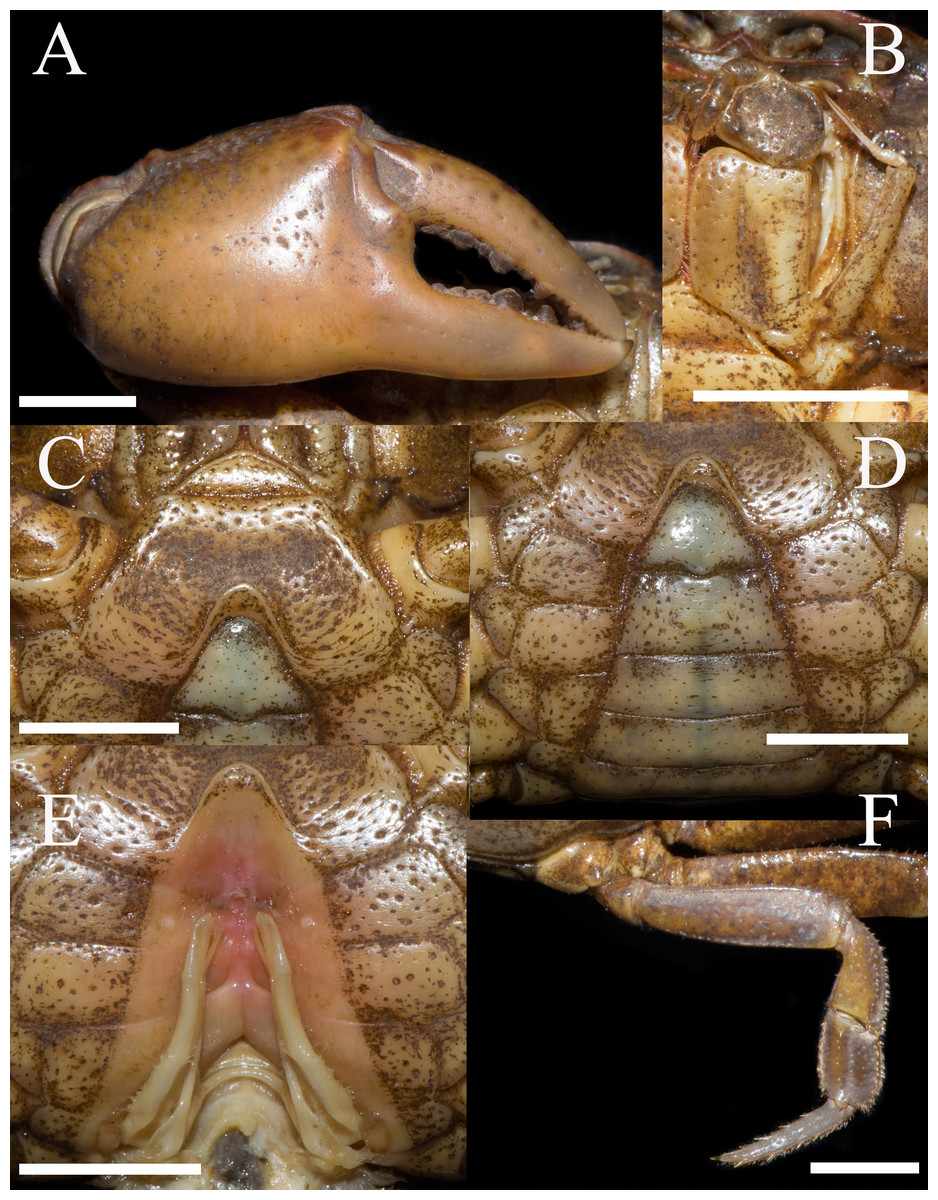
Figure 2: Heterochelamon huidongense n. sp. Holotype male (25.8 × 23.1 mm) (NCU MCP 423601).
(A) Outer view of larger cheliped; (B) left third maxilliped; (C) ventral view of anterior thoracic sternum; (D) ventral view of pleon; (E) ventral view of sterno-pleonal cavity with right G1 in situ; (F) right fourth ambulatory leg. Scales = 0.5 cm. Photo credit: Song-Bo Wang.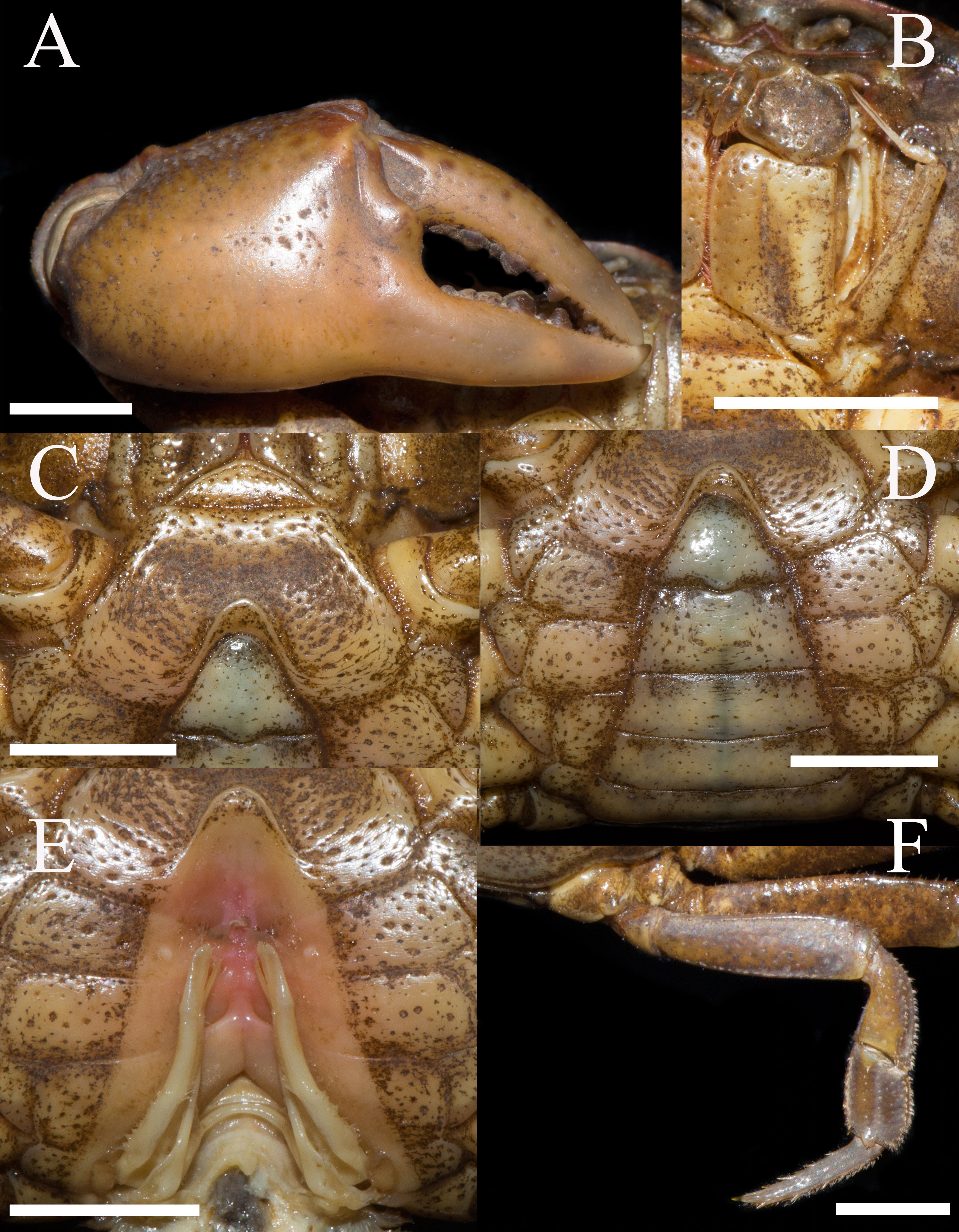{kind=link}
Description. Carapace subquadrate, width about 1.1 times as length (n = 13), regions not clearly demarcated; dorsal surface convex longitudinally, smooth (Figs. 1A, 3A). External orbital angle triangular, sharp, separated from anterolateral margin by U-shaped notch; postorbital cristae inconspicuous, postfrontal lobe slightly convex, separated from each other; cervical groove indistinct; H-shaped gastric groove indiscernible (Figs. 1A and 3A). Epibranchial tooth very sharp, distinct; anterolateral margin cristate, straight, lined with 7–8 granules, shorter than posterolateral margin; posterolateral surface smooth (Figs. 1A, 3A). Frontal, supra-, infraorbital margins cristate, lined with indistinct granules; suborbital, pterygostomial regions smooth, with a few round granules (Fig. 1B). Epistome posterior margin distinctly cristate, median lobe low, triangular, lateral margins slightly sinuous (Fig. 1B).

