
Inconsistent response of soil bacterial and fungal communities in aggregates to litter decomposition during short-term incubation
- Published
- Accepted
- Received
- Academic Editor
- Douglas Burns
- Subject Areas
- Microbiology, Soil Science
- Keywords
- Soil bacterial and fungal diversity, Soil aggregate, Litter decomposition
- Copyright
- © 2019 Li and Yang
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2019. Inconsistent response of soil bacterial and fungal communities in aggregates to litter decomposition during short-term incubation. PeerJ 7:e8078 https://doi.org/10.7717/peerj.8078
Abstract
Background
Soil aggregate-size classes and microbial communities within the aggregates are important factors regulating the soil organic carbon (SOC) turnover. However, the response of soil bacterial and fungal communities in aggregates to litter decomposition in different aggregate-size classes is poorly understand.
Methods
Soil samples from un-grazed natural grassland were separated into four dry aggregate classes of different sizes (2–4 mm, 1–2 mm, 0.25–1 mm and <0.25 mm). Two types of plant litter (leaf and stem) of Leymus chinensis were added to each of the four aggregate class samples. The CO2 release rate, SOC storage and soil microbial communities were measured at the end of the 56-day incubation.
Results
The results showed that the 1–2 mm aggregate had the highest bacterial Shannon and CO2 release in CK and leaf addition treatments, and the SOC in the <0.25 mm aggregate was higher than that in the others across the treatments. The relative abundance of Ascomycota was higher in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates in the treatment without litter addition, and the relative abundance of Aphelidiomycota was lower in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates. Also, litter addition increased the relative abundance of Proteobacteria and Bacteroidetes, but decreased the relative abundance of Acidobacteria, Gemmatimonadetes, and Actinobacteria. The relative abundance of Ascomycota and Aphelidiomycota increased by more than 10% following leaf litter addition. The bacterial Shannon index had a significantly positive and direct effect on SOC concentration and CO2 release, while the fungal Shannon index was significantly correlated with SOC concentration. Our results indicate that the soil bacterial diversity contributes positively to both carbon emissions and carbon storage, whereas soil fungal diversity can promote carbon storage and decrease carbon emissions.
Introduction
Litter decomposition is a key step in the carbon cycle (Bonan et al., 2013; Schmidt et al., 2011; Wieder, Bonan & Allison, 2013), especially in grassland ecosystems, which cover 40% of the earth’s land surface (Lu et al., 2017), and possess about 20% of the soil organic carbon (SOC) stocks globally (Schuman, Janzen & Herrick, 2002). The majority of studies have shown that litter decomposition usually depends on three main drivers: climate factors (soil moisture and temperature) (He et al., 2010; Riggs et al., 2015; Wang, Zeng & Zhong, 2016; Zhong et al., 2017), litter quality (i.e., its chemical composition) (Hishinuma et al., 2017; Zhang et al., 2016), and decomposer activity (Jia et al., 2016; Keiser & Bradford, 2017). In addition, there is increasing evidence that soil microbial communities influence litter decomposition rates over and above climate and litter quality controls (Bradford et al., 2016; Schimel & Schaeffer, 2012). Studies on litter decomposition have focused on the bacterial community composition and have confirmed the involvement of a series of colonizers in the litter decomposition process (Fanin & Bertrand, 2016). Soil fungi are also one of the major microbial decomposers of plant litter, being able to decompose the recalcitrant component (Liang et al., 2017).
Soil aggregates are the basic units of soil structure (Bronick & Lal, 2005) and are generally sub-divided into macro-aggregates (>0.25 mm) and micro-aggregates (<0.25 mm) (Yang, Liu & Zhang, 2017). Soil microbial community compositions are non-uniformly distributed in the soil aggregates and may be sensitive to changes in soil environments (Six et al., 2004). In general, the distribution of bacteria in soil varies with aggregate size (Neumann et al., 2013), and the microbial biomass associated with soil aggregates has been reported to be heterogeneously distributed (Wang, Li & Zheng, 2017). Only a few studies have used sequencing analysis to assess the bacterial communities of aggregates (Gupta & Germida, 2015). Although our previous study found the distribution of soil bacteria and fungi in aggregates to be inverse (Yang, Liu & Zhang, 2019), very little is known regarding their contribution to litter decomposition in different aggregate fractions. In addition, soil microbial processes are regulated by constraints in soil pH, which is considered to be an important factor controlling the balance of fungal to bacterial growth in the soil (Lauber et al., 2009; Rousk et al., 2010).
We designed a two-factor incubation experiment in the laboratory: one was litter quality (leaf and stem), while the other was soil aggregate size. The SOC mineralization, SOC content, soil pH, and soil microbial community were measured at the end of the incubation period. We hypothesized that (1) the soil bacterial and fungal communities are unevenly distributed in the aggregate fractions, (2) different aggregate sizes respond differently to the quality of the litter addition, and (3) the correlation between soil properties and soil microbial diversities controls litter decomposition.
Material and Methods
Material collection and preparation
The soil samples were collected from one type of ungrazed natural grassland located in Guyuan, Hebei Province, China (41°46′N, 115°41′E, elevation 1,380 m) in May of 2018, the initial stage of growing season. This area is a typical temperate zone characterized by a mean annual precipitation of 430 mm and a mean annual temperature of 1.4 °C. The minimum monthly mean air temperature is −18.6 °C in January and the site reaches a maximum of 21.1 °C in July. The site has a calcic-orthic Aridisol soil with a loamy-sand texture.
In brief, the top layer (0–15 cm) of the soil (∼200 kg) at one location was collected in plastic bags with a shovel, and was quickly transported to the laboratory by car, upon which the plant roots and leaves were carefully removed by hand and the soil was air-dried. The soil was sieved to separate large macro-aggregates (2–4 mm), macro-aggregates (1–2 mm), meso-aggregates (0.25–1 mm) and micro-aggregates (<0.25 mm) according to Yang, Liu & Zhang (2017). In brief, the undisturbed soil was shaken through four sieves (4, 2, 1 and, 0.25 mm) for 2 min, and the large macro-aggregates (2–4 mm) were collected from the two mm sieve, macro-aggregates (1–2 mm) from the one mm sieve, and meso-aggregates (0.25–1 mm) from the 0.25 mm sieve, and the micro-aggregates (<0.25 mm) were passed through the 0.25 mm sieve. Soil aggregates were stored hermetically at room temperature after until collecting the litter samples.
