The first pterosaur basihyal, shedding light on the evolution and function of pterosaur hyoid apparatuses
- Published
- Accepted
- Received
- Academic Editor
- Fabien Knoll
- Subject Areas
- Evolutionary Studies, Paleontology, Zoology
- Keywords
- Pterosaurs, Basihyal, Hyoid apparatus, Evolution, Function, Yixian Formation, China
- Copyright
- © 2020 Jiang et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2020. The first pterosaur basihyal, shedding light on the evolution and function of pterosaur hyoid apparatuses. PeerJ 8:e8292 https://doi.org/10.7717/peerj.8292
Abstract
The pterosaur is the first known vertebrate clade to achieve powered flight. Its hyoid apparatus shows a simplification similar to that of birds, although samples of the apparatus are rare, limiting the ability to make an accurate determination. In this study we reveal a new pterosaur specimen, including the first definite basihyal. Through the comparison of pterosaur hyoids, a trend has been discovered for the shortened hyoid relative to the length of the skull, indicating a diminished role of lingual retraction during the evolution of the pterosaur. The new material, possibly from a gallodactylid Gladocephaloideus, represents one of the least effective lingual retractions in all pterosaurs. Based on the structure of an elongated ceratobranchial and retroarticular process on mandibles, the function of the Y-shaped istiodactylid tongue bone is similar to those of scavenger crows rather than chameleons, which is consistent with the interpretation of the scavenging behavior of this taxon. More fossil samples are needed for further study on the function of other pterosaur hyoids.
Introduction
The pterosaur is the first known vertebrate to have obtained the ability of powered flight. The hyoid apparatus of the pterosaur shows a simplification similar to that in birds, in which the muscles play an important role in feeding and respiration (Li, Zhou & Clarke, 2018). The elaboration of the hyoid in pterosaurs is largely unknown because preserved fossils are rare despite widespread descriptions of them (Wellnhofer, 1970; Wellnhofer, 1975; Wellnhofer, 1978; Wellnhofer, 1985; Wellnhofer & Kellner, 1991; Wang & Lü, 2001; Wang et al., 2005; Lü, Xu & Ji, 2008; Padian, 2008a; Padian, 2008b; Dalla Vecchia, 2009; Jiang & Wang, 2011a; Cheng et al., 2012; Wang et al., 2012; Jiang et al., 2014; Lü, 2015).
In general, the ossified bony elements known as the hyoid that are preserved in pterosaur specimens are slender, rod-like ceratobranchial I that vary in length, curvature, and expansion of both tips (Dalla Vecchia, 2009). Williston (1903) considered the proatlas of Nyctosaurus (FMNH P 25026) as a basihyal. This interpretation was accepted in the main text but overturned in a note at the end of the monograph by Bennett (2001). Hence, except for two ceratobranchials, the other elements of hyoid apparatus have never been reported in pterosaurs.
A unique type of Y-shaped ceratobranchial has been reported in Ludodactylus (SMNK PAL 3828) (Frey, Martill & Buchy, 2003) and Liaoxipterus (JLU CAR-0018) (Lü, Xu & Ji, 2008). The extremely elongated and fused anterior parts were used to support a lingual function similar to that in chameleons (Lü, 2015). However, this understanding is challenged based on the new observation of the pterosaur hyoids.
In this study we describe a novel material found in the upper part of the Yixian Formation of Jingangshan locality, where three exceptionally preserved pterosaur and avian embryos were discovered in Jehol Biota (Wang & Zhou, 2004; Ji et al., 2004; Zhou & Zhang, 2004). This new specimen contained the first and well-preserved basihyal that had not been previously reported in any other pterosaur. We are able to approximate the hyoidean evolution in pterosaurs and discuss the function of some special types of hyoid apparatuses based on the newly discovered hyoid apparatus.
Materials and Methods
The newly discovered specimen (IVPP V 14189) has incomplete mandibular rami with a nearly complete hyoid apparatus and soft tissue impressions. The material was collected from a local farmer in the beginning of the 21st century and was prepared by a professional technician in IVPP. Two istiodactylid specimens were examined under a microscope; one is the holotype of Nurhachius ignaciobritoi (IVPP V 13288), whose hyoid was further prepared and first reported here, the other is the holotype of Liaoxipterus brachyognathus (JLU CAR-0018).
Further non-invasive observations were made using three-dimensional images of the outer and inner structures of the regions of interest. However, microtomography was not suitable in this case due to the shape of the specimen, which is composed of two broad and relatively thin plates (ca. 30 × 30 ×0.8 cm3). Micro X-ray computed laminography (Micro-CL) was conducted in the Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences. Image segmentation and visualization were performed using the software VGStudio version 2.2 and Mimix, version 17.0.
Results
Description of the new material
The novel fossil specimen was preserved in a dark grey slab of shale, with black bones and soft tissue impressions (Fig. 1). The preserved mandibular rami did not have any tooth or alveolus, indicating that it was from the posterior portion. The posterior boundaries of both rami were formed by breakage, and the articular was missing. The preserved elements of the mandible were fused quite well without any suture and only the rod-like angular can be distinguished in the medial view of the right ramus. Although a portion of the outer surface of the sample is missing, the rest is smooth with some grooves along the long axis of the mandible on the ventral margins. The preserved lengths of the left and right mandibular rami are 211.0 mm and 175.3 mm, respectively. The depth of the left ramus is 22.4 mm without apparent changes, while the right ramus slightly tapers, from 21.6 mm to 17.3 mm. The different depths of the two sides are likely caused by the different portion of the ramus or by the taphonomy, with the latter explanation preferred.
