
The multi metal-resistant bacterium Cupriavidus metallidurans CH34 affects growth and metal mobilization in Arabidopsis thaliana plants exposed to copper
- Published
- Accepted
- Received
- Academic Editor
- Todd Anderson
- Subject Areas
- Agricultural Science, Microbiology, Plant Science, Environmental Contamination and Remediation
- Keywords
- Arabidopsis, Cupriavidus, Copper, Metals
- Copyright
- © 2021 Clavero-León et al.
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2021. The multi metal-resistant bacterium Cupriavidus metallidurans CH34 affects growth and metal mobilization in Arabidopsis thaliana plants exposed to copper. PeerJ 9:e11373 https://doi.org/10.7717/peerj.11373
Abstract
Copper (Cu) is important for plant growth, but high concentrations can lead to detrimental effects such as primary root length inhibition, vegetative tissue chlorosis, and even plant death. The interaction between plant-soil microbiota and roots can potentially affect metal mobility and availability, and, therefore, overall plant metal concentration. Cupriavidus metallidurans CH34 is a multi metal-resistant bacterial model that alters metal mobility and bioavailability through ion pumping, metal complexation, and reduction processes. The interactions between strain CH34 and plants may affect the growth, metal uptake, and translocation of Arabidopsis thaliana plants that are exposed to or not exposed to Cu. In this study, we looked also at the specific gene expression changes in C. metallidurans when co-cultured with Cu-exposed A. thaliana. We found that A. thaliana’s rosette area, primary and secondary root growth, and dry weight were affected by strain CH34, and that beneficial or detrimental effects depended on Cu concentration. An increase in some plant growth parameters was observed at copper concentrations lower than 50 µM and significant detrimental effects were found at concentrations higher than 50 µM Cu. We also observed up to a 90% increase and 60% decrease in metal accumulation and mobilization in inoculated A. thaliana. In turn, copper-stressed A. thaliana altered C. metallidurans colonization, and cop genes that encoded copper resistance in strain CH34 were induced by the combination of A. thaliana and Cu. These results reveal the complexity of the plant-bacteria-metal triad and will contribute to our understanding of their applications in plant growth promotion, protection, and phytoremediation strategies.
Introduction
Copper (Cu), like many other elements, is an essential trace nutrient required for normal plant growth and development (Silver, 1998; Andresen, Peiter & Küpfer, 2018; Shabbir et al., 2020; Kumar et al., 2021). Required Cu levels in plants range between 5 and 30 mg kg−1, whereas Cu levels in agricultural soils normally range between 10 and 60 mg kg−1 (Ballabio et al., 2018). Unnatural soil copper sources include manure, sewage sludge, and mineral fertilizers and pesticides (Oorts, 2013), and copper accumulation and leaching are strongly dependent on soil type and conditions (Shabbir et al., 2020). High copper levels negatively affect plants and microorganisms thriving in soil environments (Rajput et al., 2020; Shabbir et al., 2020), leading to the development and expression of detoxification and tolerance processes (Giachino & Waldron, 2020; Kumar et al., 2021).
Because of its redox potential, Cu has the ability to cycle between oxidized Cu2+ and reduced Cu+, thus mediating electron transfer reactions as a cofactor in the active centers of several key enzymes. Cu+ reduction places Cu in crucial roles in environmental and physiological processes such as photosynthesis, respiration, reactive oxygen species detoxification, ethylene perception, and cell wall remodeling and lignification (Maksymiec, 1998; Burkhead et al., 2009; Schmidt, Eisenhut & Schneider, 2020). These processes have been widely studied in the plant model Arabidopsis thaliana (Lequeux et al., 2010; De Smet et al., 2015; Niu et al., 2019). Cu deficiency and excess cause imbalances in normal plant development and function (Pätsikkä et al., 2012; Demirevska-Kepova et al., 2004). The most common symptoms of Cu toxicity in plants are primary root length inhibition, vegetative tissue chlorosis, and the imbalanced capture of other relevant trace elements such as Fe, Mn, and Zn (Lequeux et al., 2010; Andrés-Colás et al., 2013).
In order to fully understand the homeostasis of Cu and other transition metals in plants, the role of plant-associated microorganisms must be considered (González-Guerrero et al., 2016; Wasai & Minamisawa, 2018). Soil microorganisms affect trace metal speciation, mobility, and availability in the soil matrix. Processes such as chemical transformation, chelation, and protonation increase the mobilization of trace elements, while precipitation and sorption decrease trace element availability (Gadd, 2004; Mohamad et al., 2012; Rajkumar et al., 2012; Sessitsch et al., 2013; Muehe et al., 2015). However, the role of metal-tolerant soil microorganisms in plant growth is still poorly understood, and additional studies using model microorganisms are clearly needed.
One suitable microorganism is Cupriavidus metallidurans strain CH34, a well known multi metal-resistant bacterial model that harbors at least 24 metal resistance gene clusters distributed acros four replicons: one chromosome (CHR1), one chromid (CHR2), and two megaplasmids (pMOL28 and pMOL30) (Janssen et al., 2010). This bacterium belongs to the Burkholderiaceae family, which is comprised of members that are capable of inhabiting diverse niches accross sludges, sediments, soils, rhizospheres, and plant roots (Pérez-Pantoja et al., 2012). Strain CH34 can withstand the millimolar range concentrations of heavy metal ions such as Cu+, Cu 2+, Ni2+, Zn2+, and Co2+, all of which are important for plant nutrition (Mergeay et al., 2008; Kirsten et al., 2011). Metal tolerance is mainly achieved by ion efflux, but also by metal complexation and reduction (Mergeay et al., 2003). The role of this strain as a potential plant growth promoting rhizobacterium (PGPR) deserves further study.
In this study, we describe, for the first time, the effects of this multi metal-resistant strain on plant growth parameters, and the uptake and mobilization of metals (including several trace metal plant nutrients) in A. thaliana individuals that are exposed and not exposed to Cu. We additionally explored strain CH34’s colonization of A. thaliana’s rhizosphere under these conditions and the transcriptional responses of a few specific genes of C. metallidurans CH34 in an A. thaliana-bacterium co-cultivation system.
Materials and Methods
Bacterial strains, growth conditions, and A. thaliana inoculation
C. metallidurans CH34 was obtained from the Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (Braunschweig, Germany). It was routinely grown in Dorn minimal saline medium (Dorn et al., 1974) containing 10 mM gluconate as a sole carbon and energy source in an orbital shaker (150 rpm) at 30 °C. We collected suspensions of cells that were fully grown to the mid-exponential phase (OD600 nm = 0.6), adjusted them to approximately 108 colony-forming units per milliliter (CFU mL−1), and diluted them in agar medium just prior to solidification in order to obtain 104 CFU mL−1 for A. thaliana inoculation of a gnotobiotic system. CFU were calculated based on colony counts from serial one mL dilutions (usually 10−2 to 10−8) of 100-µl aliquots plated on three R2A agar plate replicates. Ultimately, the CFU values corresponded to the average of colony counts x corresponding dilution × 10 (100 aliquot/1,000 uL dilution volume). To assess the effect of heat-inactivated bacteria, we heated an inoculum suspension at 95 °C for 20 min prior to the final dilution in agar medium. This temperature was high enough to kill all bacterial cells without destroying them (Poupin et al., 2013). Bacterial cell viability was routinely confirmed using R2A medium agar plate counting.