Figure 3: Heterochelamon huidongense. n. sp. Paratype female (26.1 × 23.5 mm) (NCU MCP 423603).
(A) Overall habitus; (B) ventral view of pleon; (C) vulvae. Scales = one cm. Photo credit: Song-Bo Wang.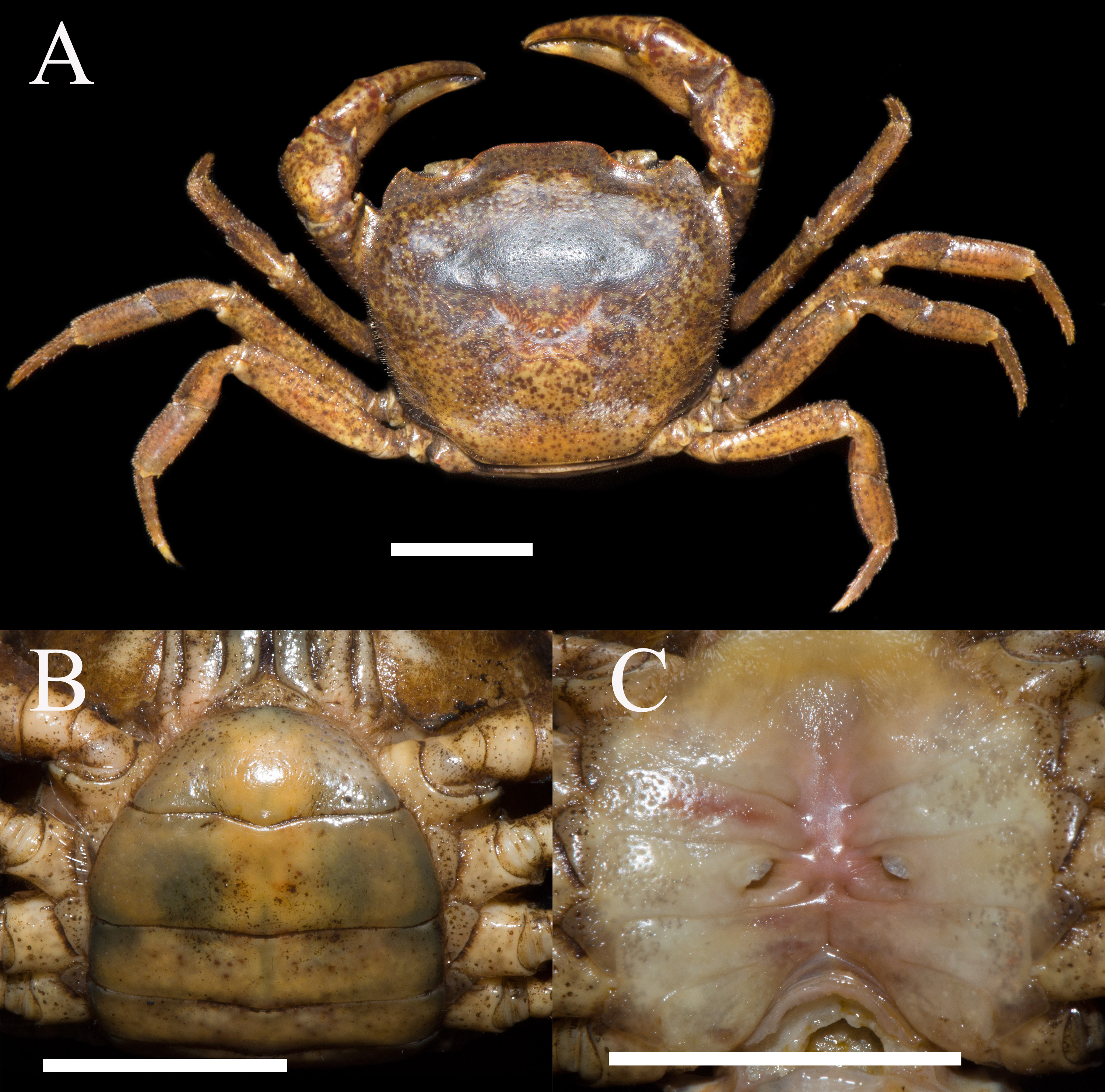{kind=link}
Third maxilliped exopod exceeding posterior margin of merus, with long slender flagellum; merus about 1.2 times as broad as long, slightly depressed medially; ischium about 1.4 times as long as broad, with distinct median sulcus (Fig. 2B).
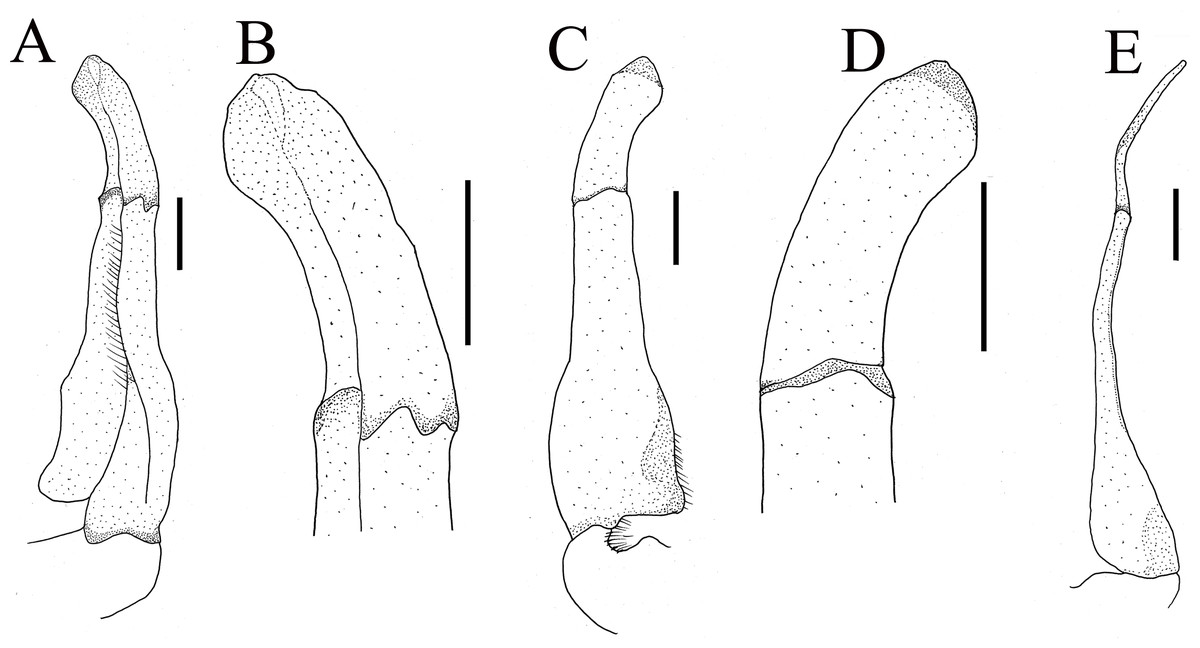
Figure 4: Gonopods of holotype of Heterochelamon huidongense n. sp.