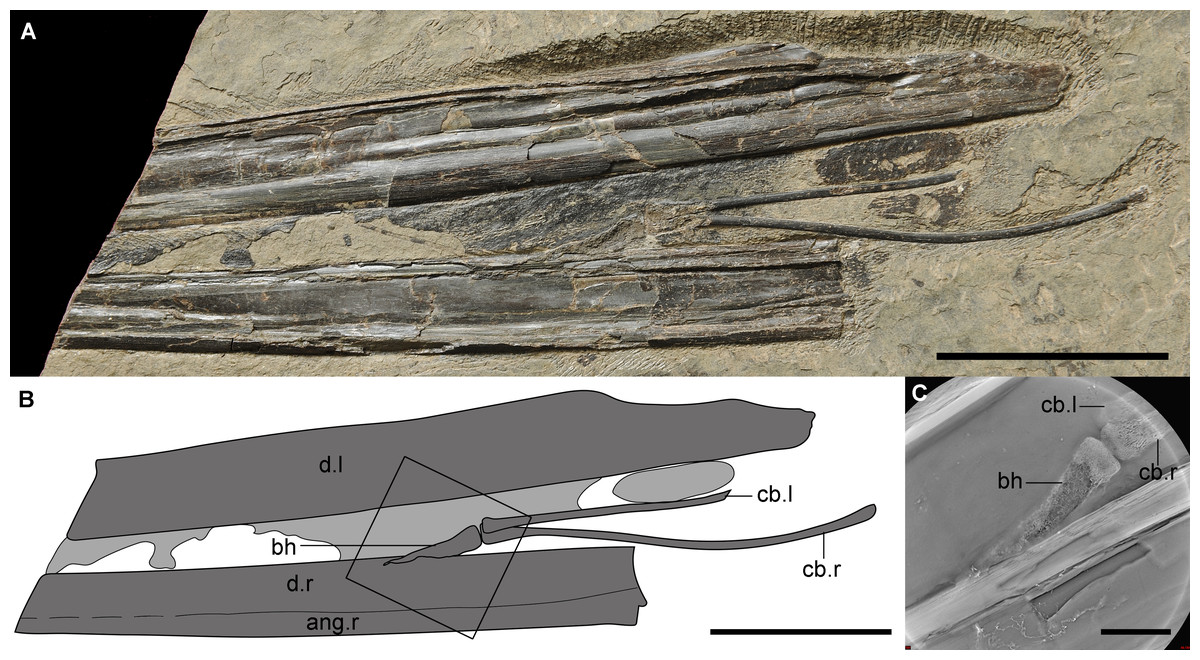
Figure 1: The new material (IVPP V 14189) of a unique hyoid apparatus in pterosaurs from the upper part of Yixian Formation.
(A) The photograph (Photo credit: Jie Zhang), scale bar, 50 mm; (B) the line drawing, potential soft tissue shown in light grey, scale bar, 50 mm; (C) the Micro-CL image of basihyal, close-up of the frame in B, scale bar, 10 mm. Abbreviation: ang, angular; bh, basihyal; cb, ceratobranchial; d, dentary; l, left; r, right.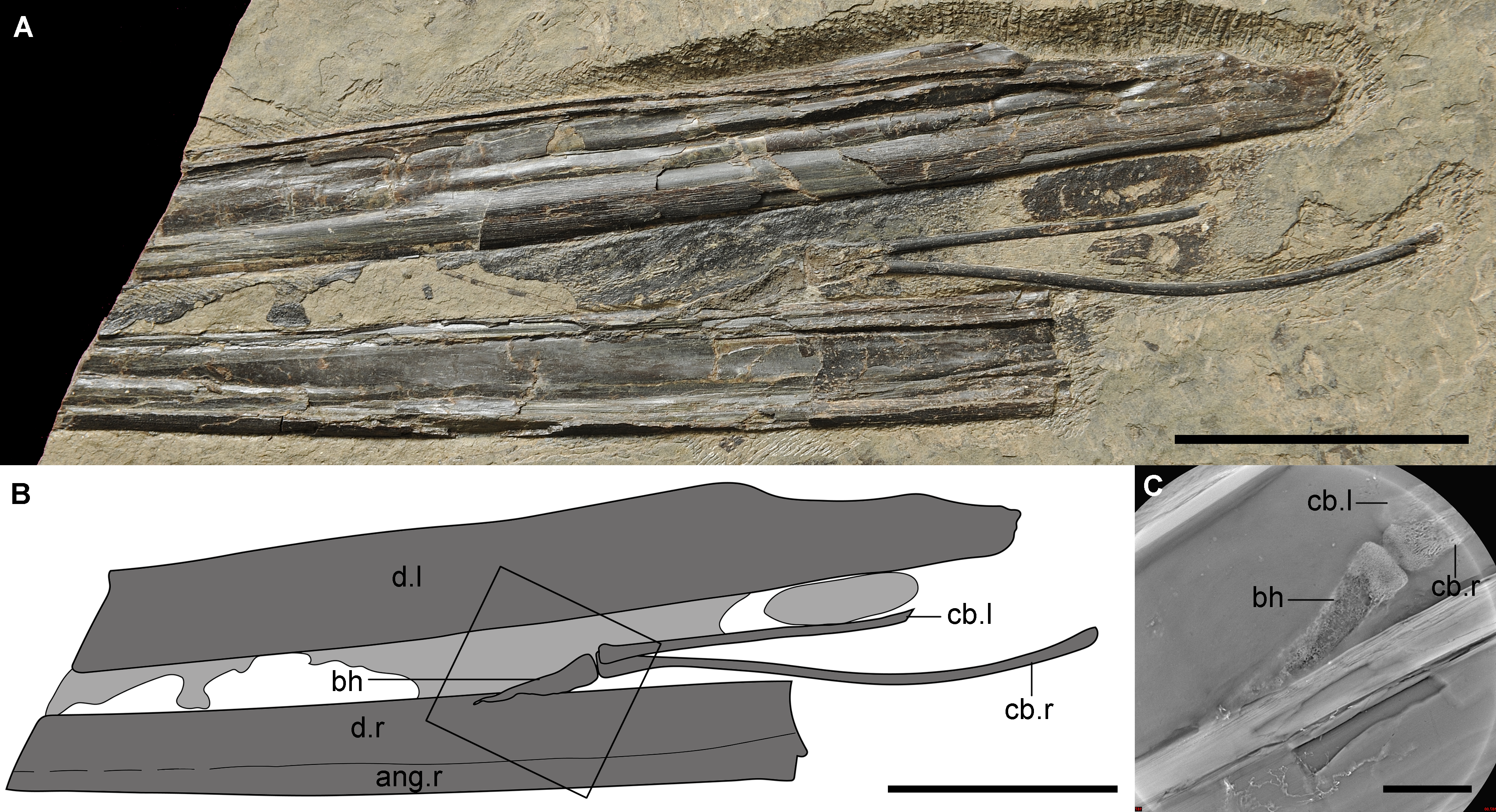{kind=link}
The hyoid apparatus includes a pair of ceratobranchial I and a middle-line element, the basihyal. The ceratobranchials are two thin, rod-like bones. The right one was complete with an expanded anterior tip (4.6 mm width) that was larger than the posterior expansion (3.0 mm width). According to the smaller posterior expansion, the possible existence of epibranchials was inferred, which has not been reported in any pterosaur; epibranchials may be similar to the cartilage seen in some modern birds. The length of the right ceratobranchial between both tips was 106.2 mm, and the width was 2.1 mm. There were shallow grooves, observable by microscope, found on the entire surface but no origin or insertion of any muscles were detected. The right ceratobranchial was slightly curved upward at the middle portion. The basihyal was an elongated triangular element, with a maximal length of 29.2 mm and a width of 6.1 mm at the base. The basihyal seemed to be twisted, indicating that it was potentially cartilaginous, or possibly cartilage that was not fully ossified, which was supported by Micro-CL imaging (Fig. 1C). The posterior aspect of the basihyal was flat as the anterior ones of ceratobranchials, inferring that the articulations between the basihyal and ceratobranchials were not flexible. The anterior end of the basihyal was pointed, inferring that the paraglossal was nonexistent, which is common in neognath birds and complicated in paleognaths (Li, Zhou & Clarke, 2018). This phenomenon has only been reported in one taxon of dinosaur (Hill et al., 2015), but was not preserved in extant crocodilians and turtles (Li & Clarke, 2015).
Identifying the new material