In vitro A. thaliana-bacteria co-cultivation assays
A. thaliana Col-0 seeds were obtained from the Arabidopsis Biological Resource Center (Ohio State University, Columbus, OH, USA). The seeds were surface sterilized with 95% ethanol commercial laundry bleach (50%), rinsed three times with sterile water, and kept at 4 °C for 4 days to synchronize germination. Sterility was demonstrated in R2A or Luria Bertani media agar plates inoculated with aliquots of the solution used to wash seeds. Square Petri dishes were prepared with half-strength Murashige-Skoog medium (MS1/2) (Murashige & Skoog, 1962), 0.8% agar, and 1.5% sucrose (a carbon source for the plant, not for strain CH34); were either inoculated or not inoculated with strain CH34; and were supplemented with 0, 25, 50, 60, and 70 µM CuSO4x5H2O. The Petri dishes were then vertically placed in a growth chamber and incubated at 20−22 °C under a 16:8 h (long day) light:dark cycle. It should be noted that the copper concentration in the MS medium was 0.1 µM. We measured the A. thaliana growth parameters 21 days after sowing (DAS) to ensure that the parameters and bacterial colonization tests were accurate (Poupin et al., 2013; Zúñiga et al., 2017). The rosette areas were calculated using Adobe® Photoshop® Cs3 software (Adobe® Systems Incorporated, San Jose, CA, USA). We measured the primary root growth using a ruler and counted the secondary roots. Fresh weight (FW) and dry weight (DW) were determined using a Shimadzu analytical balance (Shimadzu Corporation, Kyoto, Japan).
Rhizosphere and A. thaliana colonization
To quantify bacterial colonization, we removed A. thaliana individuals at 21 DAS from inoculated agar that was either supplemented or not supplemented with 50 µM CuSO4x5H2O, and used vortex agitation to wash roots in a 10 mM MgSO4x7H2O solution in order to release their attached bacteria cells. The copper concentration was chosen based on its relation with positive or negative effects on A. thaliana. The extracted liquid was serially diluted with the same solution before plating on R2A agar plates, and the CFU mg−1 FW values were determined after 24 h of incubation at 30 °C. To quantify rhizosphere colonization, we used vortex agitation to wash the roots in a 10 mM MgSO4x7H2O solution in order to release the samples of agar (MS1/2medium) that were in close contact with the roots (rhizosphere agar at a distance of less than one cm) and attached bacteria cells. We used A. thaliana plants that were grown without strain CH34 inoculation to check the sterility of the conditions. Sterility was proved after inoculation of aliquots from the solution used to wash samples of the agar material attached to roots, in rich media agar plates as indicated above. Agar plates without A. thaliana, and either supplemented with or without 50 µM CuSO4x5H2O, were used to determine A. thaliana’s influence on bacterial growth. Agar material samples were also washed with a 10 mM MgSO4x7H2O solution.
Metal quantification in A. thaliana tissues
For total metal quantification, we separated the rosettes and roots of A. thaliana individuals 21 DAS, cultivated in the presence of 50 µM CuSO4x5H2O, and either inoculated or not inoculated with strain CH34, and rinsed them six times with ultra pure water. After drying for 24 h at 50 °C, the rosettes and roots material was separated into three 8 mg DW groups and digested using a HNO3:H2O2 = 2:1 mixture in a high-performance microwave (Milestone, Ethos One, Brondby, Denmark). Metal analysis was performed using an ICP-MS Thermo Scientific X Series 2 spectrometer (ThermoFisher Scientific, Waltham, MA, USA). We calculated the translocation factor (TF) for each metal as the ratio between the rosette metal g−1 µg and the root metal g−1 µg, for each condition. Metal contents were determined by multiplying each sample concentration by the DW.
Bacterial gene expression tests in A. thaliana-bacterium co-cultivation assays
To evaluate C. metallidurans CH34 gene expression in the presence of A. thaliana, we performed co-cultivation assays in hydroponic cultures. In order to allow adequate A. thaliana growth, 60 previously sterilized seeds were sown in the sterile conditions described above: MS (100%) medium supplemented with 3% sucrose, and 0 or 25 µM CuSO4x5H2O. This Cu concentration was chosen again because no detrimental effects were observed in the other A. thaliana individuals. A. thaliana growth was carried out under the same conditions as the in vitro cultures. At 21 DAS, the hydroponic cultures were inoculated with 1 × 104 cells/mL of strain CH34. We interrupted the A. thaliana-bacterium interaction using quenching buffer (methanol 60%, 62.5 mM HEPES, and distilled water) after 30, 60, and 180 min of inoculation, and stored the biological material at −80 °C. These time periods were chosen because A. thaliana already influences bacterial gene expression (Zúñiga et al., 2013; Zúñiga et al., 2017). We used the same hydroponic systems that were inoculated with strain CH34 without A. thaliana, along with a growth medium containing gluconate supplemented with 0 or 25 µM CuSO4x5H2O, as the control to verify the effects on the induction and repression of bacterial genes in the absence of A. thaliana or Cu.
Quantitative real time polymerase chain reaction (qRT-PCR) analysis
To extract RNA, we processed 4 ml of each co-cultivation assay using the geneJET RNA purification kit (ThermoFisher Scientific, Waltham, MA, USA). The RNA was quantified using an EON™ microplate reader (BioTek®, Winoosky, VT, USA) and treated with the TURBO DNase kit (Ambion, Austin, TX, USA) to remove DNA contamination. cDNA synthesis was performed using the ImProm-II™ Reverse Transcription System (Promega Corporation, Madison, WI, USA) with 1 µg of RNA in 20 µL reactions. RT-PCR was performed using the Brilliant SYBR® Green QPCR Master Reagent kit (Agilent Technologies, Santa Clara, CA, USA) and the Eco™ Real-Time PCR detection system (Illumina®, San Diego, CA, USA). The PCR mixture (10 µL) contained 2 µL of template cDNA (diluted 1:10) and 100 nM of each primer. Amplification was performed under the following conditions: 95 °C for 10 min; followed by 40 cycles of 94 °C for 30 s, 60 °C for 30 s, and 72 °C for 30 s; followed by a melting cycle from 55° to 95 °C. Relative gene expression calculation was conducted as described in the software manufacturer’s instructions (Illumina®, San Diego, CA, USA). We calculated an accurate ratio between the expression of the gene of interest (GOI) and the housekeeping (HK) gene (16S rRNA) according to 2–(ΔCtGOI-HK). Firstly, we normalized using a ratio between the GOI and the 16S gene (as HK). Then, we used the time 0 treatment (before any treatment with MS or plant exudates) as the calibrating condition. The previous allows us to compare each treatment data with the others. It should be mentioned that the use of 16S rRNA is only for preliminary and explorative purposes and subsequent studies should include more appropriate housekeeping genes. Gene expression levels were normalized to the average expression level values in the control treatment. We also used reported primer pairs for copK, copC, copC 2, and copF gene sequences (Table 1), which were involved in the copper resistance of strain CH34 (Monchy et al., 2006). Primer pairs for aleO, iucA, aleB, piuA, phaC1, catA, pcaG, bvgA, and phcA genes (Table 1), involved in potential bacterial-plant interactions, were designed using Primer 3 version 0.4.0 (http://primer3.ut.ee). All PCR determinations were performed using at least three biological replicates and two technical replicates per treatment.
| Gene | Locus tag | Forward primer | Reverse primer |
|---|---|---|---|
| aleO | Rmet_5746 | GGAAGACGTCTATCGGACCA | CTGTGGCGTAACAGGATGTC |
| iucA | Rmet_1115 | GCATGTCGTGGATTGATCTG | GTGCAAGGGATATGGCTCAG |
| aleB | Rmet_1118 | ACGAAGAAGGACACGGTCAC | GGGAGATGGCTTGTGTTGTT |
| piuA | Rmet_4617 | TCTCGACGATTTCACGAATG | GCGGTCTCGTCATTGATCTT |
| phaC1 | Rmet_1356 | ACCAATACGACGGCAGAGAG | ACTGAAACGGTTGGGACTTG |
| catA | Rmet_4881 | CGCTGGTAAAGCAGTTCCTC | TTGAACAGATCGGTCGTCAG |
| pcaG | Rmet_4014 | GTCGTTCCGCTTCGTTACC | GCATGAACACCGACACCAT |
| bvgA | Rmet_5714 | CCATTGGAGACTCGCTCTTC | GTCACGACCTTGACCTGCTT |
| phcA | Rmet_2977 | GATGCTGGGCCTGAATATGT | AGAAACTCGCCAACCACCT |
Statistical analysis
The statistical analyses of A. thaliana growth parameters were performed using the two-way analysis of variance (ANOVA). Tukey’s honest significant difference (P < 0.05) test was used to make comparisons across different treatments.
