Screening of MYB1R1 interaction with LDOX promoter to regulate anthocyanin biosynthesis in peaches
- Published
- Accepted
- Received
- Academic Editor
- Vladimir Uversky
- Subject Areas
- Agricultural Science, Molecular Biology, Plant Science
- Keywords
- Anthocyanin synthesis, Peach, LDOX promoter, MYB, NGS, Transcription factor, Yeast one-hybrid
- Copyright
- © 2025 Wu et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2025. Screening of MYB1R1 interaction with LDOX promoter to regulate anthocyanin biosynthesis in peaches. PeerJ 13:e19975 https://doi.org/10.7717/peerj.19975
Abstract
Background
The floral color variegation of cultivar ‘Sahong Tao’ is distinctive and possesses significant ornamental value. Currently, there are no relevant reports on how MYB transcription factors (TFs) interact with LDOX promoter to regulate the flower color variegation in peach.
Methods
In this study, we screened for proteins that interact with the LDOX promoter using yeast one-hybrid (Y1H) and next-generation sequencing (NGS). The NGS data were aligned with the Arabidopsis database (TAIR10) utilizing Python 3.10.4. PlantTFDB was employed to identify TFs, while PlantRegMap was used to predict TFs that interact with the LDOX promoter. The Y1H assay verified MYB1R1 interaction with LDOX promoter, and Y1H-AOS predicted their binding sites. The physicochemical properties, structure and interacting proteins of MYB1R1 were analyzed using bioinformatics methods. Sequence alignment and phylogenetic tree analyses of MYB1R1 were performed. Finally, the tissue expression specificity of MYB1R1 and LDOX in ‘Sahong Tao’ was examined using qRT-PCR.
Results
The Y1H and NGS results indicate that 1,190 proteins interact with the LDOX promoter. Among these, 20 TFs were identified, including ERF, MYB, NF-YB, SBP, S1Fa-like, TCP, bHLH, LBD, ZF-HD, C3H, DBB, MYB-related, and HD-ZIP. Of the 1,190 proteins, 1,146 exhibit high similarity to homologs in Arabidopsis, with 332 classified as RNA binding proteins and 124 as DNA binding proteins. A comparison with the NGS results identified seven TFs that align with predictions from PlantRegMap. Based on these findings, we selected MYB44 (PRUPE_6G229000, PRUPE_1G430000) and MYB1R1 (PRUPE_5G182000) as candidate members. Y1H assays demonstrated that MYB1R1 interacts with the LDOX promoter. Y1H-AOS was used to confirm 24 interaction binding sites. MYB1R1 consists of an 897 bp full-length CDS, encoding 298 amino acids, with a predicted molecular weight of 32.49 kDa and a theoretical isoelectric point of 7.20. MYB1R1 features a typical SANT-MYB domain, and its secondary structure is predominantly composed of irregular coils. Phylogenetic analysis indicates a close evolutionary relationship between MYB1R1 from ‘Sahong Tao’ and both Prunus avium and Prunus speciosa. Promoter prediction analysis for MYB1R1 reveals multiple hormone- and stress-related cis-acting elements. MYB1R1 may interact with bHLH and other proteins to perform its functions. In variegated petals, MYB1R1 expression is higher and LDOX expression is lower compared to red petals, suggesting that MYB1R1 negatively regulates anthocyanin synthesis by interacting with LDOX. This study contributes to elucidating the function of MYB1R1 and the regulatory mechanism of MYB- LDOX in the flower color of ‘Sahong Tao’.
Introduction
Peach (Prunus persica), a small deciduous tree belonging to the genus Prunus in the Rosaceae family, exhibits a rich diversity of floral colors. Variegation is a distinctive floral color pattern, characterized by red-white bicoloration in the peach cultivar ‘Sahong Tao’ (Chen et al., 2014). This phenotype holds significant ornamental value, and is notably observed in other horticultural plants such as Prunus mume, Rhododendron, Antirrhinum majus and Petunia hybrida (Nie et al., 2023; Piao, Wu & Cui, 2021; Wu et al., 2017; Yasumasa et al., 2012). The variegated phenotype of ‘Sahong Tao’ is primarily associated with the types, concentration, and distribution of anthocyanins in petal cells (Cheng et al., 2015).
Anthocyanins are water-soluble flavonoid pigments (Sun et al., 2016) that impart a diverse range of colors to plant tissues and cells (Khoo et al., 2017). Previous studies of ‘Sahong Tao’ have demonstrated that red petals possess significantly higher concentrations of anthocyanins compared to variegated petals, suggesting that the red coloration primarily arises from the accumulation of anthocyanins (Zhou et al., 2015b). The formation of anthocyanins is regulated by a collaborative interaction between structural genes and transcription factors (TFs) (Schwinn et al., 2016). Structural genes are classified into two categories: early biosynthetic genes, which include phenylalanine ammonia-lyase (PAL), cinnamate-4-hydroxylase (C4H), chalcone synthase (CHS), and chalcone isomerase (CHI). Late biosynthetic genes including dihydroflavonol-4-reductase (DFR), leucoanthocyanidin oxygenase/anthocyanidin synthase (LDOX/ANS), and UDP-glucose: flavonoid-3-O-glucosyltransferase (UFGT). This classification is based on their specific roles in the biosynthetic pathway (Chen et al., 2019; Sharma et al., 2024). The primary role of LDOX is to facilitate the conversion of colorless anthocyanins into pigmented anthocyanins (Owens & McIntosh, 2011; Zohar et al., 2015). In Arabidopsis thaliana, a reduction in the allelic mutant of LDOX results in decreased levels of anthocyanins and proanthocyanidins, leading to a lighter seed coat coloration of the seed coat (Bowerman et al., 2012). In Reaumuria trigyna, two significantly differentially expressed genes, RtLDOX and RtLDOX2, have been identified. RtLDOX2 enhances tolerance to abiotic stress by promoting the accumulation of anthocyanins and flavonols (Li et al., 2021), while the RtLDOX gene complements anthocyanidin synthesis (Zhang et al., 2016). In ‘Sahong Tao’, proteomic analyses have indicated that the protein abundance of LDOX is higher in red samples than in variegated samples (Zhou et al., 2015a). Further analysis demonstrates that LDOX gene expression and enzyme activity are significantly greater in red samples (Wu et al., 2020). These findings suggest that LDOX plays a critical role in the formation of variegated flowers in ‘Sahong Tao’. Currently, there is limited research on the interaction between MYB and the LDOX promoter in regulating anthocyanin synthesis in peach.
Various TFs participate in the anthocyanin biosynthesis pathway by regulating the expression levels of structural genes (Erika et al., 2015). Anthocyanin biosynthesis refers to the series of enzymatic reactions that lead to the formation of anthocyanin pigments from the precursor phenylalanine in plants (Sharma et al., 2024). These TFs can modulate anthocyanin synthesis either independently or by forming complexes, such as the ternary complex composed of an R2R3-MYB TF, a basic helix-loop-helix (bHLH) domain protein and a WD-repeat protein, collectively known as the MYB-bHLH-WD40 (MBW) complex (An et al., 2012; Liu et al., 2024a; Xie et al., 2012). MYB proteins play significant roles in regulating the biosynthesis of anthocyanin and flavonol compound in plants. A total of 137 R2R3-MYB TFs have been identified in A. thaliana. Among these, AtPAP1 (AtMYB75), AtPAP2 (AtMYB90), AtPAP3 (AtMYB113), and AtPAP4 (AtMYB114) have been confirmed to be involved in anthocyanin synthesis (Borevitz et al., 2000; Heppel et al., 2013; Nesi et al., 2001; Ramsay & Glover, 2005; Stracke et al., 2007), while AtMYB123 regulates proanthocyanidin biosynthesis (Lepiniec et al., 2006). Furthermore, the over-expression of PpMYB108 significantly enhances anthocyanin biosynthesis in tobacco flowers and interacts with the PpDFR promoter in peach (Khan et al., 2022). PpMYB15 and PpMYBF1 are implicated in the regulation of flavonol biosynthesis in peach fruit (Cao et al., 2019). ZeMYB32, identified as an R2R3-MYB, negatively regulates anthocyanin biosynthesis in Zinnia elegans (Jiang et al., 2024). Additionally, the over-expression of MYB10.1 in tobacco modulates reproductive anthocyanin biosynthesis (Rahim et al., 2019). PbMYB1L in the peel of ‘Red Zaosu’ pear acts as a positive regulator of anthocyanin biosynthesis (Zhou et al., 2024). However, the functions of MYB proteins related to anthocyanin synthesis in ‘Sahong Tao’ remain unknown.
The yeast one-hybrid (Y1H) technique is a molecular biology method employed to investigate protein-DNA interactions involving specific sequences. (Berenson et al., 2023; Sun et al., 2017). In lilies, the Y1H assay was utilized to screen ethylene response factors (ERFs), auxin/indole-3-acetic acid (AUX/IAA), and basic transcription factor 3 (BTF3) proteins that are involved in regulating anthocyanin biosynthesis (Yuwei et al., 2021). In Qingke (Hordeum vulgare L. var. Nudum Hook. f), HvANT2 exhibited promoter-binding activity towards anthocyanin-related genes such as CHI, dihydroflavonol-3′-hydrogenase (F3′H), and UDP-glucosyltransferase (GT) (Wang et al., 2024). In blueberries, VcMYB-1 and VcAN1 have been shown to bind to the VcGSTF8 promoter, thereby regulating anthocyanin biosynthesis (Zhang et al., 2024b). DcMYB11c binds to the promoters of DcUCGXT1 and DcSAT1, contributing to anthocyanin accumulation in carrot purple petioles (Duan et al., 2023). In apples, MdNAC33 activates the expression of MdbHLH3, MdDFR, and MdANS genes. Furthermore, MdNAC33 was found to interact with MdMYB1, a positive regulator of anthocyanin biosynthesis and accumulation (Zhang et al., 2024a).
The synthesis and accumulation of anthocyanins are essential for the formation of flower variegation in ‘Sahong Tao’. LDOX is a key gene that controls the biosynthesis of pigmented anthocyanins. However, the mechanisms underlying its regulatory network remain unclear. In this study, we identified MYB TFs that interact with the LDOX promoterusing the Y1H assay and next-generation sequencing (NGS) technology. The candidate MYB members were subsequently cloned and their interactions with the LDOX promoter were verified. These findings provide a foundation for a deeper understanding of the regulatory pathways involved in anthocyanin biosynthesis in peach.
Materials & Methods
Plant materials
The plant materials utilized in this study were derived from the peach cultivar ‘Sahong Tao’, which was cultivated under natural conditions in the experimental field of Huaiyin Institute of Technology (119°2′27″E, 33°33′13″N). Red and variegated flower buds were collected during the initial balloon stage. The variegated petals (petals-V), red petals (petals-R), leaves, red sepals (sepals-R), green sepals (sepals-G), pistils, and stamens were collected individually into centrifuge tubes. The samples were promptly frozen in liquid nitrogen and stored at −80 °C until further use.
Yeast one-hybrid screening and Next-generation sequencing technology analysis
The ‘Sahong Tao’ Y1H cDNA library and the LDOX promoter bait vector (pHIS2-LDOX-QDZ) have been constructed in our laboratory (Wu et al., 2024). Y1H screening assays were performed using the Clontech Yeast One-Hybrid System, provided by ProNet Biotech Co., Ltd (Nanjing, China). All positive clones from the yeast sieve plate were scraped into 2 × YPDA liquid, then centrifuged and collected. Colony PCR was performed, and the PCR products were sequenced using the Illumina NovaSeq 6000 platform (ProNet Biotech Co., Ltd, Nanjing, China), with a typical fragment length of 150 bp. Gene IDs, CDS and protein sequences were obtained by performing a BLAST search against the peach genome database downloaded from NCBI (https://ftp.ncbi.nlm.nih.gov/genomes/all/GCF/000/346/465/GCF_000346465.2_Prunus_persica_NCBIv2/). The NGS data were aligned with the Arabidopsis database (TAIR10) using Python 3.10.4 software, applying an e-value threshold of less than 1 × 10−10 to identify homologous genes. PlantTFDB version 5.0 (https://planttfdb.gao-lab.org/prediction.php) was used to determine whether the proteins were TFs (Supplementary Data S2). Additionally, PlantRegMap (https://plantregmap.gao-lab.org/binding_site_prediction.php) was used to predict TFs that interact with the LDOX promoter (Supplementary Data S3).