In September 2018, two types of plant litter (leaf and stem) were obtained from the dominant species (Leymus chinensis) in the same location from where the soil samples were collected. The litter was brought to the laboratory, and dried at 65 °C to constant weight. In order to avoid the effects of litter size on decomposition, the plant litter was cut into ca. 1-cm-long sections for the incubation experiment. Some basic characteristics for the soils in Table 1 were cited from our previous studies (Yang, Li & Zhang, 2019; Yang, Liu & Zhang, 2017; Yang, Liu & Zhang, 2019).
| Soil aggregates | Litter type | ||||||
|---|---|---|---|---|---|---|---|
| 2–4 mm | 1–2 mm | 0.25–1 mm | <0.25 mm | Leaf | Stem | ||
| SOC (g kg−1) | 13.27(0.1)ab | 11.87(0.2)b | 5.20(0.2)c | 14.17(0.3)a | TC (g kg−1) | 411.96(0.1)c | 424.19(0.1)a |
| TN (g kg−1) | 1.67(0.03)bc | 1.73(0.03)b | 1.07(0.07)c | 1.90(0.06)a | TN (g kg−1) | 17.20(0.1)a | 14.30(0.1)c |
| C/N ratios | 7.95(0.01)a | 6.86(0.01)b | 4.86(0.02)c | 7.46(0.01)a | C/N ratios | 23.95(0.1)c | 29.66(0.1)a |
Notes:
Different letters in the same row indicate a significant difference at P < 0.05 using least significant difference tests.
Experimental design and incubation study
The three replicates of air-dried soil samples (200 g dry weight) of each aggregate size class (2–4, 1–2, 0.25–1, and <0.25 mm) were placed at the bottom of 1,000 mL jars. The jars were new and unused, and we did not sterilize them beforehand because our incubator has the function of ultraviolet sterilization. The two plant litter types (3 g of dry matter) were combined with 200 g of dry soil in the microcosms. Although air drying of soil sample is not representative of the communities that originally existed in the soil, it can represent the difference in the distribution of microbes in our incubation conditions (Yang, Liu & Zhang, 2019). No litter addition was used as the control check (CK). There were a total of 72 microcosms (4 aggregate size × 3 litter types × 3 replications × 2 sampling times). The moisture content was adjusted to 30% by weighing each microcosm and adding distilled water, and 30% is the maximum field water capacity of the soil (Yang, Liu & Zhang, 2019). Each jar was covered with perforated cling film to reduce humidity loss while allowing gaseous exchange. The jars were pre-incubated for 3 days at a constant temperature of 25 °C. After the pre incubation period, the jars were maintained for 56 days at 25 °C. During the 56-day incubation, the soil moisture of each microcosm was maintained consistently by weighing each microcosm every week and adding distilled water. After 28 and 56 days of incubation, 36 microcosms were retrieved, respectively. Litter was removed from each microcosm, cleaned with water to remove adhering soil particles, dried (65 °C, 48 h) and weighed.
Soil aggregate respiration measurements
Soil aggregate respiration was measured at the end of the incubation. In brief, small vials with five mL of 1 M NaOH were placed in the incubation jars to trap CO2 for 24 h. The soil respiration (g CO2- C g−1 soil day−1) was estimated by titrating two mL NaOH from each trap with 0.1 M HCl after adding two mL 1 M BaCl2 (1:1) and a phenolphthalein indicator using a Digital Burette continuous E (VITLAB, Grossostheim, Germany) according to Yang et al. (2018). At the end of the soil respiration incubation, 10 g of soil sample was collected immediately after the removal of the plant litter and stored at −80 °C for microbiological sequencing. The remaining soil samples were air-dried for SOC and pH assays. SOC concentration was measured using an elemental analyzer (TOC, Elementar, Germany), and soil total nitrogen (TN) was measured using the FOSS Kjeltec 2300 Analyser Unit (FOSS, Hillerød, Sweden). Soil pH was determined after shaking a soil water (1: 2.5 wt/vol) suspension for 30 min.
Soil DNA extraction and sequencing
Genomic DNA was extracted from each soil aggregate sample using an E.Z.N.A.® stool DNA Kit (Omega Bio-tek, Norcross, GA, USA) according to the manufacturer’s instructions. All extracted DNA samples were stored at −20 °C before further analysis. The V3-V4 regions of the bacterial 16S rRNA gene were amplified using universal primers 338F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) (Lane et al., 1985), and the fungal ITS gene was amplified by the ITS1 (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS2 (5′-GCTGCGTTCTTCATCGATGC-3′) primers (White et al., 1990). The PCR program was as follows: 5 min initial denaturation at 95 °C; 25 cycles of denaturation at 95 °C (30 s), annealing at 56 °C (30 s), elongation at 72 °C (40 s); and a final extension at 72 °C for 10 min. PCR reactions were performed in triplicate 25 µL mixtures containing 2.5 µL of 10 × Pyrobest Buffer, 2 µL of 2.5 Mm dNTPs, 1 µL of each primer (10 Mm), 0.4 U of Pyrobest DNA Polymerase (TaKaRa), and 15 ng of template DNA. The amplicon mixture sequenced on an Illumina HiSeq 2500 platform (Biomaker, Beijing).
Processing of sequencing data
The extraction of high-quality sequences was first conducted with the QIIME package (Quantitative Insights Into Microbial Ecology) (v1.2.1). The original sequence data were sorted into valid reads after demultiplexing and quality-filtering with the following rules: (i) 300-bp reads were truncated at any site receiving an average quality score of <20 over a 50-bp sliding window, and truncated reads shorter than 50 bp were discarded; (ii) exact barcode matching, less than two nucleotide mismatches in the primer, and no ambiguous characters in the read; (iii) only overlapping sequences longer than 10 bp were assembled according to their overlapped sequence (Chen et al., 2018). The unique sequence set was classified into operational taxonomic units (OTUs) under the threshold of 97% identity using UCLUST.
Statistical analysis
The Shannon index was calculated as follows (Begon, Harper & Townsend, 1986): where N is the total OTUs of the sample, Ni is the number of individuals in group i.
