
Association between C-Maf-inducing protein gene rs2287112 polymorphism and schizophrenia
- Published
- Accepted
- Received
- Academic Editor
- Bao-Liang Zhong
- Subject Areas
- Cognitive Disorders, Epidemiology, Public Health, Women’s Health, Medical Genetics
- Keywords
- C-Maf-inducing protein, CMIP, Haplotype analysis, Gene polymorphism, Schizophrenia
- Copyright
- © 2021 Fu et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. Association between C-Maf-inducing protein gene rs2287112 polymorphism and schizophrenia. PeerJ 9:e11907 https://doi.org/10.7717/peerj.11907
Abstract
Background
Schizophrenia is a severely multifactorial neuropsychiatric disorder, and the majority of cases are due to genetic variations. In this study, we evaluated the genetic association between the C-Maf-inducing protein (CMIP) gene and schizophrenia in the Han Chinese population.
Methods
In this case-control study, 761 schizophrenia patients and 775 healthy controls were recruited. Tag single-nucleotide polymorphisms (SNPs; rs12925980, rs2287112, rs3751859 and rs77700579) from the CMIP gene were genotyped via matrix-assisted laser desorption/ionization time of flight mass spectrometry. We used logistic regression to estimate the associations between the genotypes/alleles of each SNP and schizophrenia in males and females, respectively. The in-depth link between CMIP and schizophrenia was explored through linkage disequilibrium (LD) and further haplotype analyses. False discovery rate correction was utilized to control for Type I errors caused by multiple comparisons.
Results
There was a significant difference in rs287112 allele frequencies between female schizophrenia patients and healthy controls after adjusting for multiple comparisons (χ2 = 12.296, Padj = 0.008). Females carrying minor allele G had 4.445 times higher risk of schizophrenia compared with people who carried the T allele (OR = 4.445, 95% CI [1.788–11.046]). Linkage-disequilibrium was not observed in the subjects, and people with haplotype TTGT of rs12925980–rs2287112–rs3751859–rs77700579 had a lower risk of schizophrenia (OR = 0.42, 95% CI [0.19–0.94]) when compared with CTGA haplotypes. However, the association did not survive false discovery rate correction.
Conclusion
This study identified a potential CMIP variant that may confer schizophrenia risk in the female Han Chinese population.
Introduction
Schizophrenia (SCZ) is a severely multifactorial neuropsychiatric disorder that affects almost 1% of adults around the world. A recent study found that the lifetime prevalence of SCZ patients in China was 0.6% (Huang et al., 2019). SCZ has devastating impacts on patients’ and their families’ quality of life. It also has an enormous financial cost. SCZ is a prototypical multifactorial disorder caused by both genetic and environmental factors. Genetic factors play a major role in SCZ etiology (Owen, Sawa & Mortensen, 2016) and genetic variations in chromosome 16 are associated with a variety of neuropsychiatric disorders. Some rare, common, and copy number variants on chromosome 16p have been found to be associated with SCZ (Chang et al., 2017; Giaroli et al., 2014). Regions on chromosome 16q, highly specific to a single psychometric measure, are also associated with neuropsychiatric disorders. Previous studies found that regions on chromosome 16q may increase susceptibility to SCZ (Lewis et al., 2003), bipolar disorder (Lewis et al., 2003), and autism (Wassink et al., 2008). Furthermore, large-scale genome-wide association studies (GWAS) conducted by Bigdeli et al. (2020) and Pardiñas et al. (2018) respectively showed two (rs34753377 and rs6500603) and three (rs17465671, rs12447542 and rs2161711) single-nucleotide polymorphisms (SNPs), located on chromosome 16 that were associated with SCZ.
C-Maf-inducing protein (CMIP) is an important gene located on 16q23 that is mainly expressed in human brains, encodes an 86-kDa protein 7-9, and plays a role in the T-cell signaling pathway (Liu et al., 2015). CMIP contributes to several biological pathways and is involved in various diseases such as glioma, gastric cancer, kidney disease, and dyslipidemia (Li et al., 2019; Mo et al., 2018; Wang & Wu, 2017; Zhang et al., 2017), as well as major depressive disorder, syndromic autism spectrum disorders, and specific language impairments (Eicher & Gruen, 2015; Gedik, 2017; Luo et al., 2017; Wang et al., 2015). However, no studies have documented the relationship between CMIP and SCZ.