Candidate MYB members’ gene cloning
Total RNA was isolated from petals-V, petals-R, leaves, sepals-R, sepals-G, pistils, and stamens using the FastPure® Universal Plant Total RNA Isolation Kit (RC411, Vazyme, Nanjing, China). Then, the cDNA was synthesized using the Goldenstar™ RT6 cDNA Synthesis Kit (TSK301S, Tsingke, Beijing, China). Based on the results of NGS and predictions obtained from the PlantRegMap website, MYB44 (PRUPE_6G229000 and PRUPE_1G430000) and MYB1R1 (PRUPE_5G182000) were identified as candidate MYB members that may interact with the LDOX promoter. Using cDNA as template, Primer Premier 6.0 software was employed to design primers (Table 1) for cloning the full-length CDS sequences. The CDS sequences were cloned using the GoldenStar® T6 Super PCR Mix Ver.2 (1.1 ×) (TSE102, Tsingke, Beijing, China). PCR products were separated by 1% agarose gel electrophoresis, and the gel was observed and cut under using a multifunctional gel image analysis system (Tanon-1160, Tanon, Shanghai, China). Gel strips were purified according to the instructions of DNA Gel Extraction Kit (TSP601-50, Tsingke, Beijing, China). The target fragment was introduced into the pClone007 Blunt vector following the kit instructions (TSV-007B, Tsingke, Beijing, China). The ligation products were transformed into Escherichia coli DH5α competent cells (TSC-C01, Tsingke, Beijing, China), and positive clones were sent to Beijing Tsingke Biotech Co., Ltd. (Beijing, China) for sequencing.
| Primer name | Primer sequence (5′-3′) | Application |
|---|---|---|
| PRUPE_5G182000-C-F | ATGGCTGGCACGTGCTC | CDS full-length clone |
| PRUPE_5G182000-C-R | TCAAGCAACACTAATAATGCTATCCC | CDS full-length clone |
| PRUPE_1G430000-C-F | ATGGCTTCCACAAAGAA | CDS full-length clone |
| PRUPE_1G430000-C-R | CTACTCGATTTTGCTAATCC | CDS full-length clone |
| PRUPE_6G229000-C-F | ATGGCAGTGAGCAGAAAAGA | CDS full-length clone |
| PRUPE_6G229000-C-R | TCACTCGATCCTGCTAACAC | CDS full-length clone |
| pGADT7-F | TAATACGACTCACTATAGG | Yeast one-hybrid assay |
| pGADT7-R | GGCAAAACGATGTATAAATGA | Yeast one-hybrid assay |
| pHIS2-F | TTCGCTATTACGCCAGCTG | Yeast one-hybrid assay |
| pHIS2-R | GTTTATCTTGCCTGCTCATT | Yeast one-hybrid assay |
| PRUPE_5G182000-AD-F | GCCATGGAGGCCAGTGAATTCAT GGCTGGCACGTGCTC | Yeast one-hybrid assay |
| PRUPE_5G182000-AD-R | cagctcgagctcgatggatccTCAAGCAAC ACTAATAATGCTATCCC | Yeast one-hybrid assay |
| PRUPE_1G430000-AD-F | GCCATGGAGGCCAGTGAATTCATG GCTTCCACAAAGAA | Yeast one-hybrid assay |
| PRUPE_1G430000-AD-R | cagctcgagctcgatggatccCTACTCGA TTTTGCTAATCC | Yeast one-hybrid assay |
| PRUPE_6G229000-AD-F | GCCATGGAGGCCAGTGAATTCATG GCAGTGAGCAGAAAAGA | Yeast one-hybrid assay |
| PRUPE_6G229000-AD-R | cagctcgagctcgatggatccTCACTCGAT CCTGCTAACAC | Yeast one-hybrid assay |
| PRUPE_5G182000-E-F | ACCTTGGGACACGCAAATCT | qRT-PCR |
| PRUPE_5G182000-E-R | GATGACGATTCCCTCGGGTC | qRT-PCR |
| RPII-F | TGAAGCATACACCTATGATGATGAAG | qRT-PCR |
| RPII-R | CTTTGACAGCACCAGTAGATTCC | qRT-PCR |
| LDOX-F | GATGCAGGGAGGAGTTGAAG | qRT-PCR |
| LDOX-R | CTGCCCAGAAGCATTGTTTG | qRT-PCR |
Verification of LDOX promoter interaction with MYB
Y1H assay was performed to verify the interaction between the LDOX promoter and the candidate MYB members. The LDOX promoter sequence was constructed into the pHIS2 vector, while the CDS of candidate MYB members were inserted into the pGADT7 vector. Subsequently, the constructed vectors were transformed into the yeast strain Y187 through co-transfection and cultured on SD-TL plates. The experimental groups included pHIS2-LDOX-1+pGADT7-PRUPE_5G182000, pHIS2-LDOX-1+pGADT7-PRUPE_1G430000, and pHIS2-LDOX-1+pGADT7-PRUPE_6G229000. The control group was pHIS2-LDOX-1+pGADT7, while positive control group was pGAD53m+pHIS2-p53. The experimental, control and positive control groups were inoculated on SD-TL, SD-TLH, and SD-TLH+50 mM 3AT medium 30 °C constant temperature for 3–5 days. The interaction was confirmed based on the growth status of the yeast cells.
The predictive analysis of the MYB1R1-LDOX promoter interaction relationship
The 3D structures of MYB1R1 protein and LDOX promoter were constructed using AlphaFold2, with pLDDT values ranging from 0 to 100 to estimate prediction confidence (Jumper et al., 2021). The HDOCK (http://hdock.phys.hust.edu.cn/) was employed to obtain a confidence score, which indicates the likelihood of binding between two molecules. A confidence score exceeding 0.7 suggests a high probability of binding (Huang & Zou, 2014). Finally, PyMOL 3.1.4.1 software and PDBePISA (https://www.ebi.ac.uk/msd-srv/prot_int/pistart.html) were used to predict the binding sites of the corresponding protein sequences.
Bioinformatics analysis of MYB1R1
The MYB1R1 gene clone and sequencing information was utilized to perform a comprehensive bioinformatics analysis. The molecular weight, protein isoelectric point, instability index, aliphatic index, and grand average of hydropathicity were assessed using Expasy-ProtParam (https://web.expasy.org/protparam/). SignalP 4.1 (https://services.healthtech.dtu.dk/services/SignalP-4.1/) was utilized to forecast the signal peptide. Membrane structure analysis was conducted with TMHMM Server v2.0 (https://services.healthtech.dtu.dk/services/TMHMM-2.0/). Furthermore, prediction of subcellular localization of plant proteins using Plant-mPLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/), and phosphorylation site analysis was performed using NetPhos 3.1 (https://services.healthtech.dtu.dk/services/NetPhos-3.1/). The secondary and tertiary structures of the protein were predicted using SOPMA (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html) and SWISS-MODEL (https://swissmodel.expasy.org/interactive), respectively. Protein interaction analysis was carried out utilizing STRING (https://cn.string-db.org/).
Phylogenetic analysis and alignment of the deduced amino acid sequence of MYB1R1
The amino acid sequence of MYB1R1 was obtained from the NCBI database. Phylogenetic tree construction was carried out using MEGA 11 software, employing the neighbor-joining method (Tao et al., 2019). This phylogenetic analysis included homologous amino acid sequences of MYB1R1 from various plants, with peach serving as a reference sequence. The conserved domains were predicted and analyzed using the SMART (https://smart.embl.de/). Multiple sequence alignment and functional domain labeling were performed using DNAMAN V6 software.
MYB1R1 promoter analysis
MethPrimer (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi) was employed to identify CpG islands, criteria used: island size > 100, GC percent > 40.0, Obs/Exp > 0.6, while PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) was used to analyze the cis-acting elements within the 2 kb promoter sequence located upstream of the start codon of the MYB1R1 gene.
Analysis of the expression pattern of the MYB1R1 and LDOX
The expression profiles of MYB1R1 and LDOX in various tissues were detected and analyzed via qRT-PCR with ChamQ Blue Universal SYBR qPCR Master Mix (Q312, Vazyme, Nanjing, China). The total reaction volume was 20 µL, comprising 10 µL of 2 × ChamQ Blue Universal SYBR qPCR Master Mix, one µL of cDNA template, 0.4 µL of forward primer, 0.4 µL of reverse primer, and ddH2O to reach a final volume of 20 µL. The primer sequences are as listed in Table 1. Amplification was performed using the Bio-Rad CFX96 Real-Time PCR system (BIO-RAD, Hercules, CA, USA) with the following program: 1 cycle at 95 °C for 10 s, followed by 40 cycles at 95 °C for 5 s and 60 °C for 30 s. Dissolution curves were generated under the following conditions: 95 °C for 30 s, 95 °C for 10 s, 60 °C for 30 s, 70 °C for 2 s, and 95 °C for 2 s. RPII was utilized as the internal reference gene (Yan et al., 2012). The relative expression of MYB1R1 and LDOX was calculated using the 2−ΔΔCt method (Livak & Schmittgen, 2001).
Results
Screening out regulatory transcription factors interaction with LDOX promoter
Based on the Y1H assay and the results from NGS of positive clones, a total of 1,190 proteins were identified as interacting with the LDOX promoter (Data S1). The NGS data were compared using BLAST against the Arabidopsis database (TAIR10), resulting in the identification of 1,146 genes exhibiting high similarity to Arabidopsis. Using PlantRegMap, we predicted that 324 TFs bind to the LDOX promoter (Fig. 1A). Among the 1,190 proteins, 20 were categorized as TFs, which included four ERF (PRUPE_5G117800, PRUPE_4G176200, PRUPE_4G222300, PRUPE_3G032300), three MYB (PRUPE_2G192100, PRUPE_6G229000, PRUPE_1G430000), two NF-YB (PRUPE_6G130500, PRUPE_5G214400), two SBP (PRUPE_4G050400, PRUPE_1G224900), one S1Fa-like (PRUPE_6G239600), one TCP (PRUPE_5G088800), one bHLH (PRUPE_4G201500), one LBD (PRUPE_4G105700), one ZF-HD (PRUPE_3G267900), one C3H (PRUPE_3G080200), one DBB (PRUPE_1G398700), one MYB_related (PRUPE_5G182000), and one HD-ZIP (PRUPE_6G193400) (Fig. 1B). Of the total 1190 proteins analyzed, 332 were classified as RNA-binding proteins, while 124 were classified as DNA-binding proteins. Additionally, 13 proteins were found to belong to both RNA-binding and DNA-binding proteins (Fig. 1C). Comparison with the NGS results identified seven TFs that were consistent with PlantRegMap predictions, including three ERF (PRUPE_4G176200, PRUPE_4G222300, PRUPE_3G032300), one HD-ZIP (PRUPE_6G193400), one MYB_related (PRUPE_5G182000), two MYB (PRUPE_6G229000, and PRUPE_1G430000).
