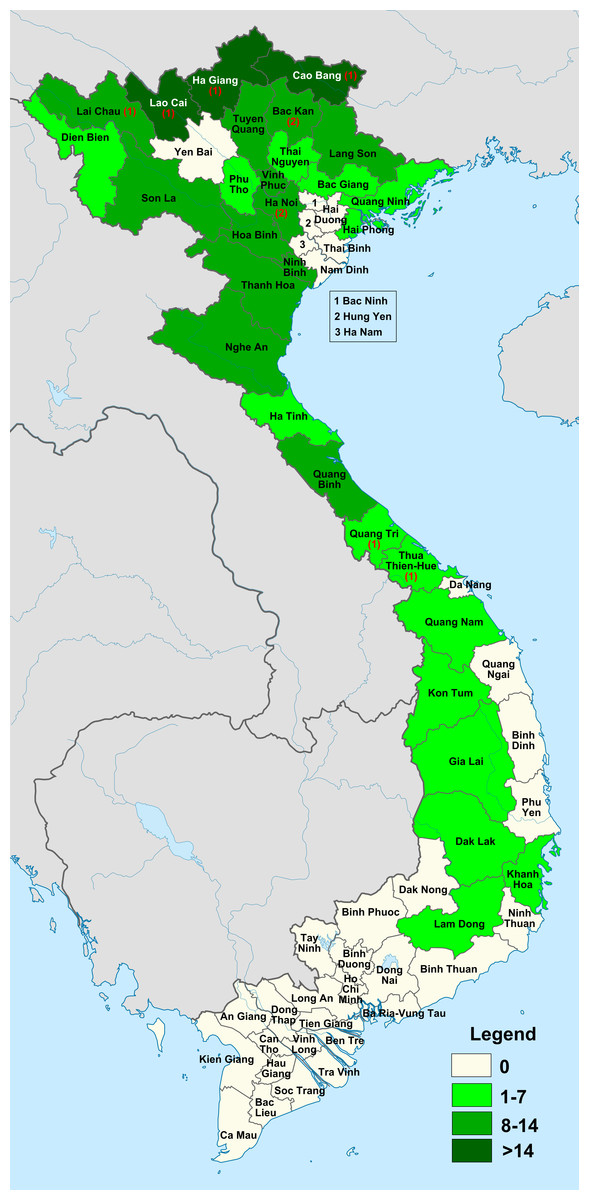
Appendix. Checklist of Elatostema in Vietnam
Species are listed in alphabetical order. Basionyms and synonyms are cited on a separate line in alphabetical order. This is followed by the species’ global distribution, elevation range within Vietnam and the exsiccatee seen for this study. New records for Vietnam are denoted by an ‘*’ next to the name.
Elatostema acuminatum (Poir.) Brongn.
Procris acuminata Poir.
Boehmeria acuminata Pers., Elatostema lowii Stapf., Elatostema membranaceum Hassk., Elatostema membranifolium Kurz, Langeveldia acuminata Gaudich., Ramium acuminatum (Pers.) Kuntze.
Bhutan, China, Indonesia, Malaysia, Myanmar, Nepal, Sikkim, Thailand, Vietnam; 400–1,650 m.
Cao Bang Province: Ha Lang District, Dong Loan Municipality, 22°46′N103°44′E, L. Averyanov et al. CBL603 (LE); Ha Giang Province: Bac Me District, Phieng Luong Municipality, 22°39′29″N 105°19′35″E, L. Averyanov et al. HAL6540 (LE); Quan Ba District, Can Ty Municipality, 23°04′N 104°59′E, N.T. Hiep et al. NTH3622 (LE); Vi Xuyen District, Cao Bo Municipality, Tam Ve Village, 22°46′18″N 104°49′01″E, D.K. Harder et al. 5615 (K, MO); Ha Noi City: Ba Vi District, Ba Vi National Park, 21°04′22″N 105°21′53″E, L.F. Fu et al. VMN_CN35 (IBK, VNMN); Quang Binh Province: Minh Hoa District, Dan Hoa Municipality, 17°45′28″N 105°46′14″E, L. Averyanov et al. HAL12205 (LE), 17°46′34″N 105°48′56″E, L. Averyanov et al. HAL12548 (LE); Vinh Phuc Province: Tam Dao District, L. Averyanov et al. LX-VN864 (LE).
Elatostema acutidentatum (W.T.Wang) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia acutidentata W.T.Wang, Bull. Bot. Lab. N.-E. Forest. Inst., Harbin 6: 53. 1980. Type: China. Yunnan, Xichou county, Fadou village, C.W. Wang & Y. Liu 85536 (holotype: PE!).
China, Vietnam; 1,400–1,900 m.
Cao Bang Province: Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL195 (LE, MO); Lai Chau Province: Than Uyen District, Ho Mit Municipality, 22°06′N 103°52′E, N.T. Hiep et al. NTH2842 (LE); Lao Cai Province: Sa Pa District, Anon. TV-284 (HN); Van Ban District, 21°59′00″N 104°02′00″E, L. Averyanov et al. HAL2215 (LE).
Elatostema albopilosum W.T.Wang
a. Elatostema albopilosum var. vicinum L.D.Duan & Y.Lin
Endemic; 200–400 m.
Bac Kan Province: Cho Don District, Xuan Lac Commune, 22°21′91″N 105°32′95″E, L.Q. Li et al. 0432 (PE); Tuyen Quang Province: Na Hang District, H.N. Qin et al. 319 (PE).
This variety is very similar to E. albopilosum W.T.Wang var. albopilosum and further study may result in it being treated as a synonym.
Elatostema arcuatobracteatum L.F.Fu, V.T.Do & C.X.He
Endemic; ca. 500 m.
Ha Giang Province: Bac Me District, Bac Me Nature Reserve, 22°45′15″N 105° 13′43″E, L.F. Fu et al. VMN_CN219 (IBK, VNMN).
Elatostema atropurpureum Gagnep.
China, Vietnam; 300–1,150 m.
Cao Bang Province: Nguyen Binh District, Phia Oac—Phia Den National Park, F. Wen et al. VMN_CN532 (IBK, VNMN); Ha Noi City: Ba Vi District, Ba Vi Mountain, B. Balansa 2534 (P), 21°04.569′N 105°21.765′E, N.A. Vislobokov & S.P. Kuznetsova s.n. (MW); Ha Giang province: Quan Ba District, Tung Vai Municipality, Thang village, 23°03′13″N 104°51′49″E, L. Averyanov VR558 (LE); Lao Cai Province: Sa Pa District, Sino-Vietnam Expedition 336 (IBSC, PE); Van Ban District, Hoang Lien—Van Ban Nature Reserve, L.F. Fu et al. VMN_CN920 (IBK, VNMN); P.A. Petelot 5100 (P); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, F. Wen et al. VMN_CN600, VMN_CN606 (IBK, VNMN); Phu Tho Province: Thanh Son District, Xuan Son Municipality, Xuan Son National Park, 21°08′00″N 104°55′55″E, M.S. Nuraliev & N.A. Vislobokov 1105 (IBK, MW).
Elatostema atroviride W.T.Wang
Elatostema atroviride var. lobulatum W.T.Wang, E. leiocephalum W.T.Wang, E. papilionaceum W.T.Wang.
China, Vietnam; 200–1,500 m.
Cao Bang Province: Ha Quang District, Pac Bo, L.F. Fu et al. VMN_CN783 (IBK, VNMN); Ha Giang Province: Yen Minh District, Lao Va Chai Municipality, 23°07′N 105°08′E, N.T. Hiep et al. NTH3448 (LE); Hai Phong City: Cat Hai District, Y.G. Wei et al. VMN_CN14 (IBK, VNMN); Cat Hai District, Cat Ba Island, N. Arnautov et al. LX-VN3626, LX-VN3774, LX-VN3793, LX-VN3834 (LE); between Meo Vac District and Dong Van District, 23°15′16″N 105°23′12″E, L.F. Fu et al. VMN_CN190 (IBK, VNMN), 22°45′23″N 105°13′46″E, L.F. Fu et al. VMN_CN218 (IBK, VNMN); Hoa Binh Province: Mai Chau District, Pa Co Municipality, 20°44′32.3″N 104°56′26.9″E, V.X. Phuong et al. HNK639 (K); Nghe An Province: Quy Chau District, Pu Huong Natural Reserve, F. Wen et al. VMN_CN478 (IBK, VNMN); Con Cuong District, Cay Da Valley, Truong Huong Hamlet, Yen Khe Commune, 19°00′39.3″N 104°51′38″E, V.D. Nguyen et al. HNK2960 (K); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2177 (TAI); Quang Binh Province: Minh Hoa District, Thong Hoa Municipality, 17°40′N 105°57′E, L. Averyanov et al. VH4797 (LE); Tuyen Hoa District, Lam Hoa Municipality, 17°55′41″N 105°49′43″E, L. Averyanov et al. CPC2620 (LE); Son La Province: Moc Chau District, Van Ho Municipality, Hua Tat Village, 20°46′18″N 104°47′45″E, D.K. Harder et al. 5774 (HN, K, MO), 20°46′18″N 104°47′29″E, N.T. Hiep et al. HAL9311 (LE); Thanh Hoa Province: Quy Chau District, Pu Huong Natural Reserve, F. Wen et al. VMN_CN427 (IBK, VNMN); Thuong Xuan District, Bat Mot Municipality, 20°01′12″N 104°57′48″E, L. Averyanov et al. CPC6655 (LE).
*Elatostema attenuatoides W.T.Wang
China, Vietnam; 500–800 m.