Based on chromosome 16’s biological function and previous studies on CMIP, we hypothesized that CMIP may have a relationship with SCZ. Additionally, gender-specific associations between gene SNPs (i.e., RELN, GABRB3 and MTHFR) and SCZ have been found in several other studies (Sozuguzel & Sazci, 2019; Wan et al., 2019; Liu et al., 2018). We conducted a genetic association study stratified by gender to examine the association between tag SNPs of the CMIP gene and SCZ in the Han Chinese population.
Materials & Methods
Study sample
A total of 761 SCZ patients and 775 healthy controls without any personal or family history of mental illness were enrolled in this study. More details of the data collection are described in a previous paper (Fu et al., 2020). All subjects were recruited after providing written informed consent. The study was performed in accordance with the protocols approved by the Ethics Committee of Jilin University, China (2014-05-01).
SNP analysis
We searched for tag SNPs of CMIP using the Haploview program (http://hapmap.ncbi.nlm.nih.gov/). We found a total of 235 tag SNPs and selected four tag SNPs (rs12925980, rs2287112, rs3751859 and rs77700579) that were associated with some neuropsychiatric disorders in order to determine the associations with SCZ. We searched for minor allele frequencies (MAF) for each SNP across 1,000 genomes. The four SNPs’ MAF threshold was set above 0.05 for the Chinese Han population (CHB).
Genomic DNA was extracted from five mL of peripheral blood collected from each subject using a commercial DNA extraction kit (Kangwei Biotech Company, Beijing, China) according to the manufacturer’s instructions. SNP genotyping was performed using matrix-assisted laser desorption/ionization time of flight mass spectrometry (MALDI-TOF-MS). The forward and reverse primers for these four SNP amplifications are listed in Table 1.
| SNP | Primer sequence (5′–3′) | |
|---|---|---|
| Forward | Reverse | |
| rs2287112 | ACGTTGGATGATCAGCAAGAGCCTCAAACC | ACGTTGGATGTGGTTGCTGGTCTGCTTTTC |
| rs77700579 | ACGTTGGATGAGGATAGTGAGCACTTACCC | ACGTTGGATGGACAATGACAGCACCACCTC |
| rs3751859 | ACGTTGGATGTTTCCACCAGTGCTCAGGG | ACGTTGGATGGTTCTCCAGGTTCAAATGTC |
| rs12925980 | ACGTTGGATGCCCTTCCCCCATTGATACTC | ACGTTGGATGCACTAACTTCTTCAGCCCTC |
Statistical analysis
We compared the demographic variables and allele and genotype distributions between patients and controls using Pearson’s chi-square (χ2) test and Student’s t-test. Multiple logistic regression was used to test the association between SCZ and alleles or genotypes. IBM SPSS (version 24.0) was used for the statistical analyses mentioned above and R software (version 3.2.3) was used for type I error correction using the false discovery rate (FDR) method. In both case and control groups, we used the goodness of fit χ2 test to test the Hardy–Weinberg equilibrium (HWE) by online software SNPStats (https://www.snpstats.net/snpstats/start.htm). Haploview 4.2 and SNPStats were then used for linkage disequilibrium (LD) and haplotype analysis. Finally, we used Quanto 1.2.4 software to calculate the statistical power for each SNP according to the MAF (rs12925980: 0.495, rs2287112: 0.175, rs3751859: 0.369 and rs77700579: 0.131). SCZ prevalence was presupposed to be 1% according to previous studies. All tests were two-sided and a Padj-value less than 0.05 was considered statistically significant.
Results
Demographic characteristics
The case group consisted of 761 SCZ patients (58.2% males, mean age = 34.61 ± 12.02 years) and the control group consisted of 775 healthy people (56.2% males, mean age = 34.74 ± 11.41 years). Cases and controls were matched by sex (χ2 = 0.681, P = 0.409) and age (t = 0.221, P = 0.825). All SNPs were in accordance with the HWE in the control group (Table 2).
| SNP | Case | Control | |||||||
|---|---|---|---|---|---|---|---|---|---|
| H0 | He | χ2 | P | H0 | He | χ2 | P | ||
| rs12925980 | 0.483 | 0.486 | 0.026 | 0.871 | 0.488 | 0.481 | 0.186 | 0.666 | |
| rs3751859 | 0.449 | 0.441 | 0.278 | 0.598 | 0.415 | 0.435 | 1.739 | 0.187 | |
| rs2287112 | 0.212 | 0.241 | 9.975 | 0.002 | 0.228 | 0.224 | 0.298 | 0.585 | |
| rs77700579 | 0.173 | 0.173 | 0.006 | 0.937 | 0.188 | 0.191 | 0.199 | 0.655 | |
Note:
Ho, observed heterozygosity; He, expected heterozygosity.