The mandibular rami were partially preserved, which was identified as the toothless caudal segment. Two pterosaur taxa have been reported so far in the Jingangshan locality, namely the gallodactylid Gladocephaloideus jingangshanensis (Lü et al., 2012), and a questionable Yixianopterus jingangshanensis, in which most of the cranial bone was artificially reconstructed (Lü et al., 2006). The new material would be best assigned to Gladocephaloideus rather than Yixianopterus due to the location in which it was found. The depth of the posterior end of the new material was four times large as that of the holotype of Gladocephaloideus, which indicates that the sizes are quite different between these two specimens. The holotype was considered to be an adult individual (Lü et al., 2012), but was later reconsidered to be a late juvenile or sub-adult individual (Lü, Kundrát & Shen, 2016). It is reasonable to believe that the new material is from an adult G. jingangshanensis and is quite larger than the holotype.
Unpublished collections from the same location housed at IVPP (IVPP V 12695A&B, V 13335, V 14235, V 14427, and V 26616) reveal some pterosaurs with different characteristics from Gladocephaloideus, suggesting the diversity of pterosaurs in this vicinity. The new material can be excluded as being from istiodactylids and Guidraco because of the different shapes of the hyoids. The length/depth ratio of the preserved toothless mandibular rami in the new material was 9.4, and the accurate value should be even larger because of the incompleteness of the sample, while the ratios of tapejarids, chaoyangopterids, anurognathid, boreopterids, anhanguerids, and some other pteranodontoids (sensu Kellner, 2003) found in Jehol Biota were less than 10; only ctenochasmatid and gallodactylid pterosaurs have a greater ratio than this value. The ratios are 10.6 and 10.2 in Gegepterus and Forfexopterus, respectively (Wang et al., 2007; Jiang et al., 2016). These are the only two published ctenochasmatids with nearly complete mandibles from the Jehol Biota. The incompleteness of the new sample and the slightly larger ratios in ctenochasmatids lead us to believe that G. jingangshanensis, a gallodactylid pterosaur, is the best available interpretation.