(A) Ventral view of the left G1; (B) ventral view of the terminal segment of left G1; (C) dorsal view of the left G1; (D) dorsal view of the terminal segment of left G1; (E) ventral view of the left G2. Scales = one mm. Photo credit: Song-Bo Wang.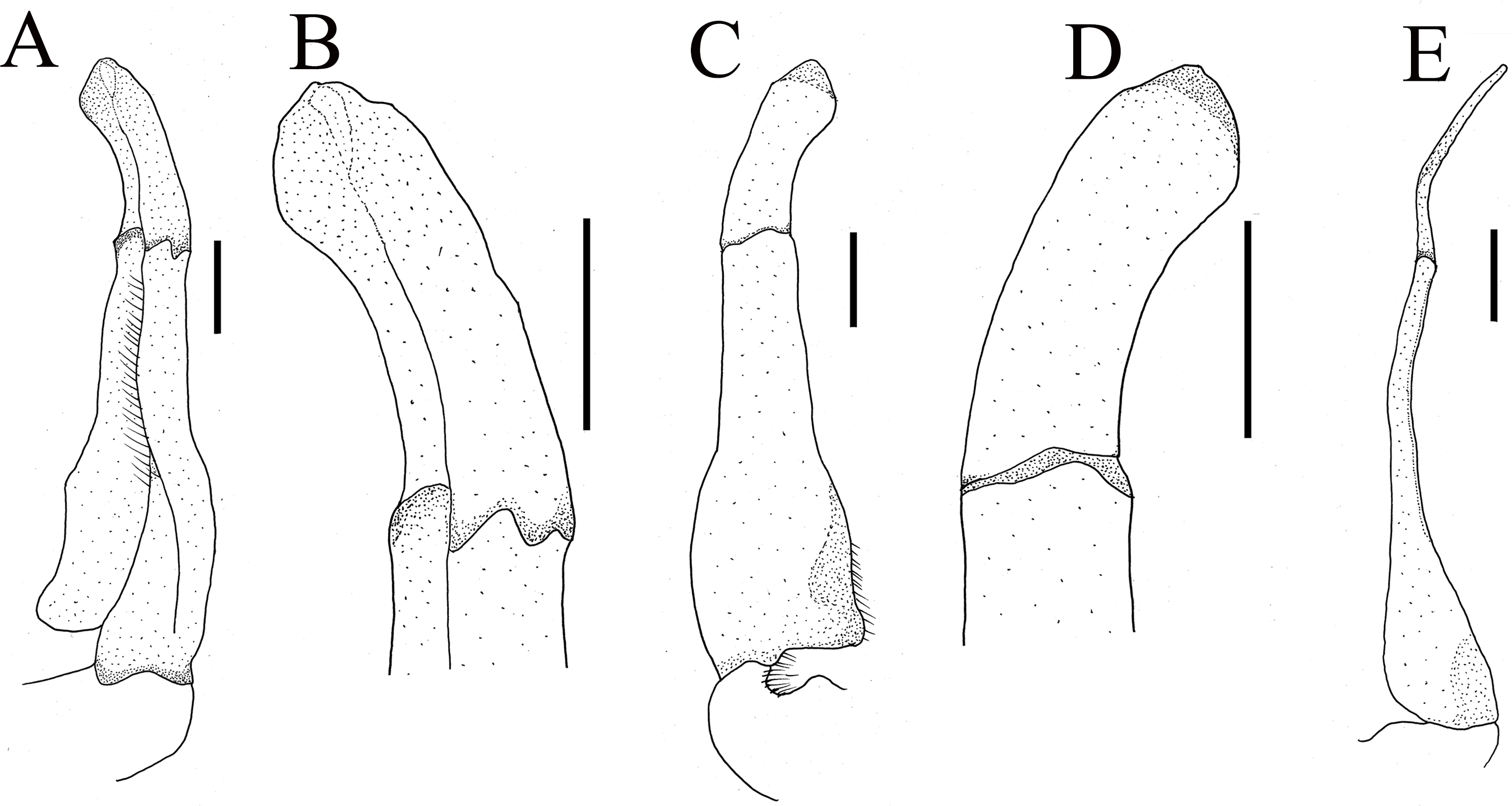{kind=link}
Male chelipeds unequal (Fig. 1A). Merus surface smooth without any pits; carpus with inner angle produced into strong spine, surface sparsely covered with pits (Fig. 1A). Cutting edges of larger cheliped fingers with row of large blunt teeth, with narrow gap when fingers closed (Fig. 2A). Ambulatory legs slender; third legs longest when stretched laterally, fourth ambulatory leg propodus about 1.6 times as long as broad, slightly shorter than dactylus, with scattered thorn-like setae (Fig. 2E).
Male thoracic sternites 2/3 demarcated by distinct suture, sternites 3/4 demarcated by shallow groove (Fig. 2C). Male sterno-pleonal cavity deep, reaching anteriorly to mid-length of thoracic sternite 4; median groove between sternites 7/8 long (Fig. 2E). Male pleon triangular, somites 3–6 gradually decreasing in width, increasing in length in males; somite 3 widest; somite 6 trapezoidal, margins slightly oblique (Fig. 2D). Telson triangular with rounded apex, lateral margin slightly concave (Fig. 2C). Female pleon broadly ovate (Fig. 3B); vulva small, ovate, located at middle of sternite 6, opening inwards, posterior margin slightly bulged (Fig. 3C).
G1 slender; terminal segment flat, bent medially, not tapered distally, distal part rounded; exceeding pleonal locking tubercle on mid length of sternite 5, not reaching suture between sternites 4/5 (Figs. 4A–4D, 2F). Distinct boundary between terminal segment and subterminal segment, latter length about 2.5 times as former, groove for G2 in ventral surface; G2 basal segment triangular, length about 2.1 times distal segment (Fig. 4).
Etymology. The species is named after the type locality, Huidong County, Huizhou City, Guangdong Province.
Distribution. The new species is known only from the type locality presently, Huidong County, Huizhou City, Guangdong Province, southern China.
Ecology. Specimens of the new species were collected at three sites along a river using fish cages or by hand (Figs. 5B, 5C). The water of the river is relatively clear, and the water flow is slow. The width of the river is approximately 1–3 m and the depth is approximately 0.2–0.3 m. The river bed is mainly sand and soil (Fig. 5E). There are many shrubs and low hills around the river, many households located dozens of meters away from the river and some vegetable gardens along the river (Fig. 5A). At one of the collection points, we also collected specimens of the gecarcinucid Somanniathelphusa sinensis H. Milne-Edwards, 1853 (Fig. 5D), which we believe coexists with the new species in the river.

Figure 5: Habitat Environment of Heterochelamon huidongense n. sp.
(A) Surrounding environment of the collection point; (B) living specimen of H. huidongense n. sp. collected with a fish cage; (C) H. huidongense n. sp. in the water; (D) living specimen of Somanniathelphusa Bott, 1968, collected with a fish cage; (E) collection point. Photo credit: A, B, D, E from Song-Bo Wang and C from Chao Huang.{kind=link}
Remarks. Compared to congeners, Heterochelamon huidongense n. sp. can easily be separated by its G1 not reaching sternites 4/5 suture in situ and terminal segment being medially bent (Figs. 2E and 4A, Table 1) (versus reaching sternites 4/5 suture and straight in congeners, except H. tessellatum which has G1 that also does not reach the sternites 4/5 suture but is strongly bent proximally), as well as a combination of characters: very sharp epibranchial tooth , triangular external orbital angle, slightly convex anterolateral margins (Fig. 1A), triangular male telson (Fig. 2C), and inward opening female vulva (Fig. 3C). For detailed differences between this new species and congeners, see Table 1.
| H. huidongense | H. jinxiuense | H. tessellatum | H. castanea | H. purpureomanualisa | H. purpureomanualisb | H. guangxiense | H. yangshuoense | |
|---|---|---|---|---|---|---|---|---|
| External orbital angle | Triangular (Fig. 1A) | Blunt (Fig. 6A) | Acutely triangular (cf. Naruse, Zhu & Zhou, 2013: Fig. 1A) | Triangular (cf. Naruse, Zhu & Zhou, 2013: Fig. 4A) | Acutely triangular (cf. Wu, 1934: Fig. 2) | Triangular (cf. Türkay & Dai, 1997) | Blunt (cf. Türkay & Dai, 1997) | Triangular (cf. Türkay & Dai, 1997) |
| Epibranchial tooth | Very sharp, separated from external orbital angle by broad notch (Fig. 1A) | Blunt, separated from external orbital angle by narrow notch (Fig. 6A) | Very sharp, separated from external orbital angle by broad notch (cf. Naruse, Zhu & Zhou, 2013: Fig. 1A) | Very sharp, separated from external orbital angle by narrow notch (cf. Naruse, Zhu & Zhou, 2013: Fig. 4A) | Very sharp, separated from external orbital angle by narrow notch (cf. Wu, 1934: Fig. 2) | Blunt, separated from external orbital angle by narrow notch (cf. Türkay & Dai, 1997) | Very sharp, separated from external orbital angle by broad notch (cf. Türkay & Dai, 1997) | Very sharp, separated from external orbital angle by broad notch (cf. Türkay & Dai, 1997) |
| Adult male chelipeds | Unequal (Fig. 1A) | Strongly unequal (Fig. 6A) | Strongly unequal (cf. Naruse, Zhu & Zhou, 2013: Fig. 1A) | Unequal (cf. Naruse, Zhu & Zhou, 2013: Fig. 4A) | Strongly unequal (cf. Wu, 1934: Fig. 2) | Strongly unequal (cf. Türkay & Dai, 1997) | Unequal (cf. Türkay & Dai, 1997) | Strongly unequal (cf. Türkay & Dai, 1997) |
| Gap of adult male major chela fingers when closed | Narrow, wedge-shaped (Fig. 2A) | Medium, teardrop-shaped (Fig. 7A) | Very broad, oblong (cf. Naruse, Zhu & Zhou, 2013: Fig. 2B) | Narrow, wedge-shaped (cf. (Naruse, Zhu & Zhou, 2013): Fig. 5B) | Very broad, oblong (cf. Wu, 1934: Fig. 2) | Very broad, oblong (cf. Türkay & Dai, 1997) | Narrow, wedge-shaped (cf. Türkay & Dai, 1997) | Very broad, oblong (cf. Türkay & Dai, 1997: fig. 6-2) |
| G1 in situ | Not reaching sternites 4/5 suture (Fig. 2E) | Reaching sternites 4/5 suture (Fig. 8E) | Not reaching sternites 4/5 suture (cf. Naruse, Zhu & Zhou, 2013: Fig. 2A) | Reaching sternites 4/5 suture (cf. Naruse, Zhu & Zhou, 2013) | No information | Reaching sternites 4/5 suture (cf. Türkay & Dai, 1997: fig. 4-3) | Reaching sternites 4/5 suture (cf. Türkay & Dai, 1997: fig. 5-3) | Reaching sternites 4/5 suture (cf. Türkay & Dai, 1997: fig. 6-4) |
| Terminal segment of G1 | Flat, bent in middle, not tapered distally, distal part rounded (Fig. 4) | Rod-like, straight, tapered distally, distal part sharp, bent slightly at distal (Fig. 9) | Knuckle-shaped, strongly bent proximally, widening distally, widest part more than twice as wide as base (cf. Naruse, Zhu & Zhou, 2013: Fig. 3A) | Rod-like, straight, not tapered distally, distal part rounded and bent slightly (cf. Naruse, Zhu & Zhou, 2013: Fig. 6A) | No information | Rod-like, straight, not tapered distally, distal part protruding slightly in inner edge (cf. Türkay & Dai, 1997: fig. 4-4) | Rod-like, slightly bent, not tapered distally, distal part protruding in inner edge (cf. Türkay & Dai, 1997: fig. 5-4) | Knuckle-shaped, straight, widening distally, widest part more than twice as wide as base (cf. Türkay & Dai, 1997: fig. 6-5) |
Notes:
Material examined. Holotype: 1 male (24.6 × 19.8 mm) (NCU MCP 342001), from Tongfu Village (24°17′10.89″N115°5′17.28″E, 267 m asl.), Toupai Town, Jinxiu Yao Autonomous County, Laibin City, Guangxi Zhuang Autonomous Region, coll. Xi-Jiao Wei, 8th Jul. 2011. Paratypes: 3 males (20.3 × 15.8 mm, 15.8 × 12.6 mm, 17.8 × 14.8 mm) (NCU MCP 342002, TDRC 002008, TDRC 002009) and 1 female (12.9 × 10.4 mm) (NCU MCP 342003), same data as holotype. Other: 1 male (14.1 × 11.4 mm) (NCU MCP 342006), same data as holotype.