Results
C. metallidurans CH34 effects on A. thaliana growth with varied Cu concentrations
We carried out in vitro A. thaliana-bacterium co-cultivation assays using a MS1/2medium supplemented with 0 to 70 µM Cu2+ over a 21-day period in order to test the effects of strain CH34 on growth. These Cu2+ concentrations, particularly higher ones, caused mild to high stress in plants, reduced biomass, and increased chlorosis (Lequeux et al., 2010; Andrés-Colás et al., 2013). The presence of strain CH34 significantly altered A. thaliana’s rosette area, primary and secondary root growth, and FW and DW at all copper concentrations when compared with the non-inoculated group (Fig. 1A and 1B). When Cu concentrations were low to intermediate (0–50 µM), strain CH34 had a positive effect on A. thaliana, increasing its primary root length by 19.4%, 53.7%, and 19.0% in treatments with 0, 25, and 50 µM Cu2+, respectively. The number of secondary roots also increased by 26.4% at 25 µM Cu2+, and the rosette area increased by 10.3% and 21.0% at 0 and 25 µM Cu2+, respectively. These changes in plant growth parameters appear to be modest, at first instance, but truly reflect effects of the presence of a single bacterium population on the plant.
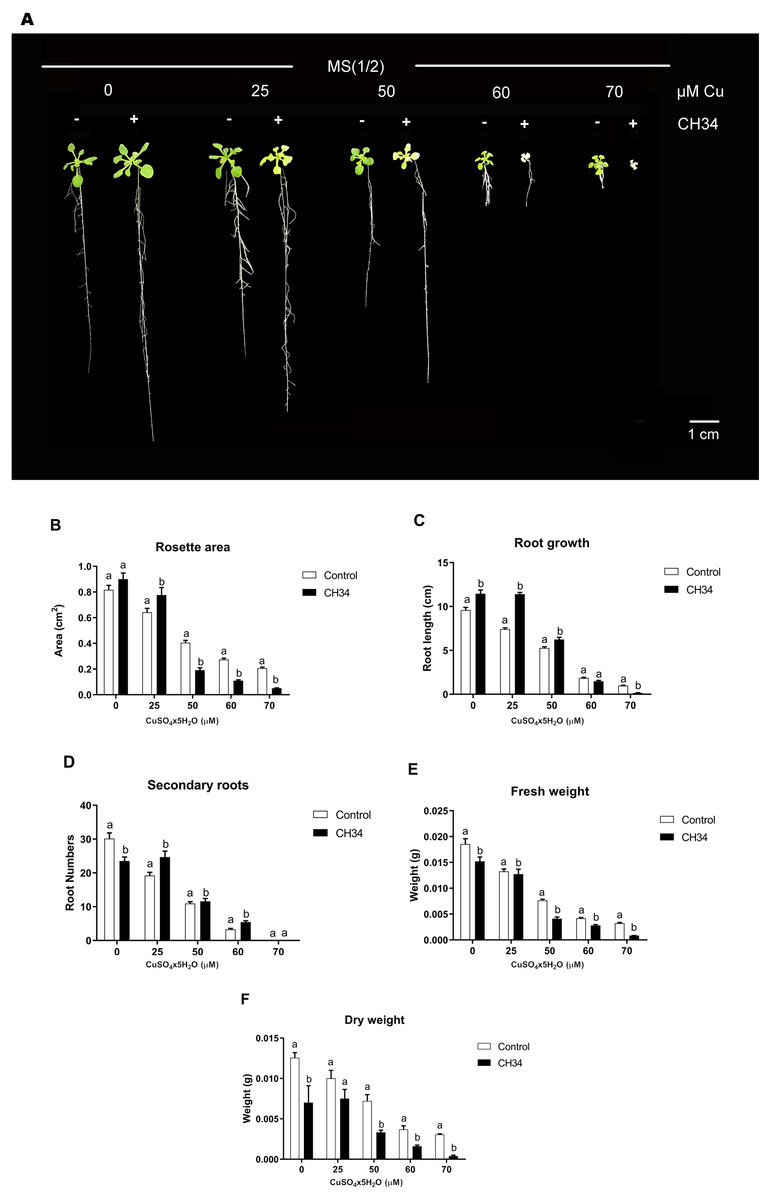
Figure 1: Effect of Cupriavidus metallidurans CH34 on growth parameters of Arabidopsis thaliana.
Rosette area, primary root growth, secondary roots and fresh and dry weights of A. thaliana Col-0 grown in in vitro gnotobiotic cultures using half strength Murashige-Skoog (MS) agar medium, inoculated with 1 × 104 colony forming units per milliliter of C. metallidurans CH34. Forty plants per condition were inoculated or not at day 1 and the half strength MS medium was supplemented with 0 to 70 µM CuSO4x5H2O. (A) Representative control and strain CH34 inoculated plants grown in the presence of different Cu concentrations. (B) Plant growth parameters registered after 21 days of sowing. Bars show mean percentage values, and error bars indicate standard deviations from experiments with 30 plants analyzed for each bacterial condition. Different letters indicate statistically significant differences among treatments at each copper concentration for each measured parameter (two way ANOVA Tukey’s HSD tests; p < 0.05).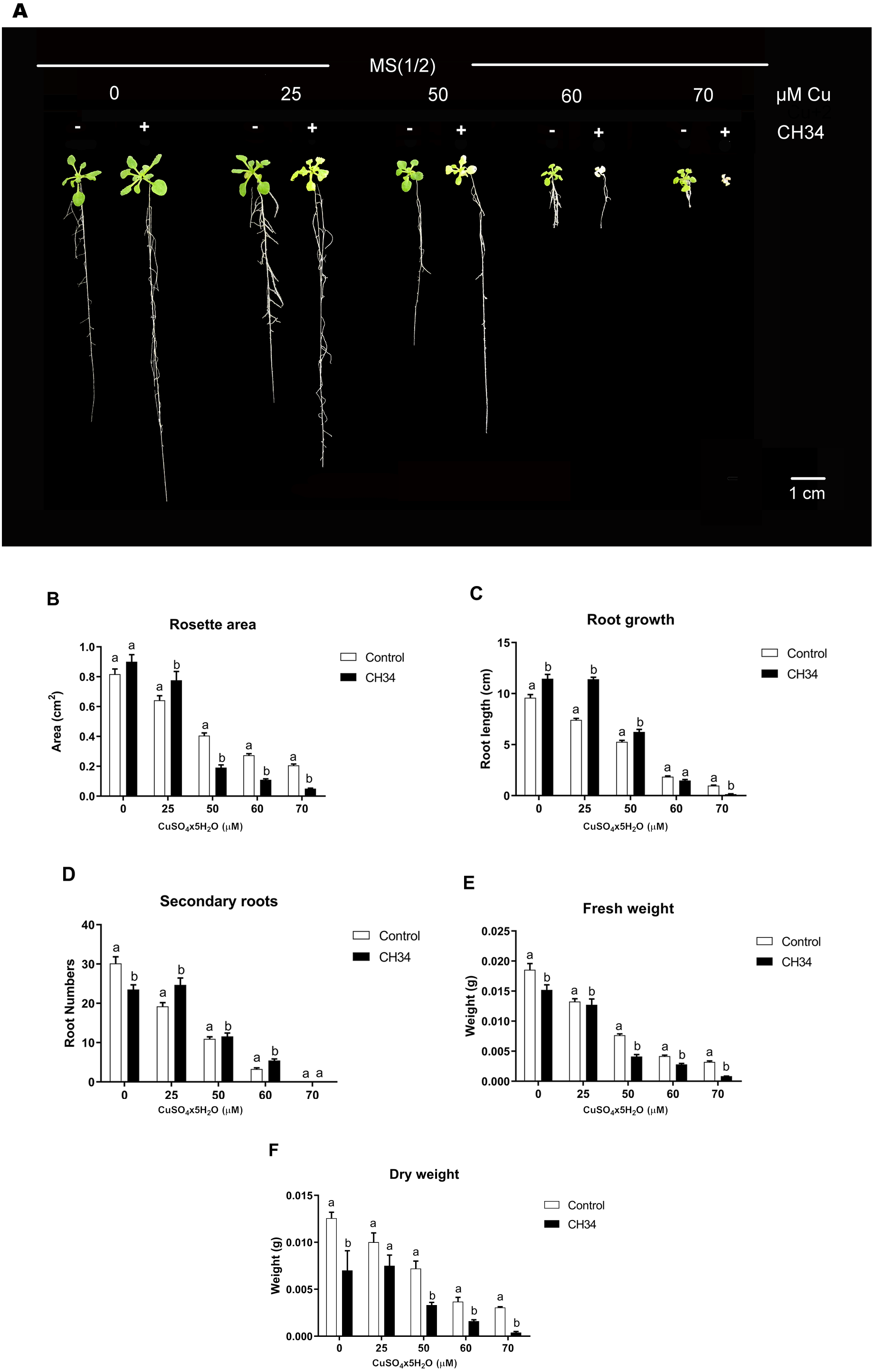{kind=link}
However, at intermediate to high Cu2+ concentrations (50–70 µM), strain CH34 had detrimental effects on primary root length, decreasing it by 20.0 and 86% in treatments with 60 and 70 µM, respectively, when compared with the non-inoculated control group. Rosette areas were reduced by 53.0, 60.0, and 75.3% at 50, 60, and 70 µM of Cu2+, respectively. FW and DW decreased in all inoculated treatments, except at 25 µM Cu2+ concentration, when compared to the non-inoculated condition (Fig. 1B). The decrease in FW and DW correlated with decreases in other A. thaliana growth parameters (Fig. 1B), except for root growth at 0 days, which might be explained by the initial reallocation of A. thaliana root resources upon bacterial inoculation. Both beneficial and detrimental effects were clearly caused by live CH34 cells, as the tests performed with heat-killed cells showed identical results as those of the non-inoculated control group (Supplementary Material, Fig. S1).
In control and Cu-exposed rosettes and roots, C. metallidurans CH34 modifies the accumulation and translocation of metals present in the A. thaliana growth medium