Discussion
The hyoidean evolution in pterosaurs
Several phylogenetic analyses have been carried out on pterosaur to date (Kellner, 2003; Unwin, 2003; Wang et al., 2009; Lü et al., 2010; Andres, Clark & Xu, 2014; Codorniú et al., 2016; Wang et al., 2017; Vidovic & Martill, 2017), although no consensus on the evolution of the pterosaur has been reached. The evolution from long-tailed taxa to short-tailed one is commonly accepted; however, this study does not address the comparisons made between the differences in published phylogenetic results. The phylogenetic results obtained from previously conducted research is included in this study done by some authors before is chosen here (Wang et al., 2017), which was also on some previous studies (Kellner, 2003; Wang et al., 2009). Some modifications were made for the discussion of hyoidean evolution (Fig. 2).
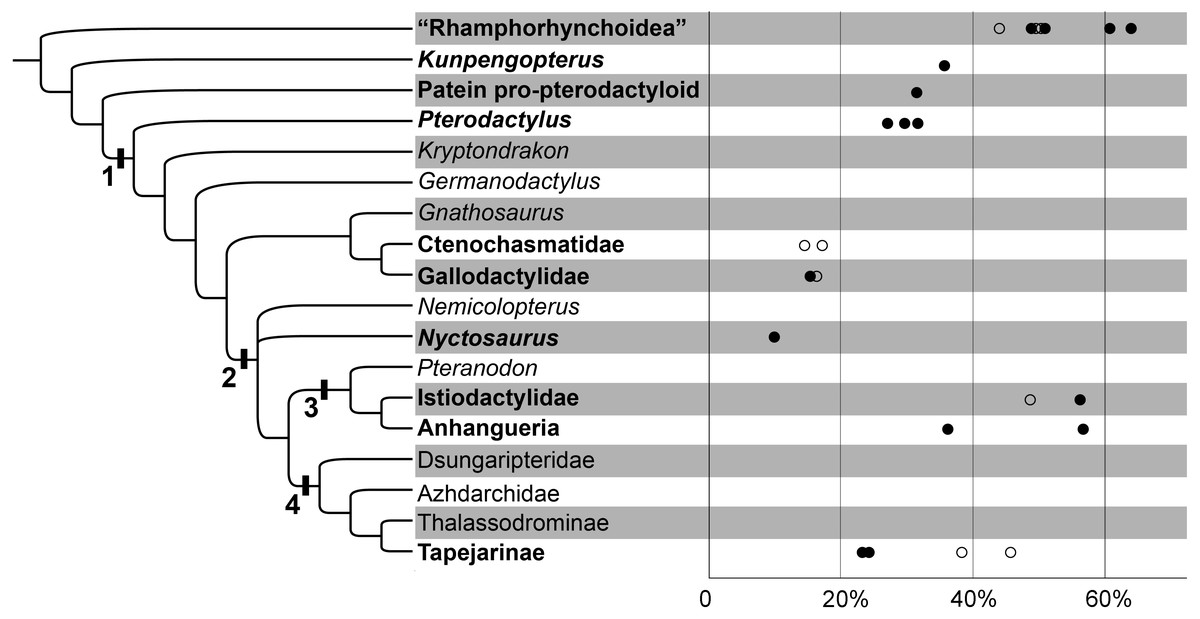
Figure 2: The variability of the ceratobranchial relative to the skull in pterosaurs.
The phylogenetic tree after Wang et al., 2017. 1. Pterodactyloidea; 2. Dsungaripteroidea; 3. Pteranodontoidea; 4. Tapejaroidea. The group of pterosaurs with hyoids are in bold. The horizontal axis indicates the ceratobranchial/skull length ratio; the circles indicate the estimated ones; details can be seen in Table 1.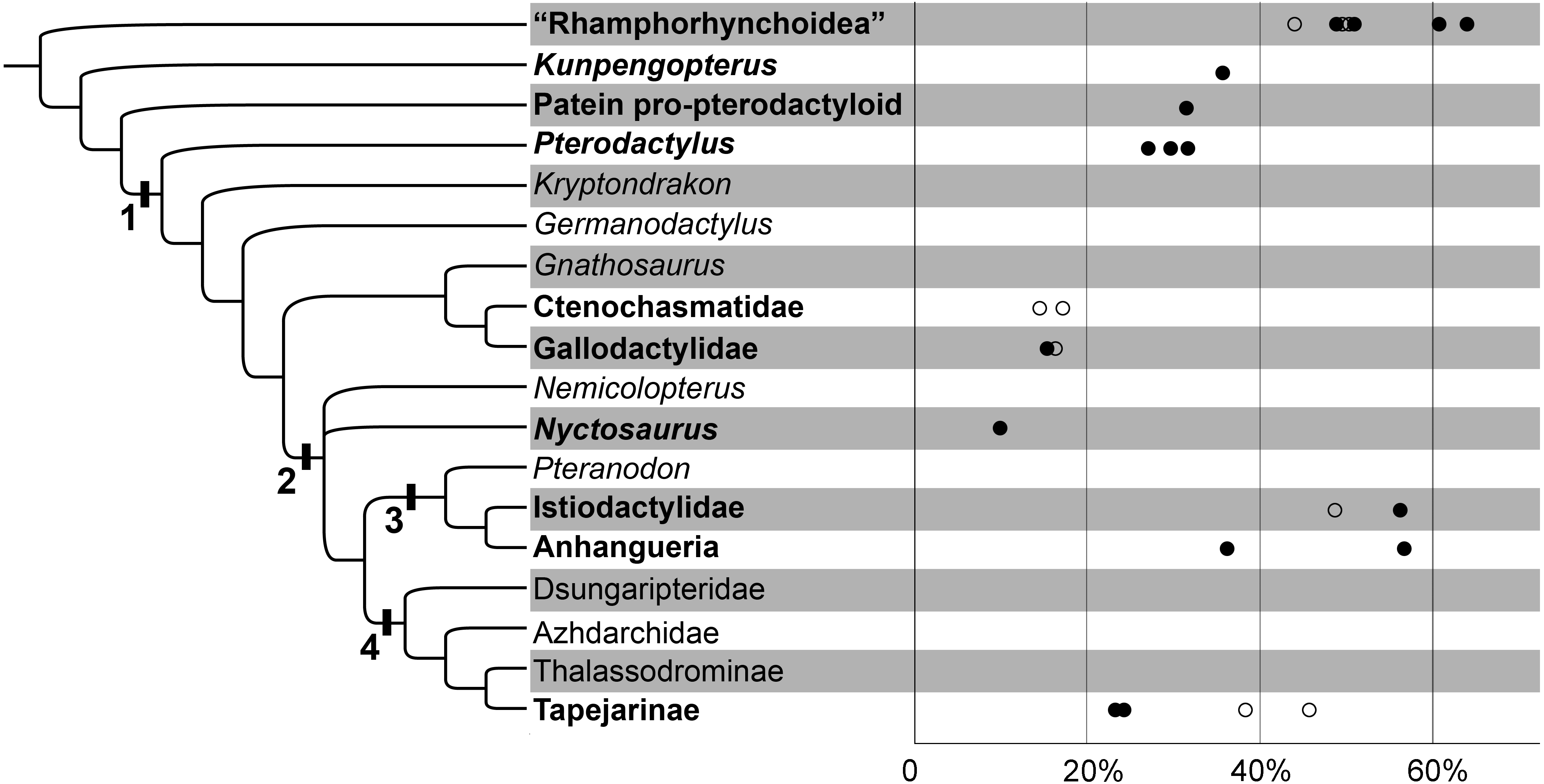{kind=link}
| Taxa | Ceratobranchial/ skull length ratio | Sources | Comments |
|---|---|---|---|
| Eudimorphodon ranzii (MCSNB 2888) | >30% | Zambelli, 1973; Paganoni, 2003 | incomplete hyoid |
| Austriadactylus cristatus (SMNS 56342) | ∼44% | Dalla Vecchia et al., 2002 | posterior most missing |
| Dorygnathus banthensis (SMNS 50914) | >34.0% | Padian, 2008a | posterior part missing |
| Dorygnathus banthensis (SMNS 50702) | 63.8% | Padian, 2008a | |
| Dorygnathus banthensis (SMNS 51827) | >30.5% | Padian, 2008a | anterior and posterior parts missing |
| Scaphognathus crassirostris (SMNS 59395) | 50.8% | Bennett, 2014 | |
| Scaphognathus crassirostris (GPIB 1304) | ∼49.4% | Bennett, 2014 | anterior most missing |
| Jianchangnathus robustodens (IVPP V 16866) | >37.6% | Cheng et al., 2012 | anterior part missing |
| Fenghuangopterus lii (CYGB-0037) | 60.70% | Lü et al., 2010 | |
| Rhamphorhynchus muensteri (BSP AS VI 34) | >36.3% | Wellnhofer, 1975 | anterior part missing |
| Rhamphorhynchus muensteri (BSP 1927 I 36) | ∼50.5% | Wellnhofer, 1975 | anterior most missing |
| Darwinopterus robustodens (HGM 41HIII-0309A) | 49.1% | Lü et al., 2011 | |