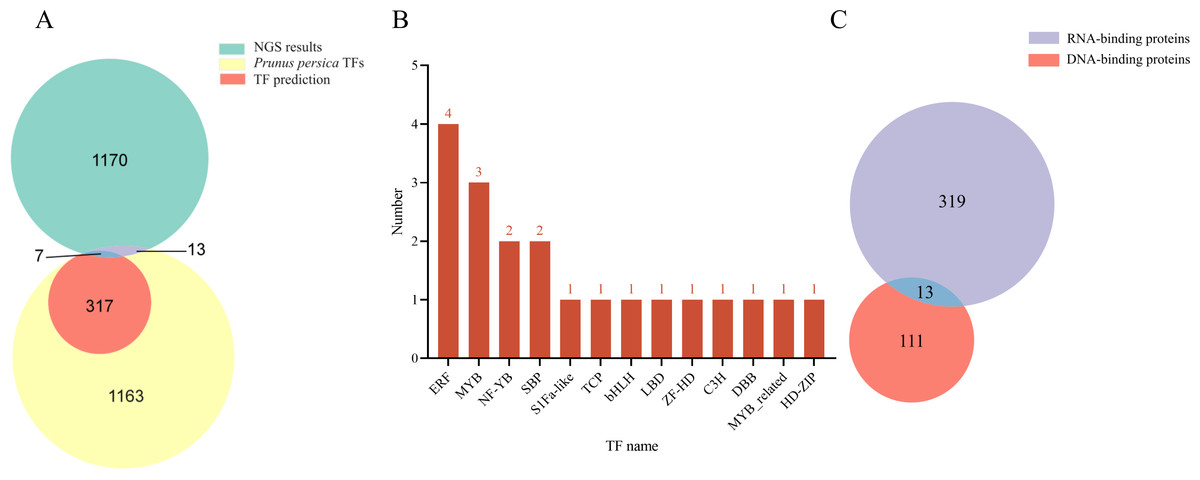
Figure 1: Analysis of the TFs screened out by NGS.
(A) Venn diagram comparing NGS results with Prunus persica TFs and predicted TFs for binding to LDOX promoter. (B) Distribution map illustrating the TFs with in each family. (C) Identification results for DNA-binding proteins and RNA-binding proteins.{kind=link}
Candidate MYB members’ gene cloning
The CDS sizes for PRUPE_5G182000, PRUPE_1G430000, and PRUPE_6G229000, were 897 bp (Fig. 2A), 939 bp (Fig. 2B), and 996 bp (Fig. 2C), respectively. Sanger sequencing results of the positive clone colonies aligned with the analysis of NGS results conducted using DNAMAN V6 software. The correct CDS were then employed to construct the prey vectors pGADT7-PRUPE_6G229000, pGADT7-PRUPE_1G430000, and pGADT7-PRUPE_5G182000, which were used to verify their interactions with the LDOX promoter.
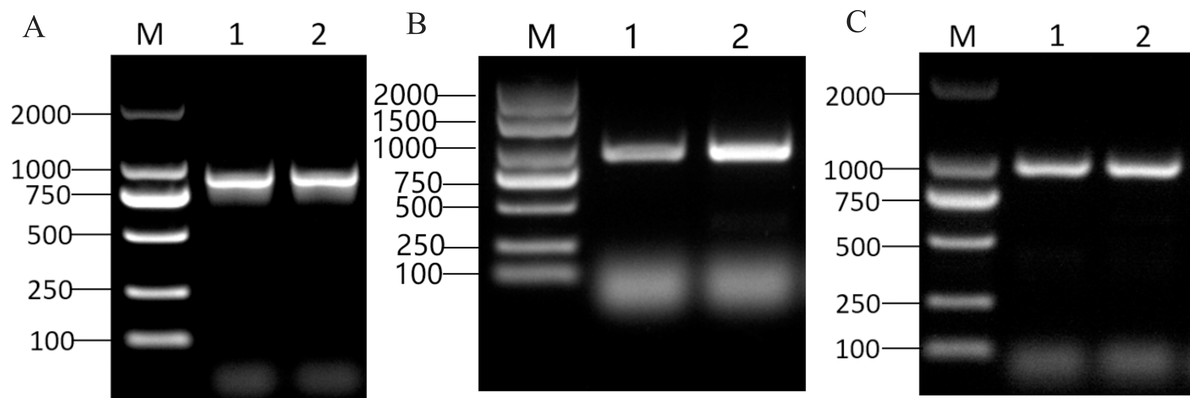
Figure 2: Cloning of candidate MYB TFs.
M: DNA marker DL2000. (A) 1–2: CDS clone of the PRUPE_5G182000, (B) 1–2: CDS clone of the PRUPE_1G430000, (C) 1–2: CDS clone of the PRUPE_6G229000.{kind=link}
Identification of MYB1R1 as a potential regulator of LDOX
The results indicated that the positive control exhibited normal growth (pGAD53m+pHIS2-p53) on SD-TL, SD-TLH, and SD-TLH+50 mM 3AT plates. In the control group (pHIS2-LDOX-1+pGADT7), normal growth was observed on SD-TL and SD-TLH plates. However, no growth occurred on SD-TLH+50 mM 3AT plates. The combination of pHIS2-LDOX-1+pGADT7-PRUPE_5G182000 displayed weak growth observed on SD-TLH+50 mM 3AT plates, indicating positive growth (Fig. 3A). Conversely, pHIS2-LDOX-1+pGADT7-PRUPE_1G430000 (Fig. 3B) and pHIS2-LDOX-1+pGADT7-PRUPE_6G229000 (Fig. 3C) did not exhibit growth on SD-TLH+50 mM 3AT plates. These results demonstrate that pGADT7-PRUPE_5G182000 interacts with pHIS2-LDOX-1.

Figure 3: Interaction validation results of pGADT7-PRUPE_5G182000 (A), pGADT7-PRUPE_1G430000 (B), and pGADT7-PRUPE_6G229000 (C) with pHIS2-LDOX-1.
(+): pHIS2-P53+pGAD53m as positive control, (-): pHIS2-LDOX-1+pGADT7 as negative control. SD-TL: -trp, -leu; SD-LH: -leu, -his; SD-TLH: -trp, -leu, -his. 3AT: a competitive inhibitor of yeast HIS3 protein synthesis, used to inhibit leaky expression of the HIS3 gene.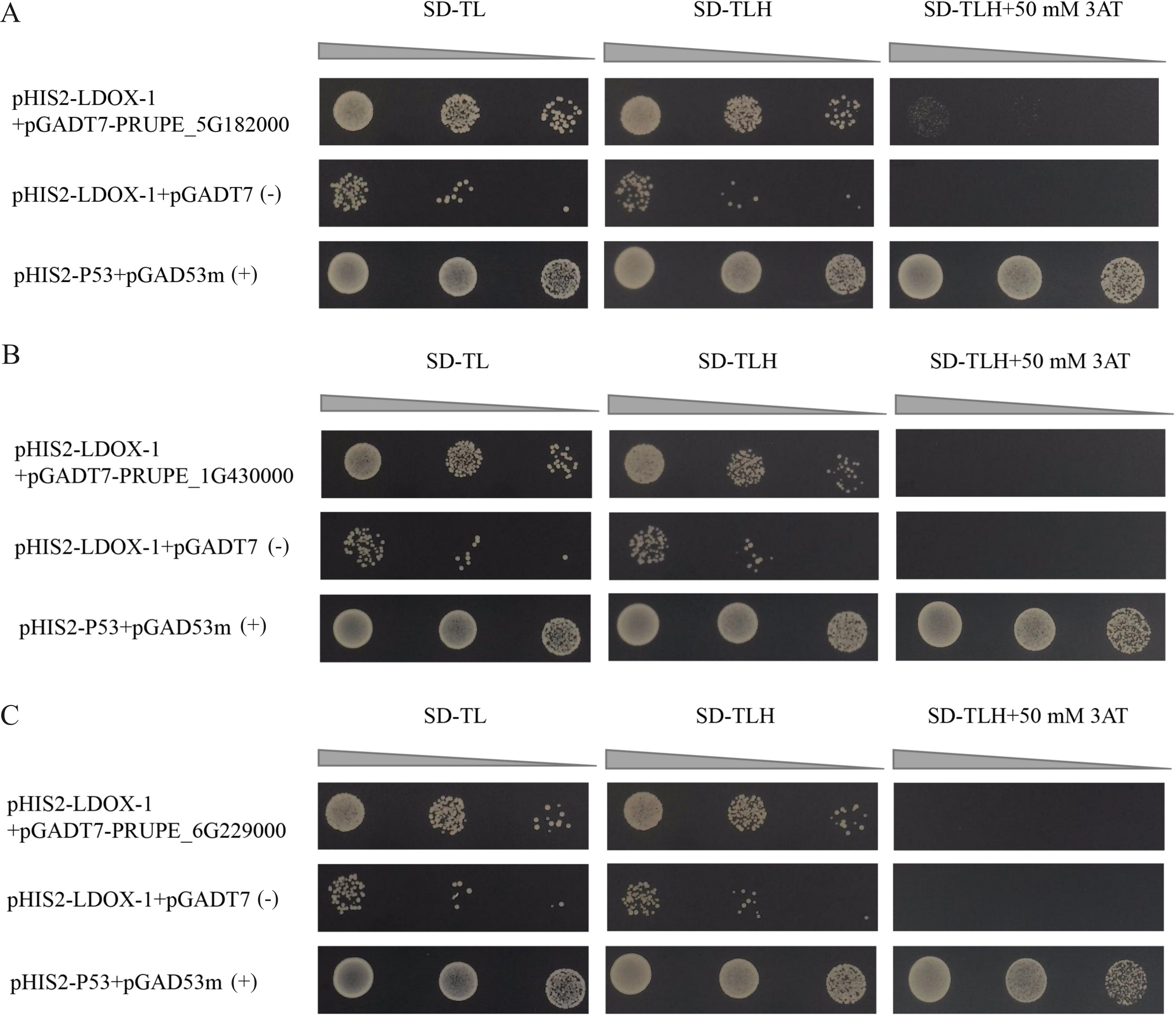{kind=link}
Y1H-AOS predicted analysis
Y1H-AOS stands for analysis of confidence in nucleic acid-protein interactions. Y1H-AOS models the structure of the LDOX promoter and the MYB1R1 protein through homology modeling to predict their interaction. The predominant elements of the 3D structure of the LDOX promoter exhibit a double helical structure (Fig. 4A), while the MYB1R1 protein includes α-helix and irregular coil (Fig. 4B). The pLDDT value obtained from AlphaFold2 was 60.63, and the confidence score from HDOCK was 0.9763, indicating the overall high reliability and stability of this nucleic acid protein composite structure. Although the pLDDT values from AlphaFold2 suggest some local uncertainty or flexibility in certain regions, the high confidence values from HDOCK further support the overall stability and quality of this composite structure. The analysis identified 24 binding sites (Fig. 4C; Table 2) where MYB1R1 protein amino acid residues interact with the LDOX promoter to form hydrogen bonds, with no disulfide, covalent, or salt bridges detected at these binding sites (Fig. 4D).
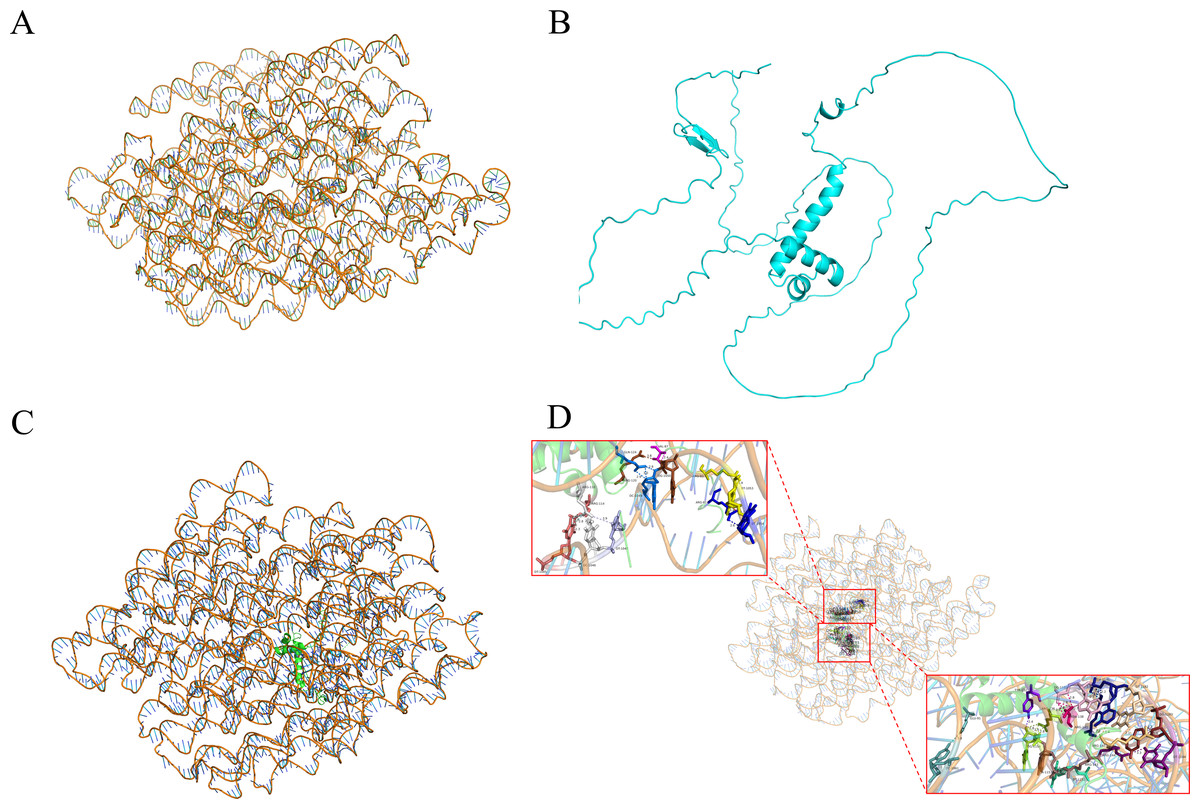
Figure 4: MYB1R1 predictive analysis.