Cao Bang Province: Ha Lang District, An Lac Municipality, 22°43′N 103°36′E, L. Averyanov et al. CBL786 (LE, MO); Tra Linh District, Quoc Toan Municipality, 22°45′15″N 106°17′54″E, D.K. Harder et al. 4336 (K); 22°45′30″N 106°17′22″E, L. Averyanov et al. VH4854 (HN, LE, MO), VH4949 (LE, MO); 22°45′N 106°19′E, L. Averyanov et al. CBL1256 (LE, MO).
*Elatostema austrosinense Y.H.Tseng & A.K.Monro nom. nov.
non E. macrophyllum Brongn.
Pellionia macrophylla W.T.Wang, Acta Bot. Yunnan. 5: 271. 1980. Type: China. Yunnan, Luchun county, Huanglianshan, D.D. Tao 845 (holotype: KUN!).
China, Vietnam; ca. 1,650 m.
Lao Cai Province: Sa Pa District, Hoang Lien National Park, L.F. Fu et al. VMN_CN884 (IBK, VNMN); Van Ban District, 21°58′42″N 104°00′37″E, L. Averyanov et al. HAL2072, HAL2088 (LE).
We selected ‘austrosinense’ as epiphet as it coincides with the origin of its distribution area, South China.
Elatostema backanense (Gagnep.) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia backanensis Gagnep., Bull. Soc. Bot. France 75: 918. 1928. Type: Vietnam. Bac Kan, Yen Lac District, P.A. Eberhardt 4669 (holotype: P!).
Endemic; elevation unknown.
Bac Kan Province: Yen Lac District, P.A. Eberhardt 4669 (P).
*Elatostema backeri H.Schroet.
China, Indonesia, Vietnam; elevation unknown.
Cao Bang Province: Bao Lac District, Dinh Phung Municipality, L.F. Fu et al. VMN_CN814 (IBK, VNMN); Bao Lam District, Tan Viet, Na Dang, L.F. Fu et al. VMN_CN829 (IBK, VNMN); Thanh Hoa Province: Quan Hua District, Pu Luong Nature Reserve, F. Wen et al. VMN_CN551 (IBK, VNMN).
Elatostema balansae Gagnep.
Elatostema balansae var. hispidum W.T.Wang, E. platyphyllum Wedd. var. balansae (Gagnep.) Yahara.
China, Malaysia, Thailand, Vietnam; 1,100–1,700 m.
Cao Bang Province: Nguyen Binh District, Phia Oac—Phia Den Nature Reserve, 22°37′49″N 105°53′11″E, L.F. Fu et al. VMN_CN170 (IBK, VNMN); Ha Giang Province: between Meo Vac District and Dong Van District, 23°09′16″N 105°24′52″E, L.F. Fu et al. VMN_CN192 (IBK, VNMN); Hoa Binh Province: Mai Chau District, Hang Kia Municipality, 20°43′39″N 104°53′43″E, N.T. Hiep et al. HAL751 (HN); Lai Chau Province: Sin Ho District, Ta Ngau Municipality, 22°18′N 103°15′E, N.T. Hiep et al. NTH2783 (LE); Lam Dong Province: Lac Duong District, Da Chais Municipality, 12°08′N 108°39′E, L. Averyanov et al. VH4511 (LE, MO); Lac Duong District, Da Chais Municipality, Bi Doup mountains, 12°11′07.0″N 108°42′55.9″E, A.N. Demidova & N.G. Prilepsky 431 (MW); Son La Province, Yen Chau District, Muong Lum Municipality, On Oc Village, 20°59′44″N 104°30′03″E, D.K. Harder et al. 7229 (HN).
Elatostema brachyodontum (Hand.-Mazz.) W.T.Wang
Elatostema ficoides Wedd. var. brachyodontum Hand.-Mazz.
China, Vietnam; elevation unknown.
Province unknown: Phuong-Lam, B. Balansa 2535 (P).
Elatostema brevifolium (Benth.) Hallier f.
Pellionia brevifolia Benth.
Amaranthus polychroa Raeusch., Elatostema radicans var. minimum (Makino) H.Schroet. Pellionia minima Makino.
China, Japan, Vietnam; 60–1,900 m.
Cao Bang Province: Nguyen Binh District, Phia Oac—Phia Den Nature Reserve, 22°36′29″N 105°52′15″E, L.F. Fu et al. VMN_CN161 (IBK, VNMN); Lao Cai Province: Sa Pa District, Hoang Lien National Park, Ton Forest Station, L.F. Fu et al. VMN_CN915 (IBK, VNMN); Sa Pa District, San Sa Ho Municipality, 22°21′10″N 103°46′30″E, M.S. Nuraliev & D.D. Sokoloff 263 (IBK, K, MW).
*Elatostema brunneinerve W.T.Wang
China, Vietnam; 1,200–1,400 m.
Ha Giang province: Quan Ba District, Tung Vai Municipality, Thang village, 23°03′42″N 104°50′42″E, L. Averyanov et al. VR591 (LE).
Elatostema caulialatum (S.Y.Liou) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia caulialata S.Y.Liou, Guihaia 3(4): 317. 1983. Type: China. Guangxi, Guiping County, Dapingshan, S.Y. Liu et al. D00322 (holotype: GXMI!, isotypes: PE!).
China, Vietnam; 200–415 m.
Bac Kan Province: Cho Don District, Xuan Lac Community, 22°21.91′N 105°32.95′E, L.Q. Li et al. 0386 (HN); Na Ri District, Kim Hy Nature Reserve, 22°17′54″N 106°01′40″E, L.F. Fu et al. VMN_CN128 (IBK, VNMN); Ha Giang Province: Meo Vac District, L.F. Fu et al. VMN_CN859 (IBK, VNMN); Thai Nguyen Province: Vo Nhai District, Truong Lung Municipality, 21°48.75′N 105°58.09′E, L.Q. Li et al. 0142 (HN); Tuyen Quang Province: Na Hang District, Anon. TV-93 (HN).
Elatostema chapaense (Gagnep.) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia chapaensis Gagnep., Bull. Soc. Bot. France 75: 920. 1928. Type: Vietnam. Lao Cai, Sa Pa District, H. Lecomte & A. Finet 415 (holotype: P!).
Endemic; elevation unknown.
Lao Cai Province: Sa Pa District, H. Lecomte & A. Finet 415 (P).
Elatostema cochinchinense (Gagnep.) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia cochinchinensis Gagnep., Bull. Soc. Bot. France 75: 921. 1928. Type: Vietnam. Haut Donnai, Con-heu Mountain, L. Pierre 4814 (holotype: A!).
Endemic; elevation unknown.
Province unknown: Haut Donnai, Con-heu Mountain, L. Pierre 4814 (A).
*Elatostema crassiusculum W.T.Wang
China, Vietnam; ca. 200 m.
Cao Bang Province: Bao Lam District, Nam Quang, L.F. Fu et al. VMN_CN838 (IBK, VNMN).
*Elatostema crenatum W.T.Wang
China, Vietnam, elevation unknown.
Phu Tho Province: Ha Ho District, Ao Chau, V.X. Phuong 3483 (HN).
Elatostema crispulihirtellum (W.T.Wang) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia crispulihirtella W.T.Wang, Bull. Bot. Res., Harbin 3(3): 57. 1983. Type: China. Yunnan, Pingbian, Shantou county, C.W. Wang 82383 (holotype: PE!).
China, Vietnam; 710 m.
Lao Cai Province: On the way from Lao Cai to Sa Pa, Sino-Vietnam Expedition 856 (PE).
Elatostema cyrtandrifolium (Zoll. & Mor.) Miq.
Procris cyrtandrifolia Zoll. & Mor.
Elatostema herbaceifolium Hayata, E. sessile var. cyrtandrifolium (Zoll. & Mor.) Wedd., E. sessile var. pubescens Hook.f., E. herbaceifolium Hayata.
Bhutan, China, India, Indonesia, Malaysia, Myanmar, Thailand, Vietnam; 200–1,050 m.
Cao Bang Province: Tra Linh District, Luu Ngoc Municipality, 22°47′N 106°18′E, L. Averyanov et al. CBL1203 (LE, MO); Dien Bien Province: Tua Chua District, Xa Nhe Municipality, 21°52′35″N 103°24′21″E, L. Averyanov et al. CPC2221 (LE); Ha Giang Province: between Meo Vac District and Dong Van District, 23°13′45″N 105°24′48″E, L.F. Fu et al. VMN_CN244 (IBK, VNMN); Meo Vac District, Sung Cha Municipality, 23°11′N 105°17′E, N.T. Hiep et al. NTH3302 (LE); Hai Phong City: Cat Hai District, Cat Ba Island, N. Arnautov et al. LX-VN3796 (LE); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2187 (TAI); Quang Binh Province: Minh Hoa District, Thong Hoa Municipality, 17°40′N 105°57′E, L. Averyanov et al. VH4793 (HN, LE, MO); Son La Province: Moc Chau District, Chieng Hac Municipality, 20°51′N 104°38′E, N.T. Hiep et al. NTH2937 (LE); Thanh Hoa Province, Quy Chau District, Pu Huong Natural Reserve, F. Wen et al. VMN_CN421 (IBK, VNMN).
Elatostema dissectum Wedd., Arch. Mus. Hist. Nat. 9(1–2): 314. 1856. Type: India, Khasia, D. Hooker & T. Thomson s.n. (lectotype: designated by Lin, Duan & Yang, 2009: 1910, P!).
Elatostema paragungshanense W.T.Wang, Pellionia tonkinensis Gagnep.