The SOC, TN, and C/N ratios for the soil aggregates and plant litter were analyzed using a one-way analysis of variance (ANOVA) with a least significant difference (LSD) test at a significance level of P < 0.05 using SPSS, version 19.0. Two-way ANOVA was used to test the effects of soil aggregate size and litter type on bacterial Shannon, fungal Shannon, SOC, CO2 release, soil pH and litter mass loss. The structural changes in the soil bacterial and fungal phyla were tested by nonmetric multidimensional scaling (NMDS) based on Bray-Curtis similarity matrices using CANOCO, version 5.0. The relationship between environmental variables (pH and SOC) and bacterial/fungal communities were tested by redundancy analysis (RDA) using CANOCO. For RDA analysis, the significance of the effect of each variable, based on its eigenvalue, was tested using the Monte Carlo Permutation test, and the resulting significance level was determined by the F ratio and P- value. Hypothetical relationships among SOC, soil pH, bacterial diversity, fungal diversity, and soil respiration were quantified by structural equation modeling (SEM) using AMOS, version 21.0, and we used the non-significant chi-square (χ2) test (the model has a good fit when 0 ≤χ2 ≤ 2 and 0.05 < P ≤ 1.00) and the root mean square error of approximation (RMSEA, the model has a good fit when 0 ≤ RMSEA ≤ 0.05 and 0.10 < P ≤ 1.00) to test the goodness of the model according to Yang, Liu & Zhang (2019).
Results
Microbial diversities and properties of the soil aggregates
The microbial diversities and properties of the soil are listed in Table 2. According to the two-way ANOVA, the 1–2 mm aggregate had the highest bacterial Shannon and CO2 release in CK and leaf addition treatments, and the SOC in the <0.25 mm aggregate was higher than that in the others across the treatments. Soil pH in the 0.25–1 mm aggregate was higher than that in the others across the treatments. In two litter addition treatments, the litter mass loss in the 1–2 mm and 2–4 mm aggregate was significantly higher than that in 0.25–1 mm and <0.25 mm soil aggregate on both day 28 and day 56.
| Litter addition | Sizes | Bacterial Shannon | Fungal Shannon | Soil organic carbon (g kg−1) | CO2 release (mg C kg−1 day−1) | Soil pH | Litter mass loss (%) | |
|---|---|---|---|---|---|---|---|---|
| Day 28 | Day 56 | |||||||
| CK | 2–4 mm | 6.75(0.10) | 4.06(0.16) | 13.19(0.05) | 61.78(4.9) | 7.93(0.01) | ||
| 1–2 mm | 6.96(0.03) | 3.19(0.07) | 11.39(0.10) | 89.47(3.5) | 8.02(0.01) | |||
| 0.25–1 mm | 6.83(0.04) | 3.52(0.30) | 5.12(0.01) | 23.28(3.5) | 8.25(0.01) | |||
| <0.25 mm | 6.87(0.04) | 4.01(0.04) | 14.10(0.04) | 43.28(3.5) | 8.02(0.01) | |||
| Leaf | 2–4 mm | 6.77(0.03) | 2.43(0.04) | 14.65(0.10) | 213.95(3.8) | 7.77(0.01) | 29.3(3.2) | 39.4(2.0) |
| 1–2 mm | 6.87(0.06) | 2.47(0.05) | 14.66(0.10) | 281.60(0.8) | 7.82(0.01) | 32.4(1.0) | 43.3(1.5) | |
| 0.25–1 mm | 6.72(0.02) | 2.51(0.08) | 8.60(0.10) | 148.68(1.4) | 7.87(0.01) | 25.9(2.0) | 33.9(2.1) | |
| <0.25 mm | 6.73(0.03) | 2.45(0.08) | 16.87(0.20) | 178.68(1.4) | 7.73(0.01) | 16.9(2.1) | 21.9(2.0) | |
| Stem | 2–4 mm | 6.81(0.04) | 2.77(0.02) | 14.29(0.12) | 160.42(4.9) | 7.87(0.01) | 26.3(3.0) | 36.3(1.5) |
| 1–2 mm | 6.82(0.03) | 2.90(0.1) | 13.61(0.10) | 207.17(5.1) | 7.91(0.01) | 29.8(2.0) | 40.8(3.0) | |
| 0.25–1 mm | 6.65(0.05) | 2.80(0.04) | 9.00(0.50) | 24.75(7.3) | 8.03(0.02) | 22.1(2.1) | 30.1(3.0) | |
| <0.25 mm | 6.85(0.01) | 2.47(0.15) | 16.66(0.04) | 49.75(7.3) | 7.85(0.01) | 10.9(3.0) | 15.9(2.0) | |
| Two-way ANOVA | ||||||||
| Litter | * | *** | *** | *** | *** | ** | ** | |
| Sizes | ** | ** | *** | *** | *** | *** | *** | |
| Litter × Sizes | ns | *** | *** | *** | *** | *** | *** | |
Response of soil bacteria and fungi to litter addition
The soil bacterial and fungal community structures of the four aggregates were distinct from each other in the NMDS plots in the three litter-addition treatments (Fig. 1). The relative abundances of the dominant bacterial and fungal phyla in the soil aggregates are presented in Fig. 2. Proteobacteria and Acidobacteria were the main microflora in the soil aggregates in all three treatments, accounting for about 60% of the total abundance (Figs. 2A–2C). The relative abundance of Ascomycota was higher in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates in the treatment without leaf and stem addition, and the relative abundance of Aphelidiomycota was lower in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates (Fig. 2D). However, leaf and stem addition increased the relative abundance of Ascomycota and Aphelidiomycota dramatically (Figs. 2E and 2F). The response of soil bacteria to leaf and stem addition was weak, changing less than 10% compare with the treatment lacking litter addition. Leaf and stem addition increased the relative abundance of Proteobacteria and Bacteroidetes, but decreased the relative abundance of Acidobacteria, Gemmatimonadetes, and Actinobacteria (Figs. 3A, 3B). However, the changes in soil fungi in response to leaf and stem addition were approximately 40% compared with no litter addition treatment, and the relative abundance of Ascomycota and Aphelidiomycota increased more than 10% following the addition of leaf litter (Figs. 3C, 3D).