| Darwinopterus modularis (ZNHM M8782) | >25% | Lü et al., 2010 | anterior part missing |
| Kunpengopterus sinensis (IVPP V 23674) | 35.7% | Cheng et al., 2017 | |
| Painten pro-pterodactyloid | 31.4% | Tischlinger & Frey, 2013 | |
| Pterodactylus antiquus (BSP 1883 XVI 1 / MCZ 1505) | 29.6% | Wellnhofer, 1970; Bennett, 2013 | |
| Pterodactylus kochi (BSP 1975 I 221) | 31.6% | This paper | |
| Pterodactylus (JME-SOS 4008) | 27.0% | This paper | |
| Gegepterus changae (IVPP V 11981) | ∼14.5% | Wang et al., 2007 | skull length estimated |
| Pterofiltrus qiui (IVPP V 12339) | ∼17.2% | Jiang & Wang, 2011b | anterior most missing |
| Feilongus youngi (IVPP V 12539) | 15.4% | Wang et al., 2005 | |
| Gladocephaloideus jingangshanensis (IG-CAGS-08-07) | ∼16.4% | Lü et al., 2012 | anterior most missing |
| Nyctosaurus (FMNH P 25026) | 10% | Williston, 1902; Williston, 1903; Wellnhofer, 1978 | |
| Nurhachius ignaciobritoi (IVPP V 13288) | >39.7% | Wang et al., 2005 | posterior part missing |
| Liaoxipterus brachyognathus (JLU CAR-0018) | ∼48.6% | Lü, Xu & Ji, 2008 | skull length estimated from similar taxa |
| Istiodactylid (BHGL-036) | 56.2% | This paper | |
| Ludodactylus sibbicki (SMNK PAL 3828) | 56.6% | Frey, Martill & Buchy, 2003 | |
| Guidraco venato (IVPP V 17083) | >37.4% | Wang et al., 2012 | anterior part missing |
| Haopterus gracilis (IVPP V 11726) | 36.1% | Wang & Lü, 2001 | |
| Anhanguera araripensis (BSP 1982 I 89) | >22% | Wellnhofer, 1985 | posterior part missing |
| Ikrandraco avatar (IVPP V 18406) | >16.9 | Wang et al., 2014 | posterior part missing |
| Sinopterus sp. (IVPP V 20249) | 23.3% | This paper | |
| Sinopterus sp. (IVPP V 26617) | 24.3% | This paper | |
| Europejara olcadesorum (MCCM-LH 9413) | ∼45.8% | Vullo et al., 2012 | nearly complete |
| Tapejara wellnhoferi (AMNH 24440) | ∼38.3% | Wellnhofer & Kellner, 1991 | nearly complete |
In most birds, the hyoid apparatus includes two rod-like ceratobranchials, two partially cartilaginous curved epibranchials, a small middle-line basihyal and urohyal, and a pair of cartilaginous paraglossa (Homberger, 2017). In pterosaurs, the ceratobranchials are similar; the basihyal is only preserved in the new specimen, and the other elements are absent. Based on the anterior and posterior expansions of the ceratobranchial tips, the basihyal should be more common than the condition in preserved fossils and the epibranchials likely existed in some pterosaur taxa. There is no evidence for the urohyal or paraglossal.