Pellionia tetramera Gagnep., Bull. Soc. Bot. France 75: 925. 1928, syn. nov. Type: Vietnam. Ba Vi District, Ba Vi Mountain, B. Balansa 2536 (holotype: P!).
Bhutan, China, India, Laos, Sikkim, Thailand, Vietnam; 200–1,350 m.
Bac Kan Province: Phu Thong Hoa, P.A. Eberharadt 4724 (P); Ha Noi City: Ba Vi District, Ba Vi National Park, V.D. Nguyen et al. HNK 1268 (K); Ba Vi District, Ba Vi Mountain, B. Balansa 2536 (P); Ha Giang province: Quan Ba District, Tung Vai Municipality, Thang village, 23°03′13″N 104°51′49″E, L. Averyanov et al. VR554, VR559, VR563, VR660, VR713 (LE); Hoa Binh Province: Da Bac District, Cao Son Municipality, Buoi Truong Station, 20°52′31″N 105°11′48″E, N.T. Hiep et al. HAL540 (HN, MO); Mai Chau District, F. Wen et al. VMN_CN262 (IBK, VNMN); Mai Chou District, Hang Kia-Pa Co Nature Reserve, F. Wen et al. VMN_CN326, VMN_CN331, VMN_CN337 (IBK, VNMN); Kon Tum Province: Dak Glei District, L. Averyanov et al. VH1757 (HN, MO); Lang Son Province: Bac Son District, Tan Tri Municipality, L. Averyanov et al. HAL6783 (LE); Lao Cai Province: Sa Pa District, Hoang Lien National Park, Ton Forest Station, L.F. Fu et al. VMN_CN916 (IBK, VNMN); Van Ban District, Khanh Yen Ha Municipality, to the S of Na Nheo Village, 21°59′04″N 104°15′08″E, L. Averyanov et al. HAL2814 (HN, LE); Van Ban District, Nam Xe Municipality, 22°01′27″N 104°00′25″E, D.K. Harder et al. DKH7051 (LE); Nghe An Province: Con Cuong District, Chau Khe Municipality, Pu Mat National Park, 18°55′56″N 104°35′01″E, M.S. Nuarliev 2128 (MW); Quy Chau District, Pu Huong Natural Reserve, F. Wen et al. VMN_CN486 (IBK, VNMN); Tuong Duong District, Tam Hop Municipality, 19°05′07″N 104°20′55″E, L. Averyanov et al. HLF7074 (LE); Quang Binh Province: Minh Hoa District, Dan Hoa Municipality, 17°41′50″N 105°45′54″E, L. Averyanov et al. HAL12410 (LE); Quang Tri Province: Huong Hoa District, Huong Viet Municipality, 16°51′14″N 106°34′13″E, N.T. Hiep et al. HLF5947 (LE); Son La Province: Chieng Yen District, N.H. Hien et al. VN239 (HN); Tuyen Quang Province: Bach Ngoc, P.A. Eberhardt 4818 (P); Na Hang District, Vinh Yen Municipality, 22°21′09″N 105°25′20″E, L.V. Cham et al. HLF299 (HN).
We place Pellionia tetramera in synonymy based on its shared characters with Elatostema dissectum: oblong-lanceolate lamina and staminate inflorescence with long peduncle.
Elatostema eberhardtii (Gagnep.) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia eberhardtii Gagnep., Bull. Soc. Bot. France 75: 921. 1928. Type: Vietnam. Quang Tri, Cu Bi, P.A. Eberharadt 1962 (holotype: P!).
Endemic; elevation unknown.
Quang Tri Province: Cu Bi, P.A. Eberharadt 1962 (P).
Elatostema eurhynchum Miq.
Indonesia, Vietnam; 1,600 m.
Kon Tum Province: Dak Glei District, 15°04′00″N 107°59′00″E, L. Averyanov et al. VH797 (HN, LE, MO).
*Elatostema fengshanense W.T.Wang & Y.G.Wei
China, Vietnam; elevation unknown.
Cao Bang Province: Ha Quang District, Pac Bo, L.F. Fu et al. VMN_CN786 (IBK, VNMN).
Elatostema ficoides Wedd.
Elatostema mariannae C.B.Clarke.
China, India, Nepal, Sikkim, Vietnam; 400 m.
Bac Kan Province: Na Ri District, Kim Hy Nature Reserve, 22°11′56″N 106°04′04″E, L.F. Fu et al. VMN_CN129 (IBK, VNMN). Hoa Binh Province: Crieng Xen, P.A. Petelot 2965 (P).
*Elatostema glochidioides W.T.Wang
China, Vietnam; 300–1,190 m.
Ha Giang Province: Between Meo Vac District and Dong Van District, L.F. Fu et al. VMN_CN233 (IBK, VNMN); Quan Ba District, Bat Dai Son Nature Reserve, Thanh Van commune, Mo Sai village, 23°07′46″N 104°57′56″E, L. Averyanov et al. VR417 (LE); Quan Ba District, Thang village, 23°03′13″N 104°51′49″E L. Averyanov et al. VR553 (LE).
Elatostema grijsii (Hance) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia grijsii Hance, J. Bot. 6(62): 49. 1868. Type: China. Fujian, Exsicc. 6704 (holotype: BM!).
Pellionia funingensis W.T.Wang.
China, Vietnam; 800–1,900 m.
Cao Bang Province: Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL069 (LE, MO); Nguyen Binh District, Phia Oac—Phia Den Nature Reserve, 22°36′24″N 105°52′46″E, L.F. Fu et al. VMN_CN157 (IBK, VNMN); Lang Son Province: Bac Son District, Tan Tri Municipality, L. Averyanov et al. HAL6753 (LE); Lao Cai Province: Van Ban District, Hoang Lien - Van Ban Nature Reserve, L.F. Fu et al. VMN_CN921 (IBK, VNMN); Vinh Phuc Province: Tam Dao Range, N. Arnautov et al. LX-VN4004 (LE).
Elatostema heterolobum (Wedd.) Hallier f.
Pellionia heteroloba Wedd.
Elatostema bicuspidatum Hallier f., E. griffithianum (Wedd.) Hallier f., E. henryanum Hand.-Mazz., Pellionia heteroloba var. minor W.T.Wang, P. keitaoensis Yamam., P. menglianensis Y.Y.Qian.
Bhutan, China, India, Laos, Myanmar, Sikkim, Vietnam; 400–1,900 m.
Cao Bang Province: Bao Lac District, Dinh Phung Municipality, L.F. Fu et al. VMN_CN816 (IBK, VNMN); Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL035, CBL062 (HN, LE, MO); Ha Giang Province: Meo Vac District, Vicinities of Meo Vac Town, 23°10′N 105°24′E, N.T. Hiep et al. NTH3385 (LE); Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN870 (IBK, VNMN); Ha Noi City: Ba Vi District, Ba Vi National Park, V.D. Nguyen et al. HNK1266 (K); Hoa Binh Province: Mai Chau District, Pa Co Municipality, V.X. Phuong 2397 (HN); Lao Cai Province: Sa Pa District, B. Khao & V. Trung 2545 (HN); Van Ban District, Khanh Yen Ha Municipality, 21°58′28″N 104°15′52″E, L. Averyanov et al. HAL2312 (LE, MO); Nghe An Province: Con Cuong District, Chau Khe Municipality, Pu Mat National Park, 18°55′51″N 104°36′20″E, M.S. Nuarliev et al. 2042 (MW); Quang Binh Province: Minh Hoa District, Dan Hoa Municipality, 17°41′50″N 105° 45′54″E, L. Averyanov et al. HAL12250, HAL12412 (LE); Son La Province: Van Ho District, Tan Xuan Municipality, 20°40′33″N 104°39′00″E, L. Averyanov et al. CPC7127 (LE).
Elatostema heyneanum (Wedd.) Hallier f.
Pellionia heyneana Wedd.
Cambodia, China, India, Indonesia, Sri Lanka, Thailand, Vietnam; 100–300 m.
Ha Noi City: Ba Vi District, Ba Vi Mountain, B. Balansa 2549 (P); Quang Binh Province: Tuyen Hoa District, Lam Hoa Municipality, 17°56′51″N 105°49′18″E, L. Averyanov et al. CPC2679, CPC2696 (LE); Quang Tri Province: Da Krong District, Ta Rut Municipality, 16°24′29″N 107°01′07″E, L. Averyanov et al. CPC3049 (LE).
Elatostema hookerianum Wedd., Arch. Mus. Hist. Nat. 9(1–2): 309. 1856, Type: Sikkim, J.D. Hooker s.n. (holotype: P!, isotype: MPU!).
Elatostema subfalcatum W.T.Wang.
Pellionia macroceras Gagnep., Bull. Soc. Bot. France 75: 922. 1928, syn. nov. Type: Vietnam. Lao Cai, Sa Pa District, P.A. Petelot 2294 (lectotype: P: P00601742!, designated here, isolectotype: P: P00601741!).
Bhutan, China, India, Sikkim, Vietnam; 1,200–2,100 m.
Lai Chau Province: Than Uyen District, Ho Mit Municipality, 22°06′N 103°52′E, N.T. Hiep et al. NTH2668, NTH2809, NTH2832, NTH2874 (LE); Lao Cai Province: Sa Pa District, Hoang Lien National Park, L.F. Fu et al. VMN_CN882 (IBK, VNMN), P.A. Petelot 2294 (P); Son La Province: Yen Chau District, Muong Lum Municipality, Lum Village, 21°00′17″N 104°29′08″E, D.K. Harder et al. 7217 (HN); Province unknown (Northern Vietnam): G.V. Mikeshin s.n. (MHA).