Allele and genotype distribution
rs12925980, rs3751859 and rs77700579 had 98% detection rates and rs2287112 had a 96% detection rate. Table 3 shows the genotypic and allelic distribution of the four SNPs and the associations with SCZ in the overall sample. The genotypic distribution of rs2287112 was found to be significantly different between SCZ patients and healthy controls (P = 0.016), but the difference did not survive the FDR correction adjusted for the multiple comparison (Padj = 0.128). The similar distribution differences and associations were observed in the female group (Table 4). The allelic distribution was significantly different between females in the patient and control groups (Padj = 0.008). The GG genotype (OR = 4.445, 95% CI [1.227–16.105]) and G allele (OR = 4.445, 95% CI [1.788–11.046]) were risk factors for SCZ. The statistical power for rs2281112 was 0.949. More details are shown in Table 4. Tables 3 and 4 show the associations based on the recessive genetic model, and the results of other genetic models are listed in Tables S1–S4.
| SNPs | Genotype | Case (n) | Control (n) | χ2 | P | Padj | OR (95 CI) |
|---|---|---|---|---|---|---|---|
| rs77700579 | AA+TA | 733 | 762 | 0.475 | 0.491 | 0.66 | 1 |
| TT | 7 | 10 | 0.710 [0.268–1.879] | ||||
| Allele | |||||||
| T | 1338 | 1379 | 0.988 | 0.32 | 0.66 | 1 | |
| A | 142 | 165 | 0.887 [0.700–1.124] | ||||
| rs12925980 | TT+CT | 479 | 499 | 0.193 | 0.66 | 0.66 | 1 |
| CC | 250 | 273 | 0.953 [0.771–1.179] | ||||
| Allele | |||||||
| T | 606 | 621 | 0.774 | 0.379 | 0.66 | 1 | |
| C | 852 | 932 | 0.937 [0.810–1.083] | ||||
| rs3751859 | GG+GA | 653 | 685 | 0.387 | 0.534 | 0.66 | 1 |
| AA | 75 | 87 | 0.901 [0.650–1.250] | ||||
| Allele | |||||||
| G | 979 | 1050 | 0.201 | 0.654 | 0.66 | 1 | |
| A | 477 | 494 | 1.036 [0.889–1.207] | ||||
| rs2287112 | TT+GT | 682 | 761 | 5.754 | 0.016* | 0.128 | 1 |
| GG | 24 | 11 | 2.419 [1.175–4.977] | ||||
| Allele | |||||||
| T | 1214 | 1346 | 0.914 | 0.339 | 0.66 | 1 | |
| G | 198 | 198 | 1.109 [0.897–1.370] |
Notes:
Padj represent P corrected by FDR.
OR, is abbreviation of Odds ratio; 95%CI is abbreviation of 95% confidence interval.