Figure 6: Heterochelamon jinxiuense. n. sp. Holotype male (24.6 × 19.8 mm) (NCU MCP 342001).
(A) Overall habitus; (B) frontal view of the cephalothorax. Scales = one cm. Photo credit: Song-Bo Wang.{kind=link}

Figure 7: Heterochelamon jinxiuense. n. sp. Holotype male (24.6 × 19.8 mm) (NCU MCP 342001).
(A) Outer view of larger cheliped; (B) left third maxilliped; (C) ventral view of anterior thoracic sternum; (D) ventral view of pleon; (E) ventral view of sterno-pleonal cavity with right G1 in situ; (F) right fourth ambulatory leg. Scales = 0.5 cm. Photo credit: Song-Bo Wang.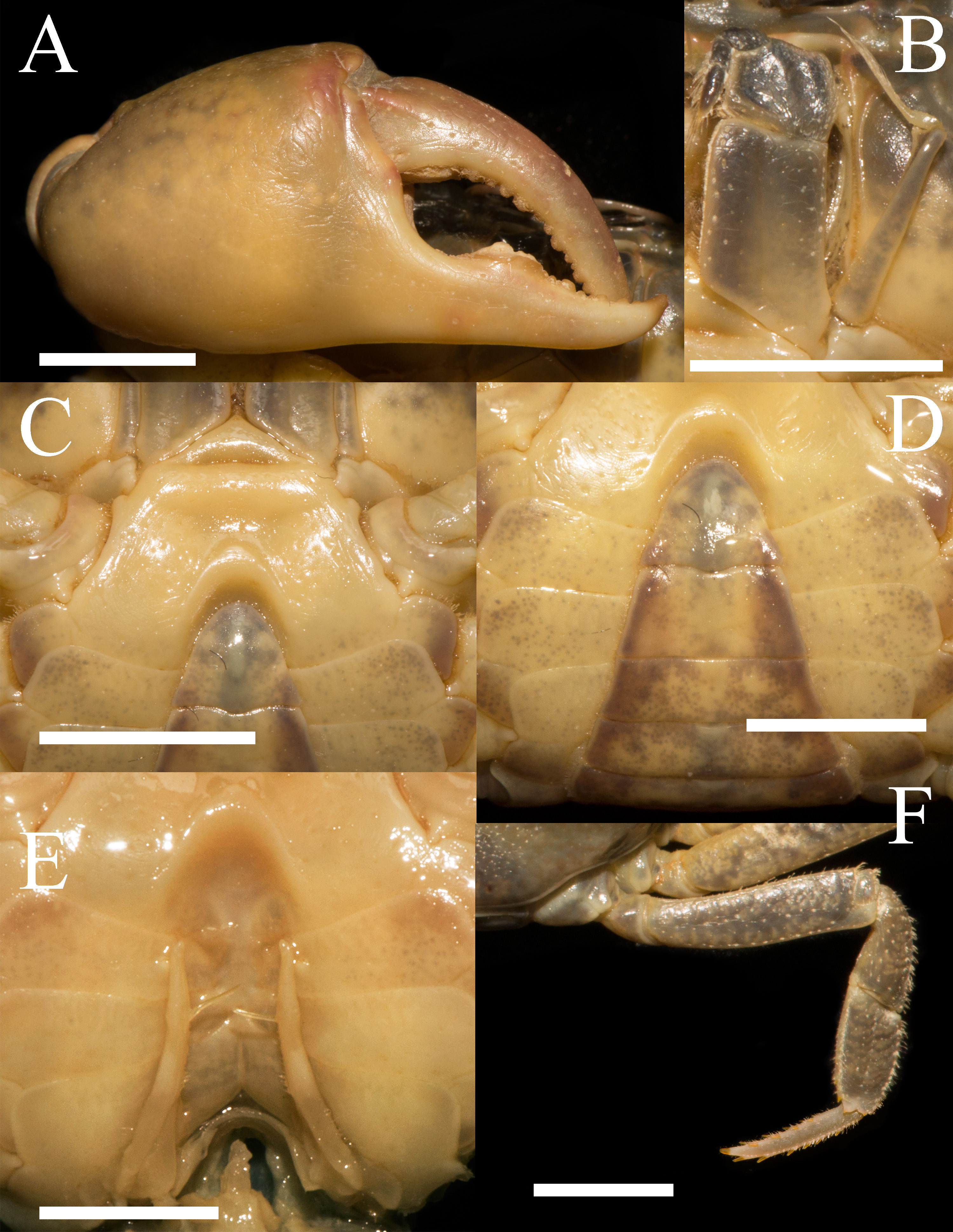{kind=link}
Comparative material. See for Heterochelamon huidongense n. sp.
Diagnosis. Carapace trapezoidal, flat, surface covered with scattered minute pits. External orbital angle blunt, postorbital cristae, postfrontal lobe indiscernible. Epibranchial tooth blunt, distinct; anterolateral margin cristate, slightly convex. Suborbital, pterygostomial regions very smooth. Median lobe of epistome posterior margin distinctly protruding. Third maxilliped exopod with long flagellum. Chelipeds strongly unequal in adult males, surface smooth, lower cutting edge of major cheliped finger with very large, blunt teeth, with medium gap when fingers closed. Ambulatory legs slender. Thoracic sternites 2/3 demarcated by distinct deep suture, anterior part of sternite 3 convex. Male sterno-pleonal cavity deep, narrow. Male telson triangular, lateral margin straight. Female vulva reaching sternites 5/6 suture. Male G1 terminal segment rod-like, tapered distally, subdistal segment suddenly inwardly bent, reaching sternites 4/5 suture.