Note: The protologue of Pellionia macroceras (Gagnepain, 1928) includes a description of the species with two type localities of ‘Chapa’ (Vietnam) and Sikkim. The former relates to the collection P.A. Petelot 2294 with two duplicates housed at P (barcodes P00601741, P00601742), while the latter is represented by the collection J. de Rémy 1863 with a single specimen at P (barcode P00601743). All of these specimens contain the name ‘Pellionia macroceras Gagnep.’ written in Gagnapain’s handwriting and match the protologue description well. The dates of these collections (1925 and 1863) are before the publication of the protologue (1928), and therefore were used during its preparation. We select the specimen P.A. Petelot 2294 (P: P00601742) as the lectotype because it contains anatomical drawing with short notes made by Gagnepain; the other one (P: P00601741) is isolectotype. The status of the specimen J. de Rémy 1863 (P: P00601743) can be indicated as a remaining syntype.
We place Pellionia macroceras in synonymy based on the same characters of falcate lamina with cordate base with Elatostema hookerianum.
Elatostema integrifolium (D.Don) Wedd.
Procris integrifolia D.Don.
Elatostema cuspidiferum Miq., E. miquelianum Wedd., E. pulgarense Elmer, E. sesquifolium (Reinw. ex Blume) Hassk., E. sesquifolium var. integrifolium (D.Don) Wedd., E. viridicaule W.T.Wang, E. zollingerianum Wedd., Procris sesquifolia Reinw. ex Blume.
Bhutan, China, India, Indonesia, Myanmar, Nepal, Thailand, Vietnam; 600–900 m.
Cao Bang Province: Bao Lac District, Dinh Phung Municipality, L.F. Fu et al. VMN_CN819 (IBK, VNMN); Hai Phong City: Cat Hai District, Y.G. Wei et al. VMN_CN06 (IBK, VNMN); Cat Hai District, Cat Ba Island, N. Arnautov et al. LX-VN3778 (LE); Hoa Binh Province: Lac Son District, Tu Do Municipality, 20°25′29″N 105°19′36″E, N.Q. Hieu et al. CPC1538 (LE); Kon Tum Province: Sa Thay District, Ro Koi Municipality, Chu Mom Ray National Park, 14°28′20″N 107°43′00″E, M.S. Nuraliev & S.P. Kuznetsova 1257 (IBK, MW); Lam Dong Province: Bao Loc District, L. Averyanov et al. III-LX-VN1183 (LE); Quang Tri Province: Da Krong District, Ta Rut Municipality, 16°24′29″N 107°01′07″E, L. Averyanov et al. CPC3039 (LE); Huong Hoa District, Huong Viet Municipality, 16°51′06″N 106°34′38″E, L. Averyanov et al. CPC2815 (LE).
Elatostema kimhyense Y.G.Wei & V.T.Do
Endemic; 350–1,200 m.
Bac Kan Province: Na Ri District, Kim Hy National Park, 22°11′56″N 106°04′05″E, Y.G. Wei & V.T. Do V-23 (IBK, PE, VNMN), L.F. Fu et al. VMN_CN194 (IBK, VNMN).
Elatostema laevissimum W.T.Wang
Elatostema laevissimum W.T.Wang var. puberulum W.T.Wang.
China, Vietnam; 780–2,200 m.
Ha Giang Province: Meo Vac District, Vicinities of Meo Vac Town, 23°10′N 105°24′E, N.T. Hiep et al. NTH3378, NTH3390 (LE); Quan Ba District, Tung Vai, L.F. Fu et al. VMN_CN871 (IBK, VNMN); between Meo Vac District and Dong Van District, 23°07′31″N 105°25′42″E, L.F. Fu et al. VMN_CN188 (IBK, VNMN); Kon Tum Province: Dak Glei District, 15°04′23″N 108°01′34″E, L. Averyanov et al. VH062 (HN, MO), 15°04′00″N 107°59′00″E, L. Averyanov et al. VH629 (HN, LE, MO), VH074, VH590 (LE, MO); Lai Chau Province: Than Uyen District, Ho Mit Municipality, 22°06′N 103°52′E, N.T. Hiep et al. NTH2812 (LE); Lao Cai Province: Van Ban District, 21°58′42″N 104°00′37″E, L. Averyanov et al. HAL2013 (LE); Nghe An Province: Que Phong District, Hanh Dich Municipality, Pu Hoat Nature Reserve, 19°44′15″N 104°48′59″E, M.S. Nuraliev 2192 (MW); Quang Nam Province: L. Averyanov et al. VH923 (LE).
Elatostema leiocarpum (W.T.Wang) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia leiocarpa W.T.Wang, Guihaia 2(3): 115. 1982. Type: China. Guangxi, Napo, Baidu, Nunghua, D. Fang et al. 25142 (holotype: GXMI!).
China, Vietnam; ca. 1,200 m.
Ha Giang Province: Quan Ba District, Bat Dai Son Nature Reserve, 23°09′18″N 105°00′09″E, D.K. Harder et al. 5029 (HN); Quan Ba District, Can Ty Municipality, 23°04′N 104°59′E, N.T. Hiep et al. NTH3618 (LE); Quan Ba District, San Chu village, 23°08′55″N 104°59′46″E, L. Averyanov et al. VR079 (LE); Quan Ba District, Tung Vai Municipality, Thang village, 23°03′41″N 104°50′42″E, L. Averyanov et al. VR575 (LE).
Elatostema lineolatum Wight
Elatostema lineolatum var. majus Wedd., E. tumidulum Koidz.
Bhutan, China, India, Myanmar, Nepal, Sikkim, Sri Lanka, Thailand; 150–1,300 m.
Cao Bang Province: Bao Lac District, Xuan Truong Municipality, L.F. Fu et al. VMN_CN809 (IBK, VNMN); Nguyen Binh District, Phia Oac—Phia Den Nature Reserve, 22°37′40″N 105°53′10″E, L.F. Fu et al. VMN_CN163 (IBK, VNMN); Dak Lak Province: Lak District, Bong Krang Municipality, Chu Yang Sin National Park, 12°25′00″N 108°23′00″E, M.S. Nuraliev et al. 925 (K, MW); Ha Giang Province: Bac Me District, Bac Me Nature Reserve, L.F. Fu et al. VMN_CN232 (IBK, VNMN); Gia Lai Province: K’Bang District, Son Lang Municipality, Kon Chu Rang Nature Reserve, 14°30′55″N 108°32′48″E, M.S. Nuraliev 2010 (MW), 14°30′59″N 108°32′27″E, M.S. Nuraliev 2020 (MW); Lang Son Province: Bac Son District, Tan Tri Municipality, L. Averyanov et al. HAL6787 (LE); Lao Cai Province: Sa Pa District, San Sa Ho Municipality, Hoang lien National Park, Cat Cat village area, 22°20′N 103°49′E, M.S. Nuraliev & D.D. Sokoloff 282 (MW); Van Ban District, Nam Xe Municipality, 22°02′35″N 103°57′23″E, D.K. Harder et al. 7010 (HN, LE); Nghe An Province: Con Cuong District, Chau Khe Municipality, Pu Mat National Park, 18°57′38″N 104°40′44″E, M.S. Nuraliev 2065 (MW); Tuong Duong District, Tam Quang Municipality, 19°03′09″N 104°36′53″E, L. Averyanov et al. HLF6758 (LE); Quang Binh Province: Bo Trach District, Hung Trach Municipality, 17°27′30″N 106°23′06″E, S.K. Wu et al. WP923 (HN); Vinh Phuc Province: N. Arnautov et al. LX-VN3985 (LE); Tam Dao District, 21°28′02″N 105°38′39″E, L.F. Fu et al. VMN_CN51 (IBK, VNMN).
Elatostema longibracteatum W.T.Wang
a. Elatostema longibracteatum var. glabricaule W.T.Wang
Endemic; 650–800 m.
Cao Bang Province: Tra Linh District, Quoc Toan Municipality, L. Averyanov et al. VH2468 (HN, LE); 22°45′25″N 106°17′58″E, D.K. Harder et al. 4288 (K); Tra Linh District, Luu Ngoc Municipality, 22°47′N 106°18′E, L. Averyanov et al. CBL1199 (LE, MO).
This variety is very similar to Elatostema longibracteatum W.T.Wang var. longibracteatum and further study may result in it being treated as a synonym.
Elatostema longipedunculatum (W.T.Wang) Y.H.Tseng & A.K.Monro, comb. nov.
Basionym: Pellionia longipedunculata W.T.Wang, Bull. Bot. Res., Harbin 2(1): 1. 1982. Type: China. Guangxi, Shangsi County, Shiwandashan, S.C. Chen 4918 (holotype: IBSC!).
Pellionia subundulata W.T.Wang var. angustifolia W.T.Wang.
China, Vietnam; 197–1,000 m.
Ha Noi City: Ba Vi District, Ba Vi Mountain, 21°04.096′N 105°21.501′E, N.A. Vislobokov & S.P. Kuznetsova 681 (MW); Ha Tinh Province: Huong Son District, Rao An-Ngam Thep village, 18°21′53″N 105°13′12″E, T.H. Nguyen et al. 184 (MO); Vinh Phuc Province: Tam Dao District, Sino-Vietnam Expedition 1996A (PE).
Elatostema longistipulum Hand.-Mazz.
China, Vietnam; 140–1,450 m.