(A) 3D structure of LDOX promoter. (B) 3D structure of MYB1R1 protein. (C) 3D structure of two sequences docking. (D) Analysis of the binding-site.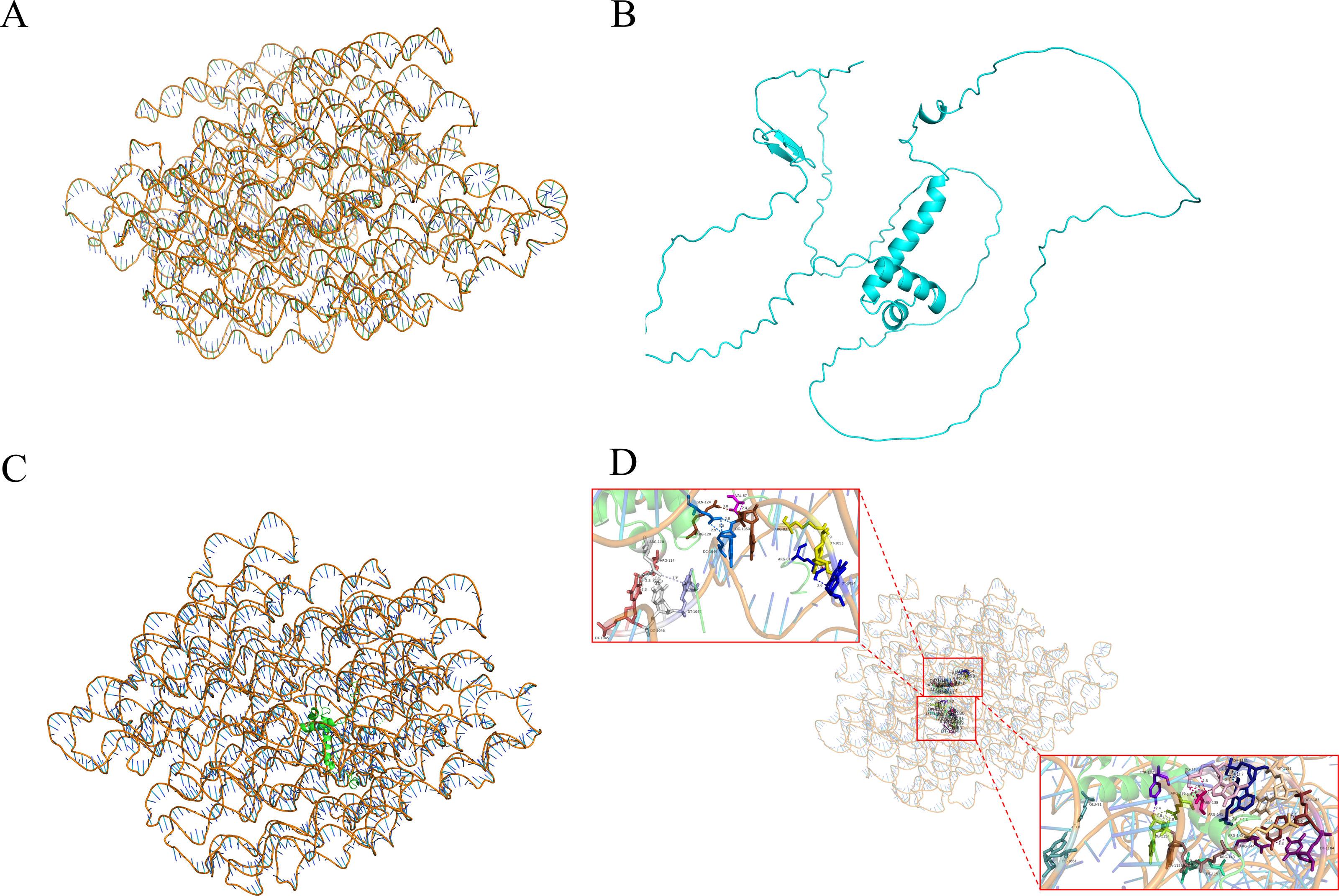{kind=link}
| Number | MYB1R1 | Dist [Å] | LDOX promoter |
|---|---|---|---|
| 1 | A:ARG 81[NH1] | 3.55 | B:DT1053[O4] |
| 2 | A:ARG 81[NH2] | 3.67 | B:DT1053[O2] |
| 3 | A:ARG 81[NH2] | 2.57 | B:DT1054[O2] |
| 4 | A:ARG 83[NH2] | 3.61 | B:DT1053[O4] |
| 5 | A:VAL 87[N] | 2.37 | B:DG1050[OP1] |
| 6 | A:ARG 110[NH1] | 2.54 | B:DT1045[O4] |
| 7 | A:ARG 110[NH2] | 3.86 | B:DT1047[O4] |
| 8 | A:ARG 110[NH2] | 2.32 | B:DC1046[N3] |
| 9 | A:ARG 114[NH1] | 2.33 | B:DT1045[O2] |
| 10 | A:ARG 120[NH1] | 3.75 | B:DG1050[OP1] |
| 11 | A:GLN 124[NE2] | 2.80 | B:DC1049[O3′] |
| 12 | A:TYR 132[OH] | 2.37 | B:DG1154[O3′] |
| 13 | A:ARG 136[NH1] | 2.83 | B:DG1154[O3′] |
| 14 | A:ASN 138[ND2] | 2.84 | B:DG1180[O5′] |
| 15 | A:ARG 141[NH1] | 3.87 | B:DG1180[O3′] |
| 16 | A:ARG 143[NH2] | 2.75 | B:DT1152[O3′] |
| 17 | A:ARG 143[NH2] | 2.01 | B:DA1153[OP1] |
| 18 | A:ARG 144[NH1] | 3.32 | B:DT1184[O4] |
| 19 | A:ARG 145[NH1] | 3.06 | B:DT1182[O2] |
| 20 | A:ARG 145[NH2] | 3.80 | B:DG1183[O3′] |
| 21 | A:ARG 145[O] | 2.78 | B:DA1181[N1] |
| 22 | A:ARG 141[O] | 3.04 | B:DA1181[N6] |
| 23 | A:ARG 145[O] | 2.84 | B:DA1181[N6] |
| 24 | A:GLU 91[OE1] | 3.81 | B:DC1861[N4] |
Analysis of MYB1R1 protein properties
In this study, we examined the MYB1R1 gene, which encodes a protein consisting of 298 amino acids (Fig. 5A). The MYB1R1 protein has a molecular weight of 32.49 kDa, with a molecular formula of C1392H2219N425O450S12, corresponding to a total atomic number of 4498. Theoretical isoelectric point value of 7.20 inferred that it is a basic protein. The minimum hydrophilic value (−4.022) was predicted at amino acid sites 20 and 21, while the maximum hydrophilic value (2.300) was found at sites 139, 140, and 141. The average total hydrophilic coefficient was calculated to be −0.599, indicating that this protein is hydrophilic (Fig. 5B). Additionally, prediction results for the protein signal peptide indicate that the MYB1R1 protein does not possess a signal peptide (Fig. 5C). The predicted number of transmembrane structures for MYB1R1 is 0, confirming that it lacks any transmembrane structure (Fig. 5D). Predict subcellular localization of MYB1R1 protein in the nucleus (Fig. 5E). Phosphorylation is predicted to occur within the MYB1R1 peptide chain, with 35 amino acid sites exhibiting scores above 0.5. This includes 21 serine, 12 threonine, and 2 thyronine phosphorylation sites (Fig. 5F).
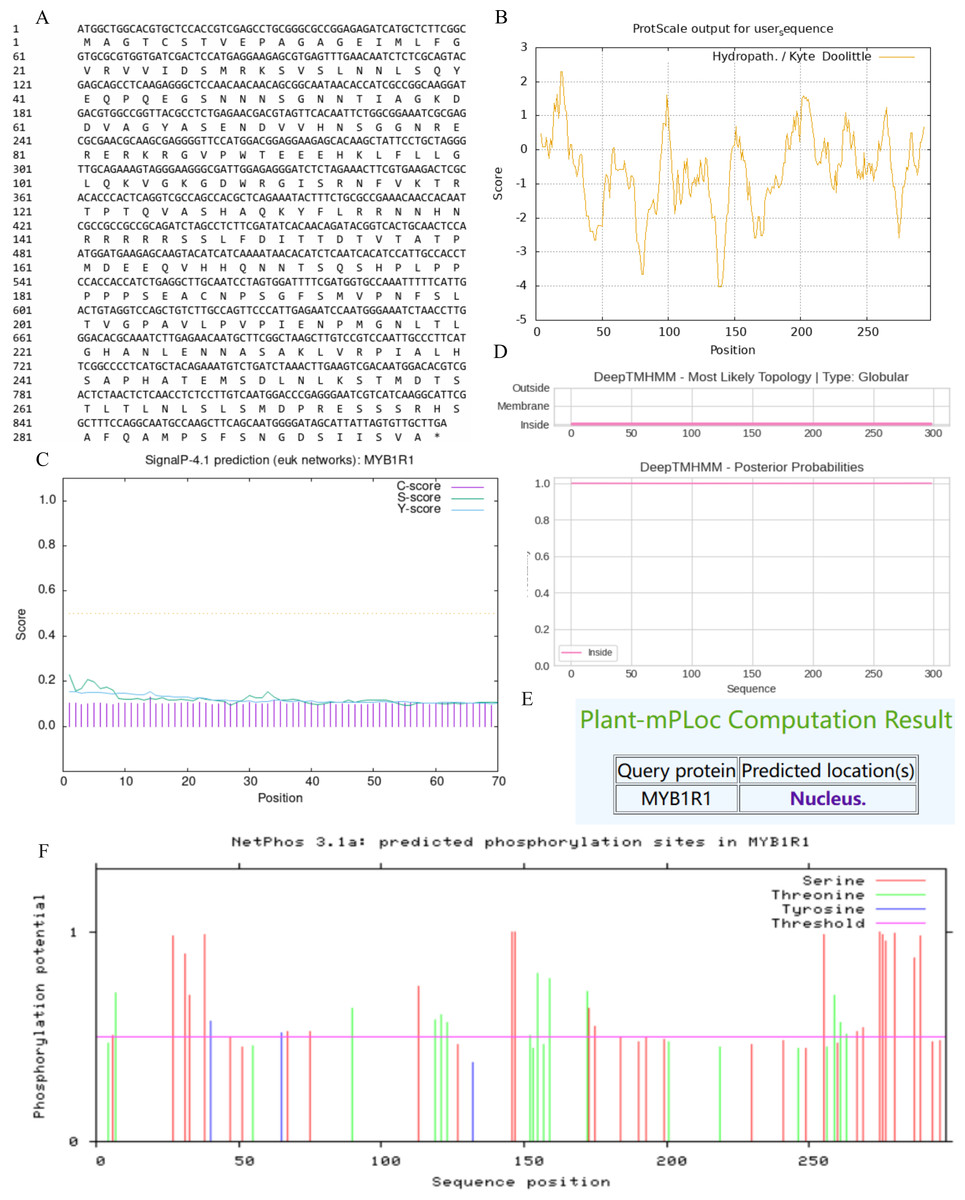
Figure 5: MYB1R1 protein physicochemical properties analysis.
