In silico identification, characterization expression profile of WUSCHEL-Related Homeobox (WOX) gene family in two species of kiwifruit
- Published
- Accepted
- Received
- Academic Editor
- Wei Ma
- Subject Areas
- Bioinformatics, Genetics, Genomics, Plant Science
- Keywords
- Actinidia chinensis, A. eriantha, Whole-genome duplication, Expression profiles, Conserved motif
- Copyright
- © 2021 Feng et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. In silico identification, characterization expression profile of WUSCHEL-Related Homeobox (WOX) gene family in two species of kiwifruit. PeerJ 9:e12348 https://doi.org/10.7717/peerj.12348
Abstract
The WUSCHEL (WUS)-related homeobox (WOX) gene family is a class of plant-specific transcriptional factors and plays a crucial role in forming the shoot apical meristem and embryonic development, stem cell maintenance, and various other developmental processes. However, systematic identification and characterization of the kiwifruit WOX gene family have not been studied. This study identified 17 and 10 WOX genes in A. chinensis (Ac) and A. eriantha (Ae) genomes, respectively. Phylogenetic analysis classified kiwifruit WOX genes from two species into three clades. Analysis of phylogenetics, synteny patterns, and selection pressure inferred that WOX gene families in Ac and Ae had undergone different evolutionary patterns after whole-genome duplication (WGD) events, causing differences in WOX gene number and distribution. Ten conserved motifs were identified in the kiwifruit WOX genes, and motif architectures of WOXs belonging to different clades highly diverged. The cis-element analysis and expression profiles investigation indicated the functional differentiation of WOX genes and identified the potential WOXs in response to stresses. Our results provided insight into general characters, evolutionary patterns, and functional diversity of kiwifruit WOXs.
Introduction
Homeobox (HB) proteins were primarily identified in Drosophila, and they belong to a large transcriptional factor (TF) family harboring a short stretch of amino acids (60–66 residues) conserved DNA-binding domain (referred to as homeodomain) (Mukherjee, Brocchieri & Bürglin, 2009; Holland, 2013; Alvarez et al., 2018). HB proteins could be found in all eukaryotic species tested (Liu et al., 2014b). The homeodomain (HD)-containing transcriptional factor family has been identified in both monocots and dicots (Ariel et al., 2007; Liu et al., 2014b). The HB superfamily can be classified into six subfamilies, including homeodomain-leucine (HD-Zip), plant homeodomain (PHD)-finger, BELL, zinc finger-homeodomain (ZF-HD), WUSCHEL (WUS)-related homeobox (WOX), and KNOTTED1-like-homeobox (KNOX) (Ariel et al., 2007; Mukherjee, Brocchieri & Bürglin, 2009; Liu et al., 2014b). Previous researches have verified that members of the WOX subfamily participate in many plant biological processes, such as the formation and maintenance of the shoot apical meristem (SAM) (Meng et al., 2019). The WOX subfamily specifically binds to the target region by the homeodomain to activate or depress the expression of the target gene in plants (Mukherjee, Brocchieri & Bürglin, 2009; Jha, Ochatt & Kumar, 2020; Tvorogova et al., 2021). Genome-wide identification of the WOX subfamily has been performed in several plants, including Arabidopsis, rice, maize, walnut, physic nut, grapes, peach, pear, apricot, coffee, and poplar (Li et al., 2020; Shafique Khan et al., 2021).
The WOX gene family has often been classified into three clades which include the ancient clade, the intermediate clade, and the modern/WUS clade (Tvorogova et al., 2021). Among the 15 WOX genes in Arabidopsis, AtWOX10, AtWOX13, and AtWOX14 belong to the ancient clade (Tvorogova et al., 2021). The AtWOX10 gene is a presumptive pseudogene owing to its undetectable gene expression in Arabidopsis plants (Deveaux et al., 2008). While AtWOX13 and AtWOX14 play central roles in regulating the development of flowers, fruits, and conductive tissues (Deveaux et al., 2008; Costanzo, Trehin & Vandenbussche, 2014; Jha, Ochatt & Kumar, 2020). However, WOX genes in the ancient clade expressed differently in different species, indicating the species-specific function of ancient clade WOX genes (Alvarez et al., 2018). The AtWOX13 gene regulates fruit development, the number of lateral roots, and flower time (Deveaux et al., 2008; Romera-Branchat et al., 2013). The AtWOX14 usually affects plant growth and the formation of conductive tissues, and the deletion of AtWOX14 induces plant dwarfism (Denis et al., 2017). The WOX genes of the ancient clade in other species functioned similarly to that in Arabidopsis (Tvorogova et al., 2021). The intermediate clade contains four AtWOX genes (AtWOX8, AtWOX9, AtWOX11, and AtWOX12) (Deveaux et al., 2008; Tvorogova et al., 2021). The AtWOX8 and AtWOX9 co-regulate the development of the apical-basal polarity axis (Wu, Chory & Weigel, 2007; Lie, Kelsom & Wu, 2012). However, the orthologs of AtWOX9 in other species are verified participating in the inflorescence development (Tvorogova et al., 2021). The AtWOX11 and AtWOX12 have similar functions in regulating the callus formation and development of adventitious roots (Liu et al., 2014a). The modern/WUS clade is the largest clade which includes eight WOX genes (AtWUS and AtWOX1-7) in Arabidopsis (Deveaux et al., 2008). Besides the homeodomain, WOX genes in the modern/WUS clade also have other conserved domains, such as the WUS motif or EAR domain (ERF-associated amphiphilic repression) (Deveaux et al., 2008; Wu, Li & Kramer, 2019). WOX genes in the modern/WUS clade participate in the regulating developments of various types of meristems (Tvorogova et al., 2021). In summary, the WOX family plays a central role in maintaining different types of meristems, regulating the formation of plant organs, and controlling cell proliferation and differentiation (Tvorogova et al., 2021).
Kiwifruit becomes a popular fruit worldwide owing to its high vitamin C content and abundant minerals (Cheng et al., 2004; Stonehouse et al., 2013). Kiwifruit belongs to the Actinidia genus, including 54 species and 75 taxa (Tang et al., 2019). The whole genome of the A. chinensis (Ac) and A. eriantha (Ae) has been reported (Pilkington et al., 2018; Tang et al., 2019). These two species are different in many vital traits, especially flowering time (Pilkington et al., 2018; Tang et al., 2019). The WOX gene family is verified to affect plant flowering and development (Tvorogova et al., 2021), whereas systematic investigations and functional analyses of the WOX gene family have not been reported in kiwifruit.
In the present study, we comprehensively identified the WOX gene family from A. chinensis and A. eriantha genome. We firstly reported the gene structure, motif compositions, chromosomal distributions of the WOX gene family for these two kiwifruit species. Further, we analyzed and compared the phylogenetic relationships and evolution patterns of the WOX gene family for the two kiwifruit species. Cis-elements analysis and expression patterns in different tissues and under different stress conditions were performed. Our results provided critical information on the structure characters, evolution patterns, and potential function of the WOX genes in the two kiwifruit species.
Materials & methods
Gene identification and analysis
The whole-genome sequences and protein sequences of the two kiwifruit species (A. chinensis and A. eriantha) were obtained from the Kiwifruit Genome Database (http://kiwifruitgenome.org/). The WOX protein sequences of Arabidopsis and rice were collected from the PlantTFDB v4.0 (http://planttfdb.gao-lab.org/). The local BLAST tool was used to construct the protein database of the two kiwifruit species. The combined protein sequences of Arabidopsis and rice WOX protein were used to query the kiwifruit protein database by the BLASTp. The candidate WOX proteins in kiwifruit were identified by BLASTp search scores of ≥ 100 and an e-value of ≤ 1 × e−10. The Conserved Domain Database (CDD) (https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml) and the simple modular architecture research tool (SMART) (http://smart.embl.de/) were used to confirm the homeodomain of the candidate WOX protein, and the candidate WOX protein harboring the homeodomain were obtained and used for further analysis.
Analysis of kiwifruit WOX protein structure
The protein length, theoretical isoelectric point (pI), grand average of hydropathicity (GRAVY), and molecular weight (MW) of the WOX gene family in the two kiwifruit species were computed using the ProtParam on ExPASy server (http://web.expasy.org/protparam/). The subcellular localization of kiwifruit WOX proteins was predicted using the online web software CELLO (v2.5, http://cello.life.nctu.edu.tw/).
Gene structure, motif features, and cis-elements analysis
The genome sequences and coding sequences of the WOX genes of the two kiwifruit species were extracted. The gene structures of WOX genes were investigated using the Gene Structure Display Server (GSDS 2.0, http://gsds.cbi.pku.edu.cn/). The conserved motifs of WOX proteins were identified using MEME (http://meme-suite.org/tools/meme) with a maximum of 10 motifs (Bailey et al., 2009). To analyze cis-elements involved in regulating WOX genes, we extracted the 1,500-bp upstream promoter regions of each WOX gene, and predicted cis-elements with the PlantCARE database (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) (Lescot et al., 2002).
Phylogenetic analysis of WOX proteins
The multiple sequence alignments of WOX proteins from Arabidopsis thaliana, Oryza sativa, A. chinensis, and A. eriantha were performed using ClustalX with default parameters (Larkin et al., 2007). The phylogenetic tree was constructed by MEGA X software using the neighbor-joining (NJ) method with 1,000 bootstrap replicates (Kumar et al., 2018).
Gene duplication and synteny analysis
The genome location of WOX genes was extracted from the corresponding GFF file using an in-house Perl script, and the chromosomal distributions were rendered using MapGene2 Chrome (http://mg2c.iask.in/mg2c_v2.0/). The duplication patterns of kiwifruit WOXs were identified using the MCScanX software with default parameters (Wang et al., 2012). The synonymous (Ks) and nonsynonymous (Ka) mutation rates of the duplicated WOX gene pairs were calculated using TBtools software (Chen et al., 2020). The syntenic analysis of kiwifruit WOXs was conducted using the MCScanX software with default parameters to produce the collinearity blocks across the whole genome (Wang et al., 2012). The collinearity gene pairs of kiwifruit WOXs were visualized using TBtools (Chen et al., 2020).
Expression analysis of kiwifruit WOXs
To investigate the expression patterns of kiwifruit WOXs in different tissues, developmental stages, or under stress treatments, we collected nine published RNA-seq data including those from leaves, roots, stems, different fruit developmental stages, fruits treated with or without ethylene, leaves infected with pathogens (PRJNA514180, PRJNA187369, PRJNA277383, PRJNA328414, PRJNA436459, PRJDB5543, and PRJNA535344) from NCBI (https://www.ncbi.nlm.nih.gov/). We further re-analyzed these transcriptome data using genomes of the Actinidia chinensis ‘Red5’ cultivar and A. eriantha white cultivar as reference genome (Pilkington et al., 2018; Tang et al., 2019). The reads alignment was performed using the HISAT2 v2.0.1 (Kim et al., 2019), and the transcripts were assembled and quantified using the STRINGTIE v2.1.5 (Pertea et al., 2015).