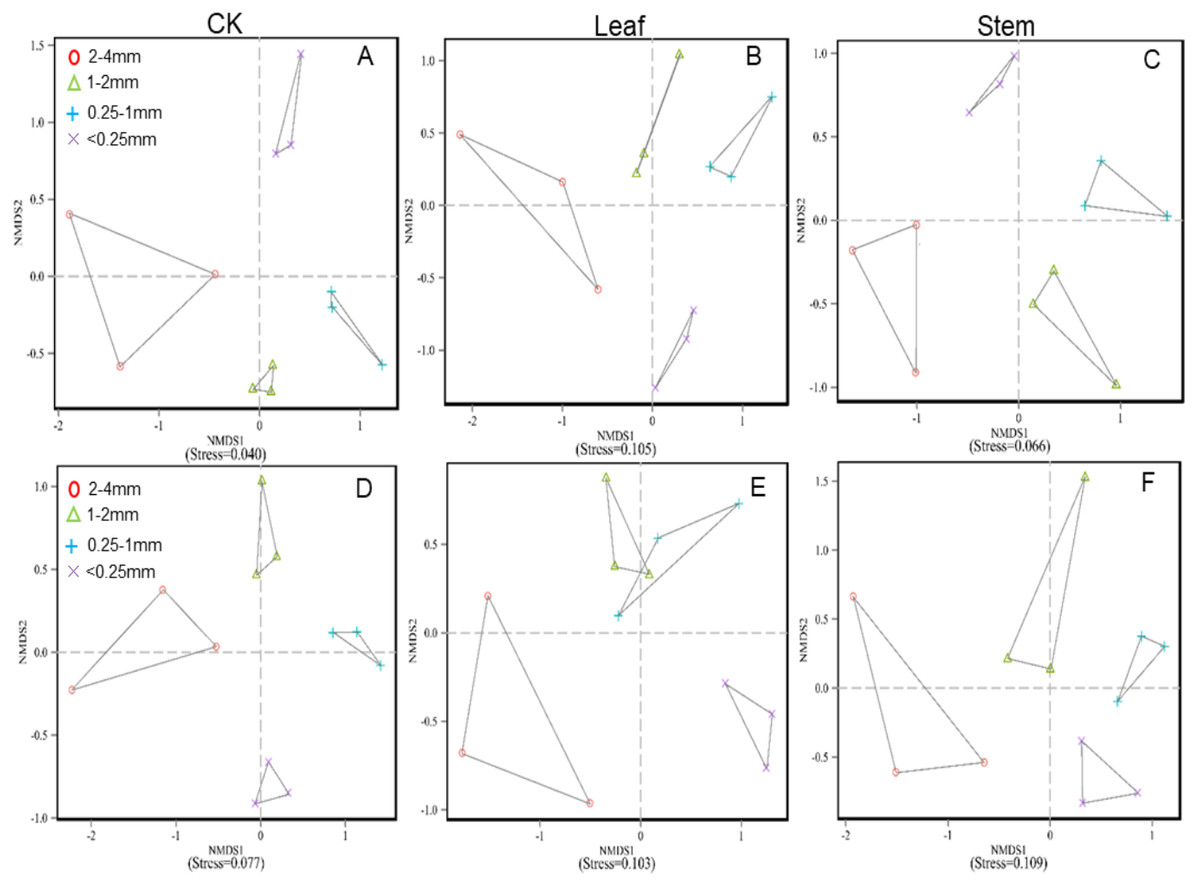
Figure 1: Nonmetric multidimensional scaling (NMDS) ordinations based on the mean abundance value of the bacterial phyla under no litter addition (A), leaf addition (B), and stem addition (C), and the fungal phyla under no litter addition (D), leaf addition (E), and stem addition (F).
Communities are indicated by colored symbols as follows: red circles, 2–4 mm; blue triangles,1–2 mm; green plus signs, 0.25–1 mm, and purple times signs, <0.25 mm.{kind=link}
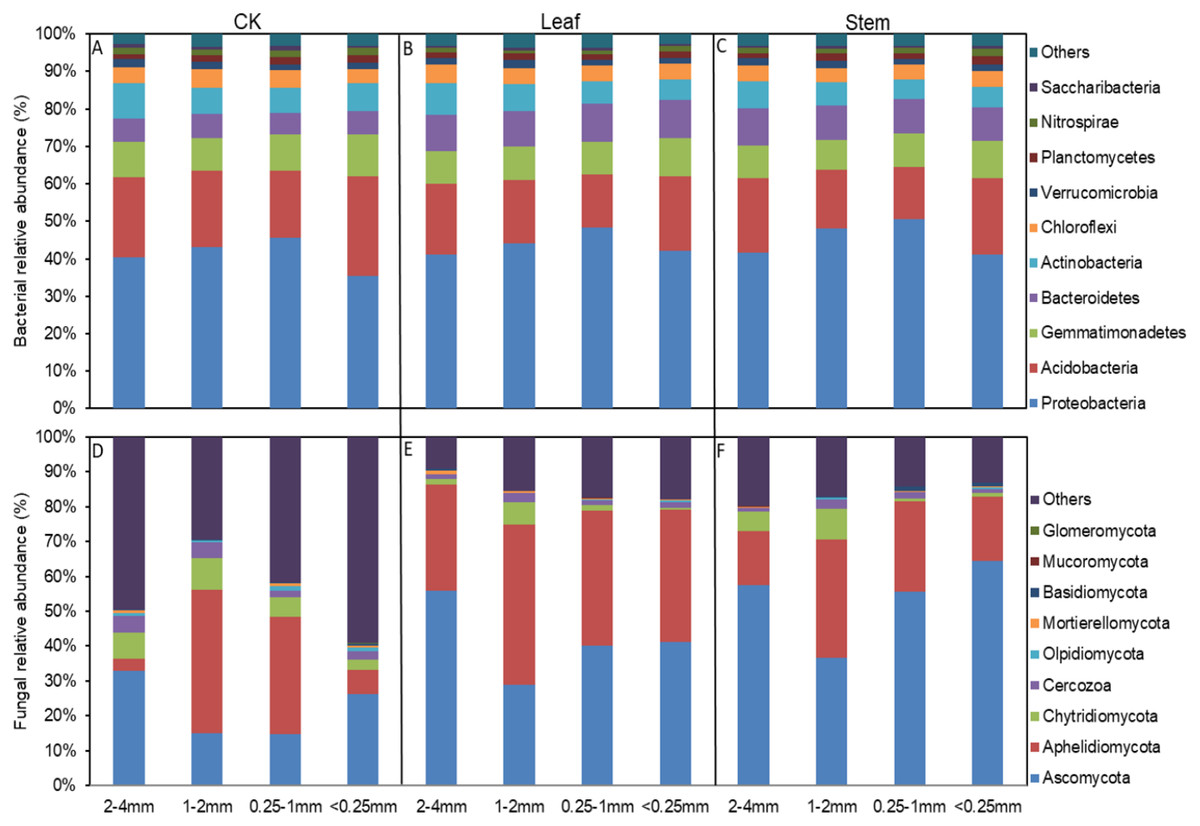
Figure 2: The relative abundances of the bacterial (A–C) and fungal phyla (D–F) in different sized aggregates.