Cao Bang Province: Ha Lang District, Thang Loi Municipality, 22°45′N 106°42′E, P.K. Loc et al. CBL1661 (LE, MO); Ha Giang Province: Bac Me District, Bac Me Nature Reserve, 22°38′37″N 105°28′36″E, L.F. Fu et al. VMN_CN220 (IBK, VNMN); Ha Noi City: Ba Vi District, Ba Vi National Park, V.D. Nguyen et al. HNK1282 (K); Hoa Binh Province: Lac Son District, Tu Do Municipality, 20°24′57″N 105°13′16″E, N.Q. Hieu et al. CPC1403 (LE); Mai Chau District, F. Wen et al. VMN_CN274, VMN_CN317, VMN_CN323 (IBK, VNMN); Lang Son Province: Huu Lung District, Huu Lien Municipality, 21°40′59″N 106°20′30″E, D.K. Harder 4116 (MO); Nghe An Province: Con Cuong District, Chau Khe Municipality, Pu Mat National Park, 18°57′42″N 104°41′40″E, M.S. Nuraliev 2077 (MW); Quy Hop District, Chau Thai Municipality, Bom Village, V.D. Nguyen et al. HNK1611 (K); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2110 (TAI); L. Averyanov et al. LX-VN1732 (LE), V.D. Nguyen et al. HNK1334 (K); Phu Tho Province: Tan Son District, Xuan Son Municipality, 21°02′55″N 104°56′51″E, L. Averyanov et al. HAL12722 (LE), 21°06′57″N 104°57′17″E, L. Averyanov et al. HAL12677 (LE); Quang Binh Province: G.V. Mikeshin, O.V. Daeva s.n. (MHA); Son La Province: Van Ho District, Tan Xuan Municipality, 20°40′33″N 104°39′00″E, L. Averyanov et al. CPC7143 (LE); Yen Chau District, Muong Lum Municipality, 21°00′35″N 104°29′06″E, L. Averyanov et al. CPC1884 (LE); Thai Nguyen Province: Vo Nhai District, Than Sa Municipality, 21°48′N 105°54′E, N.T. Hiep et al. NTH3780 (LE); Thanh Hoa Province: Quy Chau District, Pu Huong Natural Reserve, F. Wen et al. VMN_CN428 (IBK, VNMN); Quan Hoa District, Hien Chung Municipality, Xuan Lien Reserve, F. Wen et al. VMN_CN398, VMN_CN410 (IBK, VNMN); Quan Hoa District, Pu Hu Nature Reserve; F. Wen et al. VMN_CN350 (IBK, VNMN); Thuong Xuan District, Bat Mot Municipality, 19°59′11″N 104°58′32″E, L. Averyanov et al. CPC6440 (LE); Tuyen Quang Province: Na Hang District, Vinh Yen Municipality, 22°21′14″N 105°25′11″E, P.K. Loc et al. HLF-009 (HN).
Elatostema macintyrei Dunn
Bhutan, China, India, Thailand, Vietnam; 1,000–1,100 m.
Cao Bang Province: Bao Lac District, Xuan Truong Municipality, L.F. Fu et al. VMN_CN807 (IBK, VNMN); Gia Lai Province: Mang Yang District, A Yun Municipality, Kon Ka Kinh National Park, 14°13′17″N 108°18′59″E, M.S. Nuraliev et al. 1492 (IBK, MW), M.S. Nuraliev 1882 (IBK, MW); Ha Giang Province: between Meo Vac District and Dong Van District, 23°09′06″N 105°24′59″E, L.F. Fu et al. VMN_CN185 (IBK, VNMN).
*Elatostema malacotrichum W.T.Wang & Y.G.Wei
China, Vietnam; 400 m.
Cao Bang Province: Ha Lang District, An Lac Municipality, 22°46′N 103°44′E, L. Averyanov et al. CBL607 (LE); Ha Quang District, Pac Bo, L.F. Fu et al. VMN_CN784 (IBK, VNMN).
Elatostema monandrum (Buch.-Ham. ex D.Don) H.Hara
Procris monandra Buch.-Ham. ex D.Don.
Elatostema diversifolium Wedd., E. laetum Wedd., E. monandrum var. ciliatum (Hook.f.) Murti, E. monandrum var. pinnatifidum (Hook.f.) Murti, E. monandrum var. serpens (Hook.f.) Murti, E. monandrum var. zeylanicum (Hook.f.) Murti, E. muscicola W.T.Wang, E. surculosum Wight, Pellionia mairei H.Lév.
Bhutan, China, India, Myanmar, Nepal, Sikkim, Sri Lanka, Thailand, Vietnam; 2,000–2,300 m.
Lao Cai Province: Sa Pa District, P.A. Petelot 5049 (P); Nghe An Province: Ky Son District, Na Ngoi Municipality, 19°11′58″N 104°11′39″E, L. Averyanov et al. CPC6150 (LE); 19°12′23″N 104°11′34″E, L. Averyanov et al. CPC6215 (LE).
Elatostema myrtillus (H.Lév.) Hand.-Mazz., Symb. Sin. 7(1): 146. 1929. Types: China. Pin-Fa, J. Cavalerie 559 (syntype: P!); Kouy-Tchéou, E. Bodinier 1548 (syntype: E); Kan-Yen-Tong, J. Esquirol 698 (syntype: E).
Pellionia myrtillus H.Lév.
Elatostema colaniae Gagnep., Bull. Soc. Bot. France 76: 81. 1929, syn. nov. Type: Vietnam. Lang Met, P.A. Colani 2927 (holotype: P!, isotype: P!).
China, Vietnam; 500–1,600 m.
Bac Giang Province: Lang Met, P.A. Colani 2927 (P); Cao Bang Province: Tra Linh District, Quoc Toan Municipality, vicinity of Thang Heng and Lung Tao Villages, 22°45′30″N 106°17′22″E, L. Averyanov et al. VH4855 (HN, LE); Tra Linh District, Quoc Toan Municipality, 22°45′15″N 106°17′54″E, D.K. Harder et al. 4312 (K); Ha Giang Province: between Meo Vac District and Dong Van District, 23°15′02″N 105°23′09″E, L.F. Fu et al. VMN_CN197 (IBK, VNMN); Meo Vac District, Sung Cha Municipality, 23°11′N 105°17′E, N.T. Hiep et al. NTH3287 (K, LE); Meo Vac District, Sung Cha Municipality, 23°10′12″N 105°17′39″E, L. Averyanov et al. HAL8490 (LE); Quan Ba District, Bat Dai Son Nature Reserve, Thanh Van commune, Mo Sai village, 23°07′46″N 104°57′56″E, L. Averyanov et al. VR409 (LE); Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN869 (IBK, VNMN); Quan Ba District, Tung Vai Municipality, Thang village, 23°03′13″N 104°51′49″E, L. Averyanov et al. VR742 (LE).
We place Elatostema colaniae in synonymy based on the same combination of characters as in E. myrtillus: small lamina (13–28 × 6–10 mm) with auriculate base and furfuraceous stem base.
*Elatostema nanchuanense W.T.Wang
China, Vietnam; 1,050–1,150 m.
Ha Giang province: Quan Ba District, Tung Vai Municipality, Thang village, 23°03′13″N 104°51′49″E, L. Averyanov et al. VR552 (LE).
Elatostema nasutum Hook.f.
Elatostema hainanense W.T.Wang, E. stipulosum Hand.-Mazz.
Bhutan, China, Nepal, Vietnam; 1,100–2,100 m.
Cao Bang Province: Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL077, CBL171 (LE, MO); Nguyen Binh District, Phia Oac–Phia Den Nature Reserve, 22°36′29″N 105°52′18″E, L.F. Fu et al. VMN_CN162 (IBK, VNMN); Ha Giang Province: Vi Xuyen District, Cao Bo Municipality, Tam Ve village, 22°46′30″N 104°49′01″E, D.K. Harder 5425 (MO); Yen Minh District, Du Gia Municipality, [23°56′N 105°13″E], P.K. Loc et al. CBL2028, CBL2080 (LE, MO); Kon Tum Province: Dak Glei District, N.T. Hiep et al. VH1757 (LE); Lai Chau Province: Than Uyen District, Ho Mit Municipality, 22°06′N 103°52′E, N.T. Hiep et al. NTH2669, NTH2812, NTH2874 (LE); Lao Cai Province: Sa Pa District, San Sa Ho, Sin Chai Municipality, 22°20′44.3″N 103°48′77.0″E, V.X. Phuong et al. HNK470 (K); Nghe An Province: Ky Son District, Na Ngoi Municipality, 19°11′59″N 104°11′24″E, N.T. Hiep et al. CPC6286 (LE); Son La Province: Bac Yen District, Chu Village, 21°15′40.0″N 104°29′27.0″E, V.D. Nguyen et al. HNK2726 (K); Thanh Hoa Province: Thuong Xuan District, Bat Mot Municipality, 19°58′18″N 104°59′24″E, L. Averyanov et al. CPC6555 (LE); Vinh Phuc Province: Tam Dao District, Tam Dao Range, N. Arnautov et al. LX-VN3858 (LE).
Elatostema neobaviense Y.H.Tseng & A.K.Monro nom. nov.
non E. baviense Gagnep.
Pellionia baviensis Gagnep., Bull. Soc. Bot. France 75: 919. 1928. Type: Vietnam. Ha Noi City, Ba Vi District, Ba Vi Mountain, B. Balansa 2548 (lectotype: designated by Lin & Yang, 2010: 1912, P!).
Endemic; elevation unknown.
Ha Noi City: Ba Vi District, Ba Vi Mountain, B. Balansa 2548 (P).
We select ‘neobaviense’ as epiphet as it takes the place of ‘baviense’ which would be in conflict with the homonym Elatostema baviensis.
Elatostema neriifolium W.T.Wang & Z.Y.Wu
China, Vietnam; 50–750 m.