Results
Genome-wide identification and classification of kiwifruit WOXs
To identify potential WOX family members in kiwifruits, WOX protein sequences from Arabidopsis were used as queries in BLASTp homology search against Ac and Ae genomes. Totaly, we identified 17 and 10 putative WOXs from Ac and Ae genomes, respectively (Fig. 1 and Table S1). The homeodomain (PF00046 and SM00389) was verified by using Pfam and SMART databases (Fig. 1). The amino acid length of both AcWOXs and AeWOXs proteins varied greatly from 199 a.a (AcWOX11a) to 354 a.a (AcWOX3a) and from 169 a.a (AeWOX4c) to 371 a.a (AeWOX10) (Table 1; Table S1). The predicted molecular weight of AcWOX and AeWOX proteins ranged from 22.3 to 39.2 kDa and from 19.4 to 42.2 kDa (Table 1). Moreover, the theoretical isoelectric point (pI) ranged from 5.30 to 9.75 for AcWOXs and from 5.66 to 9.76 for AeWOXs (Table 1). The subcellular localization of kiwifruit WOX proteins was predicted, and all of AcWOX and AeWOX proteins were localized in the nuclear (Table 1).
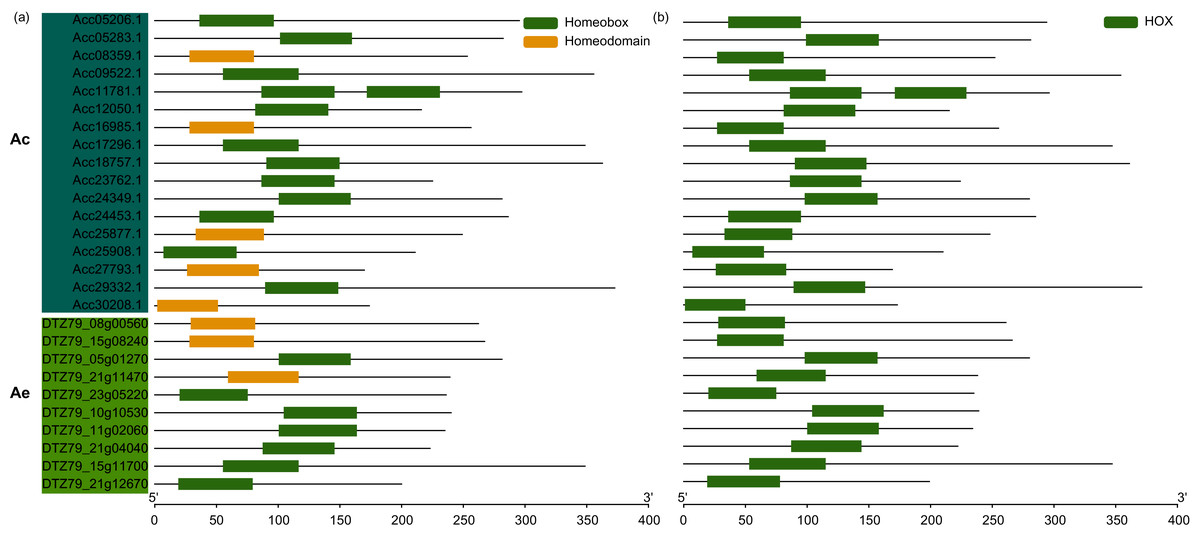
Figure 1: Conserved domain of WOX.
Conserved domain of kiwifruit WOX gene predicted by Pfam (A) and SMART (B). Ac, A. chinensis; Ae, A. eriantha.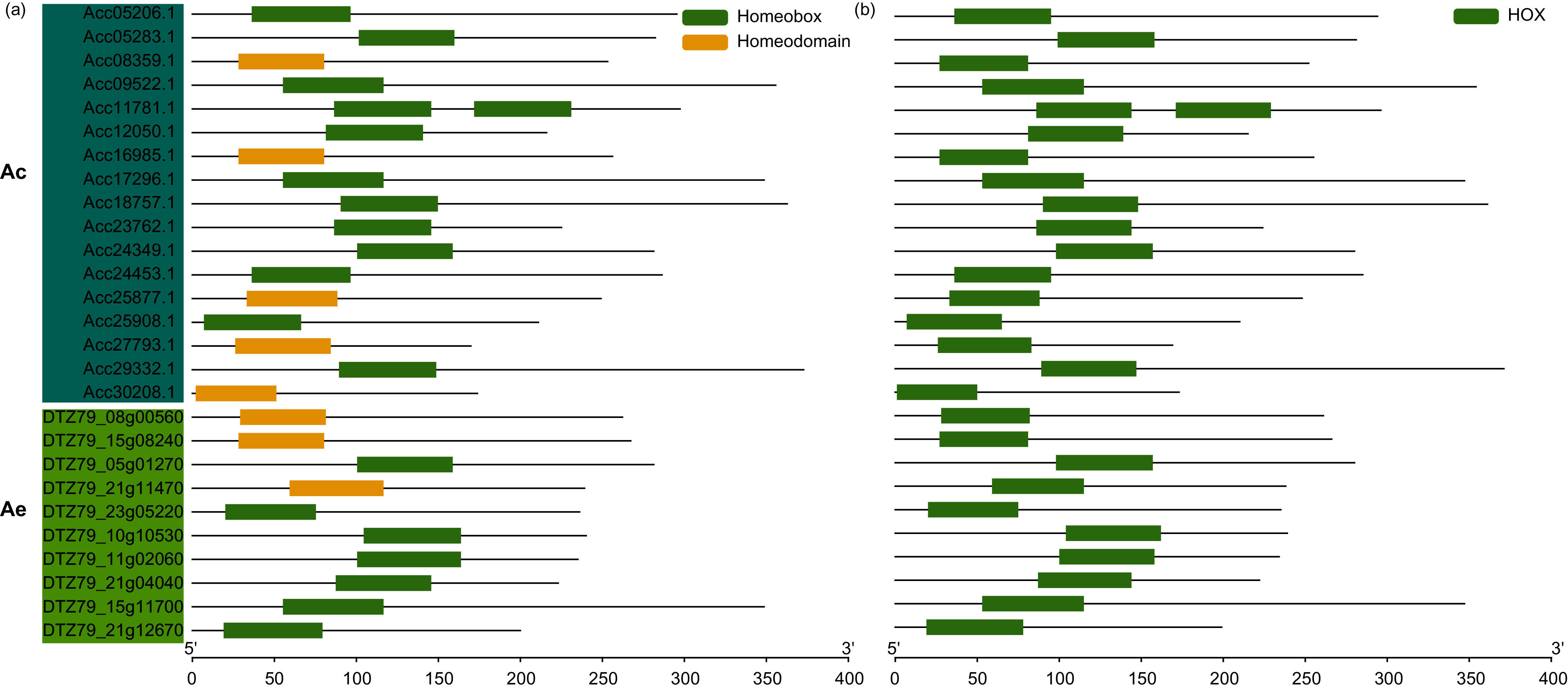{kind=link}
| Species | Genes | Genome ID | Protein Length (aa) | MW (Da) | pI | GRAVY | Predicted Localiaztion |
|---|---|---|---|---|---|---|---|
| A. chinensis | AcWUS1a | Acc05206.1 | 239 | 27,182.38 | 9.08 | −0.876 | Nuclear |
| AcWOX13a | Acc05283.1 | 266 | 29,050.87 | 5.49 | −0.448 | Nuclear | |
| AcWOX11a | Acc08359.1 | 199 | 22,315.54 | 9.75 | −0.808 | Nuclear | |
| AcWOX9a | Acc09522.1 | 280 | 32,003.07 | 5.87 | −0.974 | Nuclear | |
| AcWOX4a | Acc11781.1 | 222 | 25,554.82 | 9.51 | −0.909 | Nuclear | |
| AcWOX4b | Acc12050.1 | 261 | 28,424.23 | 5.39 | −0.423 | Nuclear | |
| AcWOX11b | Acc16985.1 | 238 | 27,037.52 | 5.30 | −1.021 | Nuclear | |
| AcWOX9b | Acc17296.1 | 235 | 26,266.99 | 9.47 | −0.829 | Nuclear | |
| AcWOX1a | Acc18757.1 | 347 | 37,969.27 | 8.30 | −0.461 | Nuclear | |
| AcWOX4c | Acc23762.1 | 236 | 27,155.85 | 5.70 | −1.118 | Nuclear | |
| AcWOX13b | Acc24349.1 | 294 | 32,910.35 | 5.79 | −0.980 | Nuclear | |
| AcWUS1b | Acc24453.1 | 281 | 32,134.20 | 5.79 | −0.996 | Nuclear | |
| AcWOX2 | Acc25877.1 | 252 | 27,361.96 | 5.55 | −0.466 | Nuclear | |
| AcWOX3a | Acc25908.1 | 354 | 39,260.64 | 8.28 | −0.606 | Nuclear | |
| AcWOX5 | Acc27793.1 | 296 | 34,123.19 | 9.46 | −0.922 | Nuclear | |
| AcWOX1b | Acc29332.1 | 215 | 24,485.44 | 9.05 | −0.817 | Nuclear | |
| AcWOX3b | Acc30208.1 | 255 | 27,862.43 | 5.38 | −0.567 | Nuclear | |
| A. eriantha | AeWOX13 | DTZ79_05g01270 | 361 | 41,243.21 | 6.74 | −0.871 | Nuclear |
| AeWOX11 | DTZ79_08g00560 | 224 | 25,913.02 | 9.20 | −0.970 | Nuclear | |
| AeWOX4a | DTZ79_10g10530 | 280 | 31,890.12 | 5.66 | −0.907 | Nuclear | |
| AeWOX4b | DTZ79_11g02060 | 285 | 31,718.21 | 6.32 | −0.925 | Nuclear | |
| AeWOX12 | DTZ79_15g08240 | 248 | 27,951.01 | 9.76 | −0.929 | Nuclear | |
| AeWOX9 | DTZ79_15g11700 | 210 | 24,383.34 | 8.58 | −0.769 | Nuclear | |
| AeWOX4c | DTZ79_21g04040 | 169 | 19,432.64 | 9.16 | −0.785 | Nuclear | |
| AeWOX10 | DTZ79_21g11470 | 371 | 42,184.49 | 9.04 | −0.875 | Nuclear | |
| AeWUS1 | DTZ79_21g12670 | 173 | 20,018.59 | 6.78 | −0.611 | Nuclear | |
| AeWOX2 | DTZ79_23g05220 | 200 | 22,510.43 | 6.84 | −0.615 | Nuclear |
Note:
Protein composition and physiochemical characteristics of kiwifruit WOX proteins.