Cu may affect A. thaliana growth parameters (Fig. 1) via metal mobilization mediated by C. metallidurans (Nies, 2016). Therefore, we quantified the rosette and root tissue distribution of the main seven metals that composed the MS medium. Although not accurate, for simplicity we refer to boron as a metal. The metals and their concentrations were optimized for plant growth in the MS medium (Murashige & Skoog, 1962). However, when present in higher levels, these seven metals are toxic plant growth inhibitors (Andresen, Peiter & Küpfer, 2018). In inoculated A. thaliana, the absence of Cu caused a significant increase in B (29 and 32%), Co (68 and 67%), Cu (11 and 18%), Fe (42 and 91%), and Mn (11 and 46%) levels in rosettes and roots, respectively. However, Mo levels decreased (61%) and Zn levels increased (38%) in roots (absolute and relative values in Tables 2 and 3, respectively). In inoculated A. thaliana, the presence of Cu2+ significantly increased Co (40 and 73%), Cu (55 and 38%), Mn (33 and 10%), Mo (60 and 64%), and Zn (21 and 71%) levels in rosettes and roots, respectively. B (30%) and Fe (9%) increased in rosettes only, Fe (31%) decreased in roots, and B levels in roots were not affected (Tables 2 and 3). In non-inoculated A. thaliana, 50 µM Cu2+ increased B (109 and 380%), Co (50 and 20%), Cu (315 and 493%), Fe (277 and 315%), Mn (33 and 59%), and Zn (54 and 30%), and decreased Mo (27% and 70%) levels in rosettes and roots, respectively (Tables 2 and 3). To evaluate the metal mobility of A. thaliana roots and rosettes, TF values were determined for each metal (Table 4). In non-inoculated A. thaliana, we found that Cu clearly altered metal mobility because in the absence of Cu2+, the TFs for Co (1.5-fold) and Mo (2.3-fold) increased, and the TFs for Cu (0.7-fold), Mn (0.8-fold), and B (0.4-fold) decreased. Strain CH34 innoculation in the absence of Cu changed the TFs of all elements except for B and Co, but in the presence of Cu, the strain modified all TFs except for Mo (Table 4). In the absence of strain CH34, Cu2+ drastically changed the metal accumulation in A. thaliana roots and rosettes, with much higher B, Cu, and Fe accumulation; mild Co, Mn, and Zn accumulation; and negative (less accumulation in the presence of 50 µM Cu2+) Mo accumulation (Table 3).
| Rosettes | Roots | |||||||
|---|---|---|---|---|---|---|---|---|
| MS1/2 | MS1/2 + 50 µM Cu2+ | MS1/2 | MS1/2 + 50 µM Cu2+ | |||||
| Control | +CH34 | Control | +CH34 | Control | +CH34 | Control | +CH34 | |
| B | 21.0 ± 1.6a** | 27.5 ± 2.2b | 44.0 ± 3.4c | 57.3 ± 5.3d | 14.6 ± 2.0a | 19.3 ± 0.9b | 70.1 ± 23.4c | 59.2 ± 23.9c |
| Co | 0.2 ± 0.0a | 0.3 ± 0.0b | 0.3 ± 0c | 0.5 ± 0.0d | 0.5 ± 0.1a | 0.9 ± 0.0b | 0.6 ± 0.1a | 1.0 ± 0.1b |
| Cu | 4.5 ± 0.0a | 4.7 ± 0.2b | 186.9 ± 6.1c | 289.0 ± 1.4d | 5.2 ± 0.1a | 6.2 ± 0.3b | 308.5 ± 13.8c | 426.2 ± 20d |
| Fe | 118.0 ± 5.4a | 167.9 ± 9.0b | 445.0 ± 34.4c | 484.1 ± 11.5d | 2671.6 ± 240.8a | 5091.6 ± 180.0b | 11098.0 ± 336.9c | 7636.7 ± 377.7d |
| Mn | 196.5 ± 9.7a | 217.7 ± 12.4b | 262.6 ± 15.4c | 350.4 ± 7.2d | 111.3 ± 9.0a | 163.0 ± 4.1b | 177.1 ± 9.7c | 195.2 ± 12.2d |
| Mo | 19.5 ± 1.4a | 18.4 ± 2.1a | 14.3 ± 0.4b | 23.0 ± 1.2c | 66.1 ± 7.3a | 25.8 ± 4.7b | 21.3 ± 4.1c | 35.0 ± 1.6b |
| Zn | 191.4 ± 17a | 191.8 ± 16.5a | 295.7 ± 15.1b | 359.2 ± 1.5c | 940.4 ± 56.2a | 1299.2 ± 55.9b | 1222.8 ± 46.7c | 2091.8 ± 107.8d |
Notes:
Standard Murashige-Skoog plant growth medium. MS1/2 + 50μM Cu2+ indicates same medium supplemented with 50 µM CuSO4x5H2O. Each condition had three technical replicates consisting of at least 8 mg of rosette or root material. Different letters represent significant differences between rosette or root levels of each metal (two-way ANOVA, p < 0.05; Tukey test, p < 0.05).