| SNPs | Genotype | Female | Male | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Case | Control | χ2 | P | Padj | OR (95% CI) | Case | Control | χ2 | P | Padj | OR (95% CI) | ||
| rs3751859 | G/G–G/A | 267 | 302 (89.1%) | 0.111 | 0.784 | 0.824 | 1 | 386 (89.6%) | 383 | 0.27 | 0.578 | 0.845 | 1 |
| (89.90%) | (88.50%) | ||||||||||||
| A/A | 30 | 37 | 0.931 [0.559–1.551] | 45 (10.4%) | 50 | 0.886 [0.577–1.358] | |||||||
| (10.10%) | (10.90%) | (11.60%) | |||||||||||
| Aelle | |||||||||||||
| G | 393 | 463 | 0.686 | 0.427 | 0.824 | 1 | 586(68%) | 587 | 0 | 0.87 | 0.87 | 1 | |
| (66%) | (68%) | (68%) | |||||||||||
| A | 201 | 215 | 1.100 [0.869–1.391] | 276 | 279 | 0.983 [0.803–1.204] | |||||||
| (34%) | (32%) | (32%) | (32%) | ||||||||||
| rs77700579 | A/A-T/A | 305 | 337 (99.4%) | 0.241 | 0.659 | 0.824 | 1 | 428 (98.6%) | 425 | 0.295 | 0.634 | 0.845 | 1 |
| (99.70%) | (98.20%) | ||||||||||||
| T/T | 1 | 2 | 1.718 [0.155–19.067] | 6 | 8 | 1.297 [0.445–3.784] | |||||||
| (0.30%) | (0.60%) | (1.40%) | (1.80%) | ||||||||||
| Aelle | |||||||||||||
| A | 556 | 613 | 0.072 | 0.824 | 0.824 | 1 | 782 | 766 | 1.217 | 0.271 | 0.733 | 1 | |
| (91% | (90%) | (90%) | (88%) | ||||||||||
| T | 56 | 65 | 1.044 [0.717–1.520] | 86 | 100 | 1.187 [0.875–1.611] | |||||||
| (9%) | (10%) | (10%) | (12%) | ||||||||||
| rs12925980 | C/C–C/T | 248 (82.9%) | 283 (83.5%) | 0.194 | 0.635 | 0.824 | 1 | 354 (82.3%) | 367 | 0.044 | 0.847 | 0.87 | 1 |
| (84.80%) | |||||||||||||
| T/T | 51 (17.1%) | 56 (16.5%) | 1.083 [0.780–1.504] | 76 (17.7%) | 66 | 1.028 [0.777–1.360] | |||||||
| (15.20%) | |||||||||||||
| Aelle | |||||||||||||
| C | 402 | 348 | 0.158 | 0.691 | 0.824 | 1 | 726(87%) | 760 | 0.434 | 0.515 | 0.845 | 1 | |
| (59%) | (58%) | (88%) | |||||||||||
| T | 276 | 250 | 1.051 [0.840–1.314] | 112 (13%) | 108 | 1.066 [0.879–1.292] | |||||||
| (41%) | (42%) | (12%) | |||||||||||
| rs2287112 | T/T–G/T | 276 (96.2%) | 335 (99.1%) | 6.148 | 0.023 | 0.092 | 1 | 406 (96.9%) | 426 (98.2%) | 1.408 | 0.275 | 1 | |
| G/G | 11 | 3 | 4.445 [1.227–16.105] | 13 | 8 | 0.733 | 1.645 [0.674–4.019] | ||||||
| (3.80%) | (0.90%) | (3.10%) | (1.80%) | ||||||||||
| Aelle | |||||||||||||
| T | 552 | 670 | 12.296 | 0.001 | 0.008* | 1 | 812 | 852 | 2.816 | 0.122 | 0.733 | 1 | |
| (96.20%) | (99.10%) | (96.90%) | (98.20%) | ||||||||||
| G | 22 | 6 | 4.445 [1.788–11.046] | 26 | 16 | 1.645 [0.875–3.094] | |||||||
| (3.80%) | (0.90%) | (3.10%) | (1.80%) | ||||||||||
Notes:
Padj represent P corrected by FDR.
OR is abbreviation of Odds ratio, 95% CI is abbreviation of 95% confidence interval.
LD and haplotype analysis
As shown in Fig. 1, the R2 values were 19 across total subjects (A) and male (B) and female subjects (C). LD was not observed across these SNPs according to the criteria (R2 > 0.8). The four SNPs’ position relationship in CMIP according to the National Center for Biotechnology Information (NCBI, https://www.ncbi.nlm.nih.gov/) gene structure are shown in Fig. 1D. We conducted haplotype association analysis with SCZ across all participants because the LD analysis results were similar between the male and female groups. The haplotype analysis results (Table 4) indicated that the haplotype made of all four SNPs (rs12925980–rs2287112–rs3751859–rs77700579) had a significantly different distribution between SCZ patients and healthy controls (Padj = 0.018). Furthermore, we estimated nine common haplotypes with a frequency >1% in detail. The results showed that the haplotype TTGT was significantly associated with SCZ (OR = 0.42, 95% CI [0.19–0.94], P = 0.032), but when FDR-adjusted the P-value was greater than 0.05 (Table 5).