(A) Amino acid sequence. (B) Prediction of hydrophilic/hydrophobicity. (C) Prediction of signal peptide. (D) Prediction of transmembrane structure. (E) Prediction of the subcellular localization. (F) Prediction of phosphorylation site.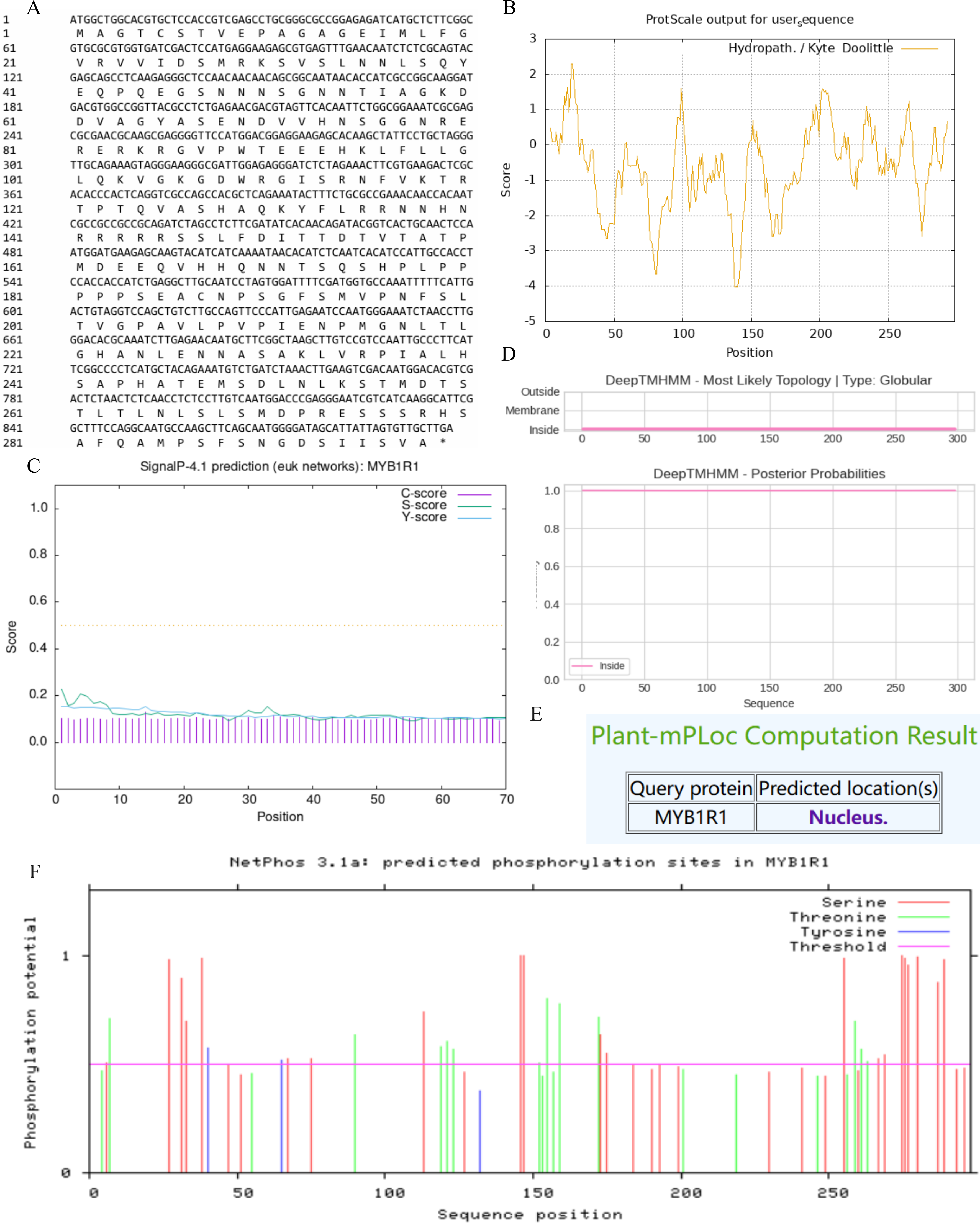{kind=link}
MYB1R1 protein structure prediction analysis
The MYB1R1 protein secondary structure prediction results indicated that its composition primarily consists of α-helix (13.76%), β-helix (2.35%), random coil (68.12%), and extended chain (15.77%) (Fig. 6A). The predicated tertiary structure exhibited a GMQE value of 0.56 and a similarity value of 97.32% (Fig. 6B). Notably, both the secondary and tertiary structure prediction results were consistent, with random coil identified as the predominant structural component of the MYB1R1 protein.
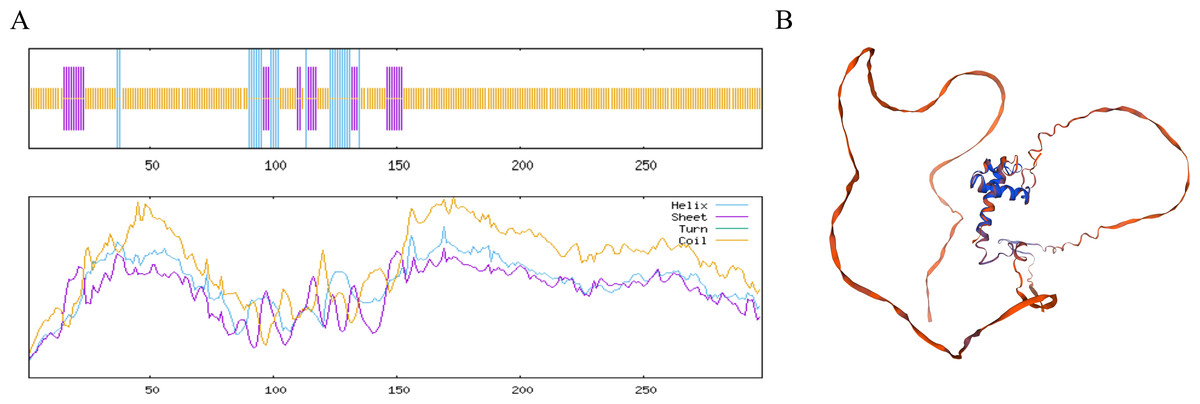
Figure 6: Structure prediction of the MYB1R1 protein.
(A) Secondary structure prediction of the MYB1R1 protein. (B) Tertiary structure prediction of the MYB1R1 protein.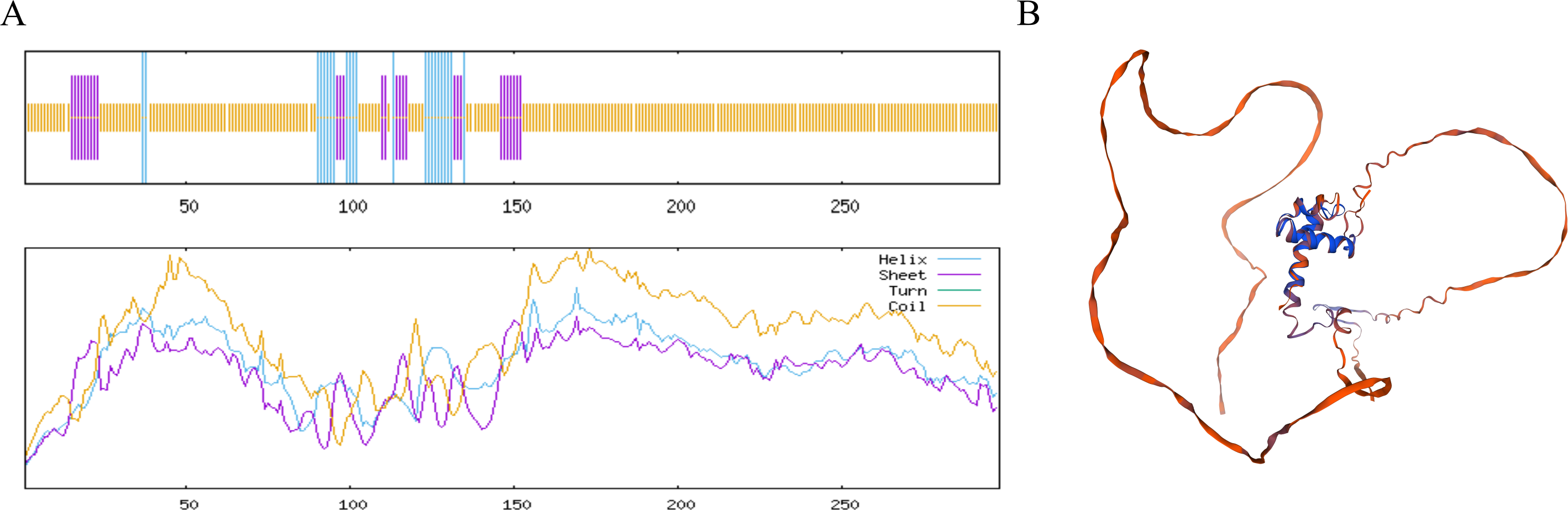{kind=link}
Analysis of the interaction proteins of MYB1R1
Using STRING to predict the interacting proteins of MYB1R1, the results showed that MYB1R1 may interact with ten proteins (Fig. 7), including bHLH domain-containing proteins (PRUPE_6G211900, PRUPE_6G212000, and PRUPE_1G434000), NAC TFs (NAC1 and NAC2), homeobox domain-containing protein (PRUPE_6G080100), cell division control protein (PRUPE_6G059500), ADF-H domain-containing protein (PRUPE_6G082200) and uncharacterized proteins (PRUPE_1G430000 and PRUPE_1G011300). This suggests that MYB1R1 may interact with additional proteins to perform various functions.
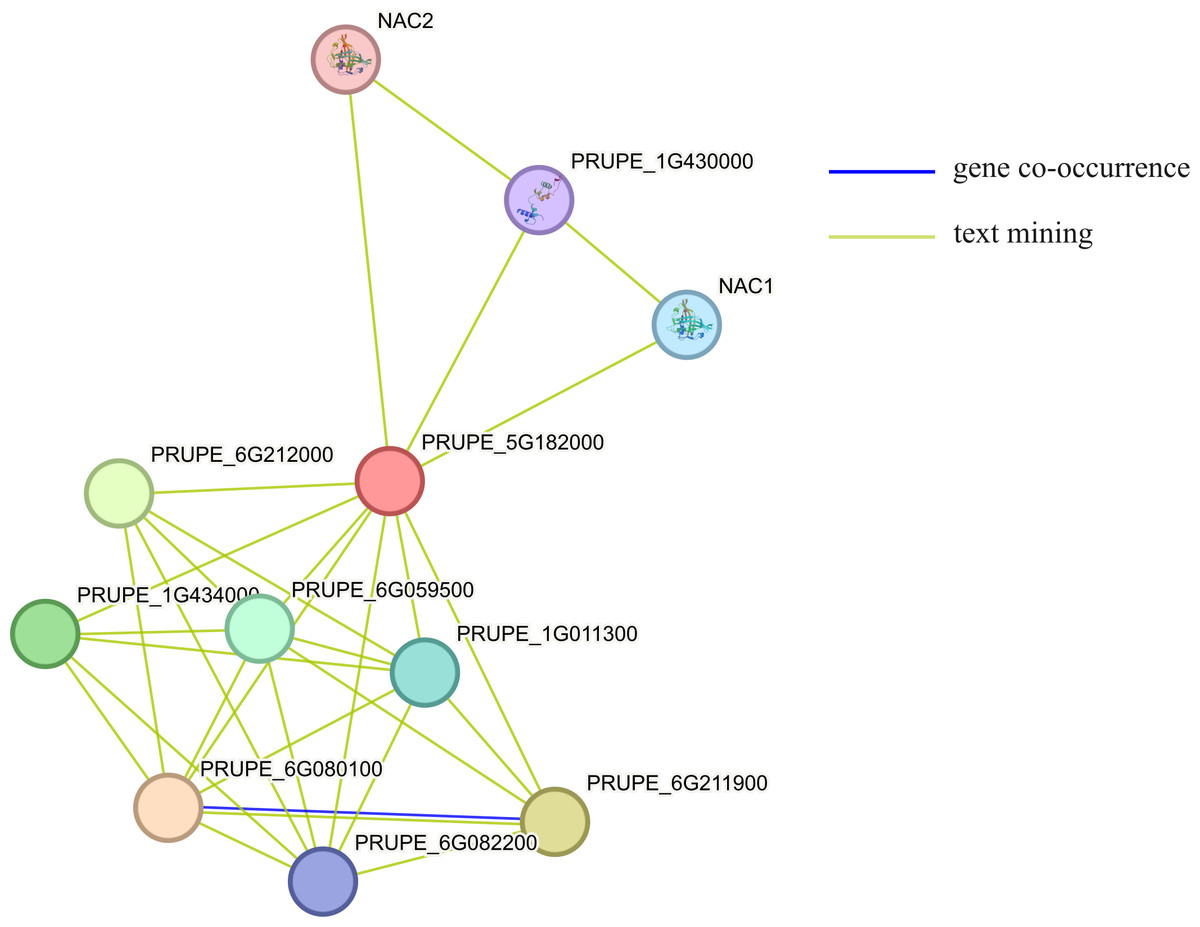
Figure 7: The protein–protein interaction network analysis of MYB1R1.