Phylogenetic and molecular evolution analysis of kiwifruit WOXs
To explore the phylogenetic relationship and evolutionary pattern of WOX genes in kiwifruit, the neighbor-joining (NJ) tree was constructed using the full-length protein sequences of the identified 17 AcWOXs, 10 AeWOXs, and previously published 15 AtWOXs and 10 OsWOXs (WOX genes from Arabidopsis and rice). Consistent with previous reports in Arabidopsis, rice, and other species (Nardmann & Werr, 2012; Lin et al., 2013; Tang et al., 2017; Shafique Khan et al., 2021), both AcWOXs and AeWOXs were classified into three clades (the ancient clade, the intermediate clade, and the modern/WUS clade) (Fig. 2). Eleven of 17 AcWOXs and five of 10 AeWOXs were assigned in the modern/WUS clade, and the ancient clade contained the least number of both AcWOX and AeWOX genes (two AcWOXs and two AeWOXs) (Fig. 2). The intermediate clade had four AcWOXs and three AeWOXs (Fig. 2). AcWOXs and AeWOXs grouped with different WOX genes in Arabidopsis and rice indicated that both AcWOXs and AeWOXs probably had abundant diversified functions similar to WOX genes in Arabidopsis and rice (Fig. 2).

Figure 2: Phylogenetic tree of WOX.
Phylogenetic tree of the WOX proteins. The full-length WOX protein sequences from rice (Os, pink gene name and circles), Arabidopsis (At, blue gene name and circles), A. chinensis (Ac, red gene name and circles) and A. eriantha (Ae, orange gene name and circles) were aligned using ClustalX 2.0 with default parameters. Then, the unrooted phylogenetic tree was constructed using MEGA X and the Neighbour-Joining method. The ancient clade, intermediate clade, and modern/WUS clade were highlighted using pink, blue, and light red sectors, respectively.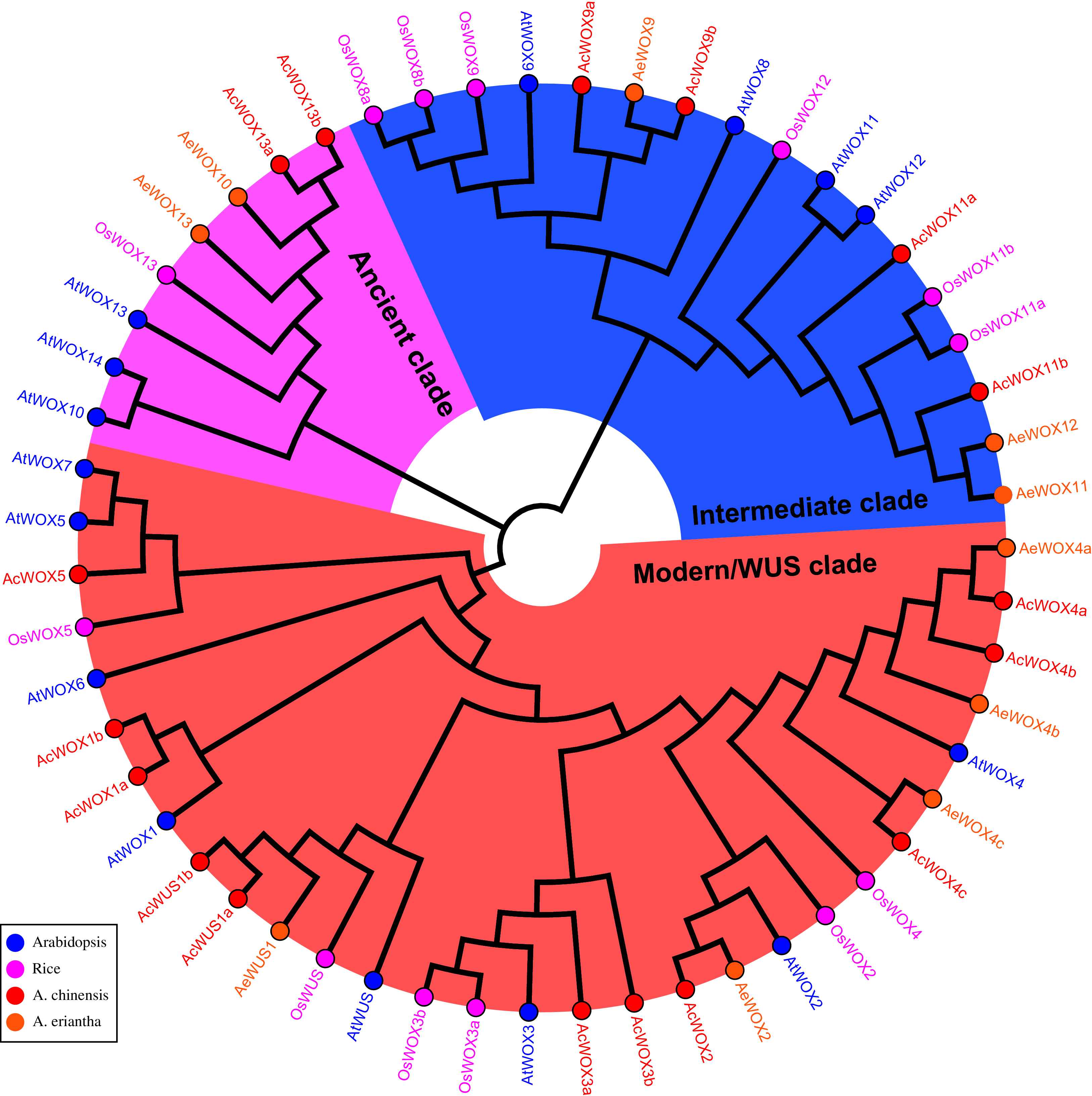{kind=link}
Chromosomal localization and structure analysis of kiwifruit WOXs
The 17 AcWOX genes were randomly distributed on 12 chromosomes of Ac (Fig. 3A), of which chromosome 8 and 21 included the most abundant WOX genes (three AcWOX genes), followed by three chromosomes (chr 5, 23, and 26) containing two genes and the other seven chromosomes (chr 3, 10, 11, 15, 16, 24, and 27) containing one AcWOX gene (Fig. 3A and Table S1). Similarly, the 10 AeWOX genes were unevenly distributed on seven chromosomes. Chr 21 and 15 contained three and two AeWOX genes, separately, and each of the other five chromosomes (chr 5, 8, 10, 11, and 23) possessed one AeWOX gene (Fig. 3B and Table S1).
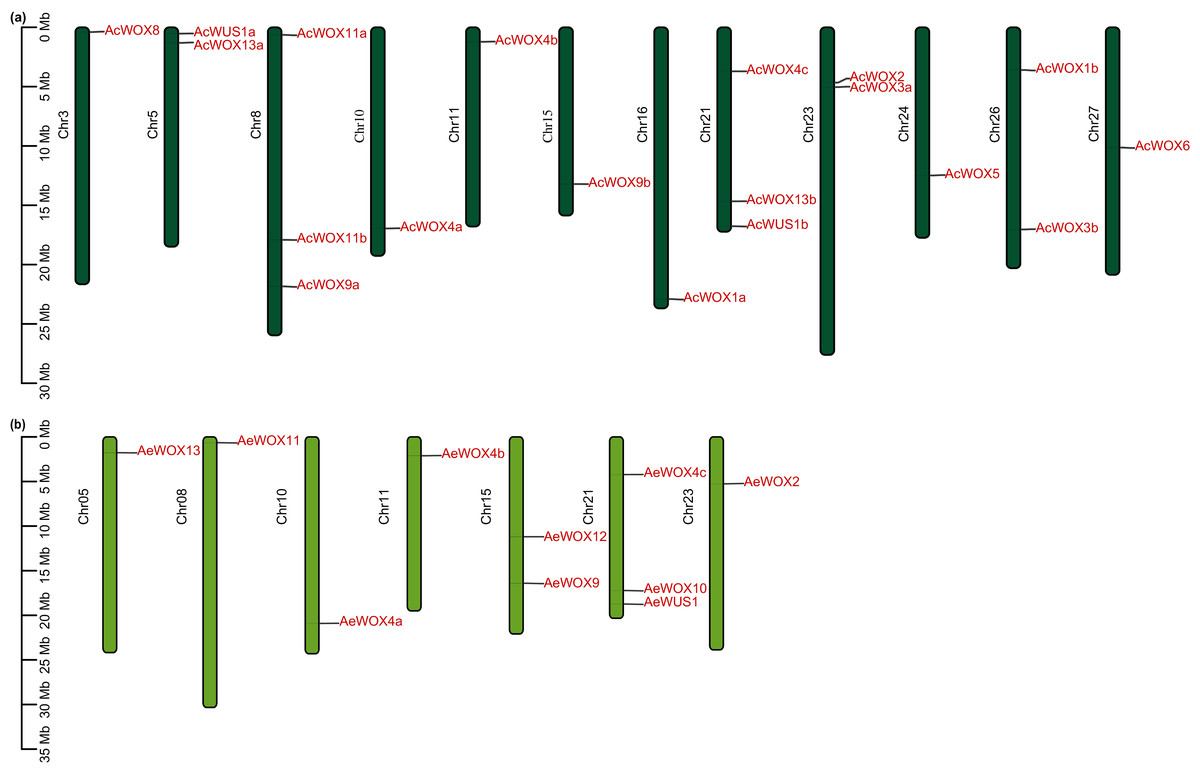
Figure 3: Chromosome location of WOX.