The abundances that accounted for <1% of all of the classified sequences belong to Others in the bar graph.{kind=link}
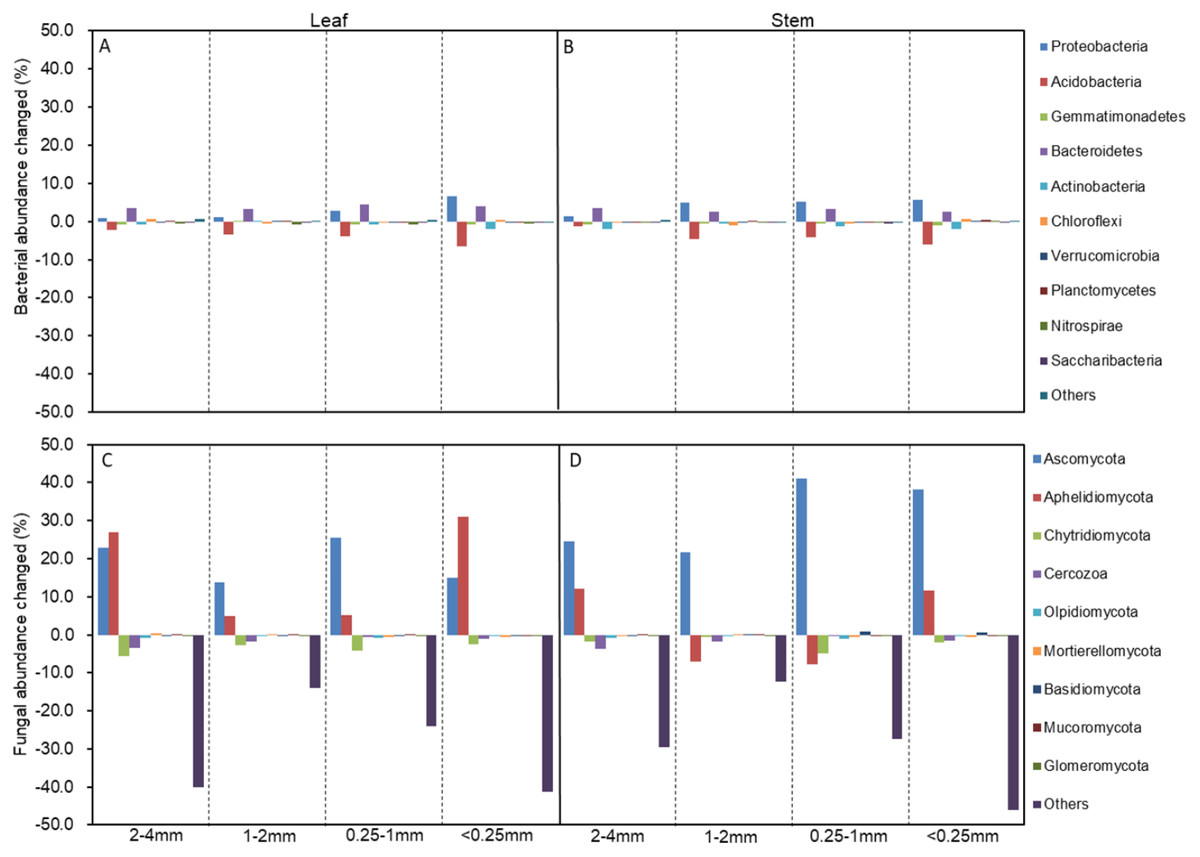
Figure 3: The changed abundances of the dominant bacterial (A, B) and fungal phyla (C, D) in different-sized aggregates under litter addition compared with no litter addition.