| Rosettes (CH34/control) | Roots (CH34/control) | Rosettes (Cu/control) | Roots (Cu/control) | |||
|---|---|---|---|---|---|---|
| Element | MS1/2 | MS1/2 + 50 µM Cu2+ | MS1/2 | MS1/2 + 50 µM Cu2+ | ||
| B | +29%** | +30% | +32% | – | +109% | +380% |
| Co | +68% | +40% | +67% | +73% | +50% | +20% |
| Cu | +11% | +55% | +18% | +38% | +315 | +493% |
| Fe | +42% | +9% | +91% | −31% | +277% | +315% |
| Mn | +11% | +33% | +46% | +10% | +33% | +59% |
| Mo | – | +60% | −61% | +64% | −27% | −69% |
| Zn | − | +21% | +38% | +71% | +54% | +30% |
Notes:
Not significant changes.
| MS1∕2 | MS1∕2 + 50 µM Cu2+ | |||
|---|---|---|---|---|
| TFControl | TFCH34 | TFControl | TFCH34 | |
| B | 1.6a | 1.3a | 0.7b | 1.1b |
| Co | 0.4a | 0.4a | 0.6b | 0.5a |
| Mo | 0.3a | 0.8b | 0.7b | 0.7b |
| Cu | 0.9a | 0.8b | 0.6c | 0.7d |
| Mn | 1.9a | 1.3b | 1.5c | 1.8d |
| Fe | 0.04a | 0.03b | 0.04a | 0.06c |
| Zn | 0.2a | 0.1b | 0.2c | 0.1d |
Notes:
TF was calculated as = (Metal concentrationRosette/Metal concentrationRoot). MS1/2 corresponds to half-diluted standard Murashige-Skoog plant growth medium. MS1∕2 + 50μM Cu2+ indicates same medium supplemented with 50 μM CuSO4x5H2O. Each condition had three technical replicates consisting of at least 8 mg of rosette or root material. Different letters represent significant differences between treatments (two-way ANOVA, p < 0.05; Tukey test, p < 0.05).
Copper-stressed A. thaliana changed C. metallidurans CH34 colonization and the transcriptional levels of some genes
To further explore Cu’s effects on the A. thaliana-C. metallidurans interaction, we conducted rhizospheric and colonization tests in the presence of 50 µM Cu2+. This concentration was chosen because it provoked intermediate effects on A. thaliana growth parameters. C. metallidurans effectively colonized (1 × 108 CFU mL−1) both A. thaliana and its rhizospheric surroundings. Adding copper produced significant changes on the bacterial colonization by both increasing bacteria directly adhered to A. thaliana roots by 3.1 times and decreasing colonization in its vicinity by 7.1 times (Table 5). It should be noted that we observed positive effects on strain CH34 colonization as a 4-log increase in CFU with respect to the non-rhizospheric conditions was recorded (Table 5).
| Treatment | Plant (CFU mg−1FW) | Rhizosphere (CFU mg−1FW) | Non-rhizosphere (CFU mg−1FW) |
|---|---|---|---|
| MS(1/2) | 5.8 × 108 ± 4.7 × 108a | 1.0 × 107 ± 8.5 × 106a | 1.7 × 104 ± 1.1 × 104a |
| MS(1/2) + 50 µM Cu2+ | 1.8 × 109 ± 7.5 × 107a | 1.4 × 106 ± 1.2 × 106b | 3.6 × 104± 1.3 × 104b |
Notes:
C. metallidurans CH34 CFU mg−1 FW were counted in 21 days after sowing plants, rhizosphere agar and non-rhizosphere MS1/2 agar, in 0 and 50 µM CuSO4x5H2O treatments. Plants were initially inoculated with 1.0 × 104 CFU ml−1. The average values of 15–20 replicates and their respective standard deviations are shown. FW corresponds to that of plant or agar material. Different letters in same column indicate statistically significant differences according to a t-test, p < 0.05.
To further understand the effects of strain CH34 on A. thaliana protection, colonization, and metal translocation, we carried out a transcriptional analysis of a small subset of genes. These experiments were performed in A. thaliana hydroponic cultures in order to fulfill the quality requirements for bacterial RNA extraction. Since Cu has a greater presence in hydroponic medium than in agar plates (Burkhead et al., 2009), we used a 25 µM Cu2+ concentration to emulate the 50 µM Cu2+ used in agar plate A. thaliana tests in order to provoke similar effects. At 21 days, C. metallidurans was inoculated and RNA was extracted after 30, 60, and 180 min from the hydroponic cultures. These bacteria—A. thaliana interaction times were selected based on a previous study that used a similar Burkholderiaceae bacterium (Zúñiga et al., 2017). Three groups of genes were chosen to perform a RT-qPCR analysis: a set of cop genes involved in Cu detoxification, a set of siderophore-related genes, and a set of colonization-related genes (Janssen et al., 2010). Figure 2 shows the relative expression levels of the genes associated with copper resistance: copK, copF, copC, and copC2. The first three genes are located in the megaplasmid pMOL30 in a cluster of 21 cop genes and they encode proteins involved in periplasmic (copK and copC) and cytoplasmic (copF) detoxification. copC2, located on a smaller cluster of cop genes (copD 2C 2B 2A 2R 2S 2), is strongly induced mainly at concentrations lower than 100 µM Cu2+ (Janssen et al., 2010). copK was clearly expressed 60 min after A. thaliana inoculation (Fig. 2), with induction still observable after 180 min in the presence, but not in the absence, of Cu2+. copC only showed a transient significant induction in the presence of Cu2+. copF gene expression levels remained essentially unchanged and copC2 expression levels noticeably decreased in the presence of A. thaliana (Fig. 2).