Distribution of WOX genes in Ac (A) and Ae (B) genomes.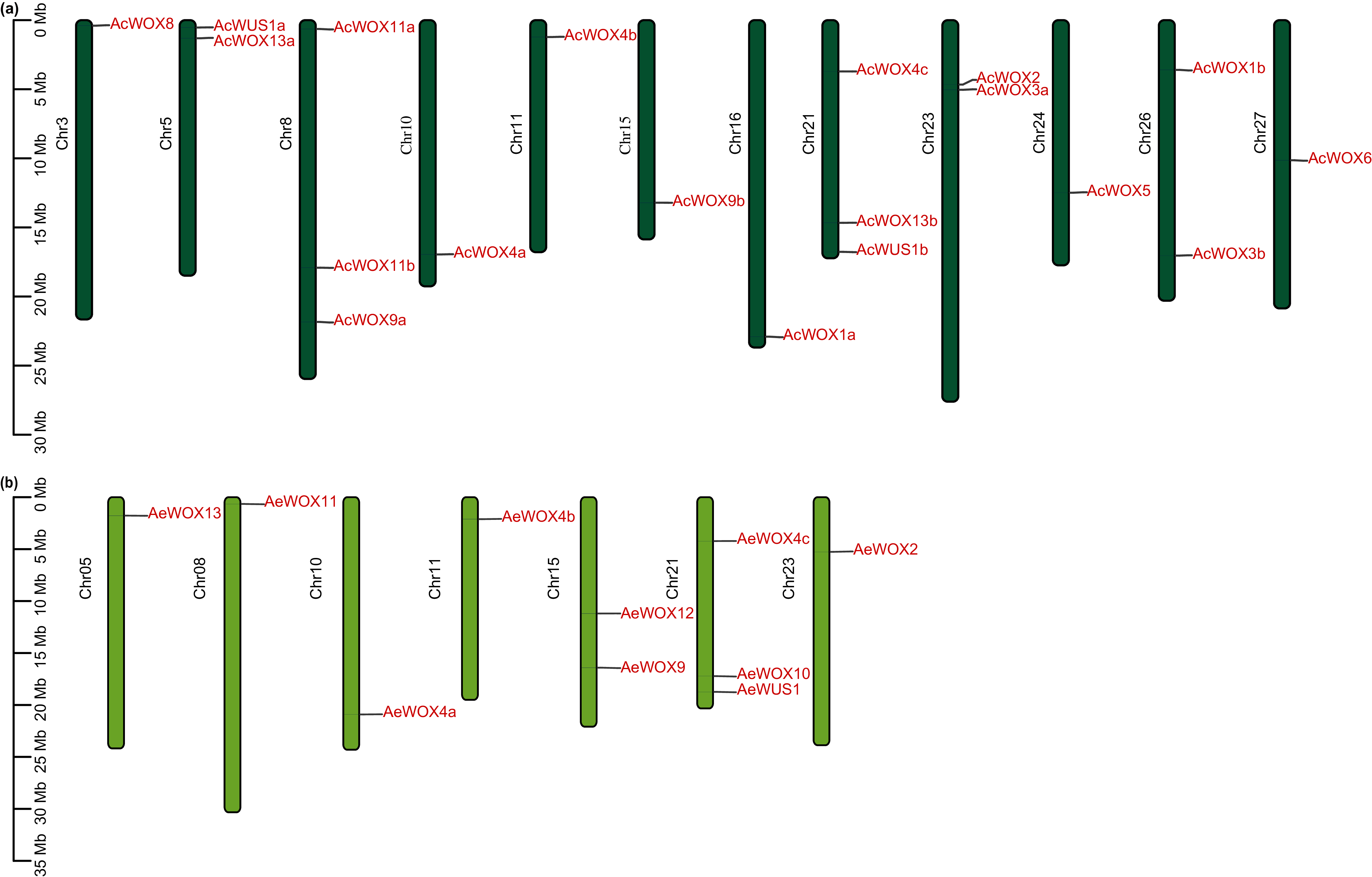{kind=link}
The exon-intron structure is a primary evolutionary feature of a gene family and provides a clue for function diversification and classification (Shiu & Bleecker, 2003). The exon number of AcWOXs and AeWOXs varied from two to four (Fig. 4). In terms of intron number and exon length, both AcWOXs and AeWOXs in the ancient clade were conserved for exon-intron structures (Fig. 4). In the intermediate clade, exon numbers of AeWOXs varied from two to four. In contrast, the AcWOXs in this clade had a fixed exon number, indicating that WOX genes had undergone varied evolutionary patterns in the two kiwifruit species. In each species, WOX genes in the same clade were more similar than those among different clades. Moreover, the exon-intron structure of both AcWOX and AeWOX genes diverged more in the modern/WUS clade, and this may result in the functional diversification of WOXs in this clade in two kiwifruits (Fig. 4).
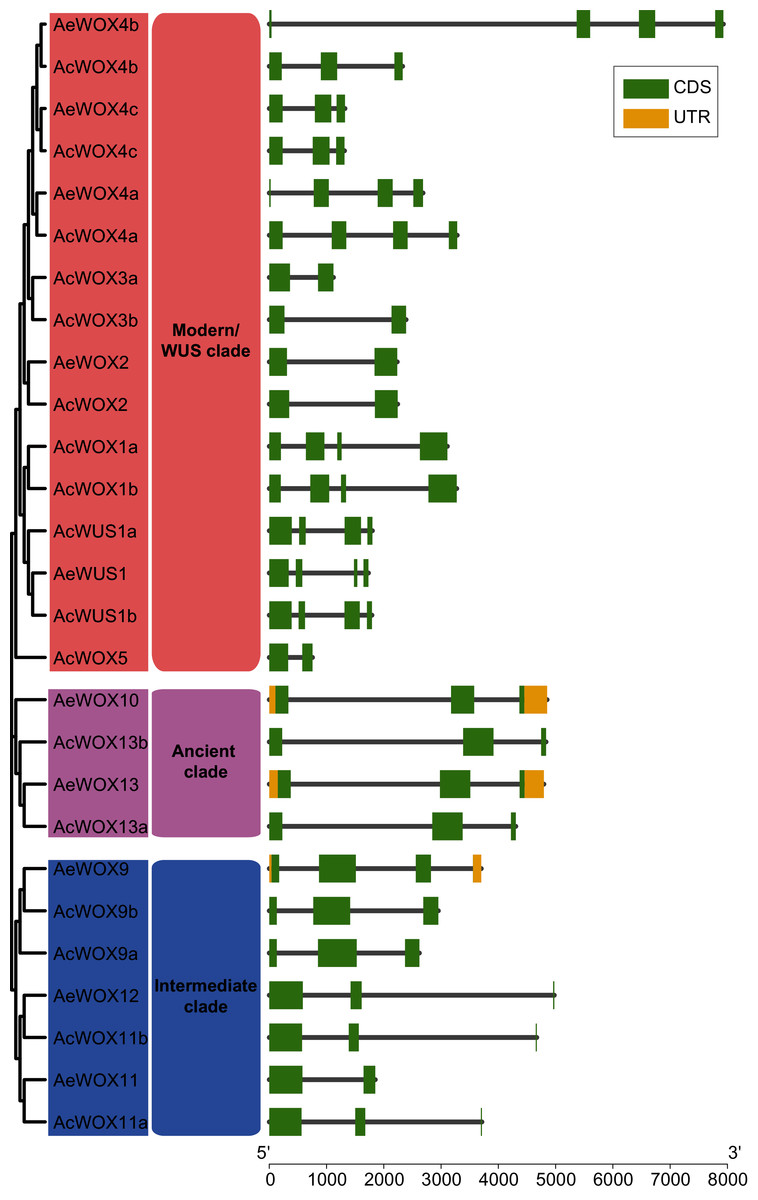
Figure 4: Structures of WOX genes.
Exon-intron structures of WOX genes in two kiwifruit species. The left panel indicated the phylogenetic tree containing AcWOX and AeWOX proteins; the middle panel showed three clades; the right panel showed exon-intron structures of kiwifruit WOX genes. The green rectangle, the yellow rectangle, and the regular line represent exons, UTRs, and introns, respectively.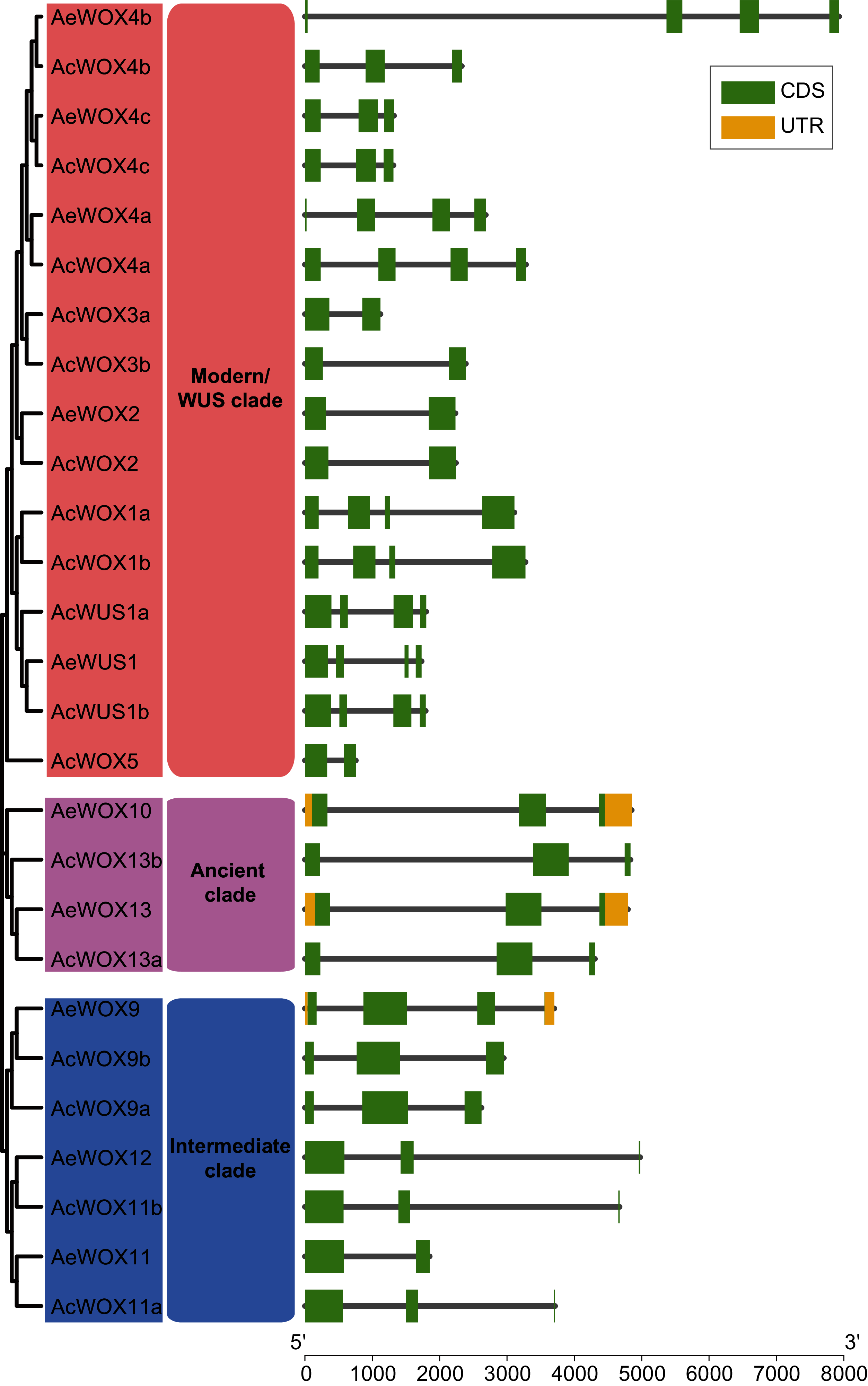{kind=link}
Domain architecture and conserved motifs analysis of kiwifruit WOXs
To infer the conserved motif architectures of kiwifruit WOXs, the motif compositions of entire sequences were predicted by MEME (Bailey et al., 2009) (Fig. 5 and Fig. S1). In total, 10 conserved motifs were identified for the kiwifruit WOXs and designated as motif 1–10 (Fig. S1). The motif number of the WOX gene ranged from two to eight in each kiwifruit (Fig. 5). Within the modern/WUS clade, the motif number varied greatly from two to eight, while motif numbers in the intermediate clades (five or six) and ancient clades (four or six) were stable (Fig. 5). All WOX genes in the two kiwifruit species contained motif 1, and we confirmed that motif 1 was the homeodomain with Pfam and SMART databases (Fig. 5). Furthermore, we found that motif 1 sequences in kiwifruit WOX genes were highly conserved (Fig. S2). Two and one clade-specific motifs were found in modern/WUS clade WOXs (motif 8 and 9) and ancient clade WOXs (motif 4), respectively (Fig. 5). Similar to results of exon-intron structure, closely phylogenetically related WOX genes showed conserved motif structures, including motif number and organization, which indicated similar functions among them (Fig. 5).