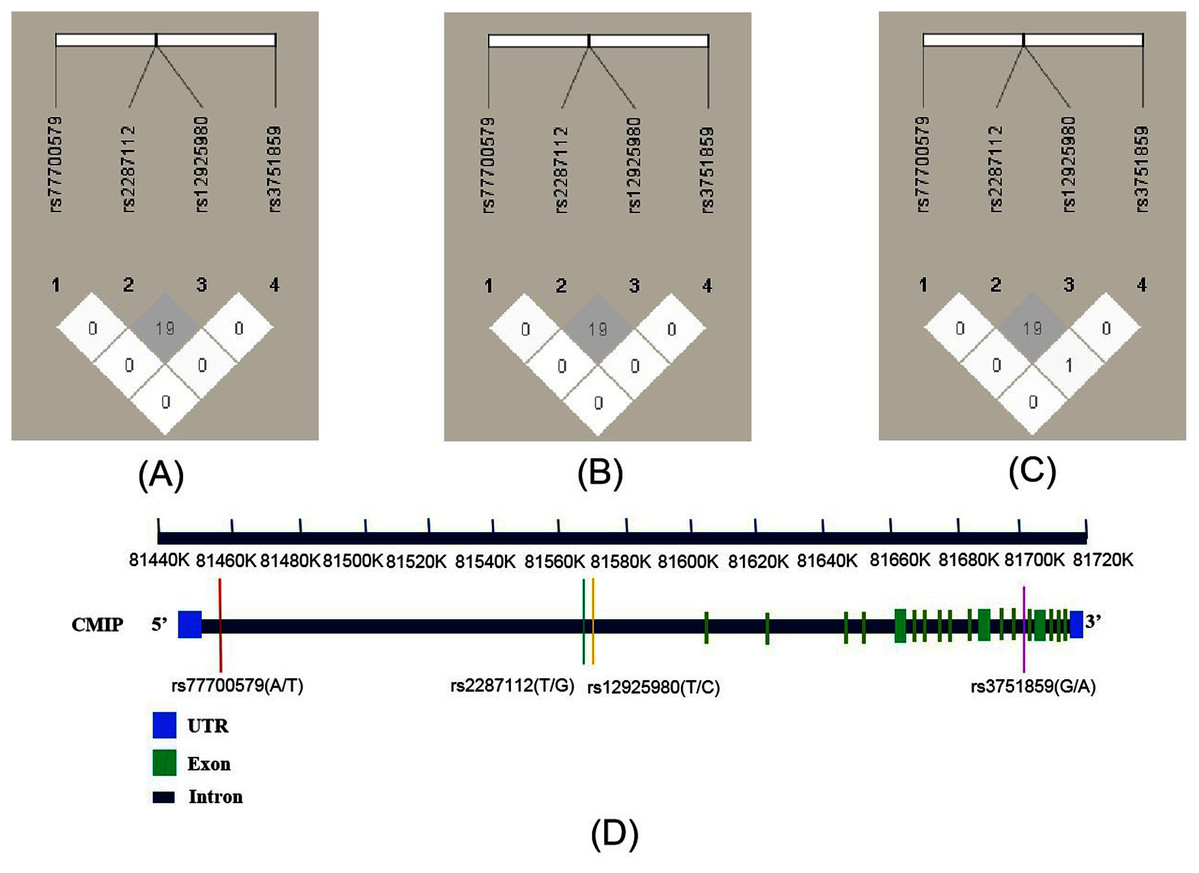
Figure 1: Linkage disequilibrium (LD) of four SNPs within CMIP in different subjects and the location of SNPs on CMIP gene structure.