The hyoids of primitive non-pterodactyloids only include the preserved ceratobranchials; this rod-like element is slender and quite long relative to the skull length (Figs. 2; 3A & 3B; Table 1). The ceratobranchial/skull length ratios are 63.8%, 50.8%, and 49.1% in Dorygnathus banthensis (SMNS 50702) (Padian, 2008b), Scaphognathus crassirostris (SMNS 59395) (Bennett, 2014), and Darwinopterus robustodens (HGM 41HIII-0309A) (Lü et al., 2011), respectively. A similar condition was found in Austriadactylus cristatus (SMNS 56342) and Rhamphorhynchus muensteri (BSP 1927 I 36), although the ceratobranchials are slightly incomplete. The ceratobranchial/skull length ratio is similar to most extant reptiles (Li & Clarke, 2015). In some non-pterodactyloids, the anterior end expanded, referring the presence of the basihyal, represented by Carniadactylus rosenfieldi (MFSN 1797) (Elsheikh & Al-Zahaby, 2014), Scaphognathus crassirostris (SMNS 59395) (Bennett, 2014), and Darwinopterus robustodens (HGM 41HIII-0309A) (Lü et al., 2011). The basihyal probably appeared in the primitive pterosaur from the Late Triassic, although this newly discovered sample from the late Early Cretaceous period is the only basihyal in pterosaurs and it is absent from the fossil records due to its cartilaginous bone tissue. The ceratobranchials are gently curved, except in some species in which it is nearly straight. For example, three specimens of Dorygnathus banthensis are straight without any bend (Fig. 3A) (Padian, 2008b), nearly excluding the influence of the taphonomy.

Figure 3: Hyoid apparatuses in pterosaurs.
(A) Dorygnathus banthensis SMNS 50914 (Photo credit: Taissa Rodrigues); (B) Scaphognathus crassirostris SMNS 59395 (Photo credit: Taissa Rodrigues); (C) Kunpengopterus sinensis, IVPP V 23674; (D) Pterodatylus, JME-SOS 4008 (Photo credit: Rui Qiu); (E) Feilongus youngi, IVPP V 12539; (F) Gegepterus changae, IVPP V 11972; (G) Haopterus gracillis, IVPP V 11726; (H) Ikrandraco avatar, IVPP V 18199; (I) Liaoxipterus brachyognathus, JLU CAR-0018, close-up of the anterior part of hyoid; (J) & (K) Ludodactylus sibbicki, SMNK PAL 3828 (Photo credit: Alexander. W.A. Kellner), K, close-up of the anterior part of hyoid; (L) & (M) Nurhachius ignaciobritoi, IVPP V 13288, M, close-up of the anterior part of hyoid; (N) Guidraco venator, IVPP V 17033; O, Tapejara wellnhoferi, AMNH 24440 (Photo credit: Alexander W.A. Kellner). The anterior ends to the left side, and E, G, J, K, N, & O shown inverted. Scale bars, 10 mm.{kind=link}
The ceratobranchial/skull length ratios have a decreasing trend from long-tailed pterosaurs to short-tailed Pterodactyloidea (Fig. 2; Table 1). In Kunpengopterus (Fig. 3C), Paiten pro-pterodactyloid, and Pterodactylus (Fig. 3D), the ceratobranchials shortens to approximately 30% of the skull length (Cheng et al., 2017; Tischlinger & Frey, 2013; Bennett, 2013; Wellnhofer, 1970), while the ratios dropped to less than 20% in the gallodactylids Gladocephaloideus (Lü et al., 2012) and Feilongus (Fig. 3E) (Wang et al., 2005), as well as in ctenochasmatid pterosaurs, such as Gegepterus (Fig. 3F) (Wang et al., 2007; Jiang & Wang, 2011a), and Pterofiltrus (Jiang & Wang, 2011b).