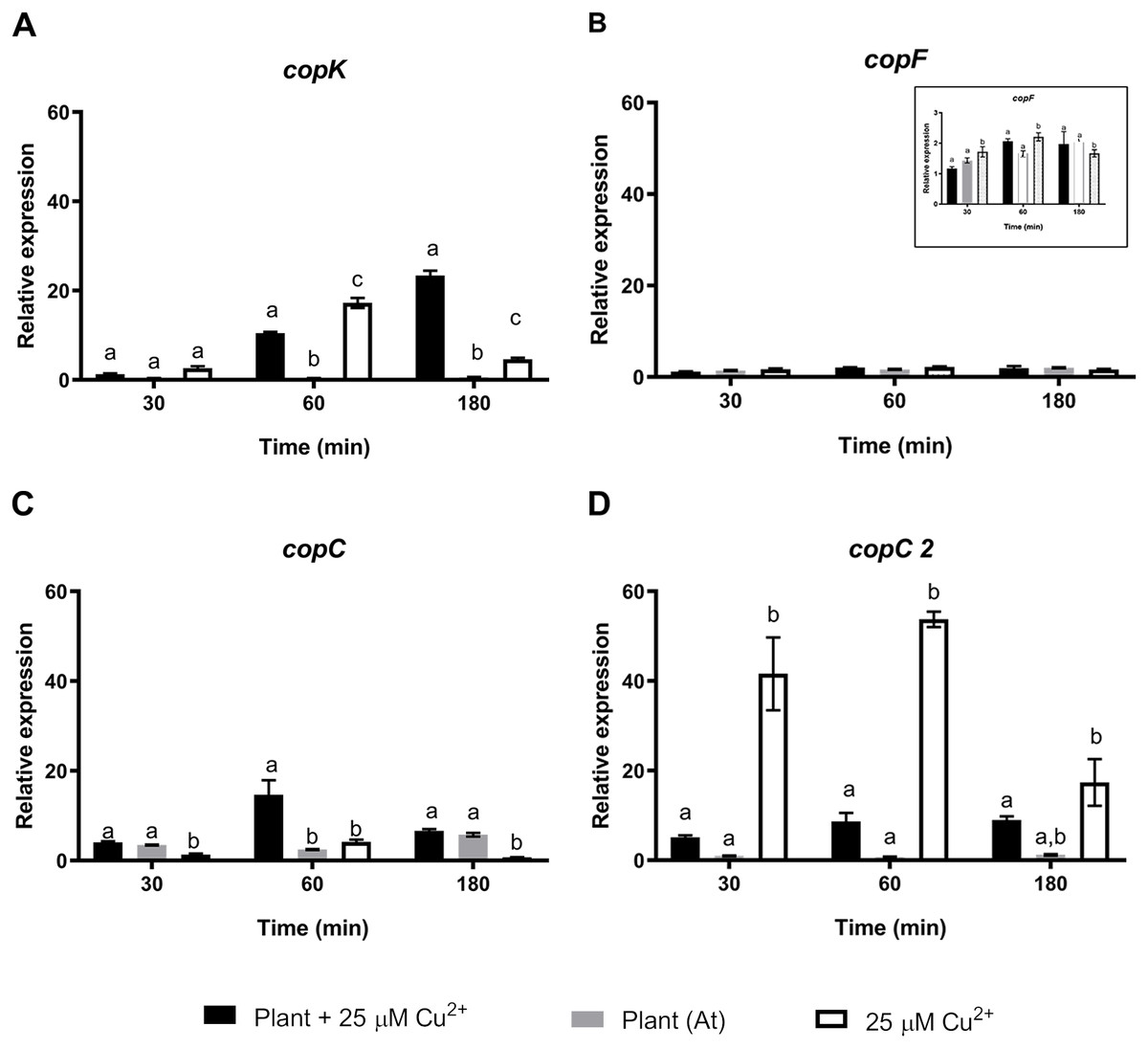
Figure 2: Relative expression levels of Cu resistance genes from Cupriavidus metallidurans CH34, in the presence or in the absence of Arabidopsis thaliana (At) or copper.
Quantitative Real Time Polymerase Chain Reactions determinations of relative expression levels of (A) copK, (B) copF, (C) copC, and (D) copC 2 genes. Expression levels were normalized respect to the housekeeping gene 16S rRNA. Data correspond to means ± standard deviations of at least three biological replicates. Different letters indicate significant differences between same time conditions (two way ANOVA Tukey’s HSD tests; p < 0.05).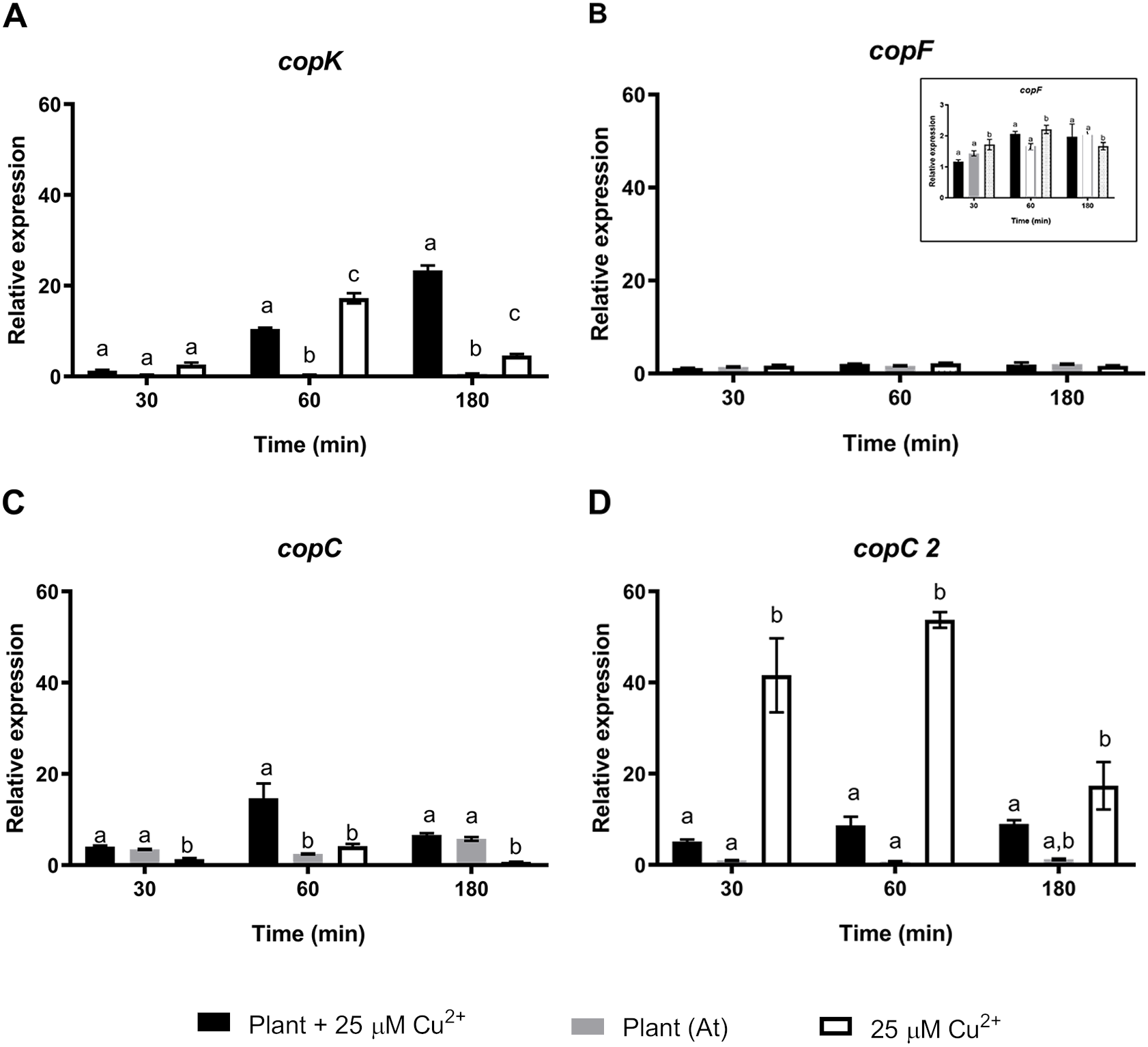{kind=link}
A second set of targeted genes were involved in the regulation, production, and sensing of C. metallidurans CH34 staphyloferrin B siderophore. A previous study found that C. metallidurans CH34 affects metal availability by producing a unique phenolate-type siderophore that has been described to bind Cd2+ (Gilis et al., 1998). The Fur protein (aleO gene) is a transcriptional regulator involved in Fe metabolism that represses staphyloferrin B synthesis under optimal Fe conditions, although it has been suggested that the transcriptional activation of other genes is also involved in Fe regulation (Ma, Jacobsen & Giedroc, 2009). After 30 and 180 min of C. metallidurans CH34 inoculation, Fur regulator expression was significantly repressed in both A. thaliana-only and A. thaliana plus 25 µM Cu2+-conditions, suggesting that Fe requirement should not be as severe as in 25 µM Cu2+-only conditions (Fig. 3). The iucA gene, one of the three genes responsible for staphyloferrin B synthesis, was only significantly induced in 25 µM Cu 2+-only conditions (Fig. 3), while the staphyloferrin B receptor aleB gene was induced after 60 and 180 min of inoculation in the presence of A. thaliana. The piuA gene encoding a siderophore/iron-complexation receptor was not involved in staphyloferrin B uptake, and was only induced in the 25 µM Cu2+-only condition (Fig. 3). Finally, we analyzed the expression changes in phaC1 (polyhydroxybutyrate synthesis), catA (catechol), pcaG (protocatechuate catabolism), bvgA (transcriptional activator of virulence and colonization factors), and phcA (virulence regulator) genes. phaC1 was the only gene that showed clear induction by A. thaliana (peaked at 30 min, Supplementary Material Fig. S2), as the other four tested genes were either not induced, or only poorly induced, by A. thaliana. Since the same medium was used across all tests, phaC1 gene’s changes could not be explained by phosphate changes.