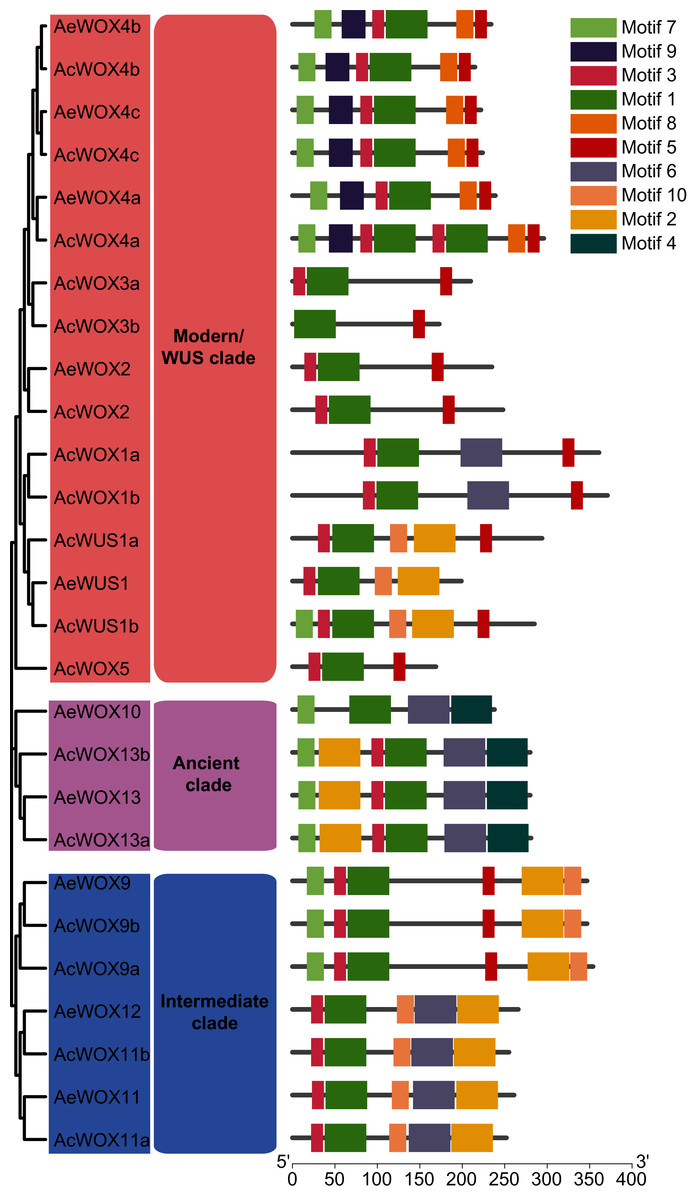
Figure 5: Motifs of WOX.
Motif architectures of kiwifruit WOX proteins. The left panel indicated the phylogenetic tree of AcWOX and AeWOX protein sequences; the middle panel showed the defined clades; the right panel showed motif architectures of WOX proteins. Rectangles with different colors represented different motifs.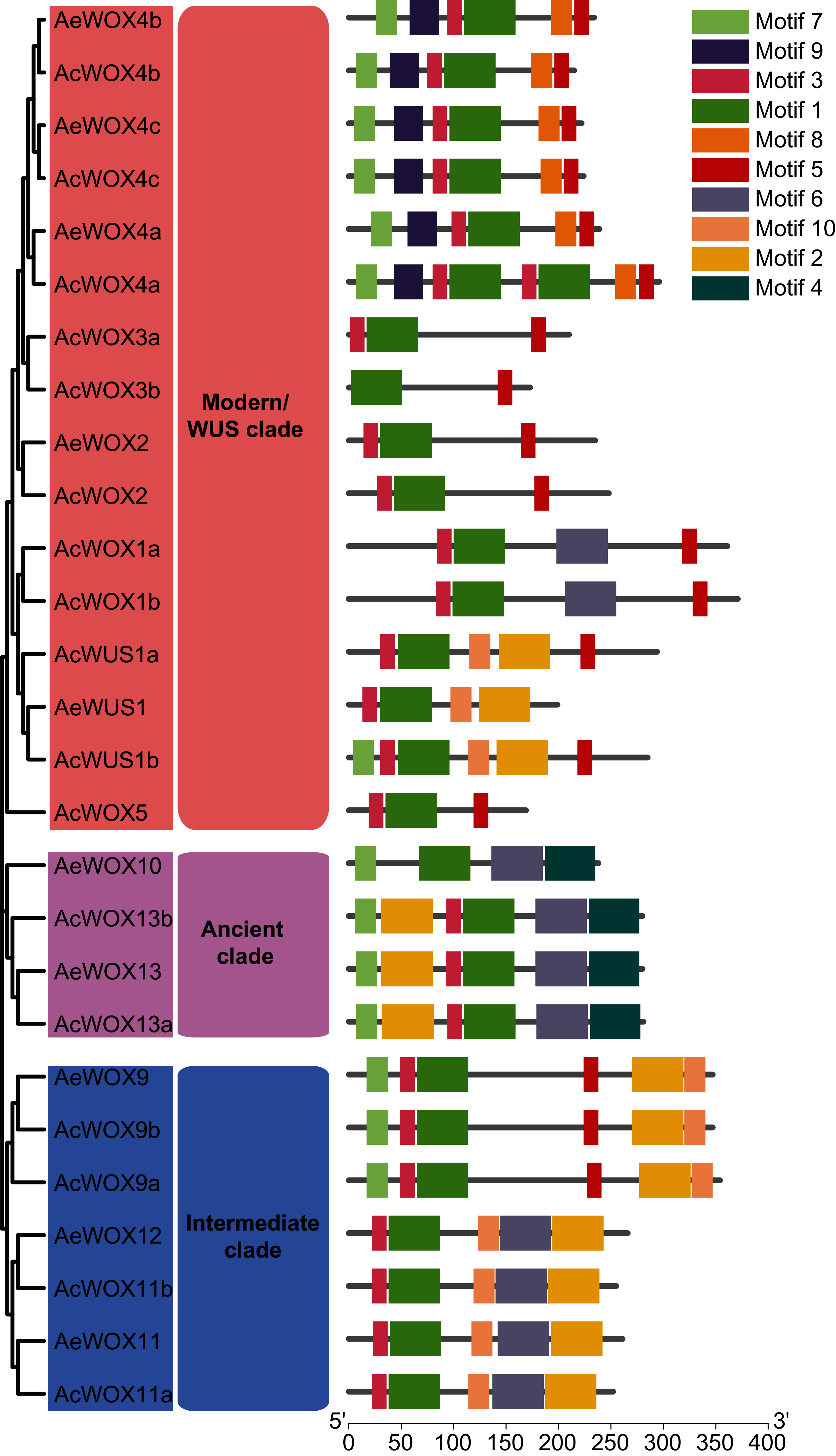{kind=link}
Synteny and duplicated gene analysis of kiwifruit WOXs
Gene duplication and loss were the main evolutionary forces driving the expansion or contraction of gene families, and duplicated genes could result in gene redundancy or new functionalization (Tang et al., 2020). To visualize the synteny relationships among homologous WOX genes and infer gene duplication events, we conducted a collinearity analysis by using MCScanX (Wang et al., 2012). Accordingly, we determined that gene pairs belonging to five types of gene duplication (singleton duplication (SD), dispersed duplication (DD), proximal duplication (PD), tandem duplication (TD), and whole-genome duplication (WGD)). We identified eight and five pairs of genes that resulted from duplication in Ac and Ae kiwifruit, respectively (Fig. 6 and Table 2). All duplicated gene pairs belonged to the modern/WUS and the intermediate clades, indicating these kiwifruit WOXs diverged considerably and had more potential to be new functionalized than that in the ancient clade (Fig. 6 and Table 2). All duplicated gene pairs were produced by the whole-genome duplication, indicating that the WGD accounted for the expansion of kiwifruit WOX families (Table 2).

Figure 6: Synteny relationship of WOX.