The circles, depicted in various colors, represent the proteins that interact with MYB1R1. Different line colors denote various active interaction sources: blue lines indicate gene co-occurrence, while yellow lines represent text mining.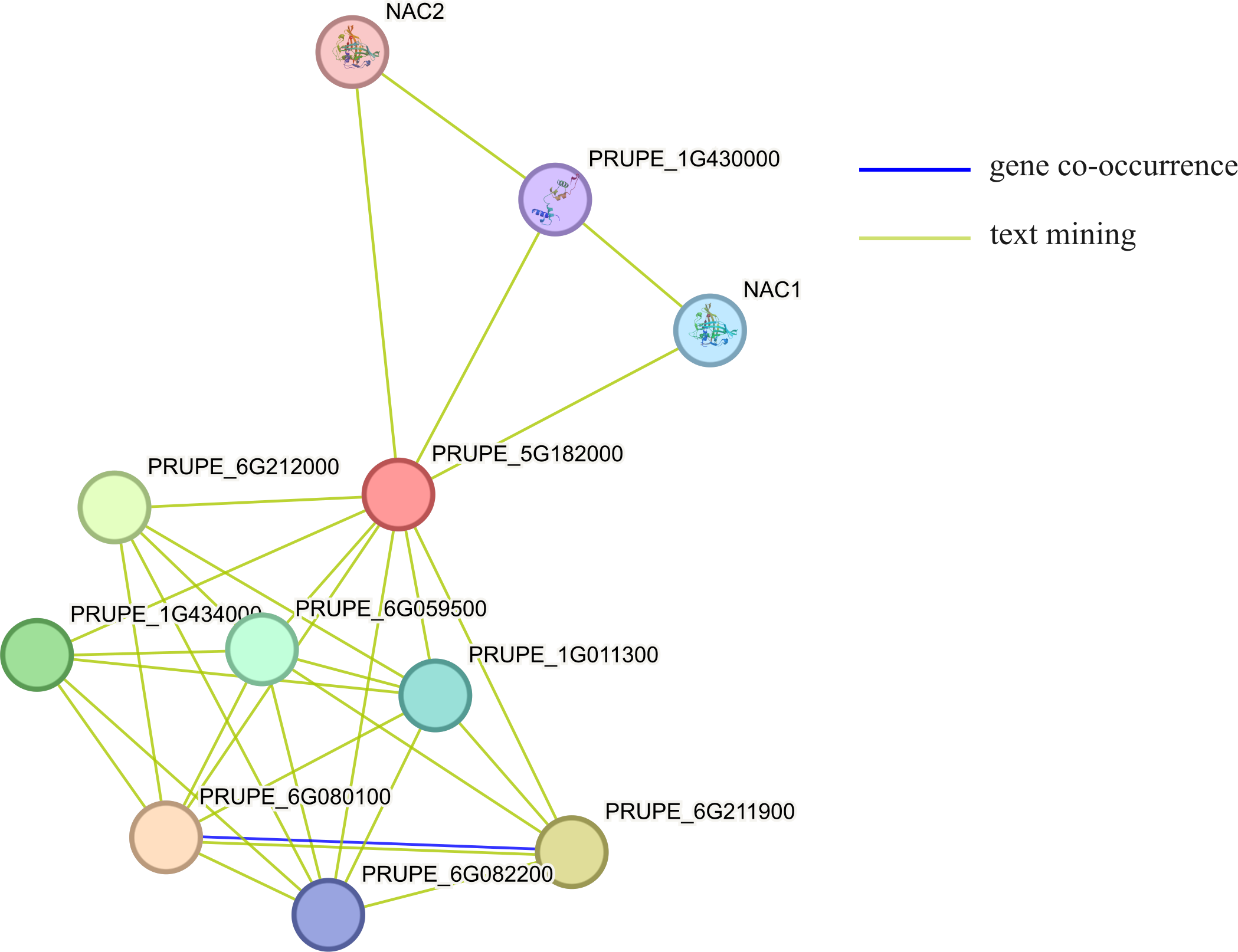{kind=link}
Sequence alignment and phylogenetic analysis of MYB1R1 gene
After aligning the amino acid sequence encoded by the MYB1R1 gene from NCBI, we performed a homology sequence alignment analysis using DNAMAN V6 software (Fig. 8A). This analysis revealed a high degree of homology (>70%) with the amino acid sequences of P. avium, P. speciosa, Pyrus communis, Pyrus× bretschneideri, Rosa chinensis, Rosa rugosa, Quercus robur, and Morella rubra. The evolutionary tree indicates that the protein encoded by the MYB1R1 gene has the closest phylogenetic relationship with P. avium and P. speciosa (Fig. 8B). Additionally, prediction and analysis of conserved domains using the SMART online tool demonstrated that the MYB1R1 gene-encoded protein shares the SANT-MYB domain with the aligned sequences from other species.
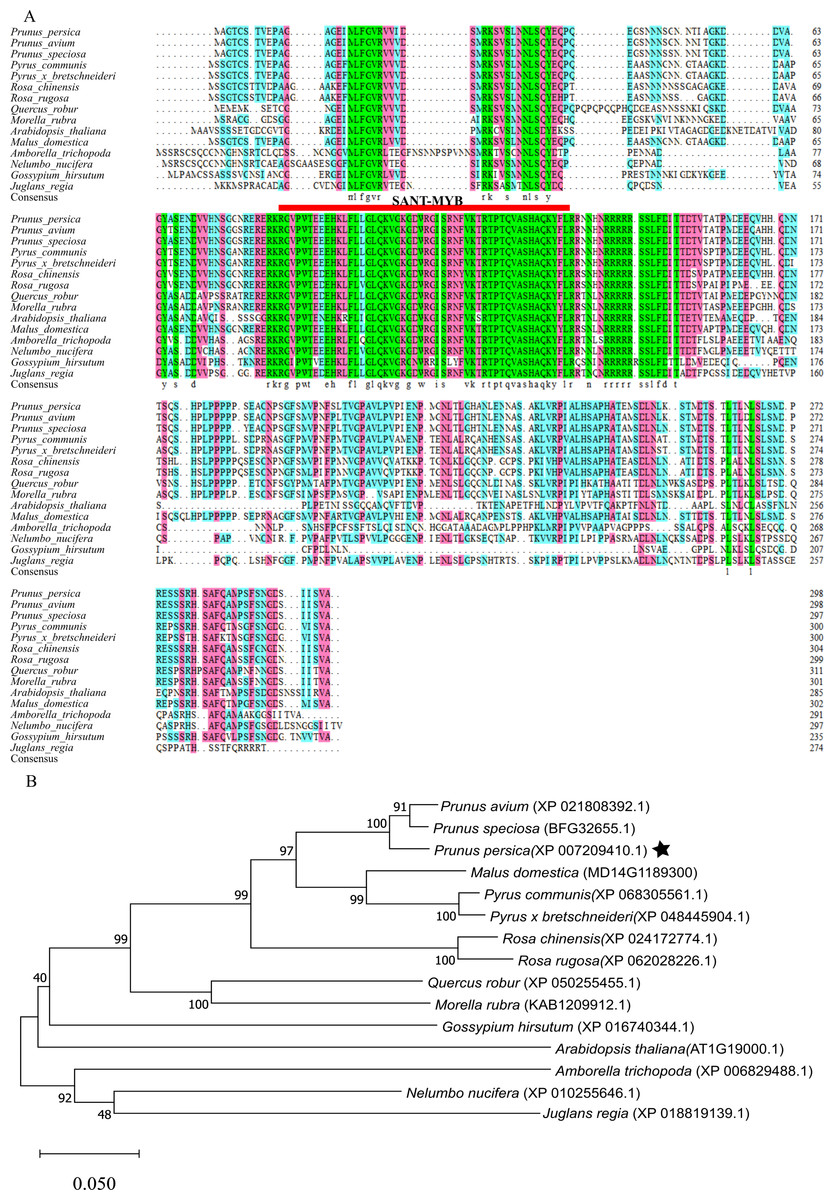
Figure 8: Multiple sequence alignment and phylogenetic analysis of MYB1R1 gene.
(A) Multiple sequence alignment of amino acid sequences among MYB1R1 and other plants. (B) Phylogenetic tree analysis of MYB1R1 protein.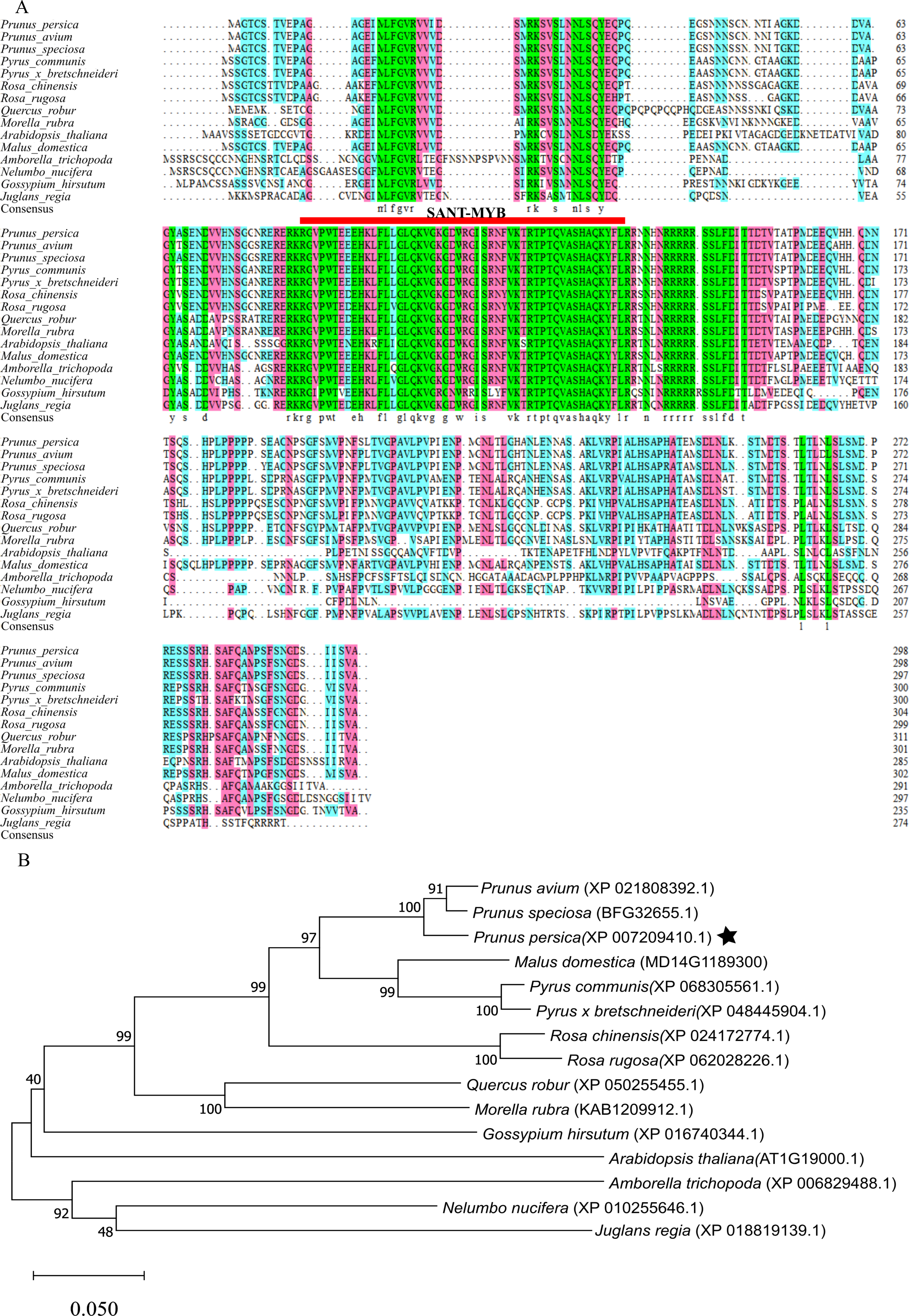{kind=link}
Prediction of CpG island and cis-acting elements in MYB1R1 promoter
The results of CpG island prediction for the promoter region of MYB1R1 indicate the presence of one CpG island within MYB1R1, which is 91 bp in length and located at the position of 471–561 bp (Fig. 9A). The results revealed that the MYB1R1 promoter contains a total of 32 distinct types of cis-acting elements. Apart from the conserved promoter elements CAAT-box and TATA-box, it includes light-responsive regulatory elements such as Box4, I-box, GATA-motif, ATCT-motif, G-box, TCT-motif, AE-box, and GT1-motif. Additionally, there are hormone-responsive elements like ABRE, TGAGG-motif, CGTCA-motif, AuxRR-core, and TCA-element. The promoter also harbors MYB TF binding elements such as MBS, as well as growth and development regulatory elements including Circadian, GCN4_motif, HD-Zip, and O2-site. Furthermore, MBS and LTR are cis-elements involved in stress responsiveness (Fig. 9B). These cis-acting elements are likely to influence the ultimate gene expression.