Lao Cai Province: Sino-Vietnam Expedition 755 (PE); Quang Binh Province: Minh Hoa District, Thuong Hoa Municipality, 17°39′22″N 105°54′59″E, N.T. Hiep et al. CPC3819 (LE); Thanh Hoa Province: Ba Thuoc District, Co Lung Municipality, Khuyn village, 20°26′39″N 105°15′33″E, L. Averyanov et al. 3130 (MO); 10 km SW of Ba Thuoc, B.A. Korotiaev s.n. (LE); Tuyen Quang Province: Na Hang District, Thanh Tuong Municipality, 22°16′26″N 105°25′58″E, P.K. Loc et al. HAL149 (HN).
*Elatostema oblongifolium S.H.Fu
Pellionia bodinieri H.Lév.
Elatostema multicanaliculatum B.L.Shih & Yuen P.Yang.
China, Vietnam; 1,050–1,400 m.
Ha Giang province: Quan Ba District, Tung Vai Municipality, Thang village, 23°03′13″N 104°51′49″E, L. Averyanov et al. VR560 (LE), 23°03′42″N 104°50′42″E, L. Averyanov et al. VR634 (LE).
*Elatostema obtusum Wedd.
Dorstenia pubescens Blanco, Elatostema delicatulum Wedd.
Bhutan, China, India, Nepal, Sikkim, Thailand, Vietnam; 1,400–1,900 m.
Cao Bang Province: Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL206 (LE, MO, P); Lao Cai Province: Sa Pa District, Hoang Lien National Park, Ton Forest Station, L.F. Fu et al. VMN_CN971 (IBK, VNMN).
*Elatostema oppositum Q.Lin & Y.M.Shui
China, Vietnam; ca. 1,100 m.
Ha Giang Province: Quan Ba District, 23°06′57″N 105°01′48″E, D.K. Harder et al. 4973 (HN, MO), 4997 (HN).
Elatostema paucidentatum H.Schroet.
a. Elatostema paucidentatum var. paucidentatum
Pellionia paucidentata (H.Schroet.) Chien.
Elatostema henryanum var. oligodontum Hand.-Mazz., Pellionia longgangensis W.T.Wang, P. paucidentata var. hainanica Chien & S.H.Wu, P. subundulata W.T.Wang.
China, Vietnam; 180–300 m.
Cao Bang Province: Bao Lam District, Bac Mieu Municipality, L.F. Fu et al. VMN_CN827 (IBK, VNMN); Ha Giang Province: between Meo Vac District and Dong Van District, L.F. Fu et al. VMN_CN240 (IBK, VNMN); Lai Chau Province: Muong Te District, Muong Te Municipality, 22°32′06″N 102°32′29″E, N.T. Hiep et al. HAL10102, HAL10123 (LE); Lang Son Province: Lung District, Huu Lien Municipality, 21°39′31″N 106°22′06″E, D.E. Atha et al. NY-HN 73 (MO); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, 20°17′33″N 105°39′54″E, D.D. Soejarto 10498 (MO), 20°17.55′N 105°39.90′E, N.T. Hiep et al. 1998 (P); Quang Ninh province: Dam Ha District, Lomg Ngong village, Sai Vong Mo Leng mt., W.T. Tsang 30277 (SING); Ha-coi, near Chuk-phai, Taai Wong Mo Shan, W.T. Tsang 29166 (SING); Thai Nguyen Province: Vo Nhai District, Thuong Lung Municipality, 21°48.75′N 105°58.09′E, L.Q. Li et al. 0078 (HN); Thanh Hoa Province: Quan Hoa District, Hien Chung Municipality, Xuan Lien Nature Reserve, F. Wen et al. VMN_CN413 (IBK, VNMN); Vinh Phuc Province: N. Arnautov et al. LX-VN3988 (LE).
b. Elatostema paucidentatum var. dolichoceras W.T.Wang
Endemic; 100–150 m.
Lao Cai Province: Sino-Vietnam Expedition 590 (PE).
*Elatostema pergameneum W.T.Wang
China, Vietnam; 100–250 m.
Lang Son Province: Huu Lung District, Huu Lien Municipality, 21°40′59″N 106°20′30″E, D.K. Harder et al. 4121 (K); Quan Binh Province: Minh Hoa District, 17°40′N 105°57′E, L. Averyanov et al. VH4611 (HN, LE).
Elatostema petelotii Gagnep.
China, Vietnam; 260–1,300 m.
Cao Bang Province: Phia Oac, P.A. Petelot 720 (P); Ha Giang Province: between Meo Vac District and Dong Van District, L.F. Fu et al. VMN_CN239 (IBK, VNMN); Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN868 (IBK, VNMN); Quan Ba District, Thang village, around point 23°03′13″N 104°51′49″E, L. Averyanov et al. VR540, VR731 (LE); Yen Minh District, Du Gia Municipality, 23°56′N 105°13′E, P.K. Loc et al. CBL2022, CBL2024 (LE); Nghe An Province: Que Phong District, Hanh Dich Municipality, Pu Hoat Nature Reserve, 19°45′19″N 104°47′45″E, M.S. Nuraliev 2142 (MW); Phu Tho Province: Thanh Son District, Xuan Son Municipality, V.X. Phuong 3930 (HN).
Elatostema platyphyllum Wedd., Arch. Mus. Hist. Nat. 9: 301. 1856. Type: India. Khasia, J.D. Hooker & T. Thomoson s.n. (lectotype: designated by Lin, Duan & Yang, 2009: 1912, K!).
E. platyphylloides B.L.Shih & Yuen P.Yang, E. platyphyllum Wedd. var. polycephalum Hara.
Elatostema baviensis Gagnep., Bull. Soc. Bot. France 76: 80. 1929, syn. nov. Type: Vietnam. Ha Noi City, Ba Vi District, Ba Vi Mountain, B. Balansa 2537 (holotype: P!).
Bhutan, China, India, Nepal, Philippines, Vietnam; 800–1,669 m.
Gia Lai Province: K’Bang District, Song Lang Municipality, Kon Chu Rang Nature Reserve, 14°31′04″N 108°36′23″E, M.S. Nuraliev et al. 1996 (MW); Ha Giang Province: between Meo Vac District and Dong Van District, 23°13′44″N 105°24′48″E, L.F. Fu et al. VMN_CN191 (IBK, VNMN); Ha Noi City: Ba Vi District, Ba Vi National Park, 21°04′23″N 105°21′36″E, L.F. Fu et al. VMN_CN33 (IBK, VNMN), Ba Vi District, Ba Vi Mountain, B. Balansa 2537 (P); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2122 (TAI); Thanh Hoa Province: Quan Hoa District, Hien Chung Municipality, Xuan Lien Nature Reserve, F. Wen et al. VMN_CN399, VMN_CN408 (IBK, VNMN).
We place Elatostema baviensis in synonymy based on the same combination of characters as in E. platyphyllum: large lamina (10–25 × 4–7.5 cm) with auriculate base, large stipule (15–25 × 3–5.5 mm) and pistillate inflorescence with short peduncle and obvious receptacle.
*Elatostema prunifolium W.T.Wang
China, Vietnam; elevation unknown.
Ha Giang Province: Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN866 (IBK, VNMN).
Elatostema pseudodissectum W.T.Wang
China, Vietnam; 1,000–1,300 m.
Cao Bang Province: Bao Lac District, Ca Thanh Municipality, 22°44′N 105°50′E, L. Averyanov et al. CBL389 (LE, MO); Vinh Phuc Province: Tam Dao District, Tam Dao National Park, 21°27′47″N 105°38′68″E, L.Q. Li et al. 559 (PE).
*Elatostema pseudolongipes W.T.Wang & Y.G.Wei
China, Vietnam; 150–600 m.
Bac Kan Province: Ba Be District, Nam Mau Municipality, Ba Be National Park, 22°26.875′N 106°37.028′E, D.E. Atha et al. DA4722 (HN); Na Ri District, Liem Thuy Municipality, 21°57.36′N 106°04.622′E, D.E. Atha et al. DA4773 (HN), Na Ri District, Kim Hy Municipality, 22°17′N 106°03′E, N.T. Hiep et al. NTH3688, NTH3691 (LE); Cao Bang Province: Tra Linh District, Quoc Toan Municipality, Thang Heng vicinity and Lung Tao Villages, 22°45′30″N 106°17′22″E, L. Averyanov et al. VH4850, VH4928, VH4929 (HN, LE, MO); Ha Giang Province: Bac Me District, Bac Me Nature Reserve, 22°45′23″N 105°13′46″E, L.F. Fu et al. VMN_CN221 (IBK, VNMN); Ninh Binh Province: Cuc Phuong National Park, L. Wu et al. VN0049 (HN, IBK); Quang Binh Province: Bo Trach District, Son Trach Municipality, 17°31′02″N 106°16′48″E, L. Averyanov et al. HAL6382 (LE); Minh Hoa District, Thuong Hoa Municipality, 17°39′12″N 105°54′53″E, N.T. Hiep et al. CPC3753 (LE); Tuyen Hoa District, Lam Hoa Municipality, 17°56′10″N 105°49′15″E, L. Averyanov et al. CPC2452 (LE); Thanh Hoa Province: Ba Thuoc District, Co Lung Municipality, Eo Dieu Village, 22°25′35″N 105°14′26″E, L. Averyanov et al. HAL3354 (HN).
* Elatostema pycnodontum W.T.Wang
China, Vietnam; 1,400–1,500 m.
Cao Bang Province: Bao Lac District, Dinh Phuong Municipality, 22°47′N 105°49′E, P.K. Loc et al. CBL1489 (LE, MO).
Elatostema radicans (Siebold & Zucc.) Wedd.
Procris radicans Siebold & Zucc.