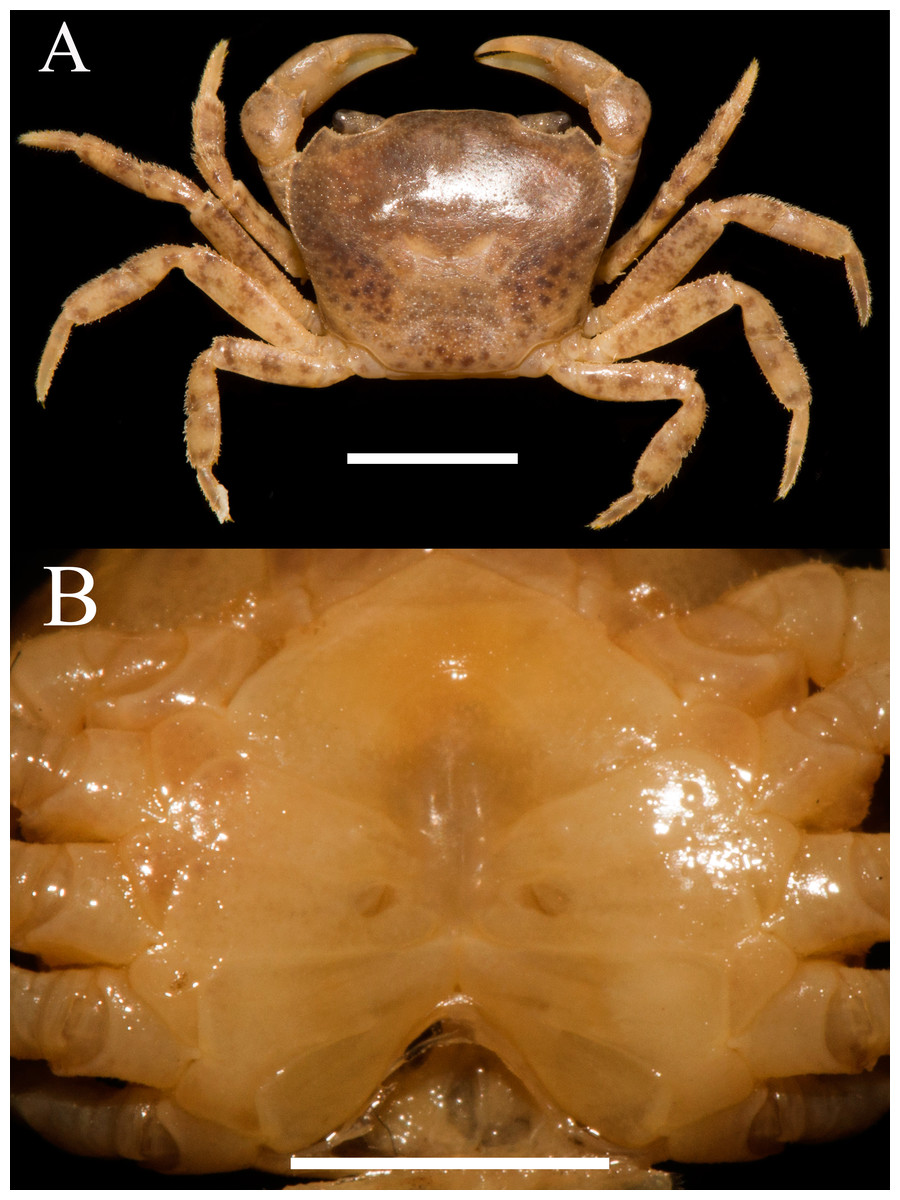
Figure 8: Heterochelamon jinxiuense. n. sp. Paratype female (12.9 × 10.4 mm) (NCU MCP 342003).
(A) Overall habitus; (B) ventral view of vulvae. Scales = one cm. Photo credit: Song-Bo Wang.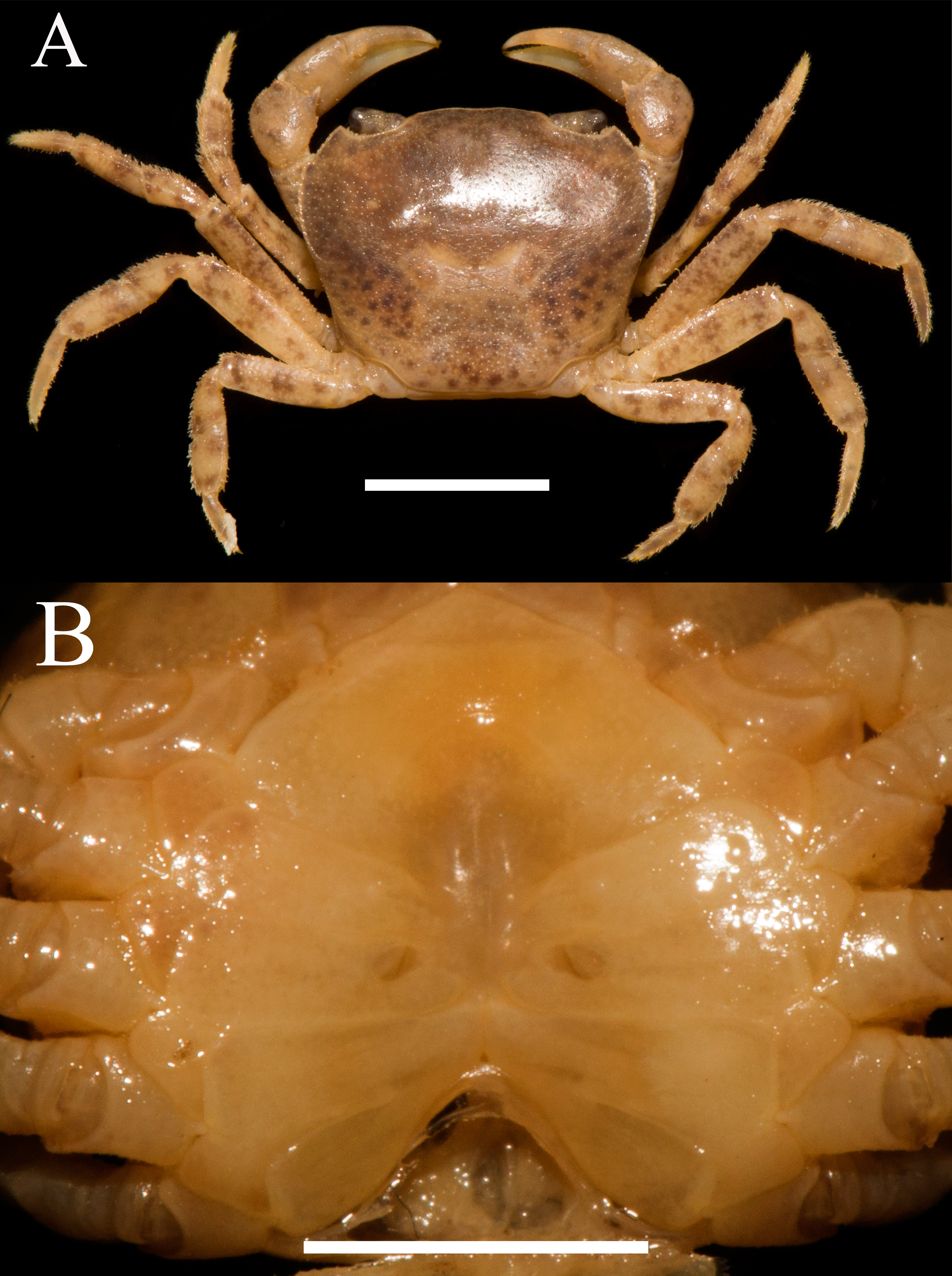{kind=link}
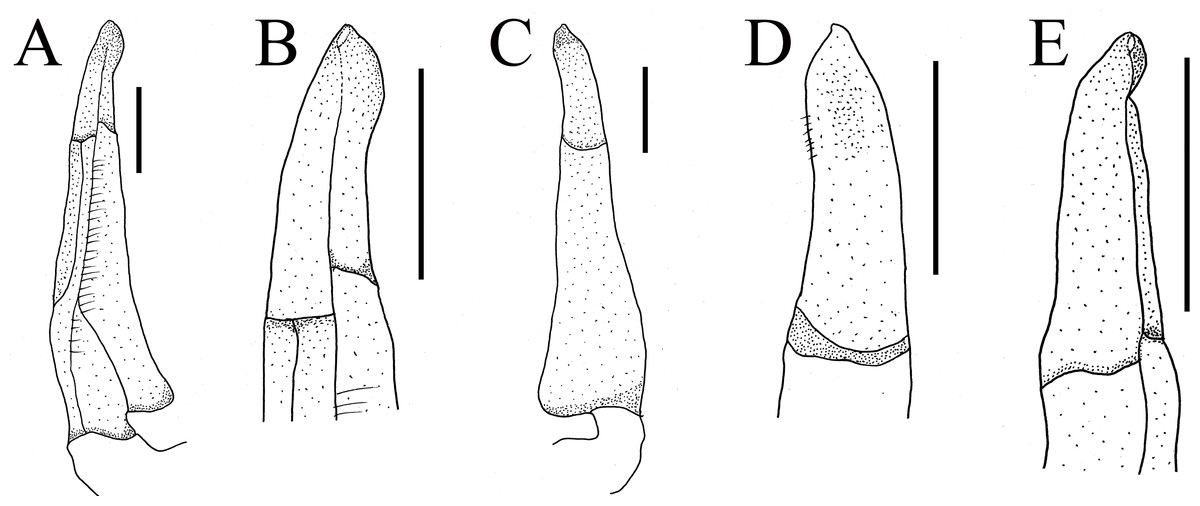
Figure 9: The first gonopods of holotype of Heterochelamon jinxiuense n. sp.