R2 values were used to estimate the LD between pairwise SNPs. (A) LD of total subjects. (B) LD of male. (C) LD of female. (D) Location of four SNPs on CMIP gene.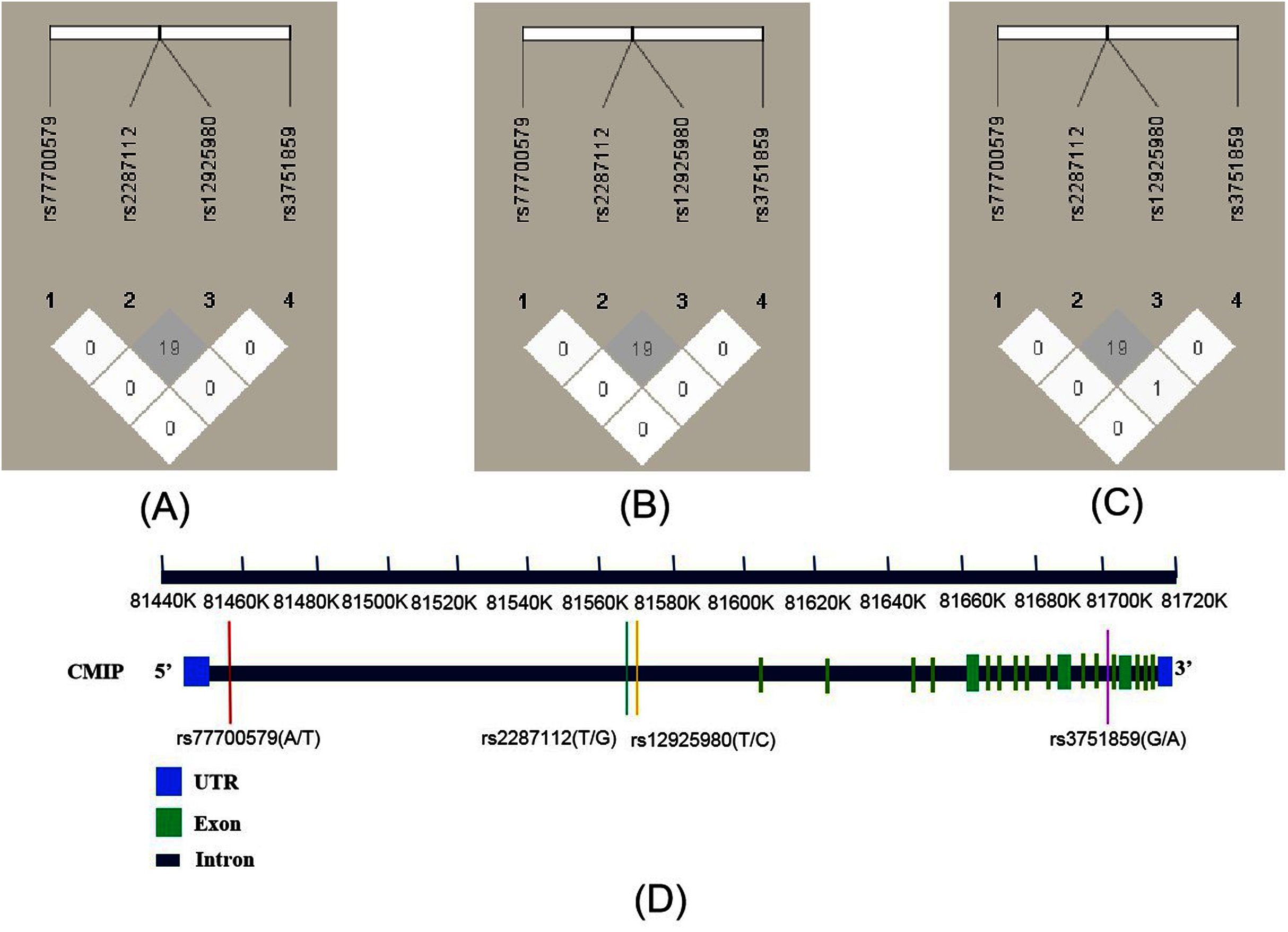{kind=link}
| rs12925980–rs2287711– | Frequency | OR (95%CI) | P | Padj | |||
|---|---|---|---|---|---|---|---|
| rs3751859–rs77700579 | Total | Control | Case | ||||
| CTGA | 0.3426 | 0.3548 | 0.329 | 1 | |||
| CTAA | 0.1782 | 0.1758 | 0.179 | 1.07 [0.83–1.38] | 0.620 | 0.620 | |
| TTGA | 0.1681 | 0.163 | 0.173 | 1.11 [0.85–1.44] | 0.440 | 0.620 | |
| TGGA | 0.0945 | 0.0908 | 0.099 | 1.10 [0.80–1.51] | 0.550 | 0.620 | |
| TTAA | 0.082 | 0.0769 | 0.09 | 1.26 [0.90–1.74] | 0.170 | 0.453 | |
| CTGT | 0.0402 | 0.0357 | 0.047 | 1.33 [0.82–2.17] | 0.250 | 0.500 | |
| TGAA | 0.0279 | 0.0309 | 0.025 | 0.83 [0.46–1.51] | 0.540 | 0.620 | |
| CTAT | 0.0249 | 0.0307 | 0.019 | 0.60 [0.30–1.22] | 0.160 | 0.453 | |
| TTGT | 0.0234 | 0.0317 | 0.013 | 0.42 [0.19–0.94] | 0.032* | 0.272 | |
Notes:
OR is abbreviation of Odds ratio, 95% CI is abbreviation of 95% confidence interval.