Elatostema cavaleriei (H.Lév.) Hand.-Mazz., E. radicans var. grande (Gagnep.) H.Schroet., Pellionia arisanensis Hayata, P. cavaleriei H.Lév., P. chikushiensis Yamam., P. iseana (Makino ex Hatus.) Hatus. ex Kimura, P. pauciflora W.T.Wang, P. radicans (Siebold & Zucc.) Wedd., P. radicans f. grandis Gagnep., P. radicans var. chikushiensis (Yamam.) S.S.Ying, P. radicans var. grandis (Gagnep.) W.T.Wang.
China, Japan, Korea, Vietnam; 600–2,700 m.
Cao Bang Province: Nguyen Binh District, Nguyen Binh Municipality, 22°37′N 105°52′E, L. Averyanov et al. CBL064 (HN, LE, MO); Nguyen Binh District, Phia Oac—Phia Den Nature Reserve, 22°36′29″N 105°52′18″E, L.F. Fu et al. VMN_CN160 (IBK, VNMN); Gia Lai Province: K’Bang District, Son Lang Municipality, Kon Chu Rang Nature Reserve, 14°31′04″N 108°36′23″E, M.S. Nuraliev et al. 1994 (MW), 14°30′59″N 108°32′27″E, M.S. Nuraliev et al. 2019 (MW), 14°30′02″N 108°34′27″E, M.S. Nuraliev et al. 2027 (MW); Ha Giang Province: Meo Vac District, vicinities of Meo Vac Town, 23°10′N 105°24′E, N.T. Hiep et al. NTH3377 (LE); Khanh Hoa Province: Khanh Son District, 12°12′N 108°44′E, L. Averyanov et al. VH4353 (LE, MO); Kon Tum Province: L. Averyanov et al. VH543 (LE, MO); Lam Dong Province: Lac Duong District, Da Chais Municipality, 12°09′N 108°41′E, L. Averyanov et al. VH2890, VH2914 (LE), 12°05′N 108°40′E, L. Averyanov et al. VH2974 (HN, LE), 12°07′N 108°41′E, L. Averyanov et al. VH3435 (LE, MO), 12°06′N 108°39′E, L. Averyanov et al. VH4493 (HN, MO); Lang Son Province: Sino-Vietnam Expedition 1597 (PE); Lao Cai Province: Sa Pa District, Hoang Lien National Park, L.F. Fu et al. VMN_CN886 (IBK, VNMN); Van Ban District, 21°58′42″N 104°00′37″E, L. Averyanov et al. HAL2094 (LE), 21°59′04″N 104°15′08″E, L. Averyanov et al. HAL2094 (LE); Nghe An Province: Ky Son District, Na Ngoi Municipality, 19°11′52″N 104°10′55″E, L. Averyanov et al. CPC6098 (LE); Ninh Binh Province: Nho Quan District, Phuoc Binh, 12°07′N 108°41′E, L. Averyanov et al. VH3369 (HN, LE); Quang Binh Province: Minh Hoa District, Dan Hoa Municipality, 17°45′27″N 105°47′46″E, L. Averyanov et al. HAL12539 (LE); Quang Nam Province: L. Averyanov et al. VH919 (HN, LE); Quang Ninh Province: Ba Mun, 21°05′N 107°35′E, L. Averyanov & E. Kudriavtseva 221 (LE); Vinh Phuc Province: N. Arnautov et al. LX-VN3941 (LE); Tam Dao District, 21°28′02″N 105°38′39″E, L.F. Fu et al. VMN_CN50 (IBK, VNMN); Tam Dao National Park, N. Arnautov et al. LX-VN3859, LX-VN3920, LX-VN4045 (LE), L. Averyanov 65-A (LE).
Elatostema ramosum W.T.Wang
Elatostema ramosum var. villosum W.T.Wang.
China, Vietnam; 500–1,400 m.
Cao Bang Province: Nguyen Binh District, Yen Lac Municipality, 22°45′34″N 105°51′49″E, L. Averyanov et al. CPC5424 (LE); Ha Giang Province: Bac Me District, Bac Me Nature Reserve, 22°45′14″N 105°13′36″E, L.F. Fu et al. VMN_CN227 (IBK, VNMN); Quan Ba District, Bat Dai Son commune, Bat Dai Son Nature Reserve, San Chu village, 23°08′55″N 104°59′46″E, L. Averyanov et al. VR065 (LE); Hoa Binh Province: Lac Son District, Tu Do Municipality, 20°25′29″N 105°19′36″E, N.Q. Hieu et al. CPC1559 (LE); Ninh Binh Province: Cuc Phuong National Park, T.H. Nguyen NTH 3015 (MO); Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2065 (TAI).
Elatostema retrohirtum Dunn
China, Vietnam; 87–1,100 m.
Bac Kan Province: Na Ri District, Kim Hy Nature Reserve, 22°11′49″N 106°03′51″E, L.F. Fu et al. VMN_CN120 (IBK, VNMN); Gia Lai Province: K’Bang District, Son Lang Municipality, Kon Chu Rang Nature Reserve, 14°30′55″N 108°32′48″E, M.S. Nuraliev 2009 (MW); Ha Giang Province: Bac Me District, Bac Me Nature Reserve, 22°45′24″N 105°13′49″E, L.F. Fu et al. VMN_CN217 (IBK, VNMN); between Meo Vac District and Dong Van District, L.F. Fu et al. VMN_CN186, VMN_CN196, VMN_CN245 (IBK, VNMN); Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN864 (IBK, VNMN); Ha Noi City: Ba Vi District, Ba Vi National Park, 21°04′22″N 105°21′36″E, L.F. Fu et al. VMN_CN34 (IBK, VNMN), 21°03′35″N 105°21′50″E, M.S. Nuraliev 1917 (IBK, MW); Hai Phong City: Cat Hai District, Cat Ba Island, Cat Ba National Park, Frog pond, 20°48.005′N 107°01.600′E, N.A. Vislobokov 13030 (MW); Hoa Binh Province: Lac Son District, Ngoc Son Municipality, 20°26′31″N 105°19′32″E, N.Q. Hieu et al. CPC1615 (LE); Lam Dong Province: Bao Loc, L. Averyanov et al. LX-VN1713 (LE); Nghe An Province: Que Phong District, Hanh Dich Municipality, Pu Hoat Nature Reserve, 19°46′19″N 104°47′00″E, M.S. Nuraliev 2180 (MW); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2099 (TAI); Son La Province: Phu Yen District, Gia Phu Municipality, Nhet Village, 21°36′27.7″N 104°04′22.8″E, V.D. Nguyen et al. HNK2469 (K); Tuyen Quang Province: Na Hang District, Vinh Yen Municipality, 22°21′07″N 105°25′06″E, N.T. Hiep et al. HLF-229 (HN).
Elatostema rhizomatosum (Gagnep.) Q.Lin
Pellionia rhizomatosa Gagnep.
Endemic; ca. 500 m.
Province unknown: Annam, col des Nuages prés Tourane, M. Poilane 7909 (P).
Elatostema rupestre (Buch.-Ham. ex D.Don) Wedd.
Procris rupestris Buch.-Ham. ex D.Don.
Elatostema zollingerianum Miq.
India, Nepal, Vietnam; elevation unknown.
Tuyen Quang Province: Na Hang District, A. Gramain et al. 649 (HN).
*Elatostema salvinioides W.T.Wang
a. Elatostema salvinioides var. robustum W.T.Wang
China, Vietnam; 1,100–1,650 m.
Dien Bien Province: Muong Cha District, Mua Ngai Municipality, 21°53′46″N 103°10′07″E, L. Averyanov et al. CPC1048, CPC1920 (LE); Lai Chau Province: Sin Ho District, Ta Ngau Municipality, 22°18′N 103°15′E, N.T. Hiep et al. NTH2772 (LE).
Elatostema scabrum (Benth.) Hallier f.
Pellionia scabra Benth.
Elatostema pellioniifolium W.T.Wang, Pellionia cephaloidea W.T.Wang, P. scabra subvar. pedunculata Yamam., Polychroa scabra (Benth.) Hu.
China, Japan, Vietnam; 700–1,800 m.
Hoa Binh Province: Da Bac District, Cao Son Municipality, Buoi Truong station, 20°54′09″N 105°12′15″E, N.T. Hiep et al. HAL592 (HN); Kon Tum Province: L. Averyanov et al. VH255 (HN, LE, MO); Phu Tho Province: Tan Son District, Xuan Son Municipality, 21°06′49″N 104°56′03″E, L. Averyanov et al. HAL12749 (LE); Son La Province: Yen Chau District, Muong Lum Municipality, 20°59′06″N 104°28′46″E, D.K. Harder et al. 7298 (HN).
Elatostema simplicissimum Q.Lin
Pellionia imbricata Gagnep.
Endemic; 200–220 m.
Lai Chau Province: 22°03′N 103°09′E, N.T. Hiep et al. NTH2747 (LE); Muong Lay District, Nam Hang Municipality, 22°08′14″N 102°58′18″E, D.K. Harder et al. 5884 (K, LE).
Elatostema sinense H.Schroet.
Elatostema sinense var. trilobatum W.T.Wang.
China, Vietnam; ca. 500 m.
Ha Giang Province: Bac Me District, Bac Me Nature Reserve, L.F. Fu et al. VMN_CN216A (IBK, VNMN); Quan Ba District, Tung Vai Municipality, L.F. Fu et al. VMN_CN872 (IBK, VNMN); Hoa Binh Province: Mai Chau District, Pa Co Municipality, F. Wen et al. VMN_CN330, VMN_CN333 (IBK, VNMN); Son La Province: Bac Yen District, Ta Sua Municipality, Chu Village, 21°15′40.0″N 104°29′27.0″E, V.D. Nguyen et al. HNK2724 (K).