(A) Ventral view of the right G1; (B) ventral view of the terminal segment of right G1; (C) dorsal view of the right G1; (D) dorsal view of the terminal segment of right G1; (E) side view of the right G1. Scales = one mm. Photo credit: Song-Bo Wang.{kind=link}
Description. Carapace trapezoidal, flat, width about 1.3 times as length (n = 6), regions not clearly demarcated; dorsal surface slightly convex, covered with inconspicuous pits (Figs. 6A, 8A). External orbital angle blunt, separated from anterolateral margin by shallow notch; postorbital cristae inconspicuous, postfrontal lobe indistinct; cervical groove indistinct; H-shaped gastric groove indiscernible (Figs. 6A, 8A). Epibranchial tooth blunt, distinct; anterolateral margin cristate, slightly convex, lined with several granules, shorter than posterolateral margin; posterolateral surface smooth (Figs. 6A, 8A). Frontal and orbital margins cristate, lined with indistinct granules, supra-, infraorbital margins glazed; suborbital, pterygostomial regions very smooth, not covered with granules (Fig. 6B). Epistome posterior margin distinctly cristate, median lobe triangular, distinctly protruding, lateral margins almost straight (Fig. 6B).
Third maxilliped exopod exceeding posterior margin of merus, with slender flagellum; merus about 1.2 times as broad as long, with median depression; ischium about 1.4 times as long as broad, longitudinal median sulcus distinct (Fig. 7B).
Chelipeds strongly unequal in adult males (Fig. 7A). Merus surface smooth without any pits; carpus inner angle forming stout spine, surface not covered with pits (Fig. 1A). Cutting edge of larger cheliped fixed finger with large rounded teeth, movable finger cutting edge lined with several small teeth, with medium gap when fingers closed (Fig. 7A). Ambulatory legs slender; third legs longest, the fourth ambulatory leg propodus about 2.0 times as long as broad, equal to dactylus in length (Figs. 6A, 7F).
Male thoracic sternites 2/3 demarcated by distinct deep suture, anterior part of sternite 3 convex, sternites 3/4 suture discernible as shallow groove, sterno-pleonal cavity reaching imaginary line joining posterior third of cheliped coxae (Fig. 7C). Male sterno-pleonal cavity deep, narrow; median longitudinal groove between sternites 7/8 medium-length (Fig. 7E). Male pleon triangular, third somite widest; somite 6 trapezoidal; somites 3–6 gradually decreasing in width (Fig. 7D). Telson triangular with rounded apex, lateral margin slightly convex (Fig. 7D). Female vulva very small, ovate, reaching sternites 5/6 suture, located at upper of sternite 6 (Fig. 8B).
Male G1 slender; terminal segment rod-like, almost straight, tapered distally; apex sharp, abruptly bent inwards subdistally; reaching beyond pleonal locking tubercle on mid length of sternite 5, to suture between sternites 4/5 (Figs. 7E, 9). Boundary between terminal segment and subterminal segment clear, latter length about 2.4 times as former, groove for G2 in ventral surface, G2 destroyed during dissection (Fig. 9).
Etymology. The species is named after the type locality, Jinxiu Yao Autonomous County, Laibin City, Guangxi Zhuang Autonomous Region.
Distribution. The new species is known only from the type locality presently, Jinxiu Yao Autonomous County, Laibin City, Guangxi Zhuang Autonomous Region, southern China.
Ecology. The collection site is a small river running behind Tongfu village. The river is mostly surrounded by dwarf mountains and many kinds of fruit trees. The river is approximately 2 m in width, the water depth is approximately 0.4 m, the riverbed mainly consists of sand and gravel and the water quality is good.
Remarks
Heterochelamon jinxiuense n. sp. can easily be differentiated from its congeners by its distally tapered G1 (Fig. 9, Table 1) (versus widening distally or not tapered distally in congeners), blunt external orbital angle (Fig. 6A, Table 1) (versus triangular or acutely triangular in congeners, except that of H. guangxiense which is also blunt), strongly unequal chelipeds in adult males and medium gap when male major chela fingers closed (Fig. 7A, Table 1) (versus strongly unequal in H. tessellatum, H. purpureomanualis and H. yangshuoense, but fingers with very broad and oblong gap when closed; other species unequal with narrow gap when closed). The detailed differences between this new species and congeners are also presented in Table 1.
The new species was collected in 2011, of which the material consisted of five males (one adult and four immature) and one female. The G2 of the only mature male had been destroyed by the time we examined the specimens. We were also unable to find this species in a subsequent collection trip. As of such, the G2 is not described here.
Phylogenetic analyses
We obtained the mitochondrial 16S rRNA gene molecular data from four species of Heterochelamon for this study. Other sequences used were downloaded from GenBank. To show where these two new species fit within the Asian freshwater crab phylogeny, 60 species of 47 genera in the family Potamidae Ortmann, 1896, were used (Table 2). The trees were constructed via the BI and ML methods and showed a high degree of consistency, and the topological structure was determined from BI with support values from both methods (Fig. 10). The phylogenetic results support the recognition of the two new Heterochelamon species, H. tessellatum and H. castanea cluster with the two new species to form an independent branch in the “China-East Asia Islands” clade (Shih, Yeo & Ng, 2009). It is possible that the two new species are in sister relationship. H. castanea is most closely related to the two new species in the two known congeneric species available for the molecular phylogenetic analysis.
| Species | MuseumCatalogue No. | Locality | GenBank No. |
|---|---|---|---|
| Heterochelamon huidongense n. sp. | NCU MCP 423604 | Guangdong, China | MN823645 |
| Heterochelamon huidongense n. sp. | NCU MCP 423605 | Guangdong, China | MN823646 |