Chromosome distribution and synteny relationship of WOX genes in two kiwifruit species. The light red and green bars indicated chromosomes for Ac and Ae, respectively. The syntenic gene pairs were connected by lines with different colors.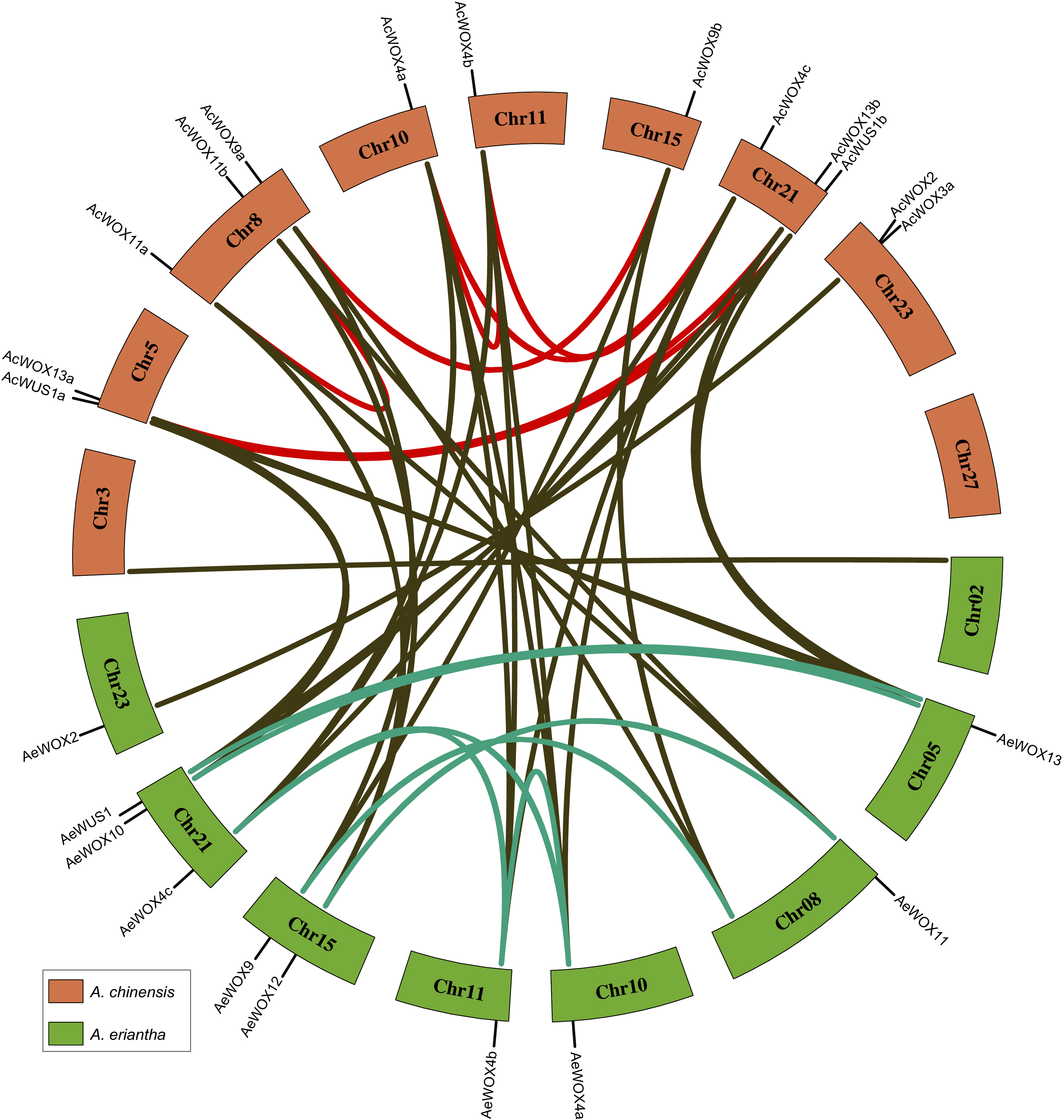{kind=link}
| Species | Duplicate pairs | Ka | Ks | Ka/Ks | Duplication |
|---|---|---|---|---|---|
| A. chinensis | AcWOX4a/AcWOX4b | 0.06322 | 0.23011 | 0.27473 | WGD |
| AcWOX4a/AcWOX4c | 0.18728 | 0.71925 | 0.26038 | WGD | |
| AcWOX4b/AcWOX4c | 0.16292 | 0.70114 | 0.23237 | WGD | |
| AcWOX9b/AcWOX9a | 0.06494 | 0.20962 | 0.30979 | WGD | |
| AcWUS1b/AcWUS1a | 0.02622 | 0.15924 | 0.16464 | WGD | |
| AcWOX13b/AcWOX13a | 0.03055 | 0.22185 | 0.13769 | WGD | |
| AcWOX3a/AcWOX3b | 0.15948 | 0.52930 | 0.30131 | WGD | |
| AcWOX11a/AcWOX11b | 0.04509 | 0.15773 | 0.28589 | WGD | |
| A. eriantha | AeWOX13/AeWOX10 | 0.02696 | 0.22560 | 0.11951 | WGD |
| AeWOX11/AeWOX12 | 0.05723 | 0.16531 | 0.34620 | WGD | |
| AeWOX4a/AeWOX4b | 0.12001 | 0.35079 | 0.34212 | WGD | |
| AeWOX4a/AeWOX4c | 0.18328 | 0.68682 | 0.26686 | WGD | |
| AeWOX4b/AeWOX4c | 0.20150 | 0.78790 | 0.25574 | WGD |
Note:
WOX duplication events identified in kiwifruits.
To estimate the selection pressure that kiwifruit WOXs experienced after the gene duplication, we calculated the Ka/Ks ratios, ratios of the rate of nonsynonymous substitution (Ka) to the rate of synonymous substitution (Ks) (Table 2 and Table S2). Generally, the Ka/Ks value reflects the selection pressure during evolution (Ka/Ks = 1: neutral selection; Ka/Ks > 1: positive selection; Ka/Ks < 1: purifying selection) (Alvarez et al., 2018). In this study, Ka/Ks ratios of WOXs ranged from 0.096 to 0.436, with an average of 0.259 (Table 2 and Table S2). These results suggested that purifying selection was the primary evolutionary force acting on kiwifruit WOXs.
Cis-element analysis of promoter regions of kiwifruit WOXs
The cis-element plays a crucial control in transcriptional regulation and significantly affects gene function (Nakashima, Ito & Yamaguchi-Shinozaki, 2009). The 1,500-bp upstream region of each kiwifruit WOX gene was extracted and employed for the cis-element prediction. Totally, 17 functional cis-elements were retained, and the core promoter elements, such as TATA-box and CAAT-box, were presented in all promoters of kiwifruit WOXs. The 17 functional cis-elements were classified into four subfamilies, including light responsiveness, plant growth and development, phytohormone responsive, and stress-responsive subfamily (Fig. 7A). Elements of the light responsiveness subfamily were the most abundant presented in the promoter regions of kiwifruit WOX genes of the two species, indicating that light could significantly affect expression patterns of kiwifruit WOXs and gene function (Fig. 7B). The number of light responsiveness cis-element in all kiwifruit WOXs ranged from 0 (AcWOX3b) to 11 (AcWOX13b) (Figs. 7A and 7B). The number of the light responsiveness cis-element within WOXs differed between two kiwifruit species (Fig. 7C). The cis-element organizations for the duplicated gene pairs were evolved divergently (Fig. 7A), suggesting the specific expression patterns and new functionalization for the duplicated gene pairs. However, the cis-element arrangements of the orthologous WOX gene pairs for the two species were highly similar, indicating that the orthologous WOX gene pairs possibly had similar functions (Fig. 7A).
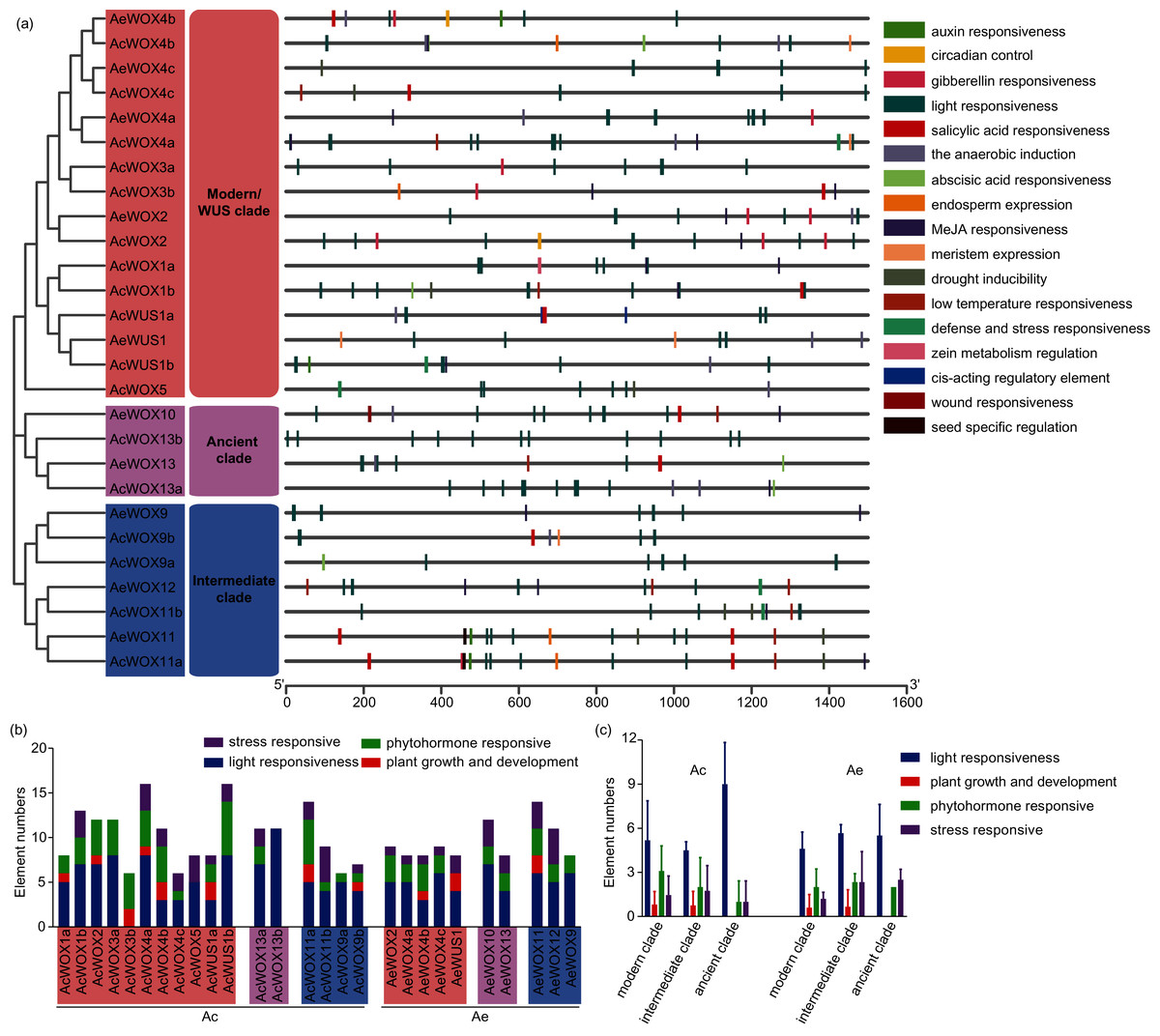
Figure 7: Cis-elements of WOX.