Elatostema sublineare W.T.Wang
China, Vietnam; elevation unkown.
Cao Bang Province: Bao Lac District, Dinh Phung Municipality, L.F. Fu et al. VMN_CN815 (IBK, VNMN); Lang Son Province: Van Lang District, Anon. 1036 (LE); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, J.M. Hu Hu2106 (TAI); Tuyen Quang Province: Na Hang District, Anon. TV-74 (HN).
Elatostema tenuicaudatum W.T.Wang
China, Vietnam; 240–1,700 m.
Cao Bang Province: Bao Lac District, Xuan Truong Municipality, L.F. Fu et al. VMN_CN808 (IBK, VNMN); Dak Lak Province: Krong Bong District, Cu Pui Municipality, 12°30′N 108°30′E, L. Averyanov et al. VH6036 (HN); Dien Bien Province: Muong Cha District, Mua Ngai Municipality, 21°53′46″N 103°10′17″E, L. Averyanov et al. CPC1921 (LE); Gia Lai Province: Mang Yang District, A Yun Municipality, Kon Ka Kinh National Park, 14°13′10″N 108°19′10″E, M.S. Nuraliev et al. 1827 (IBK, MW); Ha Giang Province: Vi Xuyen District, Cao Bo Municipality, Tam Ve village, 22°46′03″N 104°49′42″E, D.K. Harder et al. 5454 (MO), 22°46′12″N 104°49′33″E, D.K. Harder et al. 5370 (MO); Yen Minh District, Du Gia Municipality, [23°56′N 103°13′E], P.K. Loc et al. CBL2063 (LE); Hoa Binh Province: Lac Son District, Tu Do Municipality, 20°26′07″N 105°17′19″E, N.Q. Hieu et al. CPC1499 (LE); Khanh Hoa Province: Khanh Son District, 12°12′N 108°44′E, L. Averyanov et al. VH4313 (HN, LE, MO); Lam Dong Province: Lac Duong District, Da Chais Municipality, 12°09′N 108°41′E, L. Averyanov et al. VH2891 (HN, LE, MO), 12°07′N 108°41′E, L. Averyanov et al. VH3413 (LE), 12°11′N 108°43′E, L. Averyanov et al. VH4387 (LE, MO); Lao Cai Province: Van Ban District, Khanh Yen Ha Municipality, 21°57′59″N 104°14′06″E, L. Averyanov et al. HAL2462 (HN, LE); Van Ban District, Nam Xe Municipality, 22°01′27″N 104°00′25″E, D.K. Harder et al. DKH7060 (LE); Phu Tho Province: Tan Son District, Xuan Son Municipality, 21°06′57″N 104°57′17″E, L. Averyanov et al. HAL12639 (LE); Quang Nam Province: L. Averyanov et al. VH905 (HN, LE); Tuyen Quang Province: Na Hang District, Vinh Yen Municipality, 22°21′09″N 105°25′20″E, Le Van Cham et al. HLF-061 (HN); Vinh Phuc Province: Tam Dao District, 21°27′35″N 105°38′58″E, L.F. Fu et al. VMN_CN45 (IBK, VNMN); Tam Dao Range, N. Arnautov et al. LX-VN3921 (LE).
Elatostema trichosanthum (Gagnep.) H.Schroet.
Pellionia trichosantha Gagnep.
Endemic; elevation unknown.
Ha Noi City: Ba Vi District, Ba Vi Mountain, B. Balansa 2529 (P).
Elatostema tsoongii (Merr.) H.Schroet.
Polycharoa tsoongii Merr.
Pellionia cristulata Gagnep.
Cambodia, China, Vietnam; 115–1,100 m.
a. Elatostema tsoongii ssp. tsoongii
Bac Kan Province: Na Ri District, Kim Hy Nature Reserve, 22°18′01″N 106°01′38″E, L.F. Fu et al. VMN_CN139 (IBK, VNMN); Dak Lak Province: Krong Bong District, Cu Pui Municipality, 12°30′N 108°30′E, L. Averyanov et al. VH5928 (HN); Lak District, Bong Krang Municipality, Chu Yang Sin National Park, 12°23′50″N 108°21′00″E, M.S. Nuraliev 997 (K, MW); Gia Lai Province: Mang Yang District, A Yun Municipality, Kon Ka Kinh National Park, 14°12′53″N 108°18′56″E, M.S. Nuraliev et al. 1449 (IBK, MW); Ha Tinh Province: Huong Son District, Son Hong Municipality, 18°34′06″N 105°11′40″E, P.K. Loc et al. HAL5193 (HN, LE); Lam Dong Province: Bao Lam District, Lac Bac Municipality, 11°43′28″N 107°41′59″E, M.S. Nuraliev et al. s.n. (photo: MW); Lang Son Province: Huu Lung District, Huu Lien Municipality, Huu Lien Protected Area, 21°40′59″N 106°20′30″E, D.K. Harder et al. 4120 (K); Pac Mo District, Anon. 1149 (LE); Quang Binh Province: Minh Hoa District, Thong Hoa Municipality, vicinities of Yen Son Village, 17°40′N 105°57′E, L. Averyanov et al. VH4729 (HN, LE, MO); Quang Tri Province: Da Krong District, Ta Rut Municipality, 16°24′29″N 107°01′07″E, L. Averyanov et al. CPC3017 (LE); Lang Khoai, Polilane 10741 (P); Thanh Hoa Province: Quan Hoa District, Pu Hu Nature Reserve, F. Wen et al. VMN_CN352 (IBK, VNMN); Thua Thien-Hue Province: Phu Loc District, Bach Ma National Park, 16°14′11″N 107°44′28″E, N.T. Hiep et al. HLF1649 (HN, MO); Tuyen Quang Province: Na Hang District, Thanh Tuong Municipality, near Ban Bung Village, 22°16′26″N 105°25′58″E, P.K. Loc et al. HAL164 (HN); Vinh Phuc Province: Tam Dao National Park, N. Arnautov et al. LX-VN4005 (LE).
b. Elatostema tsoongii ssp. subpeltatum (Gagnep.) H.W.Li
Pellionia subpeltata Gagnep.
China, Vietnam; elevation unknown.
Bac Giang Province: Lang Met, P.A. Petelot 2929 (P).
Elatostema veronicoides (Gagnep.) H.Schroet.
Pellionia veronicoides Gagnep.
China, Vietnam; elevation unknown.
Hoa Binh Province: Da Bac District, Cho Bo, B. Balansa 2251 (P); Thanh Hoa Province: Pu Huong Natural Reserve, F. Wen et al. VMN_CN422 (IBK, VNMN).
Elatostema vietnamense Q.Lin & L.D.Duan
Endemic; elevation unknown.
Thua Thien-Hue Province: Phu Loc District, Bach Ma National Park, H.N. Qin et al. 156 (PE).
Elatostema xanthophyllum W.T.Wang
China, Vietnam; 400–1,450 m.
Bac Kan Province: Na Ri District, Kim Hy Nature Reserve, 22°11′56″N 106°04′05″E, L.F. Fu et al. VMN_CN134 (IBK, VNMN); Na Ri District, Kim Hy Nature Reserve, Y.G. Wei & V.T. Do VMN_CN24 (IBK, VNMN); Ha Giang Province: Quan Ba District, 23°06′57″N 105°01′47″E, D.K. Harder et al. 4830 (HN); Quan Ba District, Can Ti Municipality, 23°04′N 104°59′E, N.T. Hiep et al. NTH3614 (K); Quan Ba District, San Chu village, 23°08′55″N 104°59′46″E, L. Averyanov et al. VR063 (LE); Quan Ba District, Thanh Van commune, Mo Sai village, 23°07′46″N 104°57′56″E, L. Averyanov et al. VR394 (LE); Son La Province: Van Ho District, Tan Xuan Municipality, 20°40′33″N 104°39′00″E, L. Averyanov et al. CPC7117 (LE).
*Elatostema xichouense W.T.Wang
China, Vietnam; 300–1,100 m.
Ha Giang Province: Quan Ba District, Bat Dai Son Nature Reserve, 23°09′N 104°59′E, D.K. Harder et al. 5120 (HN), L. Averyanov et al. VR052 (LE), N.T. Hiep & L. Averyanov 5120 (MO); Quan Ba District, Bat Dai Son Nature Reserve, Thanh Van commune, Mo Sai village, 23°07′46″N 104°57′56″E, L. Averyanov et al. VR365 (LE); Meo Vac District, L.F. Fu et al. VMN_CN852 (IBK, VNMN); Hoa Binh Province: Lac Son District, Tu Do Municipality, 20°26′07″N 105°17′19″E, N.Q. Hieu et al. CPC1482 (LE); Mai Chau District, Van Mai Municipality, 20°35′26″N 105°02′00″E, D.K. Harder et al. 8170 (MO); Ninh Binh Province: Nho Quan District, Cuc Phuong National Park, L. Averyanov et al. LX-VN1728 (LE).
Excluded taxa
Elatostema cuneatum Wight
Elatostema approximatum Wedd., E. densiflorum Franch. & Sav., E. nipponicum Makino, E. webbianum Wedd.
Bhutan, China, India, Indonesia, Laos, Thailand.
This species was indicated by Ho (2003) and Hiep (2005) as occurring in Vietnam but no specimen was cited and we were unable to locate any specimen from Vietnam.
{kind=link}
{kind=link}

{kind=link}

{kind=link}

{kind=link}
{kind=link}
{kind=link}