Pteranodontoid (sensu Kellner, 2003) pterosaurs display the unexpanded anterior ends of ceratobranchials, suggesting the absence of the basihyal in Istiodactylidae and Anhangueria (Figs. 3H–3N). The decreasing trend was reversed in istiodactylids and Ludodactylus sibbicki (SMNK PAL 3828), whose ceratobranchials are quite elongated and slightly tapered forward (Frey, Martill & Buchy, 2003; Lü, Xu & Ji, 2008). The anterior fusion of ceratobranchials was considered to be the lingual process in Ludodactylus and Liaoxipterus (Lü, 2015). The unfused two ceratobranchials can be identified by the observation of the holotype of Liaoxipterus (Fig. 3I), and the condition is also seen in the photos of Ludodactylus (Figs. 3J & 3K). The further prepared holotype of Nurhachius (IVPP V 13288), which is the first specimen of istiodactylids from China, also shows the unfused ceratobranchials with the anterior portions close to each other (Figs. 3L & 3M). Therefore, no fusion of the ceratobranchials existed in pterosaurs, contrary to what has been described in earlier literature (Lü, 2015). Guidraco is closely related to Ludodactylus and has only one elongated ceratobranchial (Fig. 3N) (Wang et al., 2012), which may also have the Y-shaped ceratobranchials.
The information on hyoids in Tapejaroidea (sensu (Kellner, 2003) is limited to three taxa in Tapejaridae: Europejara, Sinopterus, and Tapejara (Fig. 3O). These three taxa, in a monophyletic group, shared a similar shape of ceratobranchials, but with varieties in their lengths relative to their skull (Fig. 2; Table 1). Europejara (MCCM-LH 9413) is the greatest at 45.8% (Vullo et al., 2012) and Sinopterus is the least at 25%.
The function of some pterosaur hyoids
There is a trend in the Pterosauria showing that the ceratobranchials are shortened relative to their skull length (Fig. 2, Table 1). The structure of the pterosaur hyoid apparatus is simplified as in birds, with elongated ceratobranchials. In birds, there are a pair of muscles that control the protraction and retraction of the tongue. M. branchiomandibularis (M.bm) originates from the mandibles and is inserted on the posterior end of ceratobranchials and the anterior end of the epibranchials, which active in tongue retraction (Homberger & Meyers, 1989). M. serpihyoideus (M.sh) and M. stylohyoideus (M.st) originate from the retroarticular process of the mandible and insert on the anterior of the basihyal and ceratobranchials, rarely on the paraglossal, and it can protract the tongue (Homberger & Meyers, 1989). It is assumed that pterosaurs had similar lingual muscles to those of birds, with a similar lifestyle and a simplified hyoid apparatus, which is a reasonable assumption as birds are the closest extant group to the pterosaur in most phylogenetic results (Witton, 2013). The M.bm is inserted on the posterior end of the ceratobranchial whether epibranchials are present in the pterosaur or not. If the origin of the M.bm does not change, the length of M.bm will be the same (Fig. 4), indicating that protraction is maintained when the ceratobranchials shorten. However, the lengths of the M.sh and M.st were shortened (Fig. 4), indicating that there was a decreased ability to retract the tongue and a lingual inability to transport food during the evolution of the pterosaur. The new material, which is presumed to be gallodactylid Gladocephaloideus, should have an elongated upper and lower jaw, with a short tooth row (Lü et al., 2012). The length of the ceratobranchials in the new material was one of the shortest relative to the skull in all known pterosaurs and the ability of lingual transport of food may have been the least effective.

Figure 4: Graphic models of lingual muscles of Haopterus and Liaoxipterus in dorsal view.