Discussion
Many studies have investigated the association between the CMIP gene and diseases such as mental neuropsychiatric disorder (Eicher & Gruen, 2015; Luo et al., 2017; Wang et al., 2015), cancer (Juan et al., 2019), and metabolic disease (Cao, Wang & Wu, 2018; Mo et al., 2018). In this study, we included 1,536 participants to study the association between four tag SNPs (rs12925980, rs22287112, rs3751859 and rs77700579) of the CMIP gene and SCZ. To the best of our knowledge, our study is the first of its kind to explore the correlation between CMIP and SCZ in the northeast CHB. We found that one loci (rs2287112) was associated with SCZ in females, indicating that CMIP was a potential risk genetic variant for SCZ. A large scale GWAS study conducted by Gedik (2017) found that the SNP rs77700579 in CMIP was associated with major depressive disorder (MDD), supporting the conclusion that CMIP was a potential candidate gene for neuropsychiatric disorders.
Several studies have detected sex-distinct gene polymorphisms with SCZ, including LTA, TNFA, IFNGR2 and PLA2G12A (Inoubli et al., 2018; Jemli et al., 2017; Yang et al., 2016). Yu et al. (2013) found eight genes with differential expression in female and male SCZ patients. Our research group also found a sex-specific SNP of gene RELN with SCZ in a previous study (Bai et al., 2019). Considering that SCZ’s sex-specific molecular phenotype has been observed in previous studies, we first explored the association between CMIP and SCZ in all samples and then separately tested the association for the male and female subgroups. We found that the SNP rs2287112 was significantly associated with SCZ in the whole group and female subgroup with a statistically significant value of 0.05. However, in the whole group the P value did not withstand FDR correction. The association between rs2287112 and SCZ only existed after P value correction in the female group. The association was not observed in the male group, providing more evidence that the molecular phenotype in SCZ is sex-specific. It should be noted that rs2287112 was not in HWE in the SCZ group, which suggested population stratification. The population structure evaluation showed no stratification and the control group conformed to HWE, ruling out the possibility of population admixture. The deviation from HWE may have been caused by the association with the disease that exerted a strong selection on the genome (Li et al., 2011).
Additionally, we carried out haplotype analysis to determine the association between the haplotype and SCZ and whether the combination of specific alleles could affect SCZ susceptibility. The TTGT haplotype (rs12925980–rs2287112–rs3751859–rs77700579) correlated with a lower risk of SCZ in our study population, but the association did not survive FDR correction. Similarly, the haplotype consisting of rs12929303–rs2287112–rs12925980 in CMIP was associated with developmental dyslexia in a Chinese population (Wang et al., 2015), suggesting that the haplotype including rs22287112 may contribute to disease susceptibility. The haplotype analysis further supported that rs2287112 allele G correlated with an increased SCZ risk.
Since this was a cross-sectional study, several limitations should be mentioned. First, this study was limited to interpreting the causal relationship between genetic risk factors and SCZ. Second, we only analyzed four SNPs in this study and may have missed some other loci associated with SCZ. Additionally, owing to the failure of demographic characteristic and in-depth clinical trait collection, we were not able to analyze the association of these SNPs with different SCZ clinical features. We were also limited to interaction analysis between genes and environment. Further studies that incorporate a large-scale sample size with more demographic characteristic information are warranted to further substantiate the association between CMIP gene polymorphism and SCZ susceptibility.
Conclusion
This study presented evidence that a CMIP variant is associated with SCZ susceptibility in northeast Han Chinese women. Considering the limitations of our work, additional functional genomics studies are required to further explain the role of SCZ-associated CMIP variants.
Supplemental Information
Raw data of this study.
Sex: 1 = male, 2 = female; Group: 1 = case, 0 = control