The abundances that accounted for <1% of all of the classified sequences belong to Others in the bar graph.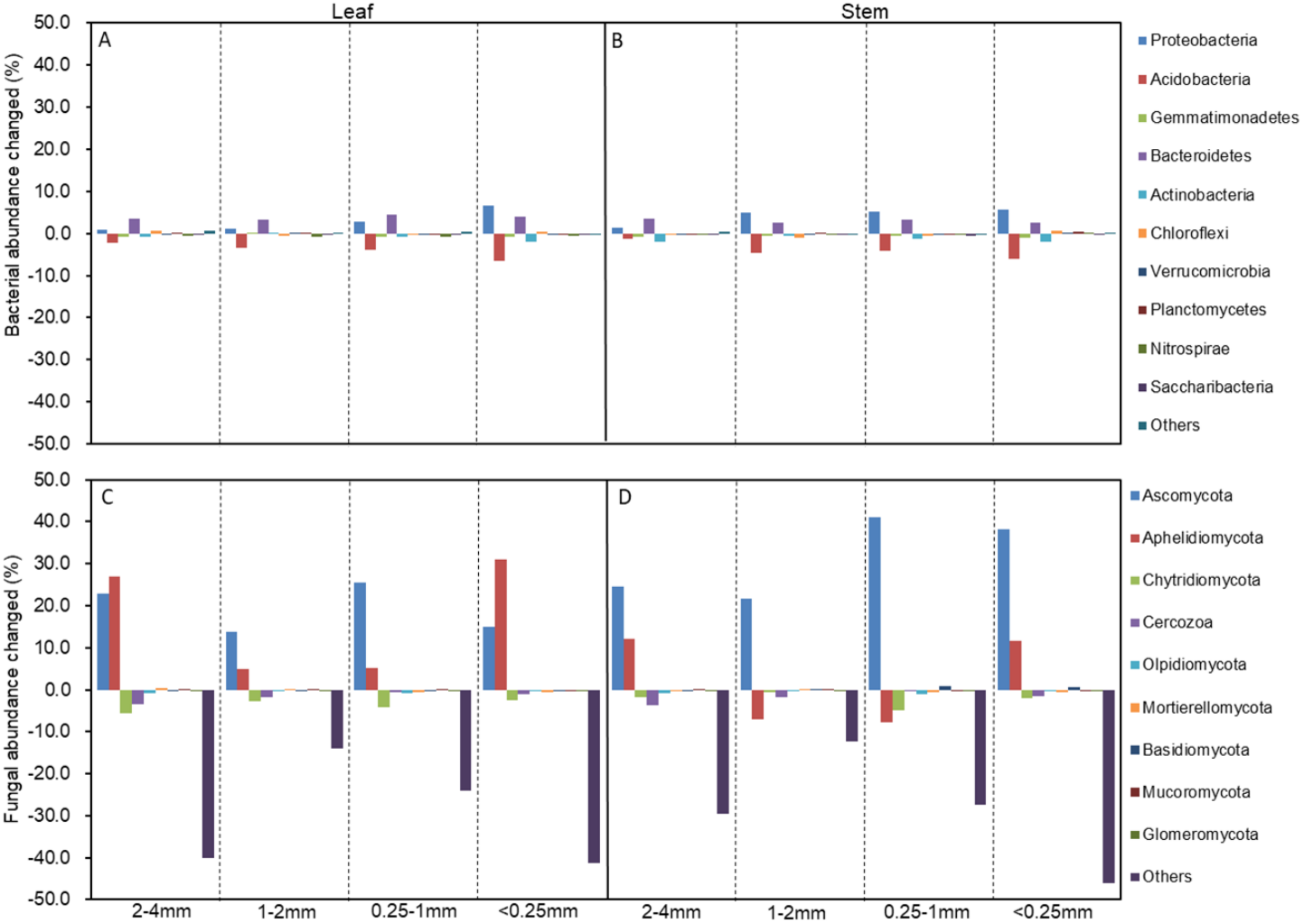{kind=link}
The relationship between soil properties and microbial diversities
RDA biplots were used to assess the physicochemical properties that influenced the abundance of the bacterial and fungal families. Overall, the combination of variables explained 65.9% and 31.5% of the bacterial (Fig. 4A) and fungal (Fig. 4B) community structure variances, respectively. Significant correlation was found between soil pH and soil bacterial communities (F = 7.51, P = 0.008), and SOC was also significantly correlated with the bacterial communities (F = 5.91, P = 0.028). In addition, the fungal communities were significantly correlated with the soil pH (F = 4.42, P = 0.042) but not by SOC (F = 2.41, P = 0.091).
The SEM showed a good fit between soil pH, microbial diversity, SOC, and respiration (Fig. 5; χ 2 = 0.07, P = 0.79; RMSEA = 0.00, P = 0.80). The fitted models explained 65% and 59% of the variance in SOC concentrations and CO2 release, respectively (Fig. 5A). Although an interaction was detected, there was no significant correlation between soil bacterial and fungal diversity with a correlation coefficient (R2) of 0.14. The SEM showed a significant interrelationship between fungal diversity and soil pH (R2 = 0.61, P < 0.001). The bacterial Shannon index indicated a significantly positive and direct effect on SOC concentrations and CO2 release (P < 0.05), while the fungal Shannon index showed significantly correlated with SOC concentrations (P < 0.001). In addition, soil pH was significantly negatively correlated with SOC concentrations and CO2 release (P < 0.001). The standardized total effects derived from the SEM revealed that SOC concentrations were mainly driven by soil pH, followed by the fungal and bacterial Shannon index, while CO2 release was mainly driven by soil pH, followed by the bacterial and fungal Shannon index (Fig. 5B)
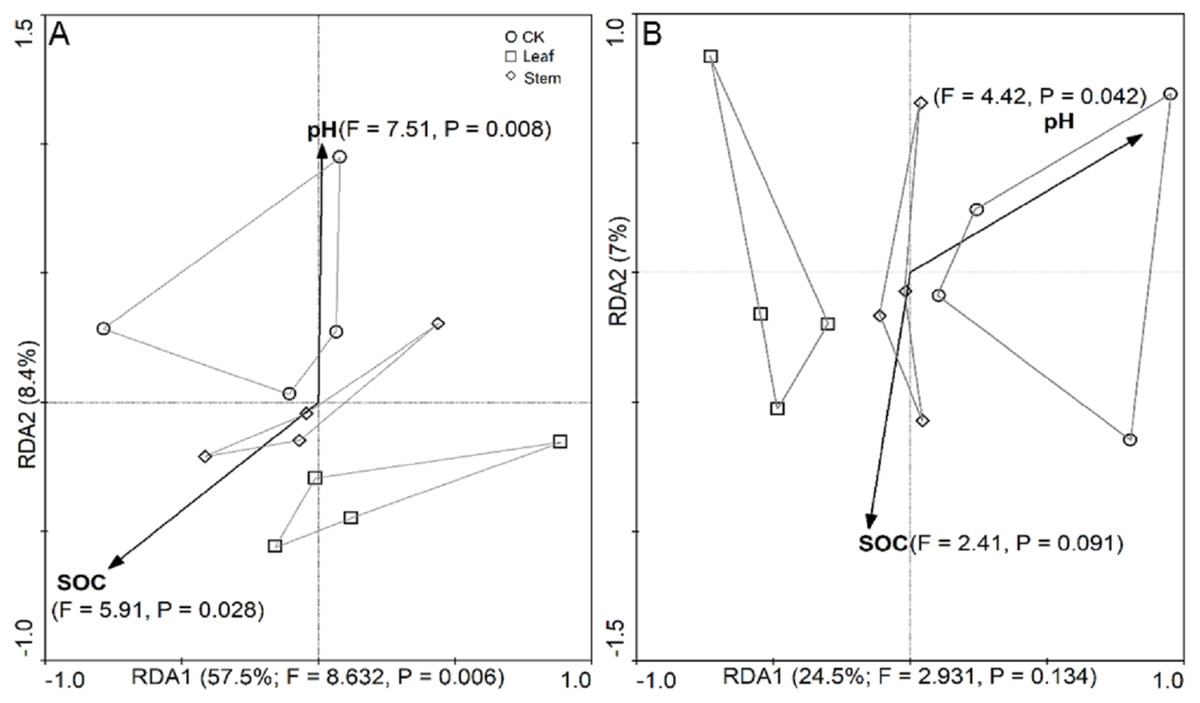
Figure 4: The redundancy analysis (RDA) showing the impact of soil properties (SOC and pH) on bacterial (A) and fungal (B) community structures.