(A) Haopterus; (B) Liaoxipterus. Red lines indicate the inferred M. sh and M. st, and blue lines indicate the inferred M.bm. The arrows indicate direction of movements and forces. The assumed epibranchials and basihyal are shown in grey within the dashed lines. Abbreviation: bh, basihyal; cb, ceratobranchial; ep, epibranchial; M.bm, M. branchiomandibularis; M. sh, M. serpihyoideus; M. st, M. stylohyoideus. Scale bar, 10 mm.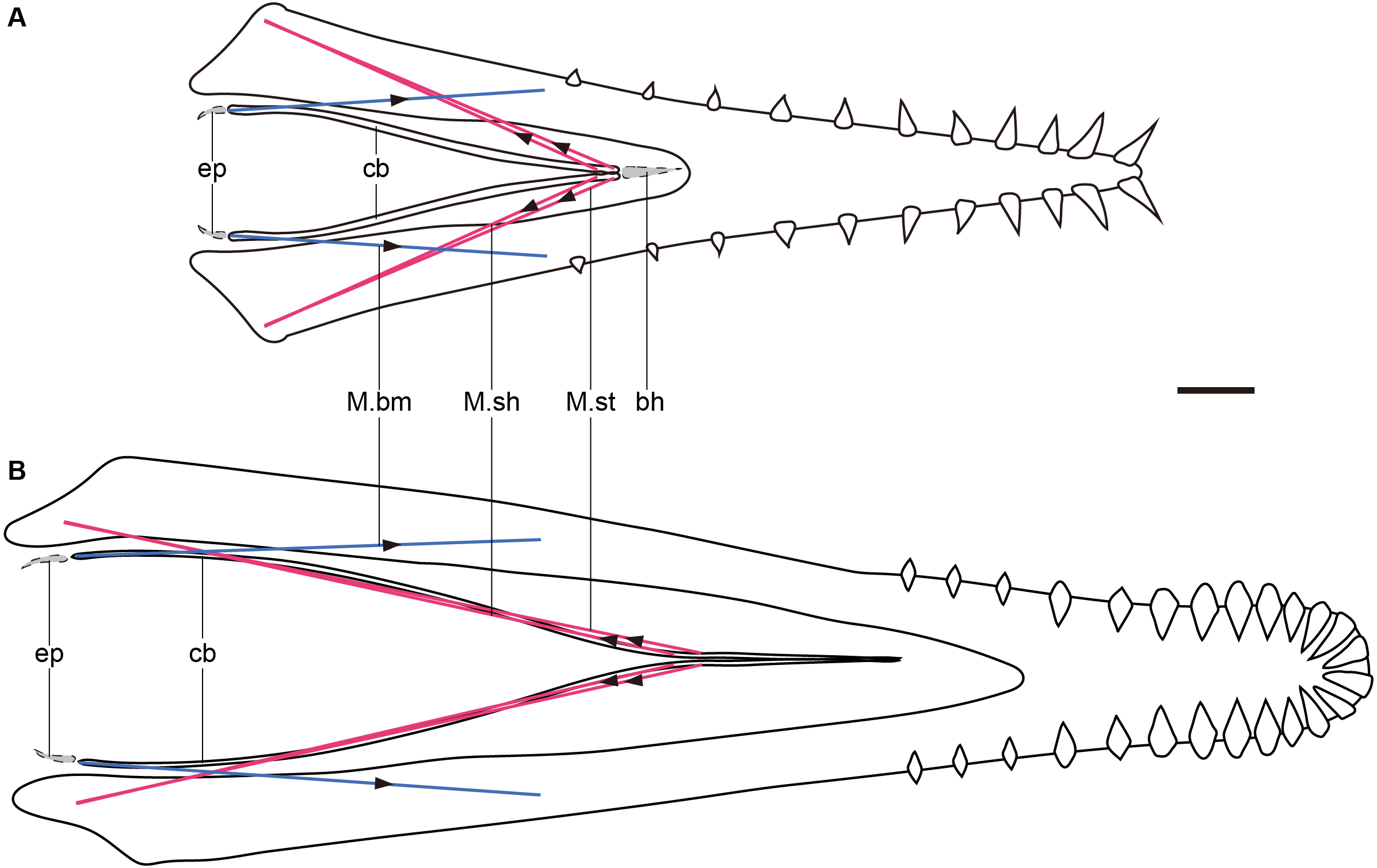{kind=link}
The elongated Y-shaped hyoid apparatuses (Fig. 4B), represented by istiodactylids, reversed the trend of the length decrease, which suggests the essential function of the tongue. Lü (2015) considered the fused (although actually unfused) anterior part of ceratobranchials as the lingual process reported in chameleons, along which the tongue slides during launch. In the prey processing of chameleons, the soft tissue of the tongue projects out of the mouth along with the hyoid element; the entoglossal process moves forward relative to the ceratobranchials and ceratohyals of the hyoid apparatus (Herrel et al., 2001). In istiodactylids, the anterior process, made up of the anterior of ceratobranchials, cannot move forward relative to the rest of the ceratobranchials, indicating the limited anteroposterior movement of the tongue. Therefore, this Y-shaped hyoid is different from that of chameleons, and istiodactylids were likely not insectivorous as was suggested in the previous literature (Lü, 2015). However, this specialized type of hyoid represents some other function of their tongues in feeding or respiration. A careful examination of this Y-shaped hyoid reveals a slight expansion in the corner of the bend in some specimens, which might be near the posterior end of the adhered anterior parts. When compared with avian muscles, this expansion is probably the insertion of M.st, which originates from the lateral and ventral aspects of the caudal mandible, primarily on the retroarticular process (Homberger & Meyers, 1989; Li & Clarke, 2016). This process is more elongated in istiodactylids than that of most other pterosaurs (Witton, 2012). The elongation of the ceratobranchials and the enlarged area for the muscular attachment indicates the development of this muscle, as well as the enhanced ability to retract the tongue, relative to powerful food transport. In some extant crows (Corvus), which are scavengers, the anterior part of the tongue can protrude out of the oropharyngeal cavity and transport food during the retraction, which is an adaption necessary for the food collection (Igwebuike & Eze, 2010; Elsheikh & Al-Zahaby, 2014). The tongue of istiodactylids may not have needed to protrude out like that of crows, because they may have used their diamond-shaped teeth to cut the food into pieces. The only elongated ceratobranchials also provide the tongue firmness as well as the bone function in crows. Therefore, istiodactylids were probably scavengers, as suggested in the previous literature (Witton, 2012). Ludodactylus also had the Y-shaped hyoid, which might have similar functions to istiodactylids. However, the presumed Y-shaped hyoid pterosaur, Guidraco, has exaggerated anterior teeth unfitting for a scavenger and the hyoid function requires further discussion.
Conclusion
The new fossil material revealed a novel type of hyoid apparatus and demonstrations the first basihyal in pterosaurs, revealed the hyoid morphology previously unseen. The hyoids in pterosaurs have a trend to shorten relative to the length of skulls, indicating the diminished role in lingual transport of food from the non-pterodactyloids to the pterodactyloids. The new gallodactylid, Gladocephaloideus, represents one of the least effective lingual retractions in all pterosaurs. Based on the elongated ceratobranchials and shape of the retroarticular process, the function of the Y-shaped istiodactylid tongue is unlike that of chameleons but closer to that of scavenging crows, which is consistent with the interpretation of the scavenging behavior in pterosaurs. More fossil samples are needed for further study on the function of other hyoids.