Cis-elements analysis in the promoter regions of kiwifruit WOX genes. (A) The cis-element architectures in the 1,500-bp promoter regions of kiwifruit WOXs. Rectangle with different colors represented different cis-elements. (B) The number of cis-elements in the promoter region of each kiwifruit WOX gene. (C) The average number of cis-elements for each clade was showed.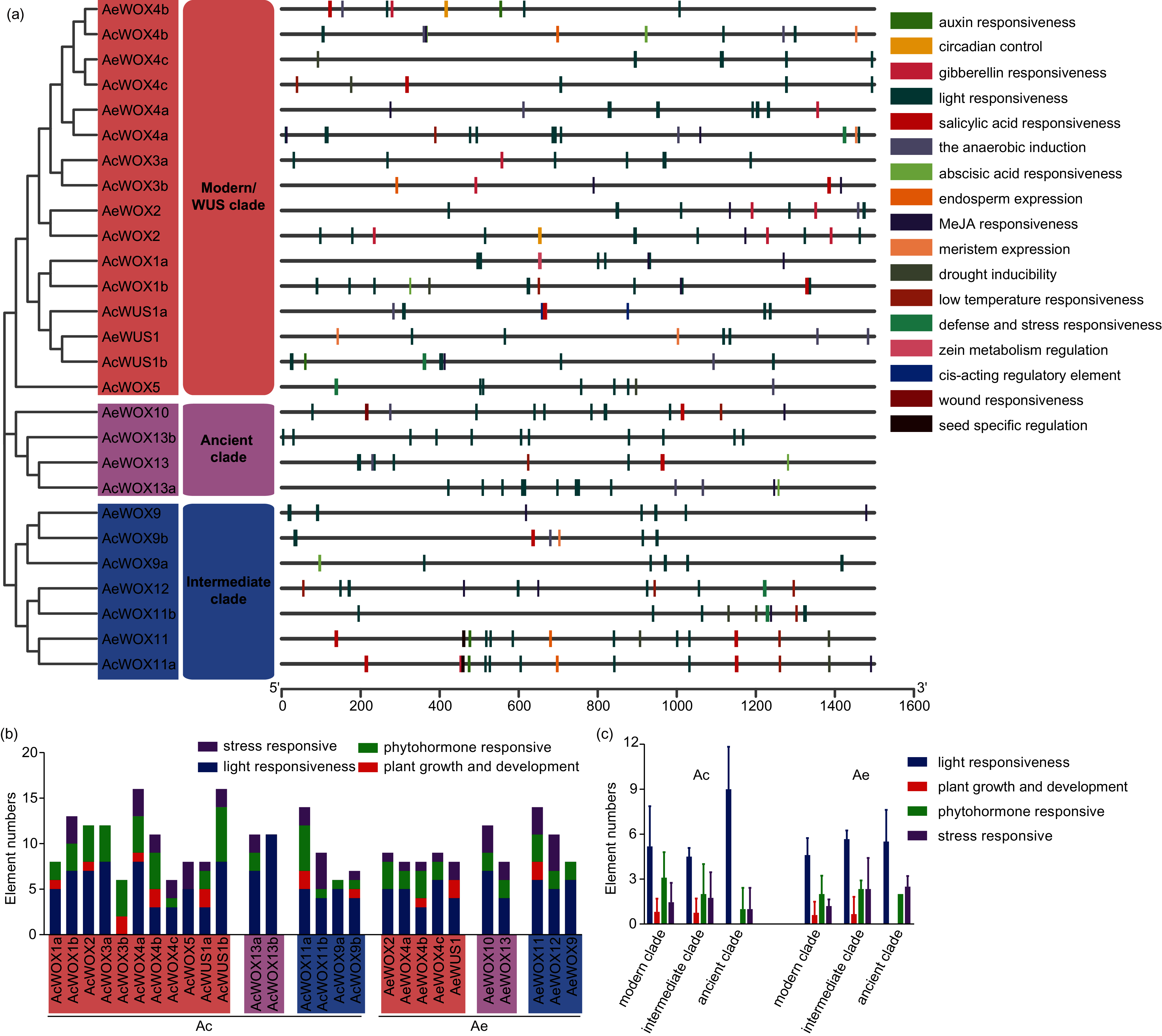{kind=link}
Expression patterns of kiwifruit WOXs
Firstly, we investigated the expression patterns of AcWOX genes in three tissues (leaf, root, and stem) based on transcriptome data (Fig. 8A). Our results showed that AcWOX genes had highly tissue-specific expression patterns (Fig. 8A). AcWOX13a and AcWOX13b highly expressed in all three tissues but more abundant in leaf and root, indicating that expression of AcWOX13a/13b might be essential for the leaf and root (Fig. S3). The other four AcWOXs, including AcWOX4a/4b/11a/11b, were expressed lowly in three tissues (Fig. S3). While the AcWOX5 is expressed explicitly in the root (Fig. 8A and Fig. S3). We further analyzed the expression profiles of AcWOX genes during fruit development. Four AcWOX genes highly expressed in fruits, and they gradually increased (AcWOX13a) or decreased (AcWOX4a/4b/13b) their expressions along with fruit development (Fig. 8B). These indicated that these four AcWOXs might affect fruit development differently (Fig. S3B).
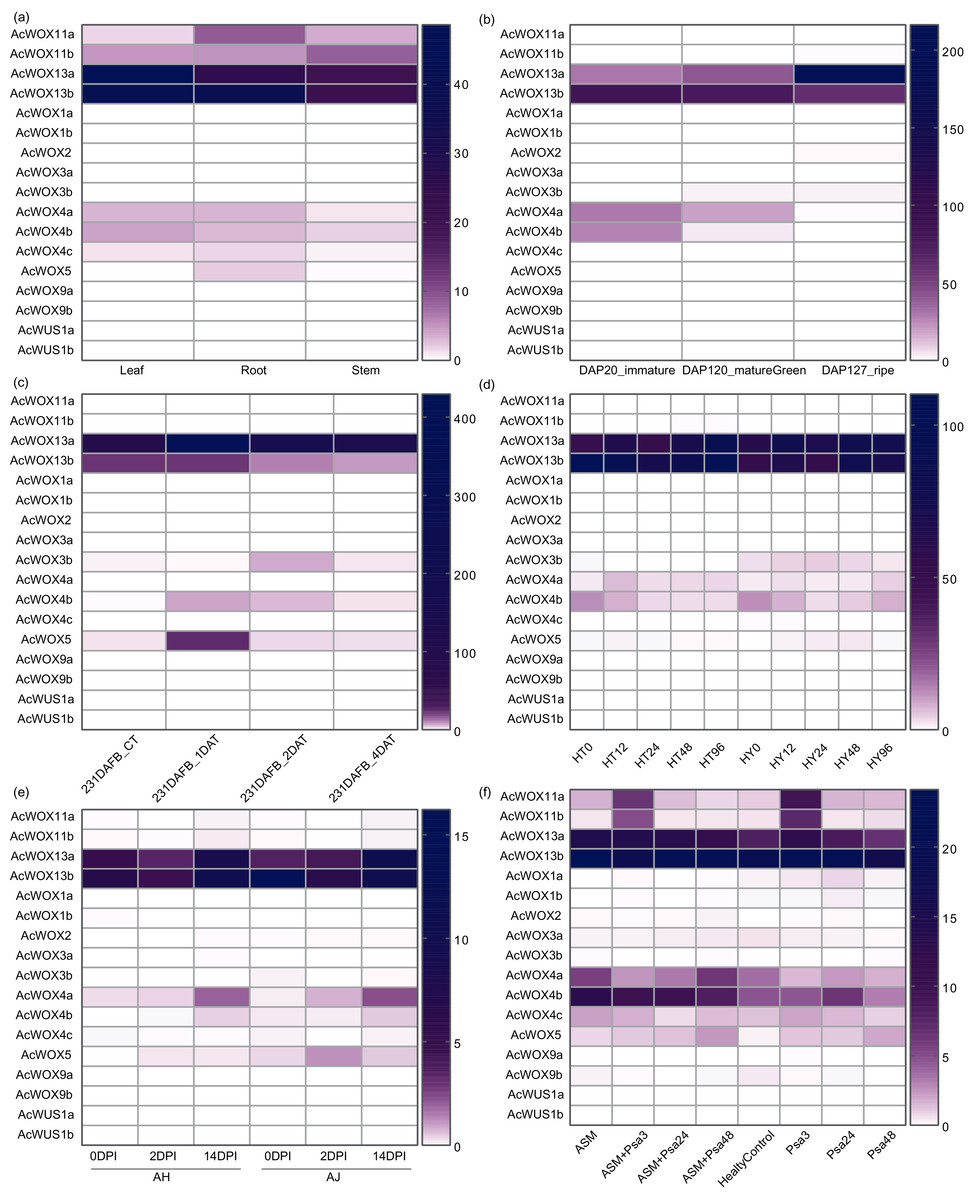
Figure 8: Expression patterns of WOX.
Expression profiles of AcWOX genes in different tissues, fruit developmental stages, and under different treatments. The heatmap indicated the FPKM (fragments per kilobase of exon model per million mapped reads) values of AcWOX genes. (A) Expression profiles of AcWOXs in three tissues. (B) Expression profiles of AcWOXs in three fruit developmental stages. DAP, days after pollination. (C) Expression profiles of AcWOXs in samples treated with ethylene. DAFB, days after full bloom of fruit; DAT, day after treated with ethylene. (D) Expression profiles of AcWOXs in two kiwifruit cultivars infected with Psa. HT and HY represented resistant and susceptible cultivar, respectively. The number following the cultivar name showed hours post the Psa invasion. (E) Expression profiles of AcWOXs in two susceptible cultivars to the invasion of Psa. DPI, days post-infection. (F) Expression profiles of AcWOXs in samples with or without Acibenzolar-S-methyl (ASM) treatment during the Psa infection. Healtycontrol represented samples without any treatment or infection. The number presented in the sample names indicated hours post the Psa infection.{kind=link}
To further confirm whether the expression of WOX genes was influenced by hormonal treatments and biotic stresses, we analyzed transcriptomic data from Ac fruits which treated with ethylene (Gunaseelan et al., 2019) or invaded with Pseudomonas syringae pv. actinidiae (Psa) which was the pathogen of kiwifruit bacterial canker disease (Michelotti et al., 2018). Totally, five AcWOX genes significantly responded to ethylene treatments (Fig. 8C). Among them, four genes (AcWOX3b/4b/5/13a) were upregulated by ethylene treatment and gradually reduced their expressions after the treatment, and the AcWOX13a had the highest expression level (Fig. S3C). Besides, AcWOX13b was downregulated by the ethylene treatment (Fig. S3C).