The significance of the effect of each variable, based on its eigenvalue, was tested using the Monte Carlo Permutation test, and the resulting significance level was determined by the F ratio and P-value. Communities are indicated by symbols as follows: circles, no litter addition; squares, leaf addition; diamonds, stem addition.{kind=link}
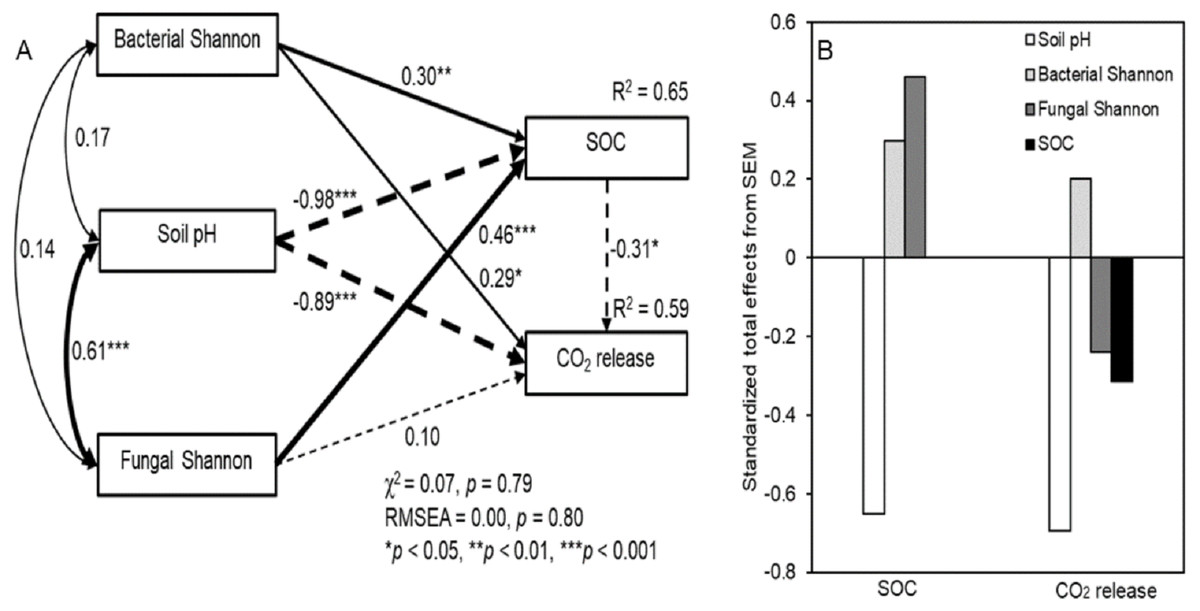
Figure 5: Structural equation model (SEM) based on the effects of the bacterial and fungal Shannon index on soil pH, SOC, and respiration under litter addition conditions (A), and their standardized total effects (direct plus indirect effects) derived from the structural equation models of SOC and CO2 release (B).
Continuous and dashed arrows indicate positive and negative relationships, respectively. The width of the arrows is proportional to the strength of the path coefficients. R2 indicates the proportion of the variance explained and appears above every response variable in the model.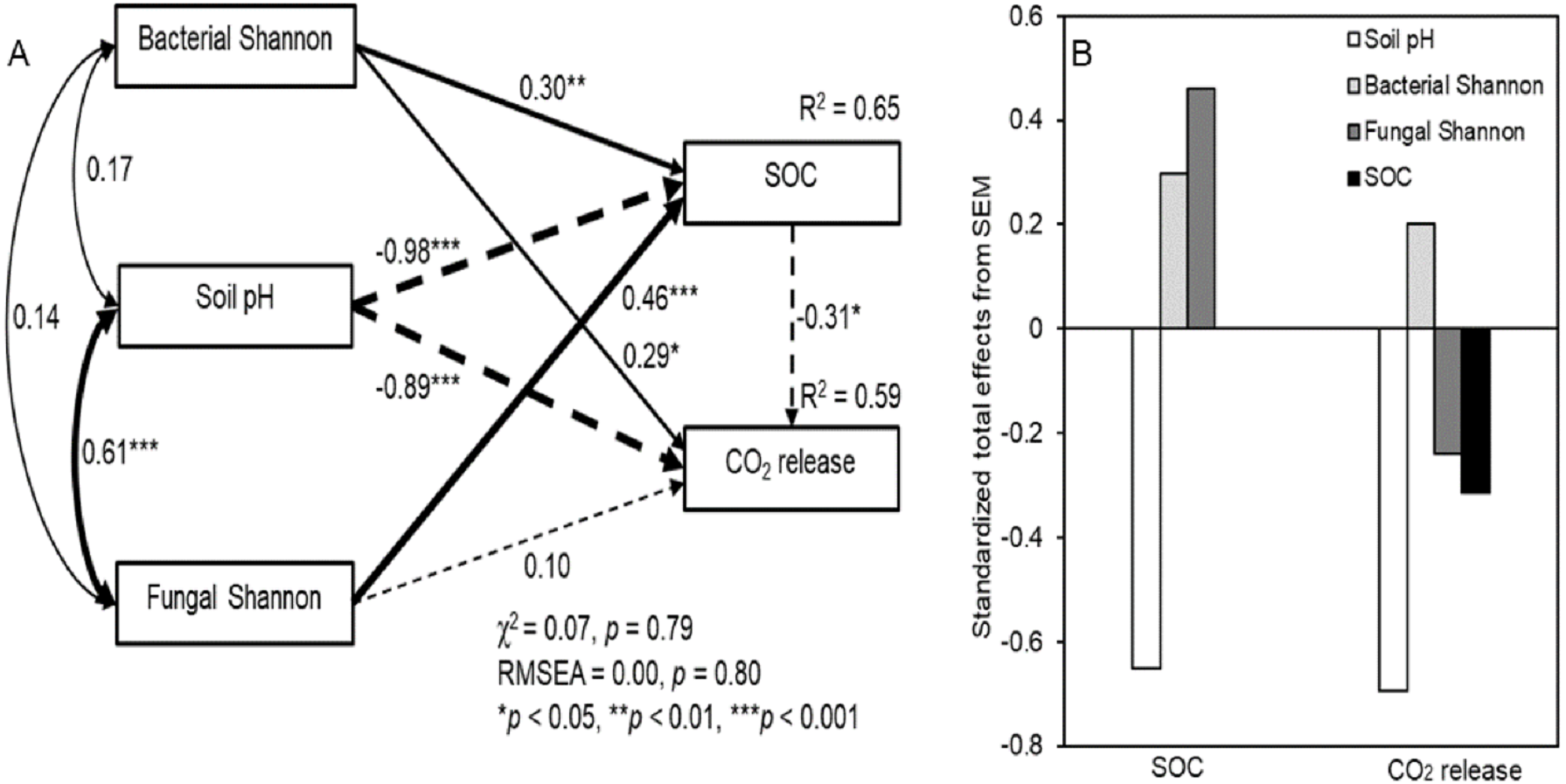{kind=link}
Discussion
Bacterial and fungal diversity in soil aggregates
Soil microbial diversity can better explain respiration than soil microbial biomass (Yang, Liu & Zhang, 2019). Furthermore, as macro-aggregates are generally dominated by soil fungi (Frey, 2005), our study considered the diversity of both the soil bacteria and soil fungi. The 1–2 mm aggregate possessed both the highest bacterial Shannon and the lowest fungal Shannon under no litter addition, implying that soil bacteria and fungi in the same resources and spaces undergo interspecies interactions at the surface of the macro-aggregates (Effmert et al., 2012), and in this aggregate, bacterial diversity is favored. Litter addition eventually resulted in increased respiration; however, the Shannon index of both the bacteria and fungi decreased in the litter-addition treatment compared with no litter addition. Bamminger et al. (2014) suggested that litter addition increases microbial biomass and results in a shift in the composition of the soil microbial community, especially for soil fungi, which is consistent with the present study. The relative abundance of individual species will increase dramatically with the addition of litter, particularly that of soil fungi, resulting in a decline in microbial diversity.
Response of soil bacteria and fungi to litter addition in soil aggregates
Adding leaf litter to the soil has a positive effect on the soil microbial community as a result of the increased carbon and nutrient resources (Fanin & Bertrand, 2016). Bacterial communities are characterized by a series of colonizers during litter decomposition, with Proteobacteria, Actinobacteria and Bacteroidetes being the most abundant taxa found over the entire decomposition process (Purahong et al., 2016; Tlaskal, Voriskova & Baldrian, 2016). Our study demonstrates that Proteobacteria, Actinobacteria, Gemmatimonadetes and Bacteroidetes are the four most-important bacterial phyla within different aggregate sizes and litter addition treatments. In addition, Sun et al. (2017) found that bacterial abundance did not show a significant change following the addition of leaf litter, which corroborates the observations made in the present study. Liang et al. (2017) suggest that the addition of residues alters the soil microbial community composition and promotes fungal growth more than bacterial growth, which is in agreement with other studies that found that soil microorganisms