| Heterochelamon huidongense n. sp. | NCU MCP 423606 | Guangdong, China | MN823647 |
| Heterochelamon jinxiuense n. sp. | NCU MCP 342004 | Guangdong, China | MN823648 |
| Heterochelamon jinxiuense n. sp. | NCU MCP 342005 | Guangdong, China | MN823649 |
| Heterochelamon tessellatumNaruse, Zhu & Zhou, 2013 | NCU MCP 153601 | Guangxi, China | MN823650 |
| Heterochelamon tessellatumNaruse, Zhu & Zhou, 2013 | NCU MCP 153602 | Guangxi, China | MN823651 |
| Heterochelamon castaneaNaruse, Zhu & Zhou, 2013 | NCU MCP 341901 | Guangxi, China | MN823652 |
| Amamiku amamense Minei, 1973 | NCHUZOOL 13125 | Amami, the Ryukyus | AB428457 |
| Aparapotamon grahami Rathbun, 1929 | ZRC YCM 0334(II) | Yunnan, China | AB428489 |
| Apotamonautes hainanensis Parisi, 1916 | ZRC | Hainan, China | AB428459 |
| Beccumon jarujini Ng & Naiyanetr, 1993 | ZRC 1991.1865 (paratype) | Chiangma, Thailand | AB428479 |
| Candidiopotamon rathbunae De Man, 1914 | NCHUZOOL | Nantou, Taiwan | AB208598 |
| Cantopotamon yangxiense Chao, Ahyong & Shih, 2017 | SYSBM 001564 | Guangdong, China | LC342050 |
| Cantopotamon zhuhaiense Chao, Ahyong & Shih, 2017 | SYSBM 001439 | Guangdong, China | LC342045 |
| Chinapotamon glabrum Dai, Song, Li & Liang, 1980 | CAS CB | Guangxi, China | AB428451 |
| Cryptopotamon anacoluthon Kemp, 1918 | NCHUZOOL 13122 | Hong Kong | AB428453 |
| Daipotamon minos Ng & Trontelj | ZRC 1996.1045 | Guizhou, China | LC198524 |
| Demanietta renongensis Rathbun, 1905 | ZRC 1998.146 | Ranong, Thailand | AB428475 |
| Diyutamon cereum Huang, Shih & Ng | SYSBM | Guizhou, China | LC198520 |
| Eosamon smithianum Kemp, 1923 | ZRC | Chantaburi, Thailand | AB428486 |
| Eosamon yotdomense Naiyanetr, 1984 | ZRC 1991.1851 | Ubon Ratchathani, Thailand | AB428485 |
| Esanpotamon namsom Naiyanetr & Ng, 1997 | ZRC 1997.776 (paratype) | Udon Thani, Thailand | AB428463 |
| Eurusanm guangdongenseHuang, 2018 | SYSBM 001408 | Guangxi, China | MG709242 |
| Flabellamon sp. | ZRC | Mae Sot, Thailand | AB428472 |
| Geothelphusa albogilva Shy, Ng & Yu, 1994 | NCHUZOOL | Pingtung, Taiwan | AB127366 |
| Geothelphusa marginata fulva Naruse, Shokita & Shy, 2004 | NCHUZOOL 13124 | Iriomote, the Ryukyus | AB428456 |
| Geothelphusa olea Shy, Ng & Yu, 1994 | NCHUZOOL 13123 | Taichung, Taiwan | AB428455 |
| Hainanpotamon fuchengense Dai, 1995 | NCHUZOOL 13128 | Hainan, China | AB428461 |
| Huananpotamon angulatum Dai & Lin, 1979 | ZRC | Fujian, China | AB428454 |
| Indochinamon tannanti Rathbun, 1904 | ZRC 1998.264 | Yunnan, China | AB428482 |
| Johora johorensis Roux, 1936 | ZRC 1990.576 | Gunung Pulai, Johor, Malaysia | AB290620 |
| Johora murphyi Ng, 1986 | ZRC 2001.2267 | Kota Tinggi, Johor, Malaysia | AB290621 |
| Kanpotamon duangkhaei Ng & Naiyanetr, 1993 | ZRC | Kanchanaburi, Thailand | AB428471 |
| Kukrimon cucphuongense Dang, 1975 | ZRC NHH9729 160997 | Ninh Binh, Vietnam | AB428483 |
| Longpotamon baiyanense Ng & Dai, 1997 | ZRC | Hunan, China | AB428470 |
| Longpotamon planum Dai, 1992 | ZRC 1998.1178 | Anhui, China | AB428469 |
| Mediapotamon leishanense Dai, 1995 | SYSBM 001094 | Guizhou, China | LC155164 |
| Mediapotamon liboenseWang, Zhou & Zou, 2019 | NCU MCP 343004 | Guizhou, China | MK820377 |
| Megacephalomon kittikooni Yeo & Naiyanetr, 1999 | ZRC 1998.22 (holotype) | Xieng Khuang, Laos | AB428462 |
| Mindoron balssi Bott, 1968 | ZRC | Mindoro, the Philippines | AB428464 |
| Minpotamon nasicum Dai & Chen, 1979 | NCHUZOOL 13121 | Fujian, China | AB428450 |
| Nanhaipotamon formosensis Parisi, 1916 | NCHUZOOL 13144 | Tainan, Taiwan | AB212867 |
| Nanhaipotamon nanriense Dai, 1997 | CAS CB05103 | Fujian, China | AB212868 |
| Neotiwaripotamon jianfengense Dai & Naiyanetr, 1994 | NCHUZOOL 13127 | Hainan, China | AB428460 |
| Ovitamon artifrons Bürger, 1894 | ZRC | Luzon, the Philippines | AB428466 |
| Parapotamon spinescens Calman, 1905 | NCU MCP | Yunnan, China | AB428467 |
| Pararanguna semilunatum Dai & Chen, 1985 | ZRC | Yunnan, China | AB428490 |
| Potamiscus yiwuensis Dai & Cai, 1998 | ZRC | Yunnan, China | AB428476 |
| Potamiscus yongshengense Dai & Chen, 1985 | NNU150951 | Yunnan, China | KY963597 |
| Pudaengon sakonnakorn Ng & Naiyanetr, 1995 | ZRC | Thailand | AB428484 |
| Pupamon nayung Naiyanetr, 1993 | ZRC 1995.558 (paratype) | Udon Thani, Thailand | AB428477 |
| Qianguimon elongatumHuang, 2018 | SYSBM 001424 | Guangdong, China | MG709240 |
| Ryukyum yaeyamense Minei, 1973 | NCHUZOOL 13126 | Iriomote, the Ryukyus | AB428458 |
| Shanphusa curtobates Kemp, 1918 | NRM 13920 | Taunggyi, Shan State, Myanmar | AB428478 |
| Socotrapotamon nojidensis Apel & Brandis, 2000 | ZRC 2000.2232 | Socotra, Yemen | AB428493 |
| Tenuipotamon huaningense Dai & Bo, 1994 | CAS CB05175 | Yunnan, China | AB428491 |
| Thaiphusa sp. | ZRC 1997.656 | Thailand | AB428474 |
| Tomaculamon pygmaeus Yeo & Ng, 1997 | ZRC 1997.326-330 (paratype) | Phitsanulok, Thailand | AB428473 |
| Trichopotamon daliense Dai & Chen, 1985 | NCHUZOOL 13130 | Yunnan, China | AB428492 |
| Yarepotamon breviflagellum (Huang, 2018) | SYSBM 001442 | Guangdong, China | MG709236 |
| Yarepotamon fossor (Huang, 2018) | SYSBM 001417 | Guangxi, China | MG709238 |
| Yarepotamon gracilipa Dai, Song, Li & Liang, 1980 | ZRC | Guangxi, China | AB428452 |
| Yarepotamon meridianumHuang, 2018 | SYSBM 001581 | Guangdong, China | MG709237 |
Notes:
- CAS CB
-
Chinese Academy of Sciences, Beijing, China
- NCHUZOOL
-
Zoological Collections of the Department of Life Science, National Chung Hsing University, Taichung, Taiwan
- NCU
-
MCP, Department of Parasitology of the Medical College of Nanchang University, Jiangxi, China
- NNU
-
College of Life Sciences, Nanjing Normal University, Nanjing, China
- NRM
-
Swedish Museum of Natural History, Stockholm, Sweden
- SYSBM
-
Sun Yat-sen Museum of Biology, Sun Yat-Sen University, Guangzhou, China
- ZRC
-
Zoological Reference Collection of the Raffles Museum of Biodiversity Research, National University of Singapore, Singapore
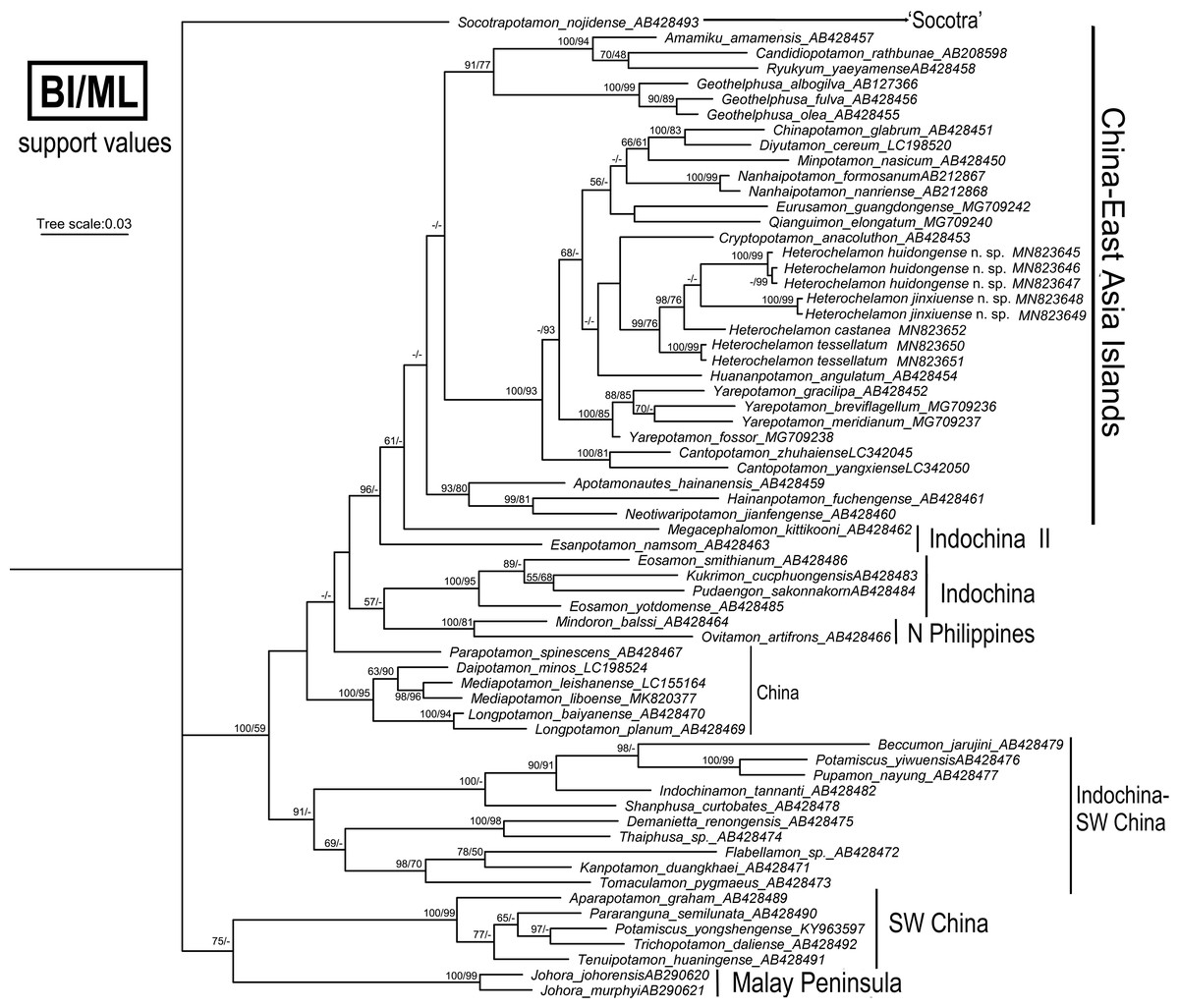
Figure 10: Phylogenetic tree.