We detected AcWOX gene expression profiles in response to Psa infection with three sets of transcriptomic comparisons, i.e., (a) comparative transcriptomes of the resistance cultivar and the susceptible cultivar (Fig. 8D); (b) two susceptible cultivars (Fig. 8E); (c) with or without Acibenzolar-S-methyl (ASM) treatment (Fig. 8F). In the first comparison, we identified five highly expressed AcWOX genes with varied transcriptional levels among different infection times (Fig. 8D). AcWOX13a and AcWOX13b displayed abundant expression in both cultivars and showed higher expressions in the resistant variety (HT) than in the susceptible variety (HY) (Fig. 8D and Fig. S3D), indicating that AcWOX13a and AcWOX13b might affect kiwifruit resistance to Psa. Similar to the results in the first set, transcriptional levels of AcWOX13a and AcWOX13b were higher than others under the infection of Psa in the second set of comparison (Fig. 8E and Fig. S3E). The third set of the comparison showed that ASM inoculated kiwifruits increased their expression of AcWOX13a/13b/4a/4b compared with samples without the ASM treatment (Fig. 8F and Fig. S3F).
Discussion
The WOX gene family is the core regulator for forming the shoot apical meristem and embryonic development, stem cell maintenance, and various other developmental processes in plants (Ariel et al., 2007; Wu, Chory & Weigel, 2007; Deveaux et al., 2008; Shimizu et al., 2009; van der Graaff, Laux & Rensing, 2009; Palovaara et al., 2010). Genome-wide identification of the WOX gene family had been accomplished in several plants (Deveaux et al., 2008; Liu et al., 2014b; Cheng et al., 2014; Tang et al., 2017; Rahman et al., 2017; Li et al., 2018, 2020; Ramkumar et al., 2018; Shafique Khan et al., 2021). However, genome-wide characterization of the WOX gene family had not been conducted in kiwifruits. In the present study, we in silico identified the genome-wide WOX gene family in two kiwifruit species (Ac and Ae). Further, we compared the characters and evolutionary patterns of kiwifruit WOXs of the two species. In addition, we analyzed and compared cis-element organizations for promoter regions of kiwifruit WOXs identified in the two species. We also investigated the expression profiles of AcWOXs in different tissues and different fruit developmental stages and the influence of hormonal treatment and biotic stresses on AcWOXs expression.
Seventeen and 10 WOX genes were identified in Ac and Ae, respectively (Fig. 1 and Table 1). Compared to the WOX family in Arabidopsis, the Ac has more while Ae has fewer family members. Interestingly, no WOX genes from Ac and Ae are classified together with AtWOX6, and no WOX genes from Ae are classified together with AtWOX1/5/7. This may be due to the gene loss in the evolution of Actinidia species. The lost of WOX6 homologous gene has also been reported in many plants including Orchidaceae species (Kanchan & Sembi, 2020; Victorathisayam et al., 2020), Solanaceae species (Li et al., 2018), and cucumber (Han et al., 2021). AtWOX6 regulates ovule development and plays a role in freezing tolerance in Arabidopsis (Park et al., 2005). However, WUS and WOX6 genes possess some similar functions and could be replaced mutually sometimes (Dolzblasz et al., 2016). WOX1 and WOX3 has been demonstated redundantly control leaves development in Petunia, Arabidopsis and tomatos (Vandenbussche et al., 2009; Nakata et al., 2018; Wang et al., 2021). WOX6 and WOX7 comprise close relatives of WOX1 and WOX5, respectively, and are possibly due to the recent whole-genome duplication (Blanc & Wolfe, 2004). These suggest that WOX gene family members could be replaced, and it may make their losing in evolution possible. WOX1 has been domenstrated that was possibly lost in monocots, where grass genomes manifested a WOX3 duplication (Nardmann & Werr, 2013).
Duplicated WOX gene pairs in both Ac and Ae were entirely owing to WGD (Fig. 6 and Table 2). Genomic analyses verified that both Ac and Ae genomes experienced three ancient WGD events (Pilkington et al., 2018; Tang et al., 2019), which supported our results. However, the difference of WOX gene number between Ac and Ae indicated that the WOX gene family in Ac and Ae had undergone inconsistent evolutionary patterns (Pilkington et al., 2018; Tang et al., 2019). Together with the asymmetric gene losing in these two kiwifruit species, WOX gene family members are species-specific in Actinidia genus. In addition, we inferred that translocation, gene retention, and gene loss post whole-genome duplications were the evolutionary forces causing variations of the WOX gene number and distribution in Ac and Ae genomes. Signals of purifying selection were found for all WOX genes in both species, which indicated the crucial roles of kiwifruit WOXs.
Homeodomain, a domain that contains a helix-loop-helix-turn-helix structure (Katoh & Standley, 2013), is conserved in the WOX gene family among different species and thus maintains the functional integrity of WOX genes. Except for the homeodomain, we additionally identified nine conserved motifs in kiwifruit WOXs (Fig. 5). Clade-specific or subclade-specific motifs in kiwifruit WOXs genes suggested the functional differentiation among WOXs in different clades (Fig. 5). However, gene structures and conserved motif organizations of most kiwifruit WOXs belonging to the same subclade were consistent. The cis-element analysis of promoter regions of kiwifruit WOXs indicated that the cis-element organizations of kiwifruit WOXs were extremely varied, even for those in the same subclade (Fig. 7). We inferred that the cis-element organizations of WOXs belonging to the same subclade regulated the functional divergence of those WOXs by regulating their expression patterns, which was verified by the results of expression analysis (Fig. 8). In total, gene structure, motif organization, and cis-element arrangement precisely controlled the functional divergent of kiwifruit WOXs.
Promoters are regions of DNA that initiate transcription of particular genes. They play prominent roles in the temporal and spatial regulation of gene expression. In the present study, a series of development-, hormone-, response- and stress- related cis-elements were detected in the promoter of kiwifruit WOXs (Fig. 7). Each WOX gene harbours at least six cis-elements belonging to at least two different functional subfamilies, except AcWOX13b which has 11 light responsiveness cis-elements (Fig. 7). Accumulating evidence has demonstrated that plant stress resistance is regulated by phytohormones (Chaiprasongsuk et al., 2018; Ditengou et al., 2018). We suspect that expression of WOX genes were influenced by stress, and it was uncovered by RNA-seq results in Ac. However, the relationship between the stress resistance and phytohormones needs more evidences.
Gene expression patterns in different tissues or under different treatments can be used to identify the functions of genes. Expression patterns of WOX genes are different either between A. thaliana and Populus tomentosa (Liu et al., 2014b) or between A. thaliana and rice (Zhang et al., 2010). In this study, expression patterns of WOX genes in kiwifruits were different from those plants informed above (Fig. 8). These implied that the functions of WOX genes might be different among monocots and dicotyledones of herbal plants, woody plants and liane.
In AcWOX and AeWOX members, few genes were mainly expressed in specific organs, indicating that they may be involved in different development processes. In Ac kiwifruit, AcWOX13a and AcWOX13b expressed higher than other WOXs, and they often respond to stresses dramatically (Fig. 8). This indicated the pivotal role of WOXs of the ancient clade in kiwifruit development and stress resistance. Similar functioned WOX gene (OsWOX11 and OsWOX13) has been reported in rice (Tvorogova et al., 2021). OsWOX11, a member of the intermediate clade, was the most reported, responding to abiotic stresses such as drought and cold reviewed in (Tvorogova et al., 2021). Minh-Thu reported that OsWOX13, an ancient clade WOX gene, enhances drought tolerance and triggers early flowering in rice (Shafique Khan et al., 2021). In poplar, over-expression of PtoWOX13 led to an increased adventitious root (AR) number, deceased AR length or increased AR roughtnees (Liu et al., 2014b). In walnut, JrWOX13 plays an important role in later root development (Chang et al., 2020). It is unclear whether AcWOX13a and AcWOX13b regulate the stress resistance via root development. Further study of the precise function of AcWOX13a and AcWOX13b will be needed in the future.
In summary, the present study firstly detected WOX genes from kiwifruit genomes. Furthermore, we identified the potential WOXs in response to hormonal treatments and biotic stresses. Our research will provide a foundation for accelerating the genetic breeding of kiwifruits.
Conclusions
In conclusion, we performed genome-wide identification and characterization of the WOX gene family in Actinidia chinensis (Ac) and A. eriantha (Ae) and conducted a detailed investigation of their evolutionary relationship, genome organization, duplication events and expression profiles. In total, 17 and 10 WOX genes have been identified from Ac and Ae genomes. They were classified into three clades (the ancient clade, the intermediate clade, and the morden/WUS clade) based on the sequence alignment and phylogenetic analysis, which was corresponding to previous report of Arabidopsis. Gene structures and motif patterns showed that WOX members in the same clade displayed more similarly. Gene duplications and selective pressure analysis indicated that WOX genes in two kiwifruit species have undergone different evolution patterns after the shared genome duplication events. Subcellular localization revealed that all WOX genes were located inside the nucleus. Furthermore, expression patterns detected tissue-specific expressed genes and hormonal or abiotic-stress responding genes. Overall, WOX genes identified in Ac and Ae genomes provided insight into general characters, evolutionary patterns, and functional diversity of kiwifruit WOXs.
Supplemental Information
Logos of motifs.
Sequence logos for the ten conserved motifs identified in the kiwifruit WOX gene family.
Homeodomain.
Sequence alignment of the homeodomain in the kiwifruit WOX proteins.
Expression profiles of WOX genes.
Expression profiles of WOX genes. AcWOXs with high expression levels in three tissues and three fruit developmental stages were shown in (a) and (b). DAP, days after pollination. AcWOXs with high expression levels in samples treated with ethylene was shown in (c). DAFB, days after full bloom of fruit; DAT, day after treated with ethylene. (d) showed AcWOXs with high expression levels in two kiwifruit cultivars infected with Psa. HT and HY represented resistance and susceptible cultivar, respectively. The number in cultivar names showed hours post the Psa invasion. (e) shows expression profiles of AcWOXs with high expression levels in two susceptible cultivars to the invasion of Psa. DPI, days post-infection. (f) was expression profiles of AcWOXs with high expression levels in samples with or without Acibenzolar-S-methyl (ASM) treatments during the Psa infection. Healtycontrol represents samples without ASM treatment and Psa infection. The number presented in sample names indicated hours post the Psa infection.