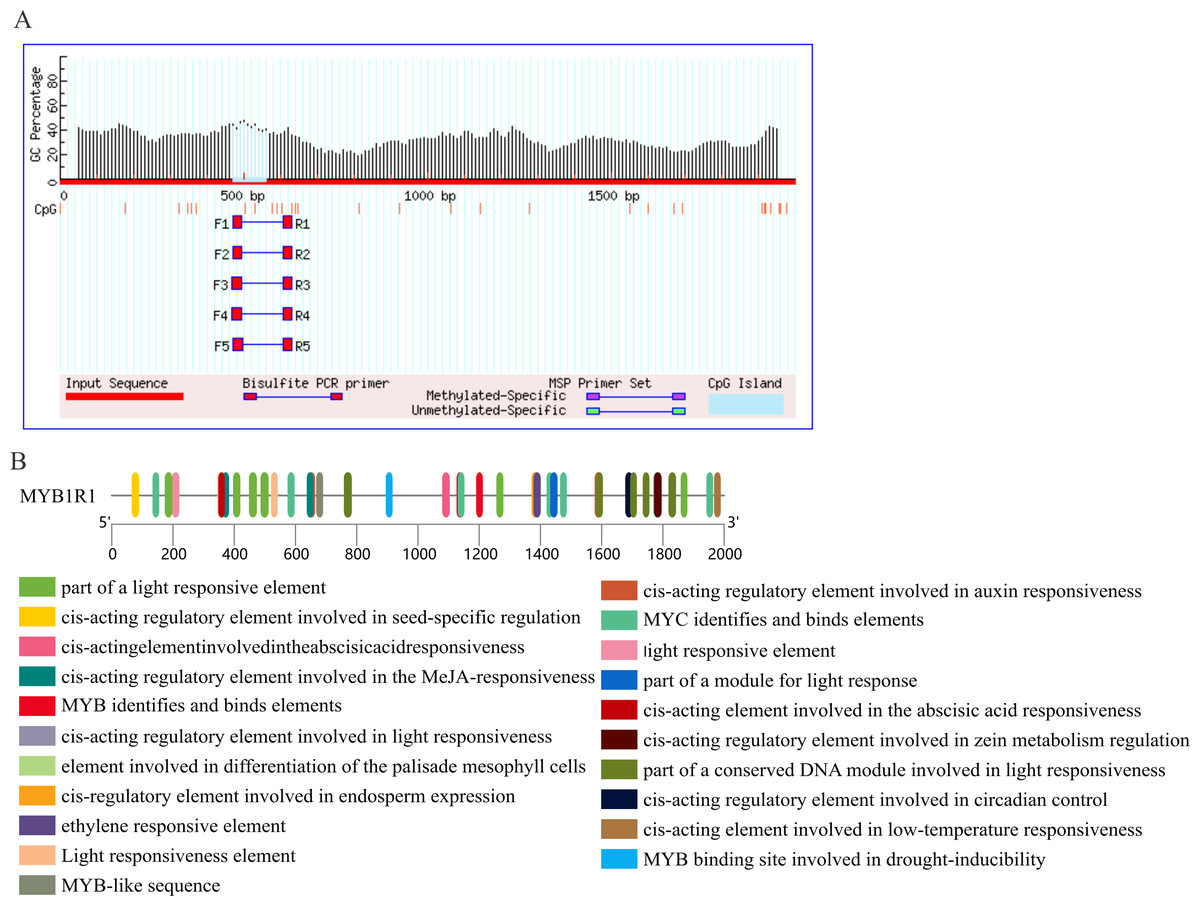
Figure 9: Prediction of CpG island and cis-acting elements in MYB1R1 promoter.
(A) Prediction of CpG island in MYB1R1 promoter. (B) Prediction of cis-acting elements in MYB1R1 promoter.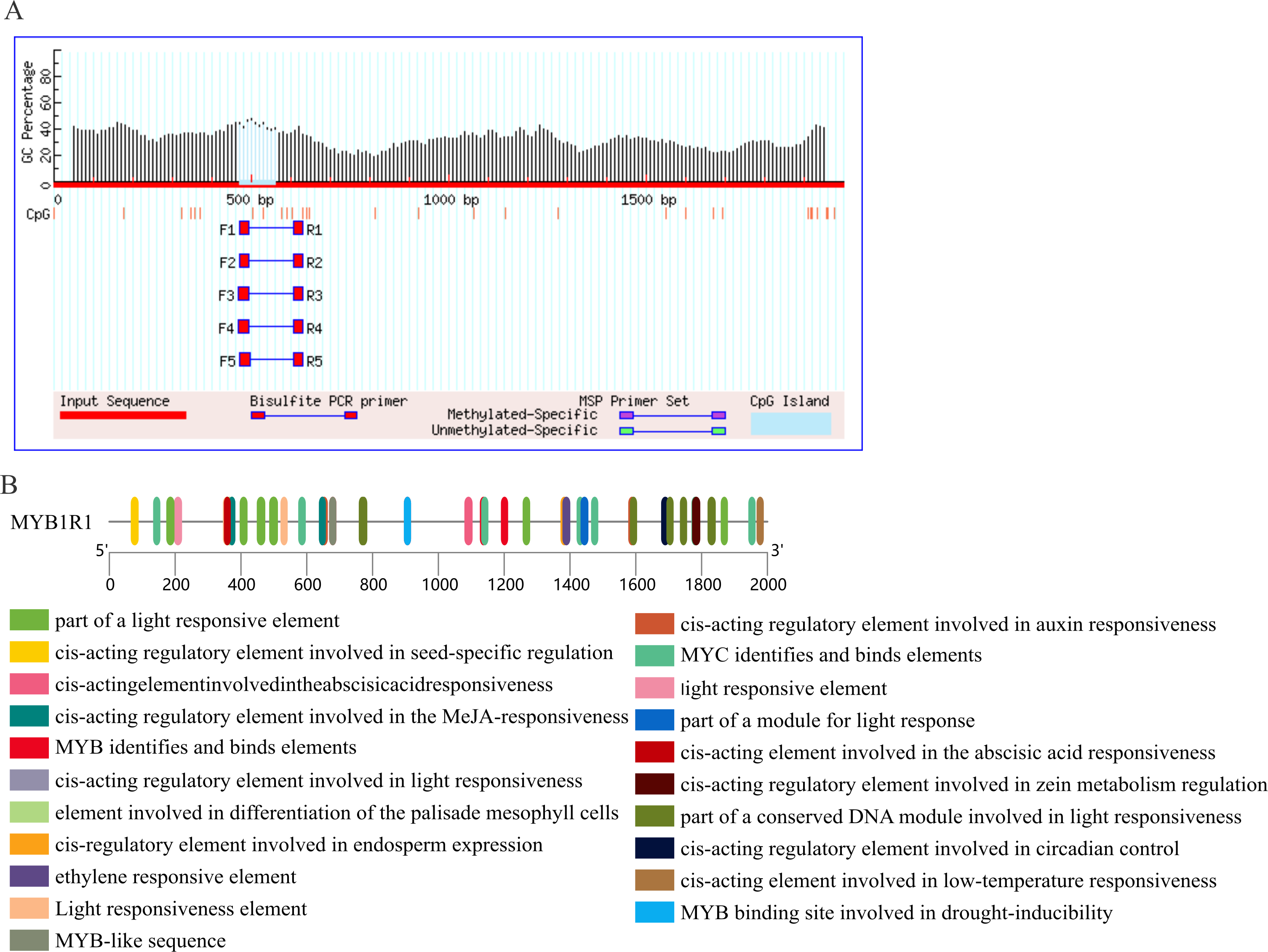{kind=link}
Gene expression analysis of MYB1R1 and LDOX
The gene expression of MYB1R1 and LDOX in various tissues of ‘Sahong Tao’ was analyzed using qRT-PCR (Fig. 10). The results showed that the expression level of MYB1R1 in variegated petals is higher than that in red petals, whereas the expression level of LDOX in variegated petals is lower than that in red petals. It is speculated that MYB1R1 interacts with LDOX to negatively regulate the synthesis of anthocyanins. Furthermore, nearly no gene expression was observed in sepals-R and sepals-G.
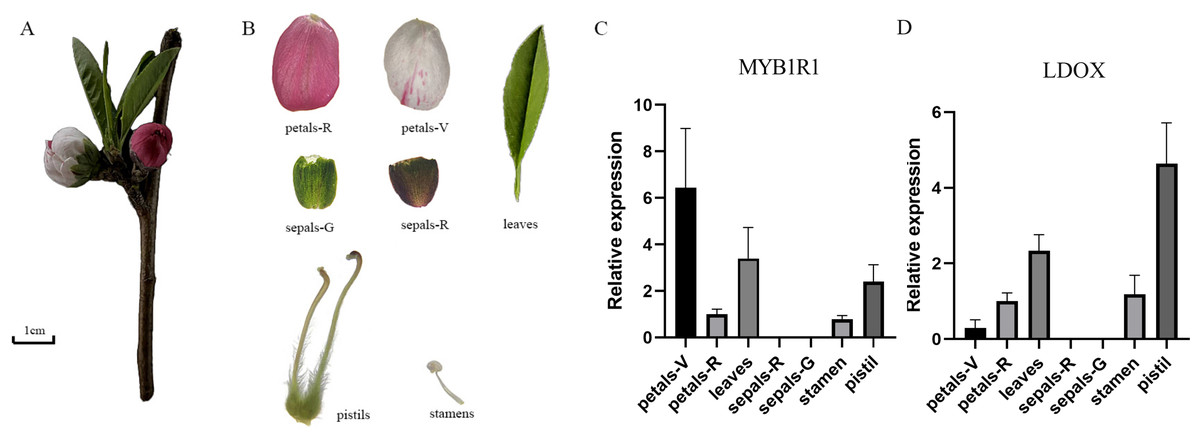
Figure 10: Plant materials and the expression analyses of two genes (MYB1R1 and LDOX).