respond to altered residue modifications by changing their community composition (Phillips et al., 2002) and also supports the observation that saprophytic fungi play a major role in the decomposition of residues (Meidute, Demoling & Baath, 2008). Indeed, the fungal relative abundances in our study changed significantly following leaf litter addition, particularly Ascomycota and Aphelidiomycota. Given that Aphelidiomycota are not typical soil fungi, but widely present in soil when litter was added, its role in litter decomposition requires further study.
Interactions between soil pH, SOC and microbial diversity after litter addition
There is increasing evidence that soil microbial communities influence litter decomposition rates over and above the climate and litter quality controls (Bradford et al., 2016; Schimel & Schaeffer, 2012). Adding leaf litter to the soil increases the SOC and soil pH (Sun et al., 2017), and the soil pH tends to be neutral after adding litter to acid soil. Similarity, in the present study, the soil pH decreased and tended to neutralize following litter addition, implying that litter addition regulates the acid–base balance of the soil. In addition, Rousk et al. (2010) suggested that soil pH has a strong influence on the diversity and composition of soil bacterial and fungal communities across a gradient. In contrast, soil microbial diversity also influences soil pH. In the present study, soil fungal diversity decreased after litter addition, which decreased the soil pH. The composition and diversity of the soil fungal and bacterial communities are thus often strongly correlated with soil pH (Lauber et al., 2009). In addition, the SEM showed a significant interrelationship between fungal diversity and soil pH, and fungal diversity showed significant correlations with SOC concentrations, especially in the microenvironment of litter addition. The standardized total effects derived from the SEM revealed that SOC concentrations were mainly driven by soil pH, followed by the fungal and bacterial diversity, while CO2 release was mainly driven by soil pH, followed by the bacterial and fungal diversity. Such datasets are valuable in advancing our understanding of the role of different groups of microorganisms in soil and how microbial activities in the soil in response to litter addition contributes to nutrient fluxes in specific soil environments.
Conclusions
Our study shows a highly different response of bacteria and fungi in soil aggregates to litter addition. Litter addition increased the relative abundance of Proteobacteria and Bacteroidetes, but decreased the relative abundance of Acidobacteria, Gemmatimonadetes, and Actinobacteria. The relative abundance of Ascomycota was higher in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates in the treatment without litter addition, and the relative abundance of Aphelidiomycota was lower in the 2–4 mm and <0.25 mm aggregates than in the 1–2 mm and 0.25–1 mm aggregates. Soil pH and SOC were found to be the determining factors shaping the bacterial communities. The bacterial Shannon index had a significantly positive and direct effect on SOC concentration and CO2 release, while the fungal Shannon index showed a significant correlation with SOC concentration. Our results indicate that soil bacterial diversity contributes positively to both carbon emissions and carbon storage, whereas soil fungal diversity can promote carbon storage and decrease carbon emissions.