Reconstructed base on the 16S rRNA genes of Heterochelamon and some species of Asian potamids sampled for comparison. Branch lengths and topologies were obtained from BI analysis. Probability values at the nodes represent support values for BI and ML. Only values > 50% are shown. Photo credit: Song-Bo Wang.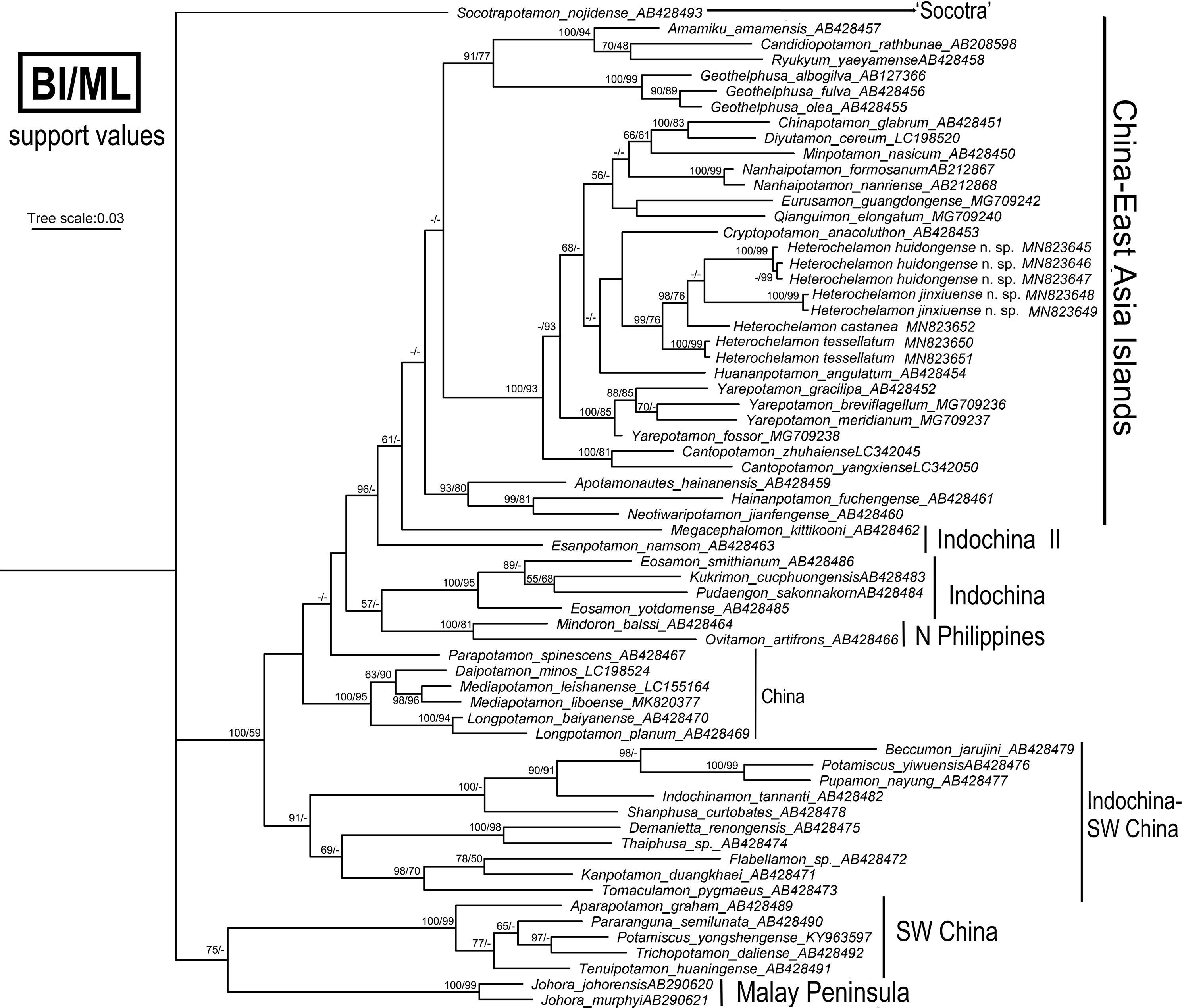{kind=link}
Discussion
The genetic analysis supports the recognition of four species of Heterochelamon which can also be differentiated morphologically. These four species form a monophyletic group with relatively high support values. Due to the lack of specimens of the other three species of Heterochelamon, we were unable to obtain their molecular data for analysis and therefore unable to determine the phylogenetic relationships of these species with the above four species.
Phylogenetically, it is possible that Heterochelamon huidongense n. sp. is more closely related to Heterochelamon jinxiuense n. sp. than any other congener (Fig. 10). However, the two can be immediately separated by the following characters: the epibranchial tooth of H. huidongense is very sharp (versus blunt in H. jinxiuense) (Figs. 1A and 6A), adult male chelipeds of H. huidongense unequal (versus strongly unequal in H. jinxiuense) (Figs. 1A and 6A), G1 terminal segment of H. huidongense bent in middle and not tapered distally (versus straight and tapered distally in H. jinxiuense) (Figs. 4A and 9A). The detailed differences among the two new species and other congeneric species are also presented in Table 1.
Türkay & Dai (1997) established Heterochelamon with Potamon (Geothelphusa) pupureomanualis (Wu, 1934) as a type species. However, the real identity of the specimens examined by Türkay & Dai (1997) is questionable, because their specimens are different from the original description by Wu (1934) in the following particulars, although we could not examine type material or topotypic material from Luocheng Country, Guangxi Zhuang Autonomous Region: the epibranchial tooth is very sharp in the type material (Wu, 1934) versus blunt in the specimens examined by Türkay & Dai (1997), and the G1 terminal segment has a bent tip in the type material (Wu, 1934) versus straight terminal segment in the specimens examined by Türkay & Dai (1997) (see also Table 1). Furthermore, the specimens examined by Türkay & Dai (1997) were said to be collected from Xiushui County, Guangxi. The locality record seems to be incorrect, because Xiushui County is in Jiangxi, not in Guangxi. Dai (1999) corrected the locality of the specimens studied by Türkay & Dai (1997) to Xiuren County, Guangxi, and we believe that this is correct locality. We concur with Naruse, Zhu & Zhou (2013) that the specimens referred to H. pupureomanualis by Türkay & Dai (1997) could actually represent an undescribed species. This matter can only to be addressed when the specimens can be re-examined.
With a few exceptions, most Chinese freshwater crab genera have restricted distributional ranges. The geographical distribution pattern of many freshwater crabs is caused by geographical isolation, which is due to the limited dispersal abilities of freshwater crabs and low fertility (Yeo et al., 2008; Ji et al., 2016; Jia et al., 2018). The distribution pattern of freshwater crabs in mainland China is closely related to the regional differentiation of China’s natural geographical environment. Heterochelamon has been reported to be mainly distributed in northern Guangxi (Türkay & Dai, 1997; Naruse, Zhu & Zhou, 2013). Noteworthy in this paper is that we describe a new species assigned to the genus from southern Guangdong, which is not in the known range of the genus, suggesting that it has a wider distributional pattern than previously thought. That being said, the distribution of Heterochelamon is still within the Huanan freshwater zoogeographical province, which mainly consists of Guangxi, Guangdong and Hainan (Huang, Ebach & Ahyong, 2020). Within this zoogeographical province, there are relatively few abiotic factors such has high mountains and rivers that may obstruct the dispersal of freshwater crabs. We therefore believe that there are other new species of Heterochelamon in this zoogeographical province.
Conclusions
This study describes two new species of freshwater potamid crabs, referred to the genus Heterochelamon. We obtained sequence data of the mitochondrial 16S rRNA gene from the two new species and two other congeneric species for genetic analysis. The analysis supports the recognition of the two new species and the monophyly of the genus. The known distribution of this genus has been heretofore restricted to northern Guangxi, but our study now shows that the genus also occurs in southern Guangdong. (GenBank: MN823645 to MN823652).