(A) ‘Sahong Tao’ plant materials used in this study. (B) ‘Sahong Tao’ pictures of the different part. (C) The relative expression level of MYB1R1 in different parts of ‘Sahong Tao’. (D) The relative expression level of LDOX in different parts of ‘Sahong Tao’.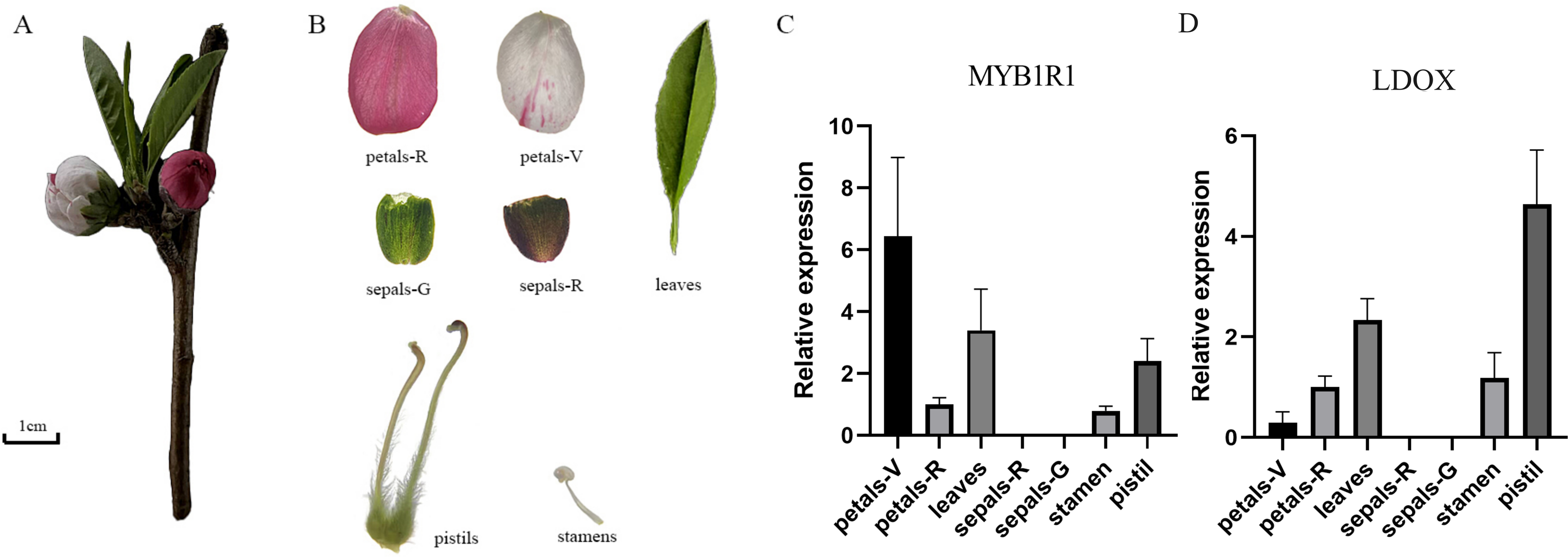{kind=link}
Discussion
Structural genes LDOX control the synthesis of anthocyanins
Anthocyanins are a group of naturally water-soluble pigments responsible for color changes in vegetative tissues and reproductive organs (Sharma et al., 2024). The color variation in the petals of ‘Sahong Tao’ is influenced by anthocyanin synthesis, which is regulated by the expression of structural genes encoding key enzymes such as PAL, CHS, CHI, F3H, F3′H, F3′5′H, DFR, LDOX/ANS, and UFGT, as well as TFs (Ferrer et al., 2008; Zohar et al., 2015). LDOX catalyzes the synthesis of unmodified colored anthocyanins from leucoanthocyanidins. During the veraison to ripening stages of grape berries, LDOX exhibits a positive correlation between gene expression and anthocyanin content (Zhao et al., 2016). In Magnolia, the expression level of LDOX genes in red petals is significantly higher than that in white petals (Shi et al., 2015). This finding aligns with observations that both the protein abundance and gene expression of LDOX in the red petals of ‘Sahong Tao’ are significantly greater than those in white petals (Zhou et al., 2015a). Variations in LDOX gene expression may affect petal color. However, the regulatory network involving LDOX is poorly understood in peach. In this study, NGS and Y1H were used to screen for TFs interacting with the LDOX promoter involved in anthocyanin synthesis in the petals of ‘Sahong Tao’. This lays a foundation for further revealing the molecular mechanisms of anthocyanin synthesis and accumulation in peaches.
The role of MYB1R1 in anthocyanin synthesis
TFs play a crucial role in regulating anthocyanin metabolism by interacting with structural genes in plants (Dubos et al., 2010). Previously reported MYB TFs, including AtPAP1 (AtMYB75), AtPAP2 (AtMYB90), AtPAP3 (AtMYB113), and AtPAP4 (AtMYB114), as well as the bHLH TT8 and GL3 (Gao et al., 2017; Salez et al., 2022), and the WRKY (Shi et al., 2022; Wang et al., 2023), have been shown to be involved in anthocyanin biosynthesis. Among these, MYB proteins represent one of the largest families of TFs in plants (Hu et al., 2020). The expression of MYB-6 and LDOX1 increases under cold stress, leading to the accumulation of major anthocyanins in purple-black carrots (Dar et al., 2022). In Arabidopsis, the expression levels of LDOX in MYB3 significantly increase under high salinity conditions, resulting in a substantial rise in anthocyanin accumulation (Kim et al., 2022). MYB forms the MYB-BHLH-TTG1 complex, which directly regulates the expression of LDOX, thereby influencing anthocyanin accumulation in Arabidopsis seedlings (Appelhagen et al., 2011). PavMYB10.1 is involved in the anthocyanin biosynthesis pathway and determines the skin color of sweet cherries (Jin et al., 2016). In peach, the MYB TF Peace is highly expressed in pink petals but shows lower expression levels in variegated petals (Uematsu et al., 2014). MYB10 and MYBPA1 regulate anthocyanin biosynthesis in the pericarp and proanthocyanidin biosynthesis in the flesh, respectively (Ravaglia et al., 2013). Previous studies have identified multiple MYB-related cis-acting elements (including MBS, MRE, MYB, and MYB recognition sites) in the LDOX promoter of the ‘Sahong Tao’ (Wu et al., 2024). In this study, 1,190 proteins interacting with the LDOX promoter were screened out through Y1H and NGS, and three MYB transcription factors were identified. Therefore, MYB TFs and LDOX may be key factors influencing anthocyanin synthesis.
The Y1H analysis is employed to investigate the interactions between proteins and DNA. In eggplant, Y1H has been utilized to substantiate the binding of SmMYB75 to the SmCHS promoter facilitating anthocyanin biosynthesis (Shi et al., 2021). Furthermore, BoMYBL2b interacts with the MRE site on ProBoDFR1, influencing anthocyanin synthesis in kale (Liu et al., 2024b). The binding of DcMYB11c to the DcUCGXT1 and DcSAT1 promoters is involved in anthocyanin synthesis in purple carrot (Duan et al., 2023). Y1H screening and qRT-PCR analysis by ABA signaling demonstrated direct binding of MYC2 and MYB1R1 to the PbFAD3a promoter in pears (Wang et al., 2022). BLAST comparison of homologous gene in Arabidopsis revealed that MYB1R1 is most similar to AT1G19000.2, which has been confirmed to be involved in proanthocyanidin accumulation (Hong et al., 2017). It is hypothesized that MYB1R1 may also have a similar function in peaches, and the current results are consistent with previous reports. Therefore, in this study, Y1H was used to verify the interaction between MYB1R1 and the LDOX promoter. Quantitative reverse transcription polymerase chain reaction (qRT-PCR) experiments demonstrated that the expression levels of MYB1R1 and LDOX are inversely correlated in variegated and red flowers of ‘Sahong Tao’, suggesting that they play a key role in the formation of petal variegation in ‘Sahong Tao’. Additionally, MYB1R1 regulates anthocyanin synthesis as a repressive transcription factor.
Bioinformatics analysis of MYB1R1 revealed that the N-terminus of it features a typical SANT-MYB domain. A family of genes containing the SANT/MYB domain has been identified in tomato, where they play a role in regulating plant growth and development (Barg et al., 2005). The protein encoded by the smh1 gene in maize contains a SANT/myb-like domain at its N-terminus and binds to repetitive sequences at the ends of DNA. This leads to the hypothesis that MYB1R1 may similarly bind to repetitive sequences at DNA termini (Marian & Bass, 2005). Results from multiple sequence alignment and phylogenetic analysis indicate that the amino acids encoded by MYB1R1 share close homology with those encoded by genes in plants such as P. avium, and Prunus speciosa. The MYB in its homologous species has been confirmed to be involved in anthocyanin biosynthesis. In addition to containing conserved promoter cis-acting elements such as TATA-box and CAAT-box, the promoter of MYB1R1 also includes cis-acting elements related to light response, hormones, growth and development, and stress response.
Future directions
DNA methylation is a crucial mechanism of genomic modification (Bartels et al., 2018), which can regulate gene expression by interactions with TFs or altering chromosomal structure. Promoter sequences recognized by TFs are prominent in the structure and contain CpG sites (Liu et al., 2013). When cytosine at these CpG sites undergoes methylation, TFs sensitive to DNA methylation are no longer able to bind to the corresponding promoter sequence, resulting in the inability to activate and express the associated gene (Curradi et al., 2002). In apples and pears, DNA methylation influences anthocyanin accumulation in the peel by regulating MYB expression. The methylation level of CHS in orchids determines the accumulation of anthocyanins in flower organs (Liu et al., 2012). In this study, MYB1R1 is shown to bind to the LDOX promoter to regulate anthocyanin synthesis, leading to petal variegation. In the whole genome methylation sequencing of ‘Sahong Tao’, the methylation level of the LDOX promoter was found to be higher in variegated samples compared to red samples. The methylation sites encompass several regulatory elements related to MYB (Wu et al., 2020). However, the difference of LDOX promoter methylation level maybe cause the differential expression of LDOX in ‘Sahong Tao’ petals, which ultimately affects anthocyanin synthesis. The effect of LDOX promoter methylation on the regulation of MYB1R1 is still unclear. Thus, the role of LDOX promoter DNA methylation in this process needs to be further investigated through techniques such as Chromatin Immunoprecipitation (ChIP) and Bisulfite Sequencing.
Conclusions
In this study, the Y1H and NGS results indicate that 1,190 proteins interact with the LDOX promoter. Among these, 20 TFs were identified. Potential LDOX promoter binding TFs were predicted, confirmed that seven TFs matched the Y1H results. We have verified the interaction between MYB1R1 and the LDOX promoter and confirmed 24 interaction binding sites by Y1H-AOS. qRT-PCR experiments showed that the expression level of MYB1R1 in variegated petals is higher than that in red petals and the expression level of LDOX in variegated petals is lower than that in red petals. The results showed that MYB1R1 interacted with LDOX promoter and negatively regulated the synthesis of anthocyanins. Our findings establish a theoretical foundation for further exploration of the regulatory role of MYB TFs in the flower color of ‘Sahong Tao’ and the biological functions of MYB1R1 gene.
Supplemental Information
NGS results
Gene ID: The gene identifier for the analyzed species; CDS: The nucleotide sequence of the gene in the analyzed species; Protein: The encoded protein sequence of the gene in the analyzed species; Read count: The number of high-throughput sequencing reads supporting the gene in the analyzed species, generally, a larger number indicates a higher likelihood of the gene being an interacting protein; A. thaliana homolog: The homolog gene ID in the model plant Arabidopsis thaliana, identified through BLAST comparison; e-value: The E-value of the BLAST comparison between this gene and its Arabidopsis thaliana homolog, with a smaller value indicating a higher similarity; A. thaliana gene symbol: The abbreviation or symbol for the Arabidopsis thaliana homolog gene; Functional annotation: The functional annotation of the gene in the analyzed species; TF/Kinase: Whether the gene in the analyzed species is a TF or a kinase.