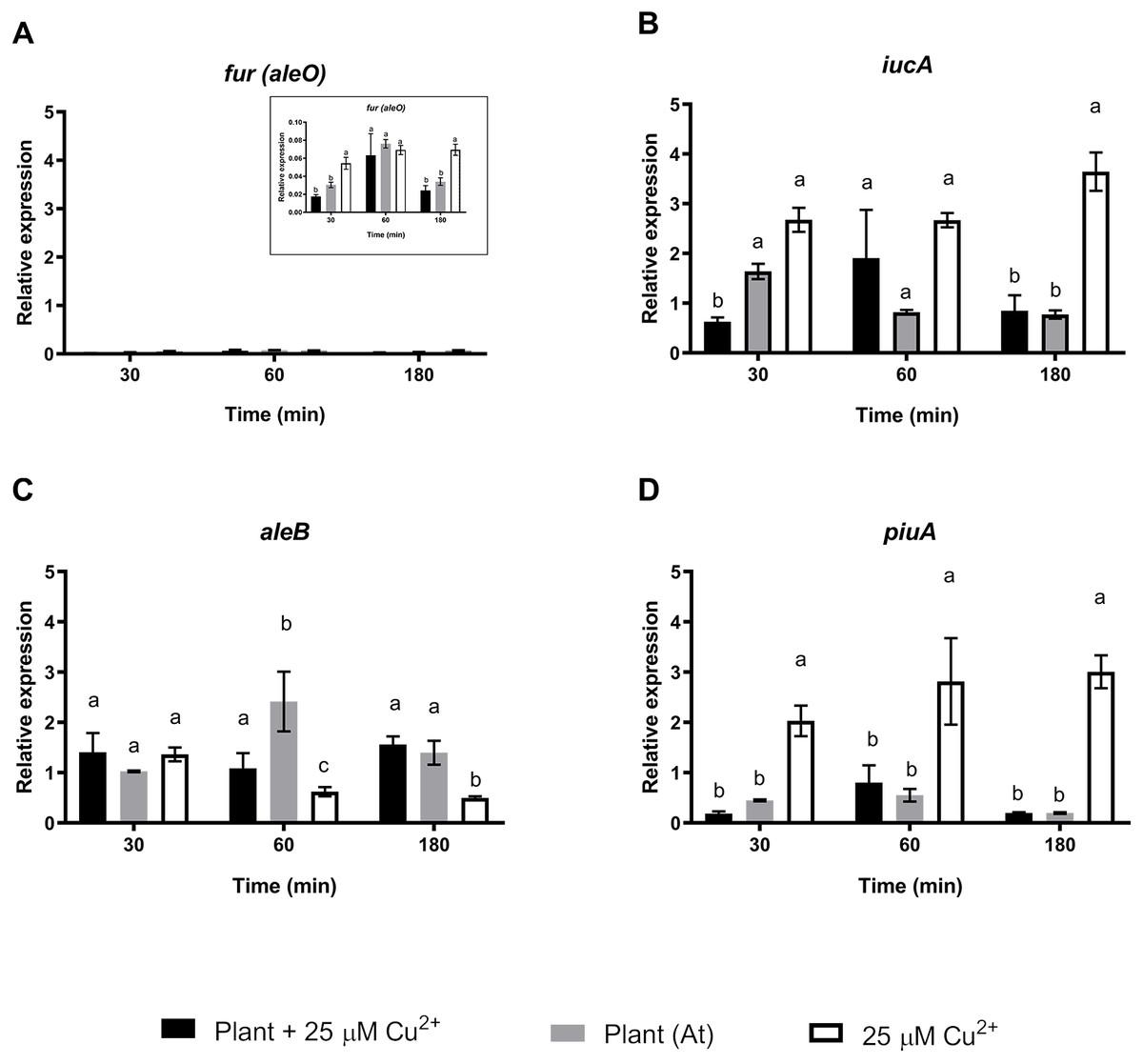
Figure 3: Relative expression levels of staphyloferrin B siderophore production genes from Cupriavidus metallidurans CH34, in the presence or in the absence of Arabidopsis thaliana (At) or copper.
Quantitative Real Time Polymerase Chain Reactions determinations of relative expression levels of (A) fur (aleO), (B) iucA, (C) aleB, and (D) piuA genes. Expression levels were normalized respect to the housekeeping gene 16S rRNA. Data correspond to means ± standard deviations of at least three biological replicates. Different letters indicate significant differences between same time conditions (two way ANOVA Tukey’s HSD tests; p < 0.05).{kind=link}
Discussion
Our results indicate that the C. metallidurans-A. thaliana interaction is characterized by a dual effect on plant growth and metal availability. A dependence on Cu2+ levels, and significant effects on of A. thaliana’s bacterium colonization and bacterial gene expression. We have previously demonstrated that closely related species such as C. pinatubonensis and P. phytofirmans have also PGPR abilities (Ledger et al., 2012; Poupin et al., 2013; Zúñiga et al., 2017) but not showing this dual effect.
C. metallidurans’ ability to act as a PGPR (see the increase in root length at low Cu2+ levels, Fig. 1), at least in A. thaliana, is not surprising as several members of the Cupriavidus genus and the Burkholderiaceae family are well-known PGPRs or, at least, interact with plants (Gyaneshwar et al., 2011; Bhattacharyya & Jha, 2012; Poupin et al., 2013; Arroyo-Herrera et al., 2020). Because PGPR traits (Zeffa et al., 2020), including 1-aminocyclopropane-1-carboxylate deaminase, auxin phytohormone production, phosphate solubilization, and nitrogen fixation (Bhattacharyya & Jha, 2012; Aeron et al., 2020), have not yet been reported in C. metallidurans, new PGPR traits would be interesting to explore in this bacterium. New PGPR traits could emerge from further in-depth study of the responses that gobernate this interkingdom relationships. Furthermore, the colonization behavior reported here is indicative of the inherent dynamics of the rhizospheric environment (Bais et al., 2006; Chaparro, Badri & Vivanco, 2014) and highlight the importance to describe and study these traits in environmental intertwined plant-microbre systems.
Few studies have addressed the effects that C. metallidurans CH34 exerts in plants and their rhizosphere environments. Because it had no apparent plant growth-promoting or protection effects, strain CH34 has been employed as a neutral bacterium (Taghavi et al., 2009). In a bioaugmentation-assisted phytoextraction procedure, strain CH34 was shown to increase Cr and Pb levels in maize shoots by a factor of 4.4 and 3.2, respectively, but it reduced Cr and Pb levels in roots by a factor of 2.9 and 4.8, respectively, illustrating how metal uptake and mobilization depend on metal species and plant tissue (Braud et al., 2009). Our results suggest a possible linkage between the plant metal uptake and the plant development when C. metallidurans CH34 is present. Microbially mediated plant growth promotion effects through metal level increases in Arabidopsis, have been scarcely explored (Lu et al., 2020).
The effects of negative Cu excess on plant growth have been previously studied (Doncheva, 1998; Burkhead et al., 2009; Kumar et al., 2021), and found that Cu mostly accumulates in root tissues (Alaoui-Sossé et al., 2004; Navari-Izzo et al., 2006; Burkhead et al., 2009). Cu provokes the reorganization of root architecture, leading to primary root growth inhibition and secondary root stimulation (Lequeux et al., 2010), although the latter effect has not been observed in A. thaliana, at least under the growth conditions used in this study. The detrimental effects of high Cu levels reported here may also be related to the uncontrolled mobilization of other micronutrients (Andresen, Peiter & Küpfer, 2018; Kumar et al., 2021).
It can be hypothesized that the dual effect of C. metallidurans on A. thaliana growth, either beneficial or detrimental depending on the Cu concentration, may be explained, although not exclusively, by the metal mobilization stimulated by this bacterium (Tables 3 and 4). This strain possesses transport mechanisms for multiple metallic ions, siderophores, and other metal-chelating molecules (Kirsten et al., 2011; Nies, 2016). At low Cu levels, strain CH34 may promote A. thaliana growth, increase micronutrient levels, and enhance uptake by roots and further mobilization to upper tissues. At high Cu2+ concentrations, such enhanced uptake and mobilization processes may cause micronutrient tissue accumulation at sub-toxic or toxic levels, impairing A. thaliana growth. It has been previously reported that Cu can alter the accumulation and translocation of transition metals (Karimi et al., 2012; Andrés-Colás et al., 2013), causing a reduction of Mn and Fe and an augmentation of Zn total content in roots. Under the conditions tested here, Cu’s effects were especially significant for B and Fe accumulation, as well as Cu (Puig et al., 2007; Puig & Peñarrubia, 2009). Mo mobilization is clearly blocked, in a less clear process (Andresen, Peiter & Küpfer, 2018).
The discrepancies in our results with those of other studies can probably be explained by our testing of micronutrient concentrations that were lower than those used in other studies (Doncheva, 1998; Alaoui-Sossé et al., 2004; Andrés-Colás et al., 2013; Saleem et al., 2020). Additionally, our experimental approach involved the addition of six metals along with Cu, which has not been previously addressed in literature and has implications for the use of C. metallidurans in phytoremediation procedures for metal-containing polluted sites (Gadd, 2004; Glick, 2010; Rajkumar et al., 2012), especially those contaminated with metal mixtures. There have been several recent studies on the use of microorganisms to remediate sites polluted with metal combinations (e.g., Cameron, Mata & Riquelme, 2018; Choinska-Pulit, Sobolczyk-Benarek & Laba, 2018). Interestingly, co-inoculation approaches have proved successful when protecting alfalfa from Cu/Zn mixtures (Jian et al., 2019), cleaning slurries containing heavy metal mixtures plus polycyclic aromatics (Subashchandrabose et al., 2019), or phytoremediating mine tailings from a Cu, Fe, Zn, and sulfur extraction operation (Benidire et al., 2021). Therefore, strain CH34 may be a good candidate to be used as a microbuial consortium member in co-inoculation schemes to take advantage of the beneficial properties and to prevent potential deleterious effects (Cillero et al. submitted, Supplementary Material).
The colonization of A. thaliana by strain CH34 cells indicated that in the presence of Cu, microenvironment conditions next to the plant increased the attraction and/or proliferation of this bacterium, which may be explained by changes produced in the composition of A. thaliana root exudates, mainly composed of sugars, amino acids, and aromatic compounds (Sasse, Martinoia & Northern, 2018; Vives-Peris et al., 2020). Strain CH34, as well as other closer Cupriavidus strains, is not able to grow on common sugars but it is able to proliferate on A. thaliana root exudates (Cillero et al. submitted, Supplementary Material). Cupriavidus strain genomes usually encode a significant group of genes encoding degradation of aromatic compounds that enable these strains to use a wide range of such organic compounds for growth (Lykidis et al., 2010; Pérez-Pantoja et al., 2015), providing ecologically relevant advantages, such as those reported in the closely-related strain C. pinatubonensis JMP134 (Ledger et al., 2012; Pérez-Pantoja et al., 2015). Additionally, Cu-induced stress in plants may also increase root exudation (Huang et al., 2016). Variations in the rhizosphere microenvironment have a mayor role modulating the rhizospheric microbiome, which is capable of recruit or discard some microbial species to dynamicly adjust benefic or detrimental interactions on the system (Zhalnina et al., 2018). C. metallidurans colonization features reported here may reflect system responses to these microenvironmental changes.
We wanted to start exploration of effects in strain CH34 gene expression of the presence /absence of the plant and Cu. The presence of A. thaliana and Cu differentially affected strain CH34 expression of some genes. For instance, cop genes involved in metal detoxification (copK and copC) were influenced by A. thaliana and Cu to a greater extent than the other cop genes, suggesting that periplasmic detoxification (Monchy et al., 2006) plays a significant role in the A. thaliana-bacterium-metal interaction. Genes involved in Fe turnover were also affected by interaction in the A. thaliana—C. metallidurans –copper triad, as has been previously reported (Puig et al., 2007). Plant-induced phaC1 gene expression was also in accordance with C. metallidurans’ interactions with A. thaliana, as polyhydroxybutyrate production is an advantage reported during early plant colonization (Kadouri, Jurkevitch & Okon, 2005; Balsanelli et al., 2016). Additional studies are clearly required as some no significant changes reported here for earlier times might not be found at later stages (7, or 14 days).
Conclusion
This work contributes to our understanding of plant-microbe interactions. At least for the metal multi-resistant C. metallidurans—A. thaliana system, we established that the dual (beneficial or detrimental) effect is dependent on copper levels, the metal root-to-shoots translocation is affected, and the validation of A. thaliana bacterial colonization and expression of some bacterial genes. Therefore, the role of metal-tolerant bacteria thriving in the rhizosphere of the plant-bacteria-metal triad deserves special attention, and plant growth promotion, protection, and/or phytoremediation strategies should be explored. Metal (Cu) levels play a key role and are probably the main factor controlling plant-microbe interactions. However, it should be kept in mind that the results reported here were derived under laboratory conditions. Further research is required to validate plant-metal tolerant bacterium-metal contents under field conditions.
Supplemental Information
Effects of inoculation of Arabidopsis thaliana with heat-inactivated Cupriavidus metallidurans cells, in the presence or in the absence of copper
Representative 21 days after sowing A. thaliana individuals, inoculated or not with heat-inactivated cells of C. metallidurans (K-CH34) in the absence (left side), or the presence (right side) of copper (50 µM CuSO4x5H2O). MS: Murashige—Skoog plant growth medium.
Relative expression levels of phaC1 gene from Cupriavidus metallidurans CH34, in the presence or in the absence of Arabidopsis thaliana or copper
Quantitative Real Time Polymerase Chain Reactions determinations of relative expression levels were normalized respect to the housekeeping gene 16S rRNA. Data correspond to means ± standard deviations of at least three biological replicates. Different letters indicate significant differences between same time conditions (Two way ANOVA Tukey’s HSD tests